



A new species of Acarothrix (Trombidiformes: Halacaridae) with notes on its sperm cells
Pepato, Almir R.  1
and Moreira, Daniel B. P.
2
1
and Moreira, Daniel B. P.
2
1Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Zoologia, Laboratório de Sistemática e Evolução de Ácaros Acariformes, Av. Antônio Carlos, 6627, Pampulha, Belo Horizonte - MG, Brazil.
2Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Zoologia, Laboratório de Sistemática e Evolução de Ácaros Acariformes, Av. Antônio Carlos, 6627, Pampulha, Belo Horizonte - MG, Brazil.
2026 - Volume: 66 Issue: 2 pages: 601-611
https://doi.org/10.24349/zk4z-iry3ZooBank LSID: 37E0C0AF-F49A-40B0-B17A-9F7416D87F7B
Original research
Keywords
Abstract
Introduction
The genus Acarothrix Bartsch, 1990 comprises six species distributed over tropical and warm temperate brackish or fluctuating salinity ecosystems around the world (Bartsch 1990, 1997, 2004, 2006a, 2015a; Procheş 2002; Chatterjee et al. 2012; Chatterjee 2019).
The genus is regarded as member of subfamily Copidognathinae Bartsch, 1983 along the diverse genus Copidognathus Trouessart, 1888 (> 359 spp), and the less diverse genera Copidognathides Bartsch, 1976 (3 spp), Werthella Lohmann, 1907 (10 spp), and the monotypic Phacacarus Bartsch, 1992 (Bartsch 2009). The genera included in the subfamily share, among other features, adults with a single pair of genital acetabula, solenidion on tarsus II dorsolateral and a single active nymphal instar, the protonymph (except for Phacacarus, for which there is no data on ontogeny) (Bartsch 2015b). The close relatedness between Copidognathus and Acarothrix was supported by molecular inference that recovered these genera in a well supported clade (Pepato et al. 2018, 2022).
Acarothrix species differ from Copidognathus due leg chaetotaxy (two instead three ventral setae on tibiae I and II) and, interestingly, genital acetabula, an osmoregulatory structure, external in males. Sexual dimorphisms that includes external genital acetabula in males and internal in females is also known from species belonging to the genus Halacarellus, Thalassarachna, Isobactrus and Halacaroides (Bartsch 2004, 2008; Pepato et al. 2011).
In this study, a new Acarothrix species is described along observations on its sperm cell morphology that is compared to the other eight halacarid species (Acaromantis vespucioi Pepato & Tiago, 2004, Copidognathus floridensis Newell, 1947, C. tupinamborum Pepato & Tiago, 2005, C. brachystomus Viets, 1940, Halacarellus thomasi (Newell, 1984), Halacaroides antoniazziae Pepato, Tiago & Rocha 2011, Rhombognathus levigatoides Pepato & Rocha 2007, and Thalassarachna basteri (Jonhston, 1836)) for which the sperm cells were described thus far (Alberti 1980; Alberti and Meyer-Rochow 2002; Pepato and Rocha 2007, 2010; Pepato et al. 2016).
Material and methods
Mites were obtained from green algae taken along Tramandaí River banks, Imbé municipality, Rio Grande do Sul State, Brazil (29°58′26.07″S, 50°08′08.55″W) on October, 2nd 2006. The locality is in the backyard of CECLIMAR (Centro de Estudos Costeiros Limnológicos e Marinhos), UFRGS (Federal University of Rio Grande do Sul).
Mites were sorted with the aid of a Stemi DV4® Carl Zeiss stereoscopic microscope. Mites preserved for DNA extraction and taxonomy were stored in Alcohol 100% at -20 °C. DNA extraction, molecular procedures and the first report on results can be found in Pepato and Klimov (2015). Mites for taxonomy were cleared with lactic acid and mounted in glycerine jelly. Drawings were made with aid of a drawing tube attached to a DM2500® Leica compound microscope; the measurements were taken by a graduated ocular micrometer.
Type material is deposited in the Acarological Collection at Centro de Coleções Taxonômicas da UFMG (CCT-UFMG). Collection acronym UFMG AC.
For Transmission electron microscopy, living male mites had their gnathosoma removed and legs detached by means of a fine entomological needle in ice-cold fixative (2.5% glutaraldehyde buffered to pH 7.2 in 0.1M cacodylate buffer). Following 2 h in cold fixative, the mite was rinsed in buffer solution and postfixed in 2% OsO4 for 2 h, rinsed again with buffer solution, immersed in uranyl acetate for 12 h, and dehydrated in a graded series of ethanol. Material was embedded in Spurr's resin. Polymerization required 24 h at 70 °C. Semi-thin sections were stained with toluidine blue and observed under a light microscope in order to allow general orientation. Ultra-thin sections through selected regions were obtained with a Leica Ultracut R® ultramicrotome, stained with lead citrate, and observed under a MET LEO 906E® Zeiss transmission electron microscope.
Abbreviations used in the description: AD, anterior dorsal plate; AE, anterior epimeral plate; GA, genitoanal plate; GO, genital opening; GP, genital plate; OC, ocular plate; PD, posterior dorsal plate; PE, posterior epimeral plate; ds, dorsal setae, from anterior to posterior: ds-1 to ds-5; gac, genital acetabula; gp, gland pores; pc, pore canaliculus; pgs, perigenital setae; sgs, subgenital setae. Legs are numbered I to IV and the segments, in proximal to distal, named trochanter, basifemur, telofemur, genu, tibia and tarsus. Palpal segments in same order are numbered P1 to P4. The chaetotaxic formula, excluding solenidia and parambulacral setae, runs from trochanter to tarsus. The position of certain structures is described according to Newell (1984), using the decimal system.
Systematics
Subfamily Copidognathinae Bartsch, 1983
Diagnosis
Dorsum with plates AD, OC and PD separate or merged; 3–4 pairs of gland pores (may be almost vestigial); 2–6 pairs of idiosomatic seta; ds-6 (if present) on anal cone. AE with pair of epimeral pores. PE with one dorsal and 2–3 ventral setae. Adults with single pair of gac. Female GA with 2-5 pairs of pgs and 0-1 pair of sgs. Male GA with 2–40 pairs of pgs; genital sclerites with 0-5 pairs of sgs. Gnathosoma normally with two pairs of maxillary setae, one pair on rostrum, one pair on gnathosomal base (rarely 2–7 pairs). Palps 3–4-segmented, attached laterally and at least slightly surpassing rostrum. Segment homologous to P-2 with one dorsal seta, that homologous to P-3 without a seta, and that representing P-4 with 0–3 setae in basal whorl; apically with setula and two spurs. Genua of legs shorter than telofemora and tibiae. Tibiae I and II with 2–3 ventral setae, tibiae III and IV with single pair of setae. Solenidion on both tarsus I and II in dorsolateral position. Tarsus I with 1–3 ventral setae, following tarsi lack ventral setae. Tarsus III and IV with 3–4 dorsal setae, both tarsi generally without ventral setae. All tarsi with paired claws. Central sclerite with minute, generally claw-like process. A single nymphal stage, the protonymph. Its GP with single pair of gac; pgs and sgs lacking (Bartsch 2015b).
Genus Acarothrix Bartsch, 1990
Diagnosis
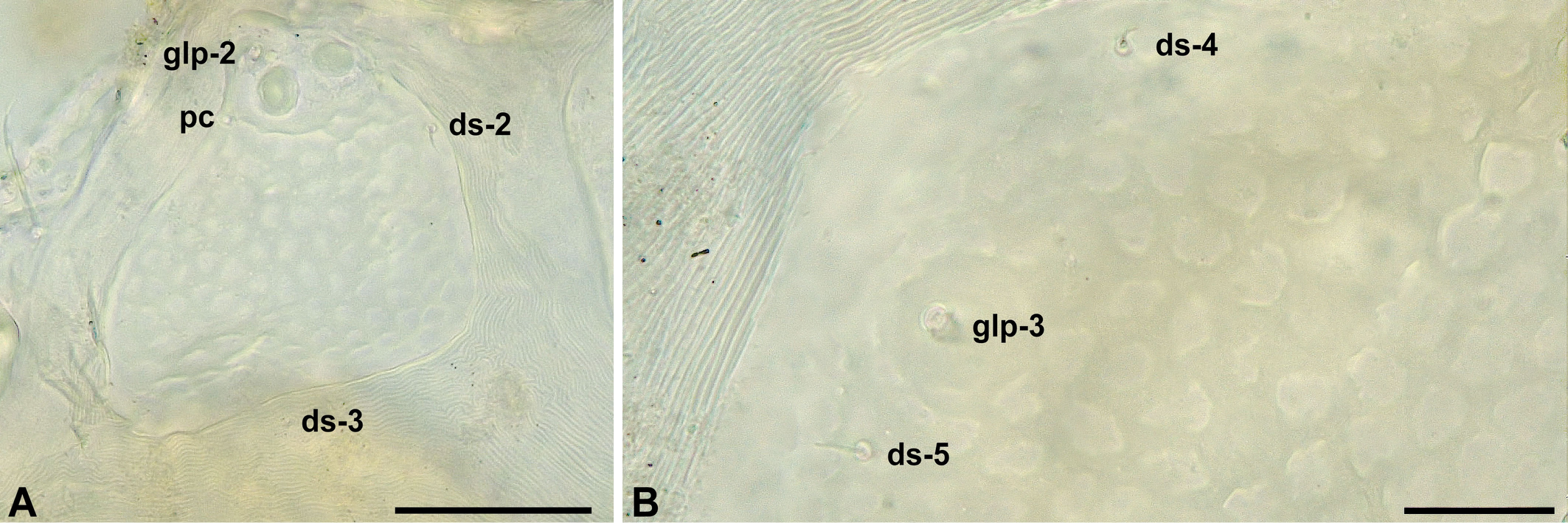
Copidognathinae, adult idiosoma 278–457 μm long. AD, OC and PD present. OC with 1–2 setae. Dorsum with 3–4 pairs of gland pores, and 5–6 pairs of idiosomatic setae; adanal setae on anal plate in either dorsal or ventral position. AE with pair of epimeral pores. PE with one dorsal seta and three ventral setae. Female GA with three pairs of perigenital setae, genital sclerites with one pair of subgenital setae. Male GA with numerous perigenital setae, its genital sclerites with four pairs of subgenital setae and one pair of external genital acetabula. Four-segmented palps attached laterally. Rostrum triangular. Gnathosomal base and rostrum each with a pair of maxillary setae. Rostral tip with two pairs of rostral setae. P-4 with three setae in basal whorl. Basifemora I–IV with 2,3,2,2 setae. Tibiae I–IV with 3–4,3–4,3,3 dorsal setae and 2,2,2,2 ventral setae, of which the ventromedial one is bipectinate on tibiae I to III or I to IV. Solenidion on both tarsus I and II in dorsolateral position. Tarsi I–IV with 2–3,0,0,0 ventral and 3,3,4,3 dorsal setae (solenidia excluded). Central sclerite with small, bidentate median claw (emended from Bartsch 2006b).
Type species: Acarothrix palustris Bartsch, 1990
Acarothrix geraldonae sp. nov.
ZOOBANK: 9BD75E14-9273-4F5C-BB07-94F073B1742F ![]()
Material examined
Holotype — ♀ (UFMGAC 1200279), from green algae along Tramandaí River banks, Imbé municipality, Rio Grande do Sul State, Brazil (29°58′26.07″S, 50°08′08.55″W), 02 October 2006, coll. A. R. Pepato.
Paratypes — 8 ♀ (UFMG AC 1200267, 1200270-1, 1200273, 1200276, 1200280, 1200282, 1200284), 1 ♂ (UFMGAC 250006), 4 Protonymphs (UFMG AC 1200272, 1200277-8, 1200283) sampling data as holotype.
Additional material — 5 ♀ (UFMG AC 1200268, 1200281, 250007-9), 2 ♂ (UFMGAC 1200285, 230020), 2 Protonymphs (UFMG AC 1200275, 1200286), sampling data as holotype.
Voucher for molecular data — UFMGAC 1200267.
Sequences deposited in GenBank — Cytochrome c Oxidase subunit I (COI), 489 bp, MG696250; Small subunit ribosomal RNA gene, 1,725 bp, KP276481; Large subunit ribosomal RNA gene, 3,461 bp, KP276405.
Description
Measurements as shown in Table I. Otherwise stated with indices, measurements were taken from the nine females, a male and four protonymphs included in the type series.
Download as
Structure
Female
Male
Protonymph
Length
Width
Length
Width
Length
Width
Idiosoma
375-457
272-397
402
326
266-320
190-251
Gnathosoma
116-1338
83-967
119
82
85-97
60-65
AD
97-118
116-142
114
128
70-84
84-95
PD
208-230
174-200
230
213
141-154
94-101
OC
101-122
78-110
101
109
60-79
55-76
AE
83-100
267-354
94
302
67-91
182-218
PE
182-251
–
221
–
117-135
–
GA or GP
157-196
138-172
202
169
62-81
71-86
GO
82-93
26-56
72
47
–
–
Rostrum
52-638
42-507
58
42
35-50
31-35

Female — Idiosoma flattened, wide (Fig. 1A-B), medial and lateral eyes with brown pigment. Mites covered by a thin layer of hazy material that partially conceal the surface ornamentation, after its removal it is possible to see that the dorsal plates are paneled (Fig. 3A-B) Dorsal plates AD, OC and PD separated by stripes of striated cuticle (Fig. 1A). AD wider than long. Pair of gland pores marginal, on rounded projections, at 0.27-0.37 of AD length, anterior to ds-1 at 0.43-0.49 of AD length. Anterior margin of AD slightly convex, posteriorly with a truncated or arcuate projection (Fig. 1A). Ocular plates with two cornea and glp-2 on raised area, pore canaliculus at posterior edge of raised area. Pair of ds-2 on anterior OC, pair of ds-3 on striated cuticle between OC and PD. PD almost uniformly paneled, costae ill defined, obscured by plate ornamentation. Pairs of ds-4 and ds-5 on PD at 0.25-0.38 and 0.56-0.60 of its length, respectively. Pair of glp-3 at 0.44-0.48 PD length and glp-4 on its posterior margin (Fig. 1A, 3B).
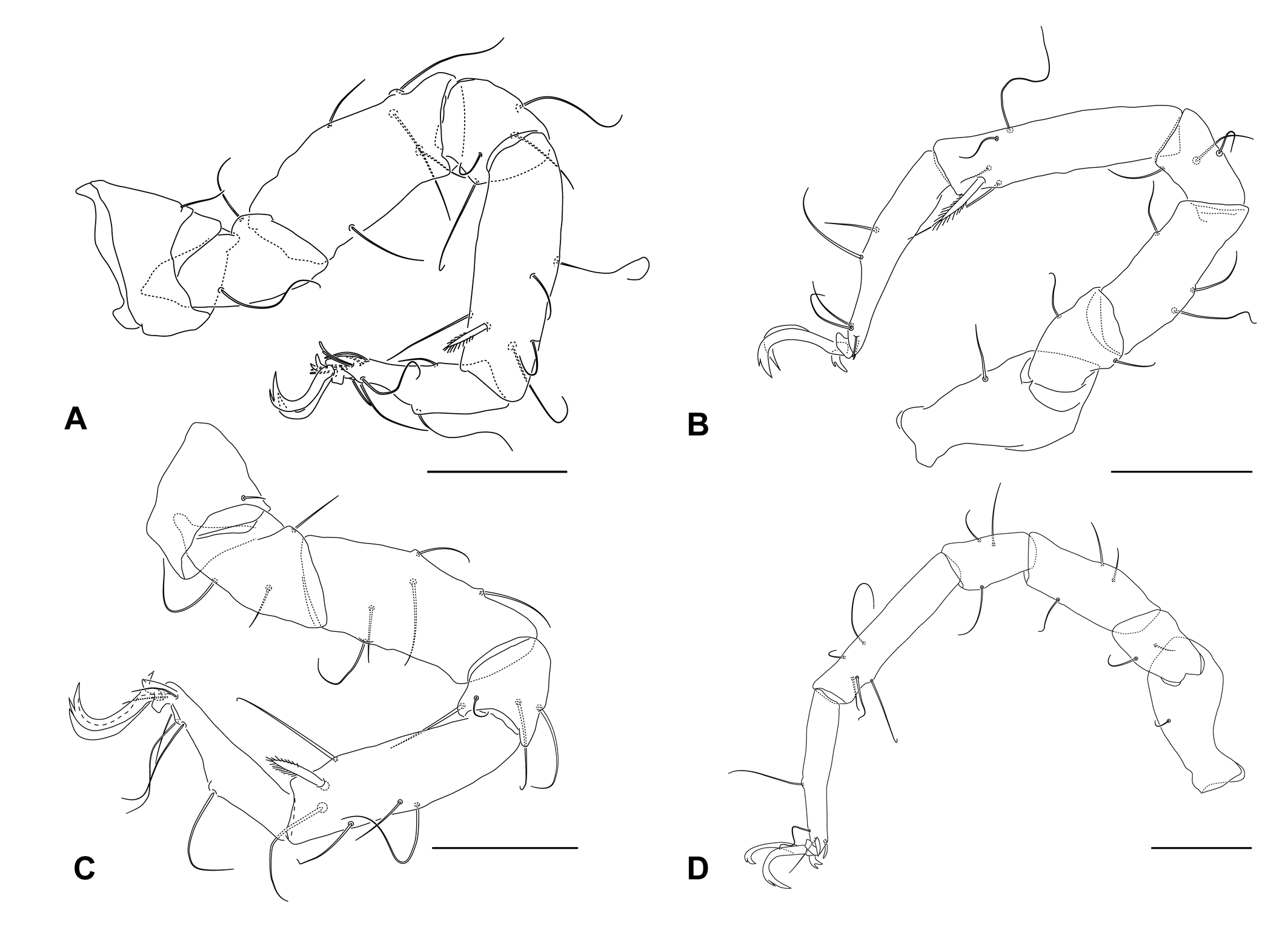
Anterior Epimeral plate posterior margin concave (Fig. 1B); three pairs of ventral setae; pair of epimeral pores lateral to apodemes between epimera I and II (Fig. 1B); pores slightly elongated, 7-14 μm long, 6-9 μm wide, its opening consisting of a complex pattern of slits. Posterior Epimeral plate with one dorsal seta and three ventral setae. Genitoanal plate anteriorly rounded, with three pairs of pgs. GO distance to anterior margin of GA equalling 0.20-0.30 of length of GO. Genital sclerites with one pair of sgs. Ovipositor extending to anterior border of genital sclerites. Anal cone ventral, bearing pair of ds-6, franked by a pair of lirifissures.
Gnathosomal rostrum is triangular, 0.43-0.54 of gnathosoma length (Fig. 1C); extending to about two thirds of P-2. Tectum arcuate. One pair of maxillary setae distal on gnathosomal base, the other pair at 0.38-0.51 of rostrum length (measured from proximal end of rostrum); two pairs of rostral setae at tip of rostrum. P-4 with three basal setae removed from basis by about length of P-3 and a minute subterminal seta.
Leg chaetotaxy (solenidia and pas excluded, bipectinated setae indicated by roman numerals within parenthesis): leg I, 1, 2, 5, 4, 6(I), 6; leg II, 1, 3, 5, 4, 6(I), 3; leg III, 1, 2, 3, 3, 5(I), 4; leg IV, 1, 2, 3, 3, 5, 3 (Fig. 2A-D). Bipectinate ventromedial seta on tibiae short (Leg I 17-21 μm, Leg II 20-24 μm, Leg III 19-26 μm) with pectens restricted to their distal half. Tarsus I with three dorsal setae; three ventral setae with a slender setae on its basal third and a pair of distal ventral setae eupathidia-like; solenidion 13-20 μm (due its position, accurate measurements of Tarsi I-II solenidia were precluded, hence values much be taken with caution) close to dorsolateral fossary seta; a papilate famulus; and a pair of distal doublet eupathidia (Fig. 1D). Tarsi II-IV lack ventral setae. Tarsus II with three dorsal setae, dorsolateral fossary seta close to tip of tarsus, adjacent to solenidion; solenidion 15-21 μm, approximately; and with a pair of doublet eupathidia (Fig. 1F). Tarsus III with four dorsal setae, eupathid medial and widened but tapering lateral pas (Fig. 1E), tarsus IV with medial singled eupathid and a lateral bristle-like pas (Fig. 1G). Claws with accessory process; pecten lacking. All tarsi with small bidentate median claw.
Male — Similar to female in most aspects. GA anteriorly rounded. GO apart from anterior border of GA by 0.46 of its length. Spermatopositor 122 μm long, 92 μm wide. GA with 21-24 pgs (Individuals UFMG AC 230020 and 250006, Fig. 1H). Genital sclerites with four pairs of sgs, arranged 2:2; both anterior sgs pairs slender and setiform, the posterior pairs spur-like and setiform (Fig. 1H). A pair of external genital acetabula immediately posterior to sgs, 8-10 μm in diameter.

Protonymphs — Dorsal plates less developed than in adults (Fig.4A-B). AE with three pairs of setae and a pair of epimeral pores. PE with two ventral setae. Genital plate without setae, with a single pair of gac. Leg IV five segmented. Chaetotaxy as follows (Fig 4 C-F): leg I: 1,2,3,4,5(I),6; leg II: 1,3,3,4,5(I),3; leg III:1,1,2,3,5(I),4; leg IV: 0,1+2,3,5,3.
Sperm cell morphology — Similar to other Acariformes, Acarothrix testes are divided in a dorsal germinative and ventral secretory portions, emptying their products in a common lumen (Fig. 5C). The glandular portion consists of cells with scattered mitochondria, well-developed rough endoplasmatic reticulum, and Golgi bodies (Fig. 5A). The lumen of the secretory border is brush-like (arrowheads in Fig. 5B). The chromatin body (the remnants of nuclei after its condensation and lost of nuclear envelope during spermiogenesis) is elongated, a complete acrosomal complex and tubules filed with an electrodense material are evident in later spermatids (Fig. 5E). The later spermatid undergoes cytoplasmic condensation, making the electrodense filled tubules give a convoluted aspect to the mature sperm cells, which present the chromatin body placed peripherally and are individually coated by a layer of secretion.

Mature sperm cells were found in a matrix of ''simple'' and ''complex'' corpuscles, the later consisting of flattened, spindle-shaped secreted bodies, with peripheric layers with distinct densities (some marked with a * in Fig. 6), while the former are denser, uniform and appears to get stuck to sperm cells (Fig. 5B-C, 6) It could be detected a accompanying secretion structured as rows of platelets (Fig. 5D) sunk in a fine grained matrix.

Etymology
The species is named after a Lahille's bottlenose dolphin (Tursiops gephyreus) female, nicknamed Geraldona by local fishermen. She is one of dolphins know to engage in cooperative fishing with humans in the same region where mites were obtained.
Remarks
Currently, six Acarothrix species are known: A. ampliata Bartsch, 2004; A. ampliumeris Bartsch, 2006; A. grandocularis Chatterjee, Marshall, Guru, Ingole & Pesic, 2012; A. longiunguis Bartsch 1997; A. palustris Bartsch, 1990; A. umgenica Procheş, 2002. Acarothrix geraldonae sp. nov. is unique in bearing the pair of ds-3 on striated cuticle between OC and PD, while A. grandocularis has it on PD, and all other species to which this feature is know on OC (A. umgenica original description does not report the placement of these setae (Procheş 2002)).
Additionally, A. geraldonae sp. nov. differs from A. ampliata by bearing six instead of five setae on tibae I-II (Bartsch 2004); A. ampliumeris has elongated OC without corneae (vs usual OC with corneae), ds-2 on striated cuticle (instead on OC), glp-3 inconspicuous (vs conspicuous), bipectinate setae present on tibia IV (vs absent), and bipectinate setae long on tibia II (vs short) (Bartsch 2006a); A. longiunguis, A. palustris, and A. umgenica lack accessory processes on their lateral claws, which are present in A. geraldonae sp. nov., being lateral claws in A. longiunguis and A. umgenica elongated (Bartsch 1990, 1997; Procheş 2002); A. grandocularis has two setae on Basifemur II (vs three), and OC elongate triangular with rounded medial corner, each with two large corneae (Chatterjee et al. 2012; Bartsch 2015a); A. longiunguis presents glp-4 pair on prominently raised cones on posterior margin of PD (Bartsch 1997); A. umgenica differs from all other congeneric species due the shape of its dorsal plates and in having the pair of ds-1 placed posteriard (Procheş 2002).
Chatterjee (2019) reviewed the occurrences of species belonging to Acarothrix. It is interesting to note that despite most species are associated to Mangroves, A. geraldonae sp. nov. type locality is further south to the southernmost occurrence of mangroves along Brazilian coast, which occurrences ends at Laguna municipality, 28.5°S (Lacerda et al. 2022). Anyway, it shares with all other species included in the genus the occurrence in a stuarine setting, in brackish water, along another mite typical for this environment, adults and deutonymphs of the pontarachnid Litarachna brasiliensis Smit, 2007 (Smit 2007). The family Pontarachnidae includes marine and brackish species (Smit 2020).
Discussion
Acarothrix geraldonae sp. nov. was sequenced for the mitochondrial gene Cytochrome Oxidase I, and nuclear ribosomal genes for the small and large subunits. These sequences were included in a series of phylogenetic inferences (Pepato and Klimov 2015; Pepato et al. 2018, 2022). In all analyses, A. geraldonae sp. nov. was recovered as sister group to the species belonging to the genera Copidognathus. The sperm cells description presented here supports this, since Acarothrix and Copidognathus spermatozoa are indistinguishable in any relevant aspect.
The description of A. geraldonae sp. nov. sperm cells adds to the description of eight other halacarid species, Acaromantis vespucioi, Copidognathus floridensis, C. tupinamborum, C. brachystomus, Halacarellus thomasi, Halacaroides antoniazziae, Rhombognathus levigatoides, and Thalassarachna basteri (Alberti 1980; Alberti and Meyer-Rochow 2002; Pepato and Rocha 2007, 2010; Pepato et al. 2016). Despite this sampling is still scanty and does not gives an appropriate representation of halacarid diversity, combined with molecular phylogenies available allows advance some hypotheses on the evolution of sperm cells and their accompanying secretions.
In the most comprehensive molecular halacarid phylogeny to date, Pepato et al. (2022) recovers the genera Acarothrix, Copidognathus, Halacarellus, Halacaroides, and Thalassarachna derivate relative to Acaromantis and Rhombognathus. It suggests that characters states such as cell membrane infoldings filed with an electrodense material, and the presence of two distinct kinds of accompanying secreted bodies are better regarded as synapomorphies shared by the former genera, contrasting with simple pleats of plasmalemma, and a single kind of accompanying secreted bodies as observed in Rhombognathus and Acaromantis.
Acknowledgments
The first author expresses his deep gratitude to all the staff at CECLIMar-UFRGS, in particular to professors Dr. Norma Luiza Würdig and Dr. Carla Ozório, for the collection stay at that center. We also express our gratitude to Dr. Marta Maria Antoniazzi, Dr. Sylvia Mendes Carneiro, and Dr. Carlos Jared (Laboratório de Biologia Celular, Instituto Butantan, Brazil) who allowed the use of the TEM, and made useful suggestions about microscopy techniques.
References
- Alberti G. 1980. Zur Feinstruktur der Spermien und Spermiocytogenese der Milben (Acari). II. Actinotrichida. Zool. Jahrb. Abt. Anat. Ontog. Tiere, 104: 144-203.
- Alberti G., Meyer-Rochow V.B. 2002. Ultrastructural investigations of testes and spermiogenesis in two species of halacarid mites (Halacaridae, Actinedida, Actinotrichida): Thalassarachna basteri from the Baltic Sea and Halacarellus thomasi McMurdo Sound (Antarctica). Arthropod Struct. Dev., 30: 315-328. https://doi.org/10.1016/S1467-8039(02)00008-7
- Bartsch I. 1990. Acarothrix palustris gen. et spec. nov. (Halacaroidea, Acari), ein Bewohner der Salzwiesen Südchinas. Zool. Anz., 224: 204-210.
- Bartsch I. 2004. Acarothrix ampliata (Arachnida: Acari: Halacaridae), a new halacarid mite from Florida, with notes on external genital acetabula. Species Divers., 9: 259-267. https://doi.org/10.12782/specdiv.9.259
- Bartsch I. 1997. Copidognathinae (Halacaridae, Acari) from northern Australia; description of four new species. In: Hanley J.R., Caswell G., Megirian D., Larson H.K. (Eds) Proceedings of the Sixth International Biological Workshop, Museums & Art Galleries of Northern Territory, Darwin, pp. 231-243.
- Bartsch I. 2006a. Copidognathines (Acari: Halacaridae) in mangroves of Singapore. I. Description of three species. Raffles Bull. Zool., 54: 83-92.
- Bartsch I. 2006b. Halacaroidea (Acari): a guide to marine genera. Org. Divers. Evol., 6 Electronic Supplement: 1-104. https://doi.org/10.1016/j.ode.2005.07.001
- Bartsch I. 2008. Halacarid mites (Acari: Halacaridae) in a freshwater influenced beach of North Stradbroke Island, Moreton Bay. Mem. Queensl. Mus., 54: 117-130.
- Bartsch I. 2009. Checklist of marine and freshwater halacarid mite genera and species (Halacaridae: Acari) with notes on synonyms, habitats, distribution and descriptions of the taxa. Zootaxa, 1998: 1-170. https://doi.org/10.11646/zootaxa.1998.1.1
- Bartsch I. 2015a. Halacaridae (Acari) amongst the epiflora fauna on trunk, branches, roots and pneumatophores on the coast of Singapore: a survey. Raffles Bull. Zool., 31 (supplement): 96-138.
- Bartsch I. 2015b. The genital area of Halacaridae (Acari), life stages and development of morphological characters and implication on the classification. Zootaxa, 3919 (2): 201-259. https://doi.org/10.11646/zootaxa.3919.2.1
- Chatterjee T. 2019. Scanning electron microscopic observation of Acarothrix grandocularis (Acari, Halacaridae) and notes on the species of the genus Acarothrix. Acta Biol., 26: 117-126. https://doi.org/10.18276/ab.2019.26-10
- Chatterjee T., Marshall D.J., Guru B.C., Ingole B, Pešić V. 2012. A new species of the genus Acarothrix (Acari, Halacaridae) from Brunei Darussalam and India. Cah. Biol. Mar., 53 (4): 541-546.
- Lacerda L.D., Ferreira A.C., Borges R., Ward R. 2022. Mangroves of Brazil. In: Das S.C., Pullaiah, Ashton, E.C. (Eds) Mangroves: Biodiversity, Livelihoods and Conservation. Springer, Singapore. https://doi.org/10.1007/978-981-19-0519-3_20
- Newell I.M. 1984. Biology of the Antartic Seas. XV Antarctic Halacaroidea. Antarct. Res. Ser., 40: 1-284. https://doi.org/10.1029/AR040
- Pepato A.R., Tiago C.G., Rocha C.E.F. 2011. Psammophilous halacarid mites (Trombidiformes, Halacaridae) from the North Coast of São Paulo State, Brazil. Zootaxa, 2823: 47-60 https://doi.org/10.11646/zootaxa.2823.1.3
- Pepato A.R., Rocha C.E.F. 2007. Description of a new species of Rhombognathus (Halacaridae, Acari), its spermiogenesis and spermatozoa. J. Nat. Hist., 41: 2433-2445. https://doi.org/10.1080/00222930701567877
- Pepato A.R., Rocha C.E.F. 2010. On spermiogenesis, sperm cell morphology and accompanying secretions of Copidognathus (Acari: Halacaridae). Zool. Anz., 249: 151-164. https://doi.org/10.1016/j.jcz.2010.08.001
- Pepato A.R., Costa S.G.S., Harvey M.S., Klimov P.B. 2022. One-way ticket to the blue: A large-scale, dated phylogeny revealed asymmetric land-to-water transitions in acariform mites (Acari: Acariformes). Mol. Phylogenet. Evol., 177: 107626. https://doi.org/10.1016/j.ympev.2022.107626
- Pepato A.R., Klimov P.B. 2015. Origin and higher-level diversification of acariform mites - evidence from nuclear ribosomal genes, extensive taxon sampling, and secondary structure alignment. BMC Evol. Biol., 15: 178. https://doi.org/10.1186/s12862-015-0458-2
- Pepato A.R., Vidigal T.H.D.A., Klimov P.B. 2018. Molecular phylogeny of marine mites (Acariformes: Halacaridae), the oldest radiation of extant secondarily marine animals. Mol. Phylogenet. Evol., 129: 182-188. https://doi.org/10.1016/j.ympev.2018.08.012
- Pepato A.R., Antoniazzi M.M., Jared, C. 2016. Spermatogenesis, sperm cell morphology and accompanying secretions from two interstitial marine mites. Acta Zool., 97: 60-66. https://doi.org/10.1111/azo.12105
- Procheş Ş. 2002. New species of Copidognathinae (Acari: Halacaridae) from southern Africa. J. Nat. Hist., 36: 999-1007. https://doi.org/10.1080/00222930110034571
- Smit H. 2020. Water mites of the world: With keys to the families, subfamilies, genera and subgenera. Leiden (The Netherlands), Nederlandse Entomologische Vereniging, 774 pp.
- Smit H. 2007. Litarachna brasiliensis n. sp., the first member of the water mite family Pontarachnidae (Acari: Hydrachnidia) from South America. Syst. Appl. Acarol., 12, 141-146. https://doi.org/10.11158/saa.12.2.8



2026-03-29
Date accepted:
2026-06-16
Date published:
2026-06-24
Edited by:
Auger, Philippe

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Pepato, Almir R. and Moreira, Daniel B. P.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)