



A new species of the genus Achipteria Berlese, 1885 (Acariformes, Oribatida, Achipteriidae) from the Kingdom of Bhutan
Oshima, Masaharu  1
; Darjee, Beejai
2
; Dorji, Tshering
3
; Choden, Tshering
4
; Pelyang, Tashi
5
; Tshewang, Tshewang
6
; Yamauchi, Takeo
7
; Nomura, Shûhei
8
; Nakano, Takafumi
9
; Dorji, Karma Dema
10
and Shimano, Satoshi
11
1
; Darjee, Beejai
2
; Dorji, Tshering
3
; Choden, Tshering
4
; Pelyang, Tashi
5
; Tshewang, Tshewang
6
; Yamauchi, Takeo
7
; Nomura, Shûhei
8
; Nakano, Takafumi
9
; Dorji, Karma Dema
10
and Shimano, Satoshi
11
1✉ Faculty of Science, Kyoto University, Kitashirakawa-oiwakecho, Sakyo-ku, Kyoto, 606-8502 Japan.
2National Biodiversity Center, Ministry of Agriculture and Livestock, Serbithang, Thimphu, 11001 Bhutan.
3National Biodiversity Center, Ministry of Agriculture and Livestock, Serbithang, Thimphu, 11001 Bhutan.
4National Biodiversity Center, Ministry of Agriculture and Livestock, Serbithang, Thimphu, 11001 Bhutan.
5National Biodiversity Center, Ministry of Agriculture and Livestock, Serbithang, Thimphu, 11001 Bhutan.
6National Biodiversity Center, Ministry of Agriculture and Livestock, Serbithang, Thimphu, 11001 Bhutan.
7Laboratory of Entomology, Graduate School of Animal and Food Hygiene, Obihiro University of Agriculture and Veterinary Medicine, Inada-cho Nishi 2-11, Obihiro, Hokkaido, 080-8555 Japan.
8Department of Zoology, National Museum of Nature and Science, 4-1-1, Amakubo, Tsukuba-shi, Ibaraki, 305-0005 Japan.
9Department of Zoology, Graduate School of Science, Kyoto University, Kitashirakawa-oiwakecho, Sakyo-ku, Kyoto, 606-8502 Japan.
10National Biodiversity Center, Ministry of Agriculture and Livestock, Serbithang, Thimphu, 11001 Bhutan.
11Science Research Center, Hosei University, Fujimi 2-17-1 Chiyoda-ku, Tokyo, 102-8160 Japan.
2026 - Volume: 66 Issue: 2 pages: 530-542
https://doi.org/10.24349/pocc-d7lmZooBank LSID: F7568F5C-C75B-4C2E-81CC-DBEECB10A892
Original research
Keywords
Abstract
Introduction
The Kingdom of Bhutan is located in the eastern Himalayan region in South Asia. The elevation of Bhutan spans a broad gradient from 95 to 7,570 m above mean sea level, increasing progressively from the southern lowlands toward the northern high mountains (Walcott 2009; WCNP and WWF 2016). Despite its relatively small land area of 38,394 km2 (NECRGB 2022), Bhutan exhibits remarkable climatic and vegetational diversity resulting from an altitudinal range exceeding 7,000 meters (Ohsawa 1987; Wangda and Ohsawa 2006). This rich natural environment supports a high biodiversity, including many endangered species. However, research on the diversity of invertebrates has not been sufficiently advanced in Bhutan (National Biodiversity Center 2017; Gyeltshen and Prasad 2022).
Chakrabarti and Wilson (1981) stated that Bhutan fosters abundant soil microarthropod fauna, as well as other taxa. Focusing on oribatid mites among soil animals, several studies were conducted in Bhutan during the 1970s–1980s (Bayoumi and Mahunka 1979a, b; Sanyal 1988; Reddy 1989). However, no research on oribatid mites has been published since then, and fewer than 30 species have been recorded so far (Oshima, unpublished data).
Specifically, in 2025, a soil fauna survey was conducted as part of a joint research project between the National Biodiversity Center in Bhutan and a Japanese research team. During the survey, specimens of an oribatid mite belonging to the family Achipteriidae Thor, 1929 were collected and, based on morphological examination, identified as an undescribed species of the genus Achipteria Berlese, 1885.
The oribatid mite genus Achipteria is the most species-rich genus in the family Achipteriidae, containing two subgenera, 40 species and two subspecies (Subías 2004, unpublished online version 2024; Ermilov and Kontschán 2025). This genus is morphologically characterized by the following features: the pteromorph has a long anterior projection; the lamella is long and wide, connected in the middle, covering most of the prodorsal area and extending beyond the rostrum; the notogaster has 10 pairs of setae and four pairs of sacculi (Weigmann 2006). Achipteriid mites have a wide distribution ranging from the Holarctic, Oriental to Neotropical regions. Several species of the family have been found in the South Asian region, but had not previously been recorded in Bhutan (see Discussion section). In this study, we describe the Bhutanese specimens as a new species of this genus. This new species represents the first record of Achipteriidae from Bhutan, and the first documented record of an oribatid mite from Bhutan since the 1980s.
Materials and Methods
Soil samples were collected from the following two locations: the humus layer, fallen leaves, of a mixed forest of evergreen broadleaf trees (mainly Quercus semecarpifolia Sm.) and coniferous trees (mainly Pinus wallichiana A. B. Jacks.) at the Tiger's Nest Monastery (Paro Taktsang) in Paro; the humus layer of a coniferous forest in nearly semi-arid conditions, with deciduous broadleaf trees interspersed among the shrubs at the Royal Takin Preserve in Thimphu, the capital of Bhutan. Then, they were placed in each Tullgren apparatus and irradiated with a 60-W electric bulb to extract the organisms into 99.5% propylene glycol for fixation. The extracted soil organisms were transferred and sorted into collecting vials with 99% ethanol under the Leica EZ4 D Microscope. The sorted oribatid mites were mounted in lactic acid on cavity slides or in Hoyer's medium on slides for microscopic observation, measurement and illustration under 200–400× magnifications. Drawings were made with an Olympus BX53 Microscope equipped with a drawing attachment and then modified in Photoshop Elements 2024 (Adobe Inc.). For Scanning-Electron-Microscopic (SEM) investigation, specimens were dehydrated and finally replaced in pentane, air-dried, and then gold vapor-deposited. Scanning electron microscopic micrographs were taken by a JEOL JCM-6000PLUS SEM.
Body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the notogaster. Body width means the maximum width of the level of pteromorphs in dorsal view. Notogastral width refers to its maximum width behind pteromorphs in dorsal aspect. The lengths of body setae were measured in lateral view. All numbers measured on specimens in the text are in micrometers (μm).
Morphological terminology used in this paper mostly follows that of the literature on Achipteriidae (e.g. Grandjean 1932, 1935; Seniczak and Seniczak 2016; Maruyama et al. 2016; Ermilov and Kontschán 2025); also, Norton (1977) for leg setal nomenclature, and Norton and Behan-Pelletier (2009) and Behan-Pelletier and Lindo (2023) for overview.
Abbreviations — Prodorsum: lam = lamella; ro, le, in, ss = rostral, lamellar, interlamellar, bothridial setae, respectively; tu = tutorial cusp; D = dorsophragma; P = pleurophragma. Gnathosoma: a, m, h, or = subcapitular setae; sup, inf, d, l, cm, acm, ul, su, it, vt = palp setae; ω = palp solenidion; cha, chb = cheliceral setae; Tg = Trägårdh's organ. Notogaster: ptm = pteromorph; len = lenticulus; r = posterior ridge; c, la, lm, lp, h1–h3, p1–p3 = notogastral setae; im, ip, ih, ips = lyrifissures; gla = opisthonotal gland opening; Sa, S1, S2, S3 = sacculi. Epimeral and lateral podosomal regions: 1a, 1b, 1c, 2a, 3a, 3b, 3c, 4a, 4b, 4c = epimeral setae; gt = genal tooth; can = capitular angle; PdI, PdII = pedotecta I, II, respectively. Anogenital region: g, ag, an, ad = genital, aggenital, anal, and adanal setae, respectively; iad = adanal lyrifissure; ψ, τ, κ = setae of ovipositor; po = preanal organ. Legs: Tr, Fe, Ge, Ti, Ta = trochanter, femur, genu, tibia, tarsus, respectively; pa = porose area; ω, φ, σ = solenidia; ɛ = famulus; d, l, v, bv, ev, ft, tc, it, p, u, a, s, pv, pl = leg setae.
Taxonomy
Family Achipteriidae Thor, 1929
Genus Achipteria Berlese, 1885
Type species: Acarus coleoptratus Linnaeus, 1758
Achipteria (Achipteria) bhutanica Oshima & Shimano sp. nov.
ZOOBANK: DCED6626-0403-472E-B2E2-8A286FCBEAA7 ![]()
(Figures 1–7)
Material Examined
Holotype: adult female, Bhutan, the Tiger's Nest Monastery (Paro Taktsang), Paro, 27°29′29″N, 89°21′25″E, 3,039 m a.s.l., 11 Oct. 2025, collected by M. Oshima from litter and humus layer of fallen leaves. Paratypes: two adult males and one adult female, same data as for holotype; two adult males and three adult females, Bhutan, forest nearby the Royal Takin Preserve, Thimphu, 27°28′57″N, 89°36′46″E, 2,627 m a.s.l., 20 Jul. 2019, collected by S. Shimano from litter of the same layer. Non-types (Figures 4–7): two adults (sex unknown), same data as for holotype.
Type deposition
Holotype [collection number: BTN031] and four paratypes were deposited in the Invertebrates Repository of the National Biodiversity Center, Thimphu [collection number: BTN032 (male): Paro, 2025; BTN033–034(female), 035(male): Thimphu, 2019]; four paratypes were deposited in the Collection of Arachnida, Department of Zoology, National Museum of Nature and Science, Tokyo (NSMT) [NSMT-Ac 14786 (male), 14787 (female): Paro, 2025; 14788 (male), 14789 (female): Thimphu, 2019].
Diagnosis—Adult
Body size: 604–672 × 466–540. Body surface almost smooth. Rostral seta bent and heavily barbed. Tutorium with long cusp. Lamella broad, with single, pointed lateral tooth; anterior margin curving downward. Lamellar seta long, weakly barbed. Interlamellar seta almost reaching distal part of lamella, roughened. Bothridial seta clavate with barbed head. Pteromorph with small lateral tooth. Posterior edge of notogaster slightly concave medially, with small tubercle-like projection. Notogastral setae short, setiform and smooth. Pedotectum I with no tooth. All tarsi tridactylous.
Description—Adult

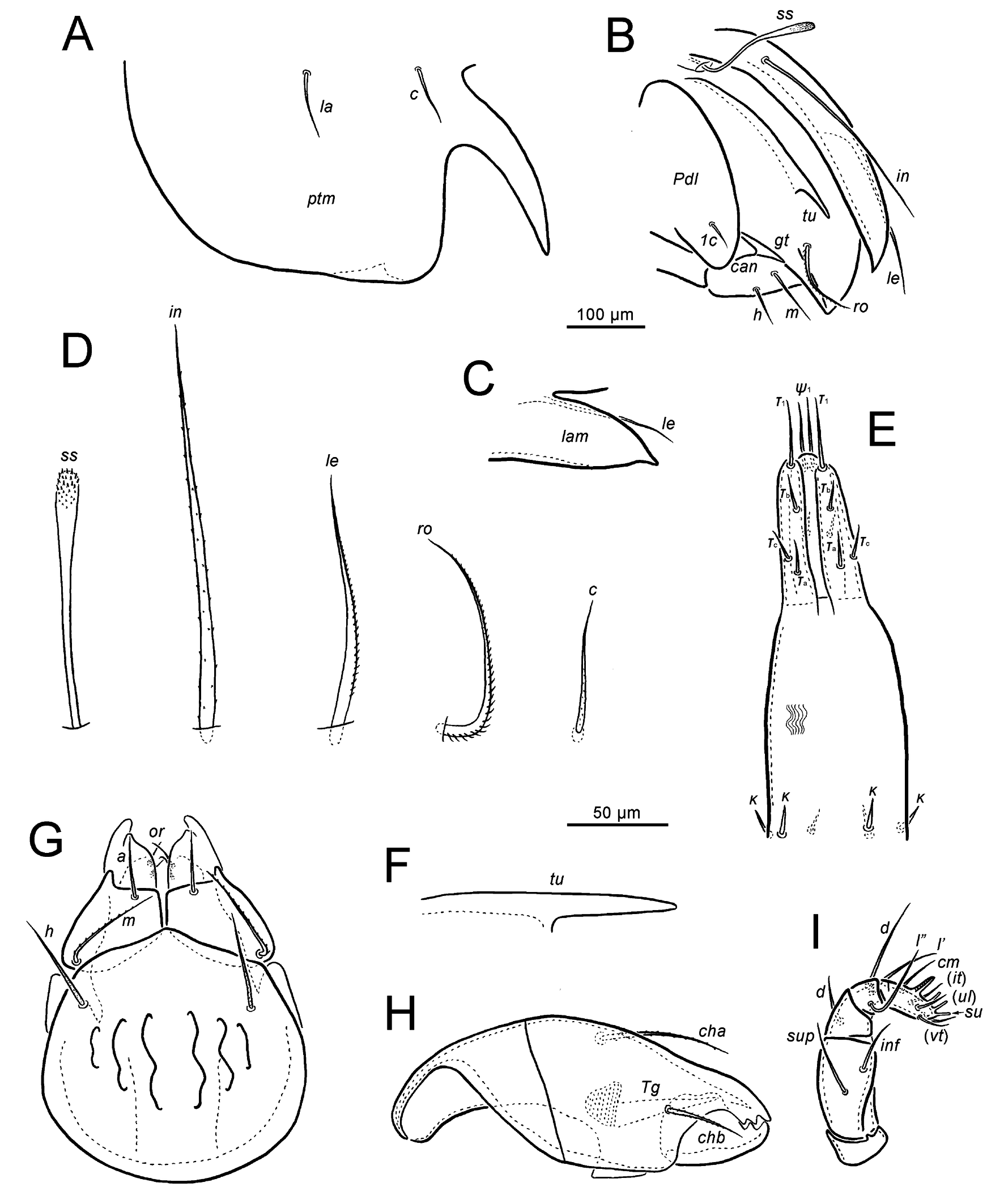



Measurements — Body length: holotype 664, male paratypes 604–639, female paratypes 644–672; body width: holotype 540, male paratypes 466–508, female paratypes 477–532; notogaster width: holotype 435, male paratypes 417–441, female paratypes 415–457.
Integument — (Figures 1A, B, 4A–C and 6A–H). Body color dark brown. Body surface almost smooth.
Prodorsum — (Figures 1A, B, 2B–D, F, 4A, C–G and 5B, D–F, 7D). Rostrum rounded, pair of ridges extends from tip backward. Rostral seta (108–114) long, heavily barbed, curved medially. Tutorium with long and narrow cusp. Lamella long and broad, fused medially, with single pointed lateral tooth, curving downward. Lamellar seta (102–127) long, setiform and weakly barbed, inserted on ventral side of lamella. Interlamellar seta (170–176) straight and roughened, almost reaching tip of lamella. Bothridial seta (93–101) clavate, with long stalk and barbed head. Exobothridial setae not observed.
Notogaster — (Figures 1A, 2A, D, 4A, B and 6D, E, G, H). Anterior margin projecting forward. Posterior end slightly concave medially, with small tubercle-like projection. Lenticulus with weak boundary. Pteromorph with small lateral tooth, numerous narrow ridges running longitudinally along reverse side. Anterior projection of pteromorph weakly curved anteromedially. Ten pairs of notogastral setae (c, la: 37–48; lm, lp, h1–h3: 8–12; p1–p3: 17–20) setiform, almost smooth. Four pairs of sacculi small, barely visible; Sa located anterolaterally to seta lm; S1 between setae lp and h3; S2 positioned anterolaterally to seta h2; S3 at midpoint of h1 and h2. Opisthonotal gland opening located posterolaterally to seta h3. Lyrifissures im, ip, ih, ips present (ia not observed).
Gnathosoma — (Figures 2G–I and 5D, E). Subcapitulum (163–175 × 118–132) with three pairs of setiform, smooth setae (a 31; m 56–60; h 51–53) and two pairs of setiform and smooth adoral setae (18, 19). Mentum with several ridges (sometimes indistinct). Palp (114–118) with five segments, setae formula: 0-2-1-3-9+ω, eupathidium acm fused with solenidion ω. Postpalpal seta (9–10) setiform, thick. Chelicera (168–170) with two setiform and barbed setae (cha 59–61; chb 35–36); cha longer than chb.
Epimeral and podosomal regions — (Figures 1B, 2B, 5A, C and 6A, B). Epimeral setal formula: 3-1-3-3; 1a, 1c, 2a (5–8) short, almost smooth; 3a, 3c, 4c (15–19) medium in length, almost smooth; 1b, 3b, 4a, 4b (51–103) long, weakly barbed. Pedotectum I large with smooth edge, narrow ridges running longitudinally along reverse side, depressed laterally; pedotectum II small. Custodium, discidium and circumpedal carina not distinct.
Anogenital region — (Figures 1B, 2E and 6A–C, E, F). Anogenital setal formula: 6-1-2-3. All genital (g1–g6: 37–48), aggenital (10–12), anal (an1, an2: 9–11) and adanal (ad1–ad3: 11–14) setae setiform, bent strongly, almost smooth. Ovipositor (length beyond middle fold 151–173, width 44–64) with three blades (65–78), each of blades with 4 setae (τ1, ψ1: 24–29; ψ2, τa, τb, τc: 10–15); six coronal setae (13–15) setiform. Lyrifissure iad located close to anterolateral margin of anal opening.

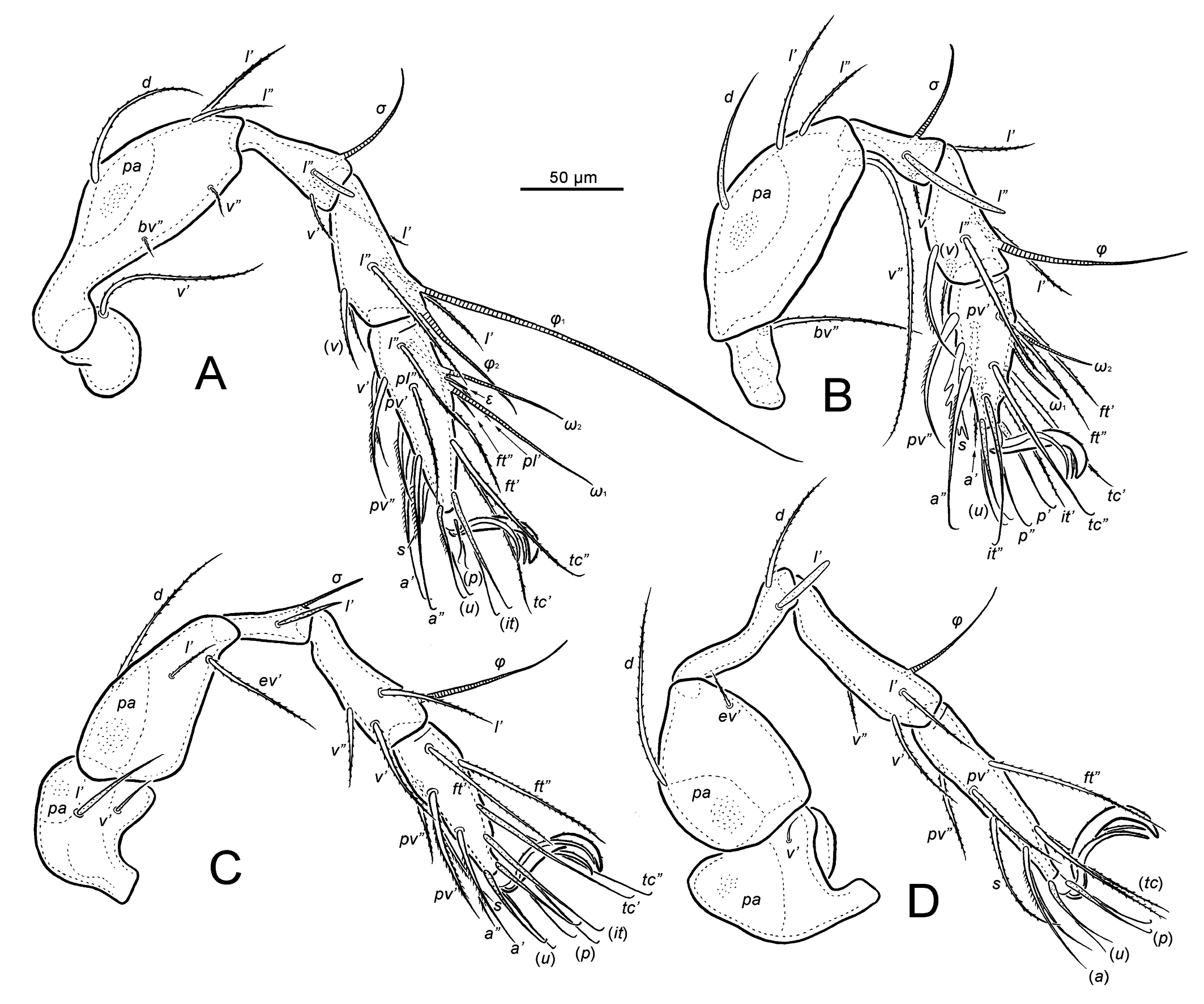
Legs — (Figures 3A–D, 6I and 7A–D). Tridactylous. All claws with small serrations on dorsal edge; lateral claws thinner than middle one. Porose area present on all femora and on trochanters III and IV. Measurements (total length from trochanter to tarsus, pretarsus excluded): I 345–392, II 325–354, III 348–386, and IV 405–434. Most setae filiform and barbed; seta s on tarsus II very thick, bearing several broad branches; setae l″ on genu I, II and setae l′ on genu III, IV clearly thick; setae ε and ft″ on tarsus I very short. Formula of leg setation (from trochanter to tarsus, including famulus, solenidia in brackets): I 1-5-3(1)-4(2)-20(2), II 1-5-3(1)-4(1)-15(2), III 2-3-1(1)-3(1)-15, IV 1-2-2-3(1)-12; homology of setae and solenidia indicated in Table 1.
Download as
Leg
Tr
Fe
Ge
Ti
Ta
I
v’
d, (l), bv”, v”
(l), v’, σ
(l), (v), φ1, φ2
(ft), (tc), (it), (p), (u), (a), s, (pv), v’, (pl), l”, ε, ω1, ω2
II
v’
d, (l), bv”, v”
(l),* v’, σ*
(l), (v), φ
(ft), (tc), (it), (p), (u), (a), s, (pv), ω1, ω2
III
l’, v’
d, l’, ev’
l’, σ
l’, (v), φ
(ft), (tc), (it), (p), (u), (a), s, (pv)
IV
v’
d, ev’
d,* l’*
l’, (v), φ
ft”, (tc), (p), (u), (a), s, (pv)
Immatures
Unknown.
Etymology
The specific name is from Bhutan.
Remarks
Achipteria bhutanica sp. nov. is similar to A. baleensis Ermilov, Rybalov & Kemal, 2011, A. catskillensis Nevin, 1977 and A. longesensillus Schweizer, 1956 in having broad lamellae with a single pointed lateral tooth. However, A. bhutanica sp. nov. differs from these species in the following characteristics: body size larger (604–672 × 466–540 vs. 558 × 378 in A. longesensillus; vs. 407–462 × 297–374 in A. catskillensis); surface of notogaster and anogenital region almost smooth (foveolate in A. baleensis; punctate in A. longesensillus); hypostomal setae present (absent in A. baleensis); rostral setae heavily barbed (slightly barbed in A. baleensis); lamellar setae not bending strongly (bending distally in A. longesensillus); heads of bothridial setae barbed (smooth in A. longesensillus and A. catskillensis); notogastral setae short (longer in A. baleensis); lateral tooth of pteromorphs present (absent in A. baleensis and A. catskillensis); lenticulus present (absent in A. catskillensis).
Discussion
Several surveys of the family Achipteriidae have been conducted in regions surrounding Bhutan. Four achipteriid species are known from the South Asia: Achipteria (A.) coleoptrata (Linnaeus, 1758) from India: Sikkim (Pearce 1906, as Oribata ovalis Nicolet, 1855); Anachipteria (A.) grandis Aoki, 1968 from Nepal (Ermilov and Martens 2014); Austrachipteria grandis (Hammer, 1967) from India: Bengal (Mondal and Kundu 1999b, as Parahypozetes); Parachipteria orientalis (Mondal & Kundu, 1999) from India: Bengal and Nepal (Mondal and Kundu 1999a, as Parahypozetes; Ermilov and Martens 2021). The occurrence of this family in Bhutan is not unexpected given its geographical proximity to surrounding regions.
Achipteriid mites are sometimes found in moss or wetlands, but mainly inhabit the forest or grassland soil (Bayartogtokh 2010; Bayartogtokh and Ryabinin 2012; Maruyama et al. 2016; Murvanidze and Mumladze 2016; Corpuz-Raros and Ermilov 2020; Schatz 2020; Behan-Pelletier and Lindo 2023). This new species also occurs in forest soils, but appears to have a habitat preference. Samples from a separate location near the type locality (approximately 700 meters away in straight-line distance from the type locality, 27°29′11″N, 89°21′42″E, 2,674 m a.s.l., 11 Oct. 2025, collected by M. Oshima from the litter) included no individuals of this species. The vegetation of this site was a pure Pinus wallichiana forest, with a somewhat wetter forest floor than that of the type locality. All of the other samples collected from various locations (from the semi-arid area to the cloud forest, 2,500–3,900 m a.s.l.) in Thimphu and Paro (except for the location where the paratypes were collected) also lack this species.
There are credible instances of the family (including genus Achipteria) occupying arboreal substrates such as tree bark, rarely epiphytic lichens and mosses, and sometimes a huge number of individuals are found (Aoki 1970; Behan-Pelletier and Winchester 1998; Lindo and Winchester 2006; Root et al. 2007; Lindo et al. 2008). Future research will focus on studying the forest across various vegetation types at different altitudinal zones within the Eastern Himalayan region of the Kingdom of Bhutan. This will include examining microhabitats involving vertical distribution within forests, from the litter layer to the canopy, with the goal of clarifying the habitat characteristics of this species and genus in the area.
Acknowledgments
We would like to express our special appreciation to Ms. Jigme Wangmo (NBC), who accompanied us and assisted with our investigation of the Tiger's Nest, and to Mr. Pema Khandu and Mr. Indra Prasad Acharja (Texas State University San Marcos, formerly affiliated with the Royal Society for Protection of Nature, Bhutan), who accompanied SS and assisted with the 2019 survey.
We are also grateful for the Research Permission code of the Kingdom of Bhutan (930275) that was granted for the research project in October 2025, and permission for the material transfer (NBC/BRD-7/2019-2020/ and PSC No. THD/BP/33; NBC/BRD-7/2025-2026/316).
This research was supported by research funds of the Asahi Glass Foundation and JSPS KAKENHI Grant Numbers 18K06392 for SS, and the Research Institute of Marine Invertebrates Grant Number KO2025-10 to MO.
References
- Aoki J. 1970. Descriptions of oribatid mites collected by smoking of trees with insecticides I. Mt. Ishizuchi and Mt. Odaigahara. Bull. Nat. Sci. Mus. Tokyo, 13(4): 585-602.
- Bayartogtokh B. 2010. Oribatid mites of Mongolia (Acari: Oribatida). Moscow: KMK. pp. 372.
- Bayartogtokh B., Ryabinin N.A. 2012. The soil mite family Achipteriidae (acari: oribatida) in Mongolia and Russian Far East. Acarologia, 52(2): 135-156. https://doi.org/10.1051/acarologia/20122044
- Bayoumi B.M., Mahunka S. 1979a. Ergebnisse der Bhutan-Expedition 1972 des Naturhistorischen Museums in Basel Acari: Oribatida (Part I). Entomol. Basil., 4: 13-24.
- Bayoumi B.M., Mahunka S. 1979b. Ergebnisse der Bhutan-Expedition 1972 des Naturhistorischen Museums in Basel Acari: Oribatida (Part II). Entomol. Basil., 4: 24-30.
- Behan-Pelletier V.M., Winchester N.N. 1998. Arboreal oribatid mite diversity: colonizing the canopy. Appl. Soil Ecol., 9: 45-51. https://doi.org/10.1016/S0929-1393(98)00052-3
- Behan-Pelletier V.M., Lindo Z. 2023. Chapter 2. Form and function. In: Behan-Pelletier V., Lindo Z. (Eds.). Oribatid Mites: Biodiversity, Taxonomy and Ecology. Boca Raton, Florida: CRC Press. p. 7-32. https://doi.org/10.1201/9781003214649
- Chakrabarti D.K., Wilson R. 1981. Preliminary studies of the soil oribatid mites (Acari) from Bhutan. Sci. Cult., 47: 132-134.
- Corpuz-Raros L., Ermilov S.G. 2020. Catalogue of oribatid mites (Acari: Oribatida) from Continental Southeast Asia. Zootaxa, 4893: 1-216. https://doi.org/10.11646/zootaxa.4893.1.1
- Ermilov S.G., Kontschán J. 2025. Taxonomic contribution to knowledge of the oribatid mite genus Achipteria (Acari, Oribatida, Achipteriidae). Int. J. Acarol., 51(2): 81-87. https://doi.org/10.1080/01647954.2024.2439799
- Ermilov S.G., Martens J. 2014. Additions to the Nepalese oribatid mite fauna, with description of two new species (Acari, Oribatida). Int. J. Acarol., 40(2): 123-132. https://doi.org/10.1080/01647954.2013.870227
- Ermilov S.G., Martens J. 2021. New faunistic and taxonomic data on oribatid mites of Nepal (Acari, Oribatida). Spixiana, 44(2): 229-236.
- Grandjean F. 1932. Observations sur les Oribates (3e série). Bull. Mus. Natl. Hist. Nat., 4: 292-306.
- Grandjean F. 1935. Observations sur les Oribates (9e série). Bull. Mus. Natl. Hist. Nat., 8: 280-287.
- Gyeltshen C., Prasad K. 2022. Biodiversity checklists for Bhutan. Biodivers. Data J., 10: e83798. https://doi.org/10.3897/BDJ.10.e83798
- Lindo Z., Winchester N.N. 2006. A comparison of microarthropod assemblages with emphasis on oribatid mites in canopy suspended soils and forest floors associated with ancient western red cedar trees. Pedobiologia, 50: 31-41. https://doi.org/10.1016/j.pedobi.2005.09.002
- Lindo Z., Clayton M., Behan-Pelletier V.M. 2008. Systematics and ecology of Anachipteria geminus sp. nov. (Acari: Oribatida: Achipteriidae) from arboreal lichens in western North America. Can. Entomol., 140(5): 539-556. https://doi.org/10.4039/n08-016
- Maruyama I., Bayartogtokh B., Shimano S. 2016. Rediscovery of Achipteria setulosa, with remarks on Japanese species of Achipteriidae and the proposal of species-groups (Acari, Oribatida). Zookeys, 578: 1-13. https://doi.org/10.3897/zookeys.578.7603
- Mondal B.K., Kundu B.G. 1999a. A new cryptostigmatid mite (Acari: Oribatei) and a few new records of cryptostigmatid fauna from forest and tea soils in Jalpaiguri District, West Bengal, India. Rec. Zool. Surv. India, 97(1): 123-132. https://doi.org/10.26515/rzsi/v97/i1/1999/160259
- Mondal B.K., Kundu B.G. 1999b. On a collection of oribatid fauna (Acari: Oribatei) from forest and tea soils in in Jalpaiguri District, West Bengal, India. Rec. Zool. Surv. India, 97(2): 79-86. https://doi.org/10.26515/rzsi/v97/i2/1999/160204
- Murvanidze M., Mumladze L. 2016. Annotated checklist of Georgian oribatid mites. Zootaxa, 4089: 1-81. https://doi.org/10.11646/zootaxa.4089.1.1
- National Environment Commission, Royal Government of Bhutan (NECRGB). 2022. First Biennial update to the UNFCCC, 2022 [Internet]. [Nov. 2022]. Thimphu: National Environment Commission, Royal Government of Bhutan; [15 Nov. 2025]. Available from: https://unfccc.int/sites/default/files/resource/BhutanBURreport.pdf
- National Biodiversity Center. 2017. Biodiversity Statistics of Bhutan 2017: A Preliminary Baseline [Internet]. [22 May 2019] Thimphu: National Biodiversity Center, Ministry of Agriculture and Forests; [15 Nov. 2025]. Available from: https://bt.chm-cbd.net/sites/bt/files/inline-files/Biodiversity%20Statistics%20of%20Bhutan%202017.pdf
- Norton R.A. 1977. A review of F. Grandjean's system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal D.L. (Ed.). Biology of oribatid mites. Syracuse: SUNY College of Environmental Science and Forestry. p. 33-61.
- Norton R.A., Behan-Pelletier V.M. 2009. Chapter 15. Suborder Oribatida. In: Krantz, G.W., Walter D.E. (Eds.). A Manual of Acarology, Third Edition. Lubbock, Texas: Texas Tech University Press. p. 430-564.
- Ohsawa M. 1987. Vegetation zones in the Bhutan Himalaya. In: Ohsawa M. (Ed.). Life Zone Ecology of the Bhutan Himalaya. Chiba: Chiba University. p. 1-71.
- Pearce N.D.F. 1906. On some Oribatidae from Sikkim Himalaya. J. Roy. Mic. Soc., 26(3): 269-273. https://doi.org/10.1111/j.1365-2818.1906.tb01643.x
- Reddy A.S. 1989. Studies on the oribatid fauna of Bhutan -1. Euphthiracarus bhutanicus sp. n. from Kanglung. Prog. Acarol., 1: 499-502.
- Root H.T., Kawahara A.Y., Norton R.A. 2007. Anachipteria sacculifera n. sp. (Acari: Oribatida: Achipteriidae) from arboreal lichens in New York State. Acarologia, 47(3-4): 173-181.
- Sanyal A.K. 1988. Two new species of the genus Scheloribates (Acarina: Oribatei) from Bhutan. Rec. Zool. Surv. India, 85(2): 237-243. https://doi.org/10.26515/rzsi/v85/i2/1988/161054
- Schatz H. 2020. Catalogue of oribatid mites (Acari: Oribatida) from Vorarlberg (Austria). Zootaxa, 4783: 1-106. https://doi.org/10.11646/zootaxa.4783.1.1
- Seniczak S., Seniczak A. 2016. Morphological ontogeny of Achipteria gigantea sp. nov. (Acari: Oribatida: Achipteriidae) from northern Spain, with comments on Achipteria. Syst. Appl. Acarol., 21: 1571-1590. https://doi.org/10.11158/saa.21.11.12
- Subías L.S. 2004. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes, Oribatida) del mundo (1758-2002). Graellsia, 60: 3-305. https://doi.org/10.3989/graellsia.2004.v60.iExtra.218
- Subías L.S. 2024. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles), unpublished 19a actualización [Internet]. [Jan. 2024]. Madrid: Universidad Complutense de Madrid; [23 Nov. 2025]. Available from: http://bba.bioucm.es/cont/docs/RO_1.pdf
- Walcott S. 2009. Geographical field notes urbanization in Bhutan. Geogr. Rev., 99(1): 81-93. https://doi.org/10.1111/j.1931-0846.2009.tb00419.x
- Wangchuck Centennial National Park and World Wildlife Fund (WCNP and WWF). 2016. Population Status and distribution of snow leopards in Wangchuck Centennial National Park, Bhutan [Internet]. [5 Oct. 2016]. Thimphu: Wangchuck Centennial National Park and World Wildlife Fund; [22 Nov. 2025]. Available from: https://wwfasia.awsassets.panda.org/downloads/wwf_sl_report_05_10_2016_super_final_compressed_1.pdf
- Wangda P., Ohsawa M. 2006. Gradational forest change along the climatically dry valley slopes of Bhutan in the midst of humid Himalayan environment. Plant Ecol., 186 (1): 109-128. https://doi.org/10.1007/s11258-006-9116-5
- Weigmann G. 2006. Hornmilben (Oribatida). In: Dahl F. (Ed.). Die Tierwelt Deutschlands part 76. Keltern: Goecke & Evers. p. 1-520.



2026-03-24
Date accepted:
2026-05-26
Date published:
2026-06-08
Edited by:
Pfingstl, Tobias

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Oshima, Masaharu; Darjee, Beejai; Dorji, Tshering; Choden, Tshering; Pelyang, Tashi; Tshewang, Tshewang; Yamauchi, Takeo; Nomura, Shûhei; Nakano, Takafumi; Dorji, Karma Dema and Shimano, Satoshi
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)