



Torrenticolid water mites of Bhutan. Genera Torrenticola Piersig, 1896 and Neoatractides Lundblad, 1941 (Acari: Hydrachnidia: Torrenticolidae)
Pešić, Vladimir  1
; Smit, Harry
2
and Gurung, Mer Man
3
1
; Smit, Harry
2
and Gurung, Mer Man
3
1✉ University of Montenegro, Cetinjski put bb, 81000 Podgorica, Montenegro.
2Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, the Netherlands.
3College of Natural Resources, Royal University of Bhutan, Lobesa, Punakha, P.O. Box 13001, Bhutan.
2022 - Volume: 62 Issue: 3 pages: 821-860
https://doi.org/10.24349/xn0u-5px2ZooBank LSID: 251E7093-B080-4E76-AA42-F285E211B055
Original research
Keywords
Abstract
Introduction
Torrenticolidae are common and distinctive water mites found worldwide, except in Antarctica. The family includes six genera, Torrenticola Piersig, 1896, Monatractides Viets, 1926, Testudacarus Walter, 1928; Pseudotorrenticola Walter, 1906; Neoatractides Lundblad, 1941, and Stygotorrenticola Pešić & Gerecke, 2014. Most of the torrenticolid mites inhabit lotic habitats (Di Sabatino et al. 2010, Pešić and Smit 2009, 2014), but some species have been reported from lentic habitats as well from interstitial waters (Di Sabatino et al. 2010). In fast flowing streams, torrenticolid mites are often the most dominant component.
The water mite fauna of the Himalayas is still insufficiently known. A recent paper of Gerecke and Smit (2022) gives an overview of previous research on water mites of the Himalayas and its foothills. More recently, aquatic biota of the Mangde Chhu river is threatened by the construction of the Mangde Chhu hydropower plant (Wangchuk et al. 2017) which accelerates the need for proper documentation of the invertebrate fauna, including water mites.
So far, only three torrenticolid mite species, i.e., Torrenticola bhutana Pesic & Smit 2007, Testudacarus tripeltatus Walter, 1928 and Monatractides tristis (Lundblad, 1941) were known from Bhutan (Pesic and Smit 2007). As a result of the present study 17 species of the genera Torrenticola and Neoatractides were identified. In this paper, descriptions of seven new species of the genus Torrenticola and one new species of the genus Neoatractides are given.
Material and methods
Water mites were collected by hand netting, sorted live in the field, and fixed in Koenike-fluid. The material of the present study is collected by the junior author (M.M.G) in 2021 in Bhutan, a landlocked country in the Eastern Himalayas. Most of the material has been collected in tributaries of the Mangde Chhu river in central Bhutan, during two collection trips, one pre-monsoon (April-May 2021) and one post-monsoon (October-November 2021). The latter river belongs to the basin of the Manas River, which drains almost all the basins of central and eastern Bhutan. Unless stated otherwise, all material is collected by Mer Man Gurung, and this is not repeated in the text. All material will be deposited in the National Biodiversity Centre, Thimphu (NBC), a part of the paratypes and the non-type material will be lodged as a long-term loan in Naturalis Biodiversity Center, Leiden (RMNH). In the section ′Material examined', collecting site abbreviations derive from the geographical database of Mer Man Gurung.
All measurements are given in μm. Measurements of palp and legs are of dorsal margin. Frontal and shoulder plates are measured on both sides and their values are given as a range. The photographs of selected structures were made using a camera on Samsung Galaxy smartphone. The following abbreviations are used: Ac-1 = first acetabulum; Cx-I–IV = first to fourth coxae; Cxgl-2–4 = coxoglandularium 2–4; dL = dorsal length; H = height; I-L-4–6 = fourth-sixth segments of first leg; L = length; mL = medial length; P-1–P-5 = palp segments 1–5; Vgl-1–2 = ventroglandularium 1–2; vL = ventral length; W = width.
Systematics
Family Torrenticolidae Piersig, 1902
Genus Torrenticola Piersig, 1896
Subgenus Torrenticola s.str
Torrenticola bhutana Pesic & Smit, 2007
Figures 1A, G
Material examined — Bhutan, MG10 Chumpigang Chhu, 27.31608°N, 90.58071°E, 1018 m asl, 8.v.2021, 1♂, 1♀ (juvenile) (1♂ dissected and slide mounted).
Description — General features – Idiosoma elongated-oval (dorsal shield L/W ratio 1.3); dorsal shield without a colour pattern (as photographed in Figure 1A); area of primary sclerotization of the dorsal plate with two dorsoglandularia; gnathosomal bay U-shaped, proximally rounded; Cxgl-4 subapical; medial suture line of Cx-II+III short; suture lines of Cx-IV in both sexes distinctly extending posteriorly beyond posterior margin of genital field, laterally curved; excretory pore and Vgl-2 close to the line of primary sclerotization, excretory pore on the level of Vgl-2; gnathosoma with a strongly curved ventral margin, rostrum elongated; P-2 longer than P-4; P-2 ventral margin straight or slightly convex, P-2 and P-3 ventrodistal protrusions bluntly pointed, P-4 with a well-developed ventral tubercle bearing one longer and three shorter setae). Male (in parentheses measurements from original description; Pesic and Smit 2007) – Idiosoma L/W 719 (875)/549 (625); dorsal shield L/W 570 (706)/444 (531); medial suture Cx-II+III L 71 (61); genital field subrectangular, L/W 156 (172)/119 (116); ejaculatory complex characteristic, proximal and distal arms short, proximal chamber large, proximal horns reduced (Fig. 1G); P-2 98 (98), P-4 77 (85).
Remarks — This species was originally described on basis of specimens collected from a stream at the Pele La mountain pass in Bhutan at an elevation of 3200 m asl (Pesic and Smit 2007).
Distribution — Bhutan (Pesic and Smit 2007; this study).
Torrenticola episce Pešić, Chatterjee, Kumar Das & Bordoloi, 2013
Figures 1B, 6D

Material examined — Bhutan, MG3 Bipgang Chhu, 27.15729°N, 90.66721°E, 586 m asl, 28.vi.2021, 2♀ (1♀ dissected and slide mounted).
Description — General features – Idiosoma roundish (dorsal shield L/W ratio 1.2–1.3); shoulder platelets fused with dorsal plate, but suture line visible; area of primary sclerotization of the dorsal plate with two dorsoglandularia; gnathosomal bay U-shaped, proximally rounded; Cxgl-4 posterior to Cxgl-2; medial suture line of Cx-II+III relatively short, 81 µm in ♂ holotype; excretory pore and Vgl-2 clearly posterior to the line of primary sclerotization; excretory pore slightly anterior to Vgl-2; suture of Cx-IV curved. Gnathosoma ventral margin curved, rostrum well developed; P-2 ventrodistal protrusion slender; ventral margins of P-2, -3 and -4 with fine, indistinct serrations; P-2 longer than P-4; P-4 with visible ventral tubercles (Figure 6D). Female (in parentheses measurements from original description; Pešić et al. 2013) – Idiosoma L 769 (806–850), W 625 (663–687); dorsal shield L 625 (700–713), W 519 (556); medial suture Cx-II+III L 29 (16–38); genital field L 178 (64–166), W 138 (136–137); L P-2 117 (120–128), P-4 95 (105–109).
Remarks — This species was described by Pešić et al. (2013) on basis of specimens found in the gut contents of the fish species Botia dario from Assam, India.
Distribution — India (Assam), Bhutan (this study).
Torrenticola dentifera Wiles, 1991
Figures 1C, 2
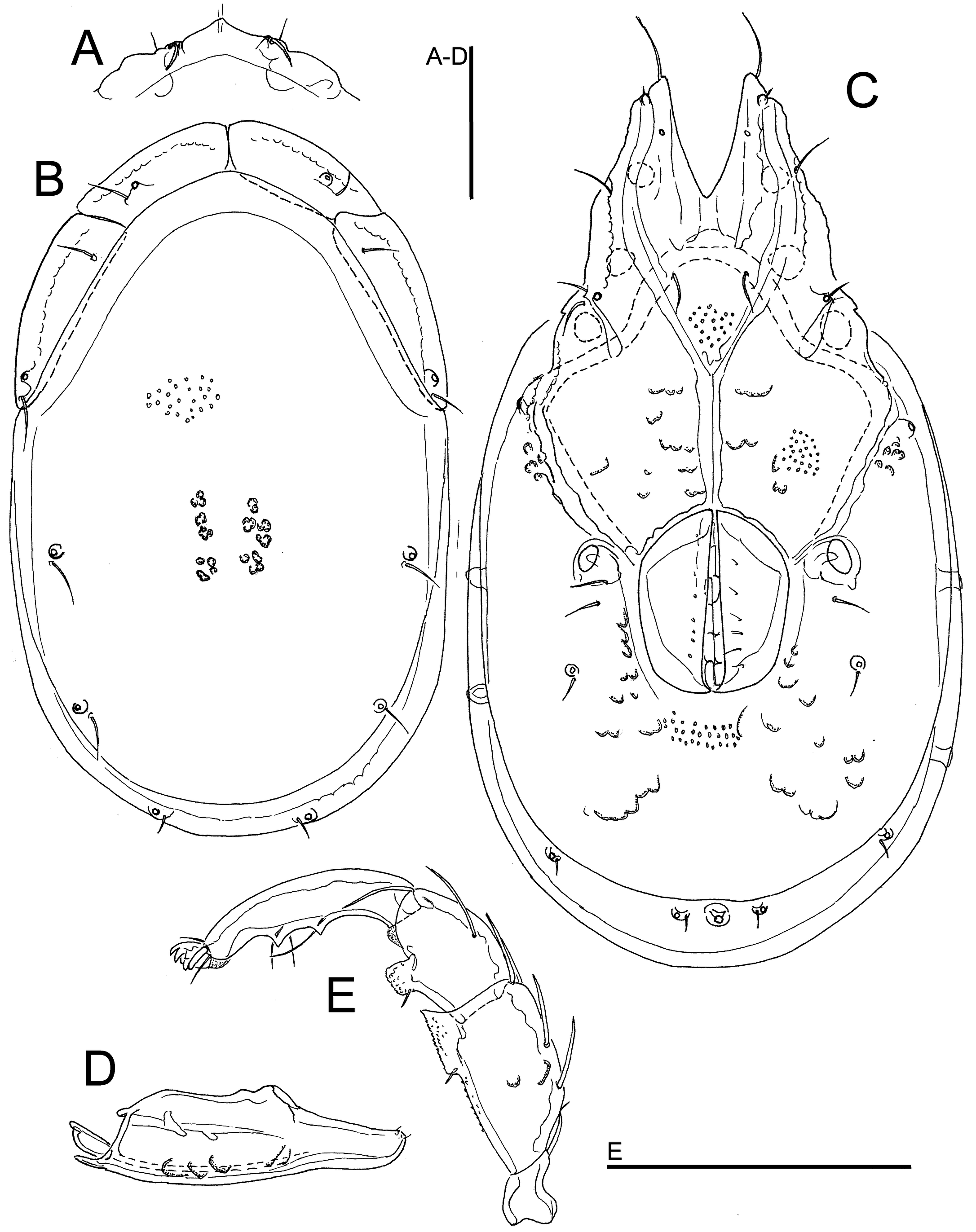
Material examined — Bhutan, MG2 Berti Chhu, 27.16264°N, 90.66003°E, 590 m asl, 25.x.2021, 1♀ (dissected and slide mounted).
Description — General features – Idiosoma elongated (dorsal shield L/W ratio 1.6); dorsal shield with a colour pattern as illustrated in Figure 1C; gnathosomal bay V-shaped (Figure 2C); Cxgl-4 subapical; medial suture line of Cx-II+III long; posterior suture line of Cx-IV in its medial part perpendicular to the longitudinal idiosoma axis, laterally distinctly curving anteriorly; excretory pore and Vgl-2 close to the line of primary sclerotization; gnathosoma ventral margin only slightly curved, rostrum well developed (Figure 2D); P-2 shorter than P-4, ventral margin of P-2 with a fine denticulation also in proximal half of the segment, distally with a laterally compressed, anteriorly directed hyaline extension and a very short, denticle-like seta laterally at base of projection; P-3 with a broad, subrectangular, apically serrated ventrodistal projection with fine denticles, and a short seta laterally at base of projection; P-4 ventral tubercles well developed and separated (Figure 2E). Female – Idiosoma L/W 594/328; dorsal shield L/W 475/292; medial suture Cx-II+III L 88; genital field L/W 122/100; L P-2 57, P-4 72.
Remarks — This species was described by Wiles (1991) on basis of two males from Selangor, Peninsular Malaysia. In the original description no information on the colour pattern of the dorsal shield was given (Wiles 1991). Later on, T. dentifera has been reported from Wuyishan, China (Gu et al. 2020a).
Distribution — Malaysia, China, Bhutan (this study).
Torrenticola dolichorrhyncha (Lundblad, 1952)
Figures 3, 4


Material examined — Bhutan: MG8 Dangdung Chhu, 27.33461°N, 90.59562°E, 1039 m asl, 30.x.2021 1♂ (dissected and slide mounted); MG9 Kartigang Chhu, 27.27896°N, 90.63088°E, 1456 m asl, 7.v.2021, 1♂, 1♀ (1♂ dissected and slide mounted); MG12 Nika Chhu, 27.526010°N, 90.29947°E, 2609 m asl., 4.xi.2021, 7♂, 6♀ (2♂ dissected and slide mounted); MG12 Nika Chhu, 27.526010°N, 90.29947°E, 2609 m asl, 4.xi.2021, 4♀; MG12 Nika Chhu, 27.526010°N, 90.29947°E, 2609 m asl, 10.v.2021 3♂, 3♀; MG12 Nikka Chhu, 27.526010°N, 90.29947°E, 2609 m asl, 10.v.2021, 2♂, 4♀; MG13 Rukhubji Chhu, 27.51174°N, 90.29711°E, 2587 m asl, 5.xi.2021, 1♂, 1♀; MG13 Rukhubji Chhu, 27.51174°N, 90.29711°E, 2587 m asl, 12.v.2021 3♂, 5♀; MG14 Chuserbu stream, 27.50246°N, 90.31782°E, 2666 m asl., 6.xi.2021, 2♂, 9♀; MG14 Chuserbu stream, 27.50246°N, 90.31782°E, 2666 m asl, 13.v.2021, 4♂, 4♀ (2♂ dissected and slide mounted); MG15 Khabab Chhu, 27.48492°N, 90.33490°E, 2500 m asl, 7.xi.2021, 1♂ (damaged; palps lacking), 2♀ (1♀ dissected and slide mounted); MG15 Khabab Chhu, 27.48492°N, 90.33490°E, 2500 m asl, 14.v.2021, 3♂, 4♀; MG16 Chendebji Chhu, 27.47692°N, 90.35265°E, 2483 m asl, 8.xi.2021 6♂, 6♀; Royal Botanical Park, Lampelri, 27.50750°N, 89.75269°E,2667 m asl, 10.viii.2016 leg. W. Klein, 6♂; springs in meadow E of Takchu, 27.35820°N, 89.286310°E, 2907 m asl, 10.viii.2016 leg. W. Klein, 3♂; Pool 3, Haa Chhu River, Domcho, 27.36472°N, 89.29564°E, 2675 m asl, 10.viii.2016 leg. W. Klein, 7♂, 4♀ (1♂ dissected and slide mounted).
Diagnosis — General features – Idiosoma roundish; dorsal shield with a colour pattern as given in Figures 4A, B; area of primary sclerotization of the dorsal plate with two dorsoglandularia; gnathosomal bay U-shaped, proximally rounded; Cxgl-4 subapical; medial suture line of Cx-II+III relatively short; excretory pore located away from the line of primary sclerotization; gnathosoma ventral margin strongly curved, rostrum strongly elongated (Figure 4E); P-2 longer than P-4; P-2 ventral margin straight, P-2 and P-3 ventrodistal protrusions blunt, laterally flattened, P-4 with a well-developed ventral tubercle bearing one longer and three shorter setae (Figure 3C). Male (measurements of one specimen from Chuserbu stream; in parentheses measurements from Lundblad 1969) – Idiosoma L 788 (759), W 581; dorsal shield L/W 647 (612)/525; medial suture Cx-II+III L 100 (107); genital field subrectangular, L/W 172(175)/128(122); suture line of Cx-IV evident, medially starting from posterior margin of genital field in a right angle to the main idiosoma axis; genital field subrectangular; ejaculatory complex conventional in shape, anterior keel, proximal and distal arms well developed (Figure 4C); L P-2 106 (103), P-4 91 (93). Female (measurements of one specimen from Khabab Chhu) – Idiosoma L 872 (758), W 675; dorsal shield L/W 700(620)/600; medial suture Cx-II+III L 63; genital field L/W 197 (168)/161 (121); suture lines of Cx-IV extending posteriorly beyond posterior margin of genital field, laterally curved; excretory pore and Vgl-2 away from the line of primary sclerotization; L P-2 106 (99), P-4 89 (94).
Remarks — The specimen from Bhutan matches the description of T. dolichorrhyncha, a species originally described on basis of specimens collected from a stream at the Kambaiti mountain pass in Myanmar (Lundblad 1941, 1969).
Distribution — Myanmar, Bhutan (this study).
Torrenticola kumari Pešić & Smit, 2020
Figures 1D–H, 5

Material examined — Bhutan, MG2 Berti Chhu, 27.16264°N, 90.66003°E, 590 m asl, 25.x.2021, 3♂ (1♂ juvenile), 3♀ (1♀ juvenile) (1♂ dissected and slide mounted); ibid., 26.iv.2021, 1♂, 1♀; MG5 Dakpay Chhu, 27.14621°N, 90.69220°E, 539 m asl, 1.v.2021 1♀ (juvenile), dissected and slide mounted.
Diagnosis — General features – Idiosoma oval; dorsal shield with a colour pattern as illustrated in Figure 1D; area of primary sclerotization of the dorsal plate with two dorsoglandularia; gnathosomal bay U-shaped, proximally rounded; Cxgl-4 subapical; medial suture line of Cx-II+III relatively short; suture lines of Cx-IV in the both sexes distinctly extending posteriorly beyond posterior margin of genital field, laterally curved; excretory pore and Vgl-2 away from the line of primary sclerotization, excretory pore on the level of Vgl-2; gnathosoma with a strongly curved ventral margin; P-2 and P-4 almost equal in length; P-2 ventral margin straight, ventral seta relatively long, P-2 and P-3 ventrodistal protrusions pointed, P-2 ventrodistal extension curved distally, P-4 with a well-developed ventral tubercle bearing one longer and three shorter setae (Figure 5D). Male (measurements of one specimen from Berti Chhu; in parentheses measurements from original description; Pešić et al. 2020) – Idiosoma L 634 (700), W 502 (565); dorsal shield L 553 (580), W 416 (469); medial suture Cx-II+III L 62 (72); genital field with obtuse, but distinct anterior angles, L/W 140 (163)/112 (127); ejaculatory complex conventional in shape, anterior keel, proximal and distal arms well developed (Figure 1H); P-2 91 (105), P-4 91 (102).
Remarks — The examined specimens from Bhutan generally match the description of T. kumari Pešić & Smit, 2020, a species recently described from Garhwal Himalayas in Uttarakhand State of India (Pešić et al. 2020). The populations of the latter species from India were originally assigned to T. semisuta Halík, 1930, a species described from Pahang, Peninsular Malaysia (Halík 1930), from which it differs in a larger size and a different colour pattern of the dorsal shield (see Pešić et al. 2020 for a discussion).
Distribution — India (Uttarakhand), Bhutan (this study).
Torrenticola lamellifera (Lundblad, 1941)
Figures 1E–I, 6A, B
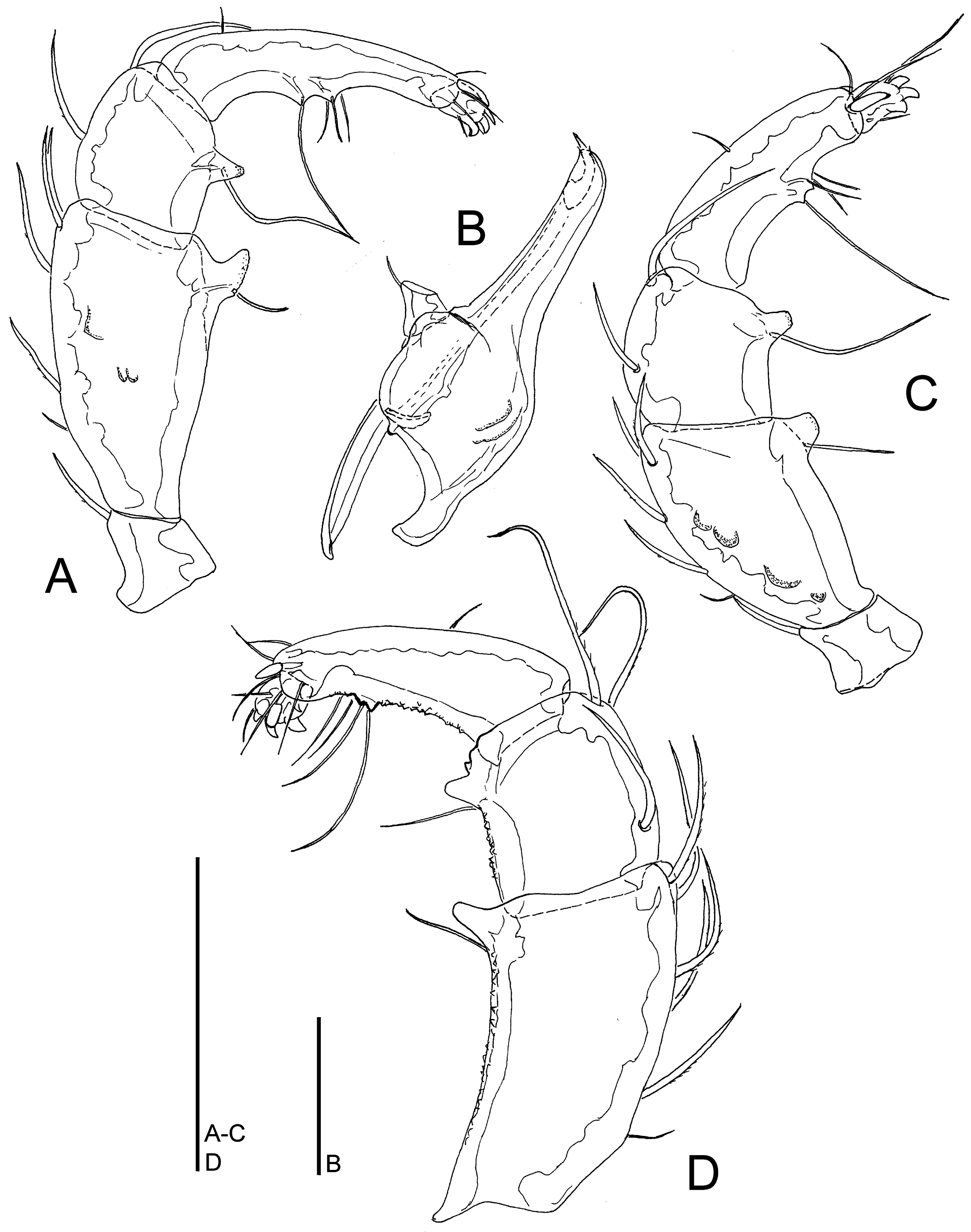
Material examined — Bhutan, MG1 Maidagang Chhu, 27.12761°N, 90.71560°E, 554 m asl, 20.x.2021, 1♂, 3♀ (1♂ dissected and slide mounted); MG3 Bipgang Chhu, 27.15729°N, 90.66721°E, 596 m asl, 28.vi.2021, 1♂; MG4 Takabi Chhu, 27.14782°N, 90.68833°E, 543 m asl, 30.vi.2021, 1♀; MG4 Takabi Chhu, 27.14782°N, 90.68833°E, 543 m asl, 26.x.2021, 1 ♂ (dissected and slide mounted)
Description — General features – Idiosoma oval; dorsal shield with a colour pattern as illustrated in Figure 1E; gnathosomal bay U-shaped, proximally pointed; Cxgl-4 subapical; medial suture line of Cx-II+III relatively short; posterior suture lines of Cx-IV extending posteriorly beyond posterior margin of genital field, laterally curved; excretory and Vgl-2 close to the line of primary sclerotization pore; gnathosomal rostrum long and slender, ventral margin in lateral view curved (Figure 6B); P-2 with a laterally compressed, longish (< 30% of ventral margin), anteriorly directed ventrodistal extension, P-3 with a short, subrectangular, apically serrated ventrodistal projection, and a long seta laterally at base of projection; P-4 slender with a well-developed ventral tubercle ending in two tips separated by a concavity, bearing one longer and three shorter setae (Figure 6A). Male (measurements of one specimen from Maidagang Chhu) – Idiosoma L/W 581/447; dorsal shield L/W 459/363; medial suture Cx-II+III L 59; genital field L/W 133/106, subrectangular; male ejaculatory complex conventional in shape, with well-developed anterior keel and proximal arms (Figure 1I).
Remarks — Torrenticola lamellifera was originally described from a single male from a stream in Dawna Hills in eastern Myanmar (Lundblad 1941, 1969). Later on, Wiles (1997) proposed to synonymize T. lamellifera with T. malaisei. Pešić et al. (2012) suggested that T. lamellifera should stand as separate species differing from T. malaisei in a longer and more slender P-4 and Cx-IV posteriorly distinctly extending posterior to the genital field.
Habitat — In Bhutan this species was collected from streams at medium altitude (500–600 m asl.). Pešić et al. (2012) reported this species from a high-elevation stream (< 1900 m asl.) in the eastern Himalayan region of India (Arunachal Pradesh).
Distribution — Burma (Lundblad 1941, 1969), Thailand (Pešić and Smit 2009), India (Arunachal Pradesh; Pešić et al. 2012), Bhutan (this study).
Torrenticola malaisei (Lundblad, 1941)
Figures 7, 24A, B, F

Material examined — Bhutan, MG8 Dangdung Chhu, 27.33461°N, 90.59562°E, 1039 m asl, 30.x.2021 1♂, 6♀ (1♂, 1♀ dissected and slide mounted).
Description — General features – Idiosoma elongated-oval (dorsal shield L/W ratio 1.3-1.4); dorsal shield with a colour pattern as illustrated in Figures 24A, B; gnathosomal bay U-shaped; Cxgl-4 subapical; medial suture line of Cx-II+III short; posterior suture lines of Cx-IV extending posteriorly beyond posterior margin of genital field, laterally curved; excretory pore and Vgl-2 away from the line of primary sclerotization; gnathosomal rostrum long and slender, ventral margin in lateral view curved (Figure 7E); P-2 longer than P-4, P-2 with a laterally compressed, longish (< 30% of ventral margin), anteriorly directed ventrodistal extension; P-3 with a short, subrectangular, apically serrated ventrodistal projection, and a long seta laterally at base of projection; P-4 short, with two small peg-like ventral tubercles separated by a concavity, bearing one longer and three shorter setae (Figures 7C-D). Male (measurements of one specimen from Dangdung Chhu) – Idiosoma L/W 644/444; dorsal shield L/W 516/377; medial suture Cx-II+III L 73; genital field subrectangular, L/W 125/95; male ejaculatory complex conventional in shape, with well-developed anterior keel and proximal arms (Figure 24F); L P-2 84, P-4 60. Female (measurements of one specimen from Dangdung Chhu; in parentheses measurements from Lundblad 1969) – Idiosoma L 746 (724), W 519; dorsal shield L 590 (578), W 440; medial suture Cx-II+III L 49 (57); genital field wider than in male and slightly tapering posteriorly, L/W 153 (157)/138(142); L P-2 95 (95), P-4 70 (81).
Remarks — The specimens from Bhutan match the description of T. malaisei, a species described by Lundblad (1941) on basis of a female collected in a stream at the Kambaiti mountain pass in Myanmar (Burma).
Distribution — Myanmar, Bhutan (this study).
Torrenticola uttarkhandensis Pešić & Smit, 2019
Figures 1F, J, 8
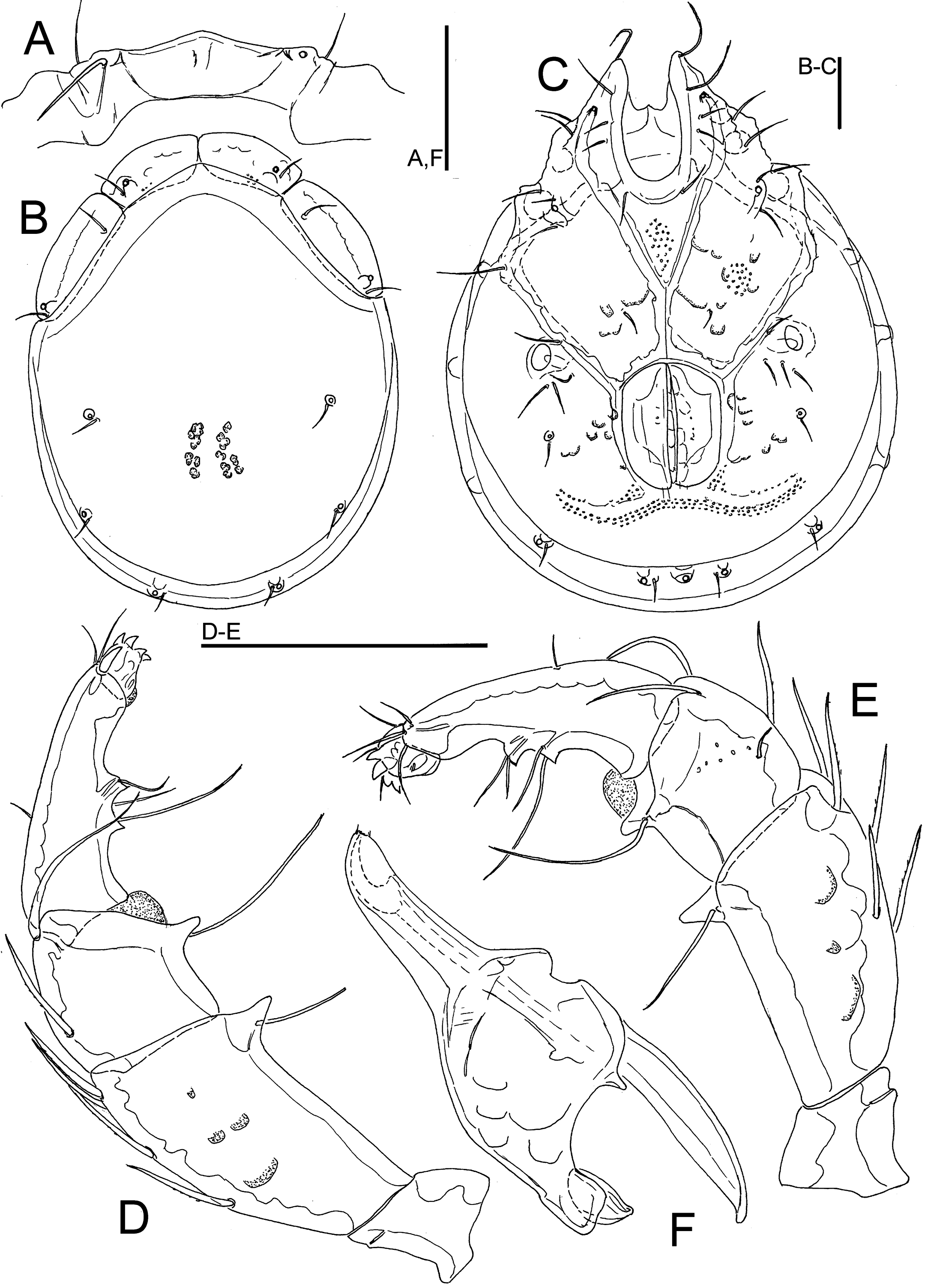
Material examined — Bhutan, MG4 Takabi Chhu, 27.14782°N, 90.68833°E, 543 m asl, 30.vi.2021, 1 ♂, 1♀ (1 ♂ dissected and slide mounted).
Description — Male (female unknown; in parentheses measurements from Pešić et al. 2019) – Idiosoma L 781 (820), W 619 (644), roundish (dorsal shield L/W ratio 1.29); dorsal shield, L/W 669 (678)/519 (527), with colour pattern as illustrated in Figure 1F; area of primary sclerotization of the dorsal plate with four dorsoglandularia; gnathosomal bay U-shaped, proximally rounded; Cxgl-4 posterior at margin of Cx-I/II, near insertion of I-L (Figure 8C); medial suture Cx-II+III L 99 (103); suture line of Cx-IV distinct, medially starting from posterior margin of genital field in a right angle to the main idiosoma axis; genital field subrectangular in shape, L/W 172 (172)/136 (144); ejaculatory complex conventional in shape, anterior keel, proximal and distal arms well developed (Figure 1J); excretory pore and Vgl-2 located close to the line of primary sclerotization, excretory pore on the level with Vgl-2; gnathosoma with strongly curved ventral margin, rostrum elongated (Figure 8F); P-2 longer than P-4, L P-2 107 (113), P-4 97 (103), P-2 ventral margin almost straight, P-2 and P-3 ventrodistal protrusions bluntly pointed and cone-shaped, P-4 with well-developed ventral protuberances, separated by a concavity (Figures. 8D, E).
Remarks — The single male from Bhutan generally matches the description of T. uttarkhandensis, a species described on basis of a male specimens collected in a stream in the Garhwal Himalaya of India (Pešić et al. 2019a). In regard to an elongated gnathosomal rostrum and Cxgl-4 shifted posterior to I-L insertion, the latter species resembles Torrenticola birmana (Lundblad, 1941) from which it differs by the ventral projections of P-2-3 not pointed, P-4 tubercles widely separated, and in a different colour pattern of the dorsal shield (Pešić et al. 2019a).
Distribution — India (Uttarakhand State; Pešić et al. 2019a); Bhutan (this study).
Torrenticola cf. turkestanica (Sokolow, 1926)
Figures 6C, 9

Material examined — Bhutan, MG1 Maidagang Chhu, 27.12761°N, 90.71560°E, 554 m asl, 20.x.2021, 1♂ (dissected and slide mounted); MG4 Takabi Chhu, 27.14782°N, 90.68833°E, 543 m asl, 26.x.2021, 7♂ (1♂ dissected and slide mounted); MG5 Dakpay Chhu, 27.14621°N, 90.69220°E, 539 m asl, 27.x.2021, 5♂, 4♀ (1♂ dissected and slide mounted); MG5 Dakpay Chhu, 27.14621°N, 90.69220°E, 539 m asl, 1.v.2021 5♂, 1♀
Description — General features – Idiosoma oval; dorsal shield with a colour pattern as illustrated in Figures 9A-C; gnathosomal bay U-shaped, proximally pointed; Cxgl-4 subapical; posterior suture lines of Cx-IV medially starting from the genital field at a 45°angle to main idiosoma axis, distinctly extending posteriorly beyond posterior margin of genital field; excretory pore and Vgl-2 well away from the line of primary sclerotization, excretory pore on the level of Vgl-2; ejaculatory complex conventional, with well-developed anterior keel and distal and proximal arms (Figure 9J); gnathosoma with a curved ventral margin (Figure 9L); P-2 and P-3 with a subrectangular ventrodistal projection, P-2 ventral seta relatively long, P-4 stocky with a well-developed ventral tubercle ending in two tips separated by a concavity, bearing one longer and three shorter setae (Figure 6C). Male (measurements of one specimen from Maidagang Chhu; in parentheses measurements of T. cf. turkestanica from Uttarakhand, India, from Pešić et al. 2019a) – Idiosoma L 606 (656), W 463 (486); medial suture Cx-II+III L 81 (94); genital field L/W 147 (145)/113 (116); L P-2 (83), P-4 (88).
Remarks — The examined specimens from Bhutan generally match the description of T. cf. turkestanica from Uttarakhand state of India (Pešić et al. 2019a). As mentioned by the latter authors, the assignment of populations from Uttarakhand to T. turkestanica, a species originally described from Tajikistan (Sokolow 1926) is still provisional, based ''mainly on the approved non-identity with alternative species'' (Pesic et al. 2007), and should be ''tested with the application of molecular techniques in case data are available from the type locality.'' (Pesic et al. 2019a: 1871). Therefore, the same conclusion applies also to populations from Bhutan.
Specimens from Thailand, provisionally assigned by Pesic and Smit (2009) to T. turkestanica, were described recently as a separate species, i.e. T. rangareddyi Pešić & Smit, 2019, differing from the former species by an elongated gnathosomal rostrum and a shorter ventral seta on P-2 (Pešić et al. 2019a).
Distribution — India (Uttarakhand; Pešić et al. 2019a); Bhutan (this study).
Torrenticola longiseta sp. nov.
ZOOBANK: 28E8E90D-E1EC-4473-9A2B-188D5C93BE21 ![]()
Figures 10, 11, 14A–C
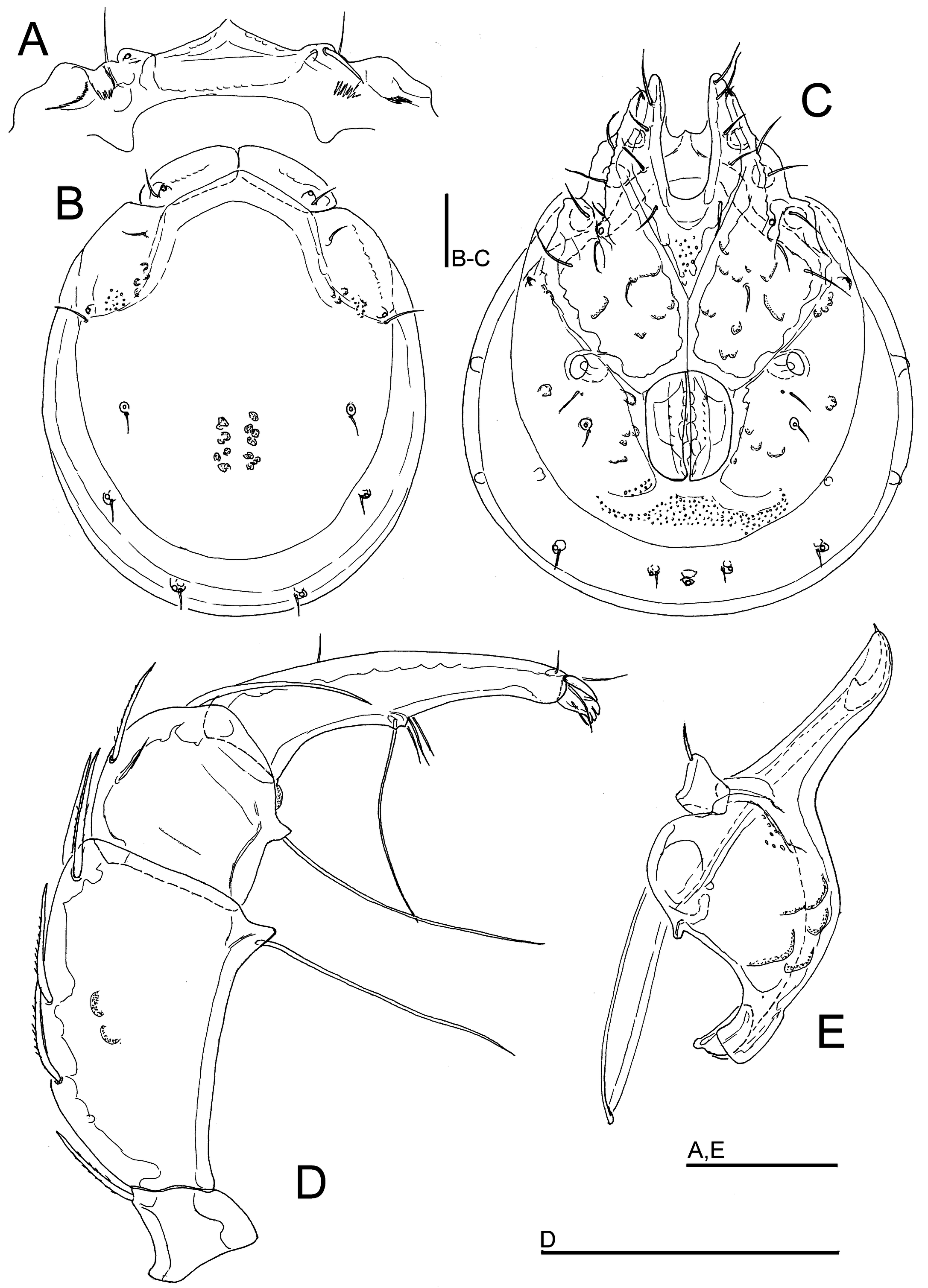

Material examined — Holotype ♂, dissected and slide mounted, Bhutan, MG5 Dakpay Chhu, 27.14621°N, 90.69220°E, 539 m asl, 27.x.2021. Paratypes: 4♂, 4♀, same data as holotype, 1♀ dissected and slide mounted; 2♀, same data as holotype, 1.v.2021, 1♀ dissected and slide mounted; 1♂ (juvenile), 1♀ (juvenile), MG1 Maidagang Chhu, 27.12761°N, 90.71560°E, 554 m asl, 20.x.2021; 1♂, MG4 Takabi Chhu, 27.14782°N, 90.68833°E, 543 m asl, 26.x.2021.
Diagnosis — Shoulder platelets fused with dorsal plate; dorsal shield with colour pattern as illustrated in Figures 14A, B; Cxgl–4 subapical; medial suture line of Cx-II+III relatively short; P-2 ventrodistal extension relatively short, on a large base, triangular, P-2 ventral seta long, clearly exceeding width of segment.
Description — General features – Idiosoma oval; shoulder platelets fused to dorsal plate, but suture line visible; dorsal shield with a colour pattern as illustrated in Figures 14A, B; area of primary sclerotization of the dorsal plate with two dorsoglandularia; gnathosomal bay U-shaped, proximally rounded; Cxgl-4 subapical; suture lines of Cx-IV distinctly extending posteriorly beyond posterior margin of genital field; excretory pore and Vgl-2 away from the line of primary sclerotization; gnathosoma with a strongly curved ventral margin, rostrum elongated (Figure 10E); P-2 ventral margin straight or slightly concave, ventral seta long, exceeding the segment width, P-2 and P-3 ventrodistal protrusions relatively short, P-2 ventrodistal extension slightly curved distally, P-3 ventral seta long, P-4 distally tapering, with a ventral tubercle bearing one long and three shorter setae (Figures 10D, 11B). Male – Medial suture line of Cx-II+III relatively short; genital field subrectangular; ejaculatory complex conventional in shape (with well-developed anterior keel and proximal arms; Figure 14C).
Measurements. Male (holotype) – Idiosoma (ventral view: Figure 10C) L 734, W 606; dorsal shield (Figures 10B, 14A) L 638, W 516, L/W ratio 1.24; dorsal plate L 603; frontal plate L 145–147, W 53–55, L/W ratio 2.7–2.8. Gnathosomal bay L 167, Cx-I total L 297, Cx-I mL 128, Cx-II+III mL 94; ratio Cx-I L/Cx-II+III mL 3.16; Cx-I mL/Cx-II+III mL 1.36. Genital field L/W 142/114, ratio 1.24; distance genital field-excretory pore 134, genital field-caudal idiosoma margin 184. Ejaculatory complex L 178. Gnathosoma vL 294, chelicera L 368; palp total L 356, dL/H, dL/H ratio: P-1, 36/36, 0.99; P-2, 119/64, 1.88; P-3, 61/59, 1.04; P-4, 123/34, 3.6; P-5, 17/11, 1.6; L ratio P-2/P-4 0.93. dL of I-L-2–6: 66, 81, 98, 103, 103; I-L-6 H 30; dL/H I-L-6 ratio 3.4.
Female (paratype from Dakpay Chhu, n = 1) – Idiosoma (ventral view: Figure 11A) L 887, W 700; dorsal shield (Figure 14B) L 731, W 613, L/W ratio 1.19; dorsal plate L 688; frontal plate L 167–170, W 59, L/W ratio 2.8–2.9. Gnathosomal bay L 194, Cx-I total L 334, Cx-I mL 138, Cx-II+III mL 44; ratio Cx-I L/Cx-II+III mL 7.6; Cx-I mL/Cx-II+III mL 3.15. Genital field L/W 169/150, ratio 1.13; distance genital field-excretory pore 222, genital field-caudal idiosoma margin 322. Gnathosoma vL 338, chelicera L 438; palp total L 421, dL/H, dL/H ratio: P-1, 41/44, 0.95; P-2, 147/79, 1.87; P-3, 70/69, 1.02; P-4, 141/39, 3.6; P-5, 22/13, 1.75; L ratio P-2/P-4 1.05. dL of I-L-4–6: 106, 111, 102; I-L-6 H 30; dL/H I-L-6 ratio 3.4.
Etymology — The species is named for its long setae of the palp.
Discussion — The new species is most similar to Torrenticola minuta (Lundblad, 1941), a species originally described from Java (Lundblad 1941), and later on reported from the Malay Peninsula (Wiles 1997). Both species have the dorsal shield with the shoulder platelets partially fused with the dorsal plate, a Cxgl-4 situated subapically and a relatively short median suture line of Cx-II–III in the male. The new species from Bhutan can be separated from T. minuta by P-2 with a shorter ventral projection and a longer ventral seta (T. minuta has a shorter ventral seta, not exceeding width of the segment, see Lundblad 1956, Figure 4F, and Wiles 1997, Figure 13C) and by a different colour pattern of the dorsal shield (in T. minuta typically with a broad posterior band of blue/violet pigment, Wiles 1997).
Distribution — Bhutan.
Torrenticola brachypalpis sp. nov.
ZOOBANK: 70A3C10B-4F5E-44A5-A2E1-3CACCAF0D0E8 ![]()
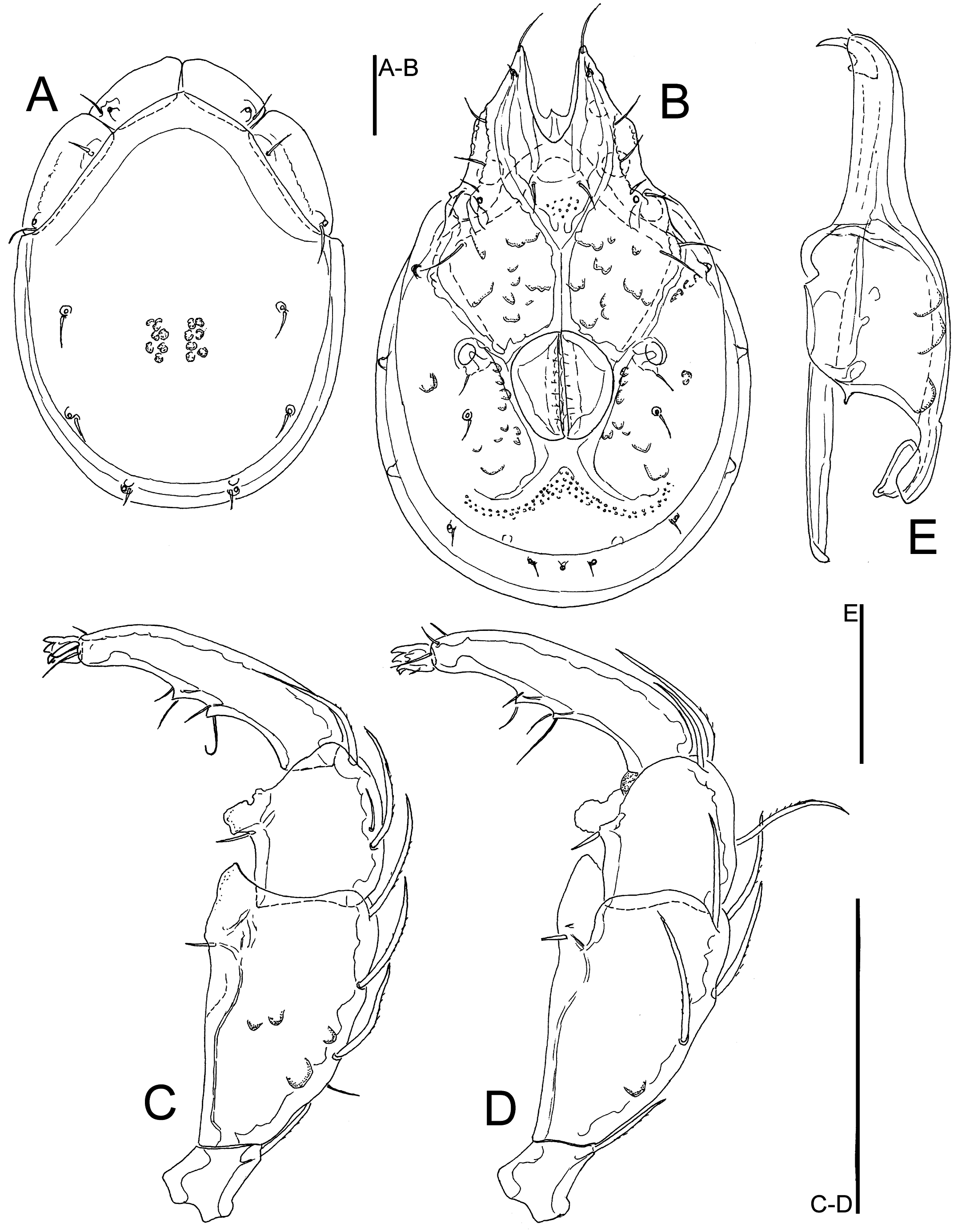
Figures 12, 13, 14D-I
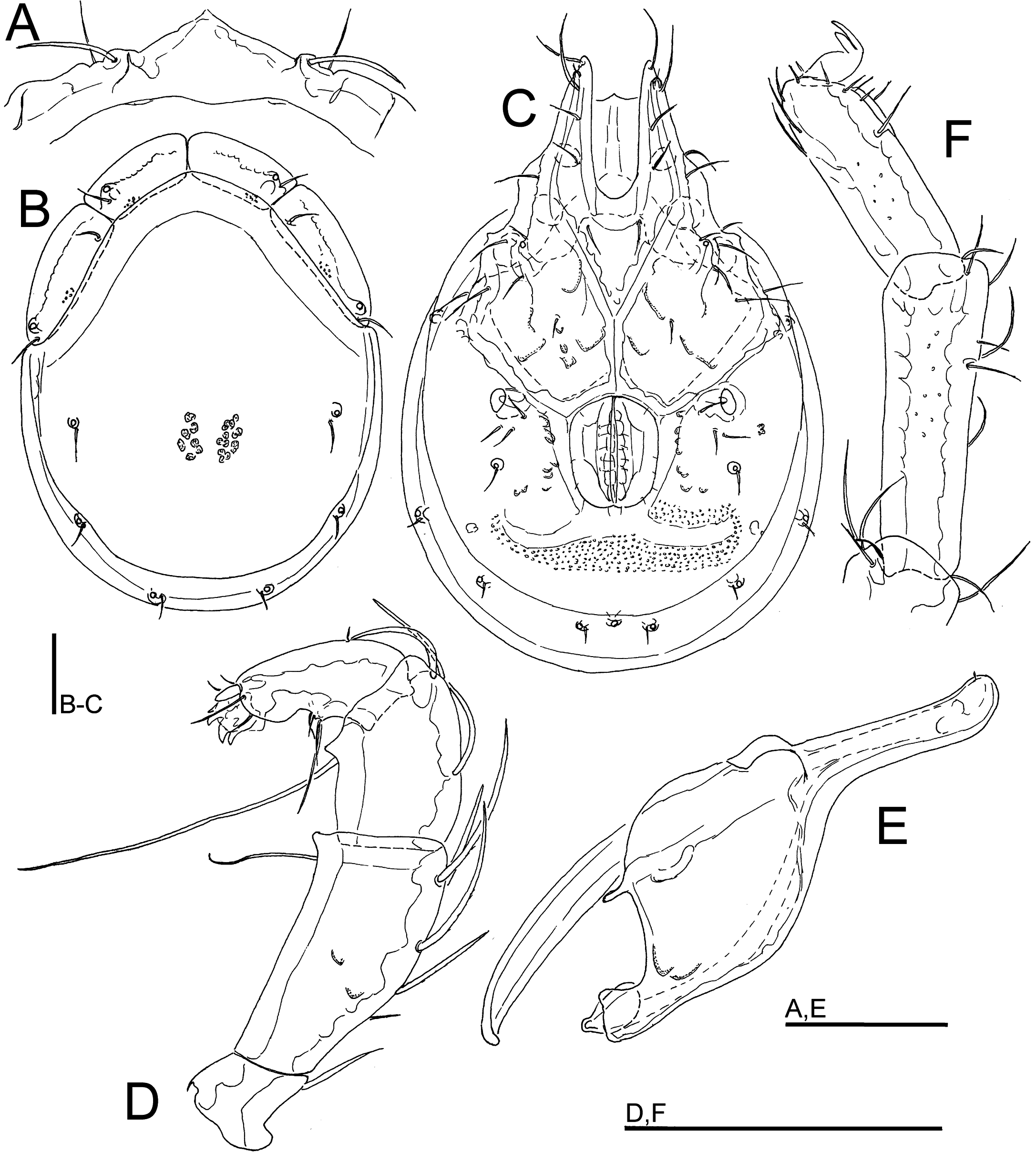

Material examined — Holotype ♂, dissected and slide mounted, Bhutan, MG3 Bipgang Chhu, 27.15729°N, 90.66721°E, 586 m asl, 25.x.2021. Paratypes: 1 ♂ (juvenile), MG3 Bipgang Chhu, 27.15729°N, 90.66721, 586 m asl, 28.vi.2021; 1 ♀, Bhutan, MG4 Takabi Chhu, 27.14782°N, 90.68833°E, 543 m asl, 30.vi.2021, dissected and slide mounted; 1 ♂, 1 ♀, MG4 Takabi Chhu, 27.14782°N, 90.68833°E, 543 m asl, 26.x.2021, dissected and slide mounted; 1 ♂ (damaged, dorsal shield lacking), MG5 Dakpay Chhu, 27.14621°N, 90.69220°E, 539 m asl, 1.v.2021.
Diagnosis — Gnathosomal rostrum long and very slender; P-2 ventral seta relatively long (reaching almost width of the segment) and slightly below the ventrodistal projection, P-3 with a very long seta located at the base of the projection, P-4 relatively short and stout, L ratio P-2/P-4 1.5–1.6.
Description — General features – Idiosoma oval-elongated (dorsal shield L/W ratio 1.3-1.4); dorsal shield without a colour pattern (as photographed in Figures 14 D-E); area of primary sclerotization of the dorsal plate with two dorsoglandularia; gnathosomal bay U-shaped, proximally pointed; Cxgl-4 subapical; suture lines of Cx-IV distinctly extending posteriorly beyond posterior margin of genital field, laterally curved; excretory pore and Vgl-2 located close to the margin of primary sclerotization; gnathosoma with a curved ventral margin, rostrum elongated and slender (Figure 12E); P-2 ventral margin straight, ventral seta slightly below the ventrodistal projection, P-2 and P-3 ventrodistal protrusions small and bluntly pointed, P-3 with a very long seta located at the base of the projection, P-4 relatively short and stout, P-4 with a ventral tubercle bearing one longer and three shorter setae (Figures 12D, 13C). Male – Medial suture line of Cx-II+III relatively short; genital field subrectangular; ejaculatory complex conventional in shape (with well-developed anterior keel and proximal arms, Figure 14F). Female – Genital field wider than in male and slightly tapering posteriorly.
Measurements. Male (holotype) – Idiosoma (ventral view: Figure 12C) L 752, W 531; dorsal shield (Figure 12B) L 591, W 445, L/W ratio 1.33; dorsal plate L 550; shoulder plate L 188–195, W 68–69, L/W ratio 2.77–2.84; frontal plate L 150, W 63, L/W ratio 2.4; shoulder/frontal plate L 1.25–1.3. Gnathosomal bay L 175, Cx-I total L 322, Cx-I mL 146, Cx-II+III mL 94; ratio Cx-I L/Cx-II+III mL 3.43; Cx-I mL/Cx-II+III mL 1.55. Genital field L/W 138/109, ratio 1.26; distance genital field-excretory pore 142, genital field-caudal idiosoma margin 198. Ejaculatory complex L 168. Gnathosoma vL 334, chelicera L 394; palp total L 244, dL/H, dL/H ratio: P-1, 28/27, 1.03; P-2, 86/41, 2.1; P-3, 58/39, 1.48; P-4, 55/27, 2.1; P-5, 17/13, 1.3; L ratio P-2/P-4 1.57. dL of I-L-2–6: 75, 75, 102, 102, 87; I-L-6 H 28; dL/H I-L-6 ratio 3.1.
Female (paratype from Takabi Chhu, n = 1) – Idiosoma (ventral view: Figure 13B) L 884, W 575; dorsal shield (Figure 13A) L 659, W 478, L/W ratio 1.38; dorsal plate L 613; shoulder plate L 203, W 72, L/W ratio 2.8; frontal plate L 156–159, W 64–66, L/W ratio 2.4–2.5; shoulder/frontal plate L 1.27–1.3. Gnathosomal bay L 197, Cx-I total L 359, Cx-I mL 163, Cx-II+III mL 69; ratio Cx-I L/Cx-II+III mL 5.2; Cx-I mL/Cx-II+III mL 2.36. Genital field L/W 160/141, ratio 1.14; distance genital field-excretory pore 188, genital field-caudal idiosoma margin 269. Gnathosoma vL 363, chelicera L 438; palp total L 278, dL/H, dL/H ratio: P-1, 30/31, 0.97; P-2, 98/50, 1.97; P-3, 67/45, 1.5; P-4, 66/30, 2.21; P-5, 17/14, 1.23; L ratio P-2/P-4 1.5. dL of I-L-4–6: 108, 108, 91; I-L-6 H 27; dL/H I-L-6 ratio 3.4.
Etymology — Named for the relatively short palp.
Discussion — In regard to the similar shape of gnathosoma and a P-4 shortened and stout, the new species resembles Torrenticola yunnanensis Gu, Jin & Guo, 2020, a species described from a single female from Yunnan Province (Mengla, China). In regard to the shape of the genital field (subrectangular as illustrated in figure 3B in Gu et al. 2020b) it is likely that the illustrated female of T. yunnanensis represent a male specimen. Torrenticola yunnanensis differs in having a shorter ventral setae on P-2 and P-3, and the ventral seta on P-2 is located proximally to the ventral extension in T. brachypalpis sp. nov.
Distribution — Bhutan (this study).
Torrenticola trongsaensis sp. nov.
ZOOBANK: 67C92662-2D1B-4678-8E87-E6C0AFFA9F29 ![]()
Figures 15, 16, 19H–J


Material examined — Holotype ♂, dissected and slide mounted, Bhutan, MG15 Khabab Chhu, 27.48492°N, 90.33490°E, 2500 m asl, 14.v.2021. Paratypes: 1♂, 4♀, same data as holotype; 1♀, MG14 Chuserbu stream, 27.50246°N, 90.31782°E, 2666 m asl, 13.v.2021, dissected and slide mounted; 1♀, MG12 Nikka Chhu, 27.52601°N, 90.29947°E, 2609 m asl, 10.v.2021.
Diagnosis — Idiosoma large in size, < 1000 µm in both sexes; Cx-I relatively short, anteriorly broad; gnathosomal rostrum short, less than width of gnathosoma; in both sexes suture lines of Cx-IV extending posteriorly beyond posterior margin of genital field; ejaculatory complex with well-developed anterior keel and proximal arms.
Description — General features – Idiosoma roundish; dorsal shield without a colour pattern (as photographed in Figures 19H–I); area of primary sclerotization of the dorsal plate with four dorsoglandularia; frontal platelets broad, frontal platelets broad, relatively short; Cx-I relatively short, anteriorly broad (Figure 15C); gnathosomal bay U-shaped, proximally rounded; Cxgl-4 subapical; medial suture line of Cx-II+III relatively short; suture lines of Cx-IV in both sexes distinctly extending posteriorly beyond posterior margin of genital field, laterally curved; excretory pore and Vgl-2 away from the line of primary sclerotization, excretory pore on the level of Vgl-2; gnathosomal rostrum short, less than depth of gnathosoma (Figure 16E); P-2 ventral margin nearly straight, P-2 and P-3 ventrodistal protrusions bluntly pointed, P-4 with a ventral tubercle bearing one long and three shorter setae (Figures 15D, E). Male – Genital field elongated, subrectangular; ejaculatory complex conventional in shape (Figure 19J). Female – Genital field large and pentagonal in shape.
Measurements. Male (holotype) – Idiosoma (ventral view: Figure 15C) L 1065, W 853; dorsal shield (Figure 15B) L 909, W 738, L/W ratio 1.23; dorsal plate L 825; shoulder plate L 288–294, W 125–131, L/W ratio 2.2–2.3; frontal plate L 194, W 113–116, L/W ratio 1.68–1.72; shoulder/frontal plate L 1.48–1.52. Gnathosomal bay L 206, Cx-I total L 397, Cx-I mL 191, Cx-II+III mL 96; ratio Cx-I L/Cx-II+III mL 4.1; Cx-I mL/Cx-II+III mL 2.0. Genital field L/W 234/162, ratio 1.45; distance genital field-excretory pore 200, genital field-caudal idiosoma margin 313. Gnathosoma vL 334, chelicera L 384; palp total L 360, dL/H, dL/H ratio: P-1, 38/39, 0.96; P-2, 116/64, 1.81; P-3, 69/56, 1.22; P-4, 114/34, 3.32; P-5, 23/14, 1.7; L ratio P-2/P-4 1.01. dL of I-L-2–6: 94, 109, 134, 153, 144; I-L-6 H 48; dL/H I-L-6 ratio 3.0.
Female (paratype from Chuserbu stream, n = 1) – Idiosoma (ventral view: Figure 16B) L 1108, W 906; dorsal shield (Figure 16A) L 947, W 768, L/W ratio 1.23; dorsal plate L 869; shoulder plate L 294–300, W 134–138, L/W ratio 2.17–2.2; frontal plate L 194, W 119, L/W ratio 1.63; shoulder/frontal plate L 1.52–1.55. Gnathosomal bay L 206, Cx-I total L 397, Cx-I mL 191, Cx-II+III mL 40; ratio Cx-I L/Cx-II+III mL 9.9; Cx-I mL/Cx-II+III mL 4.8. Genital field L/W 247/209, ratio 1.18; distance genital field-excretory pore 250, genital field-caudal idiosoma margin 397. Gnathosoma vL 353, chelicera L 366; palp total L 366, dL/H, dL/H ratio: P-1, 38/40, 0.96; P-2, 118/61, 1.94; P-3, 72/58, 1.25; P-4, 117/36, 3.37; P-5, 21/14, 1.5; L ratio P-2/P-4 1.0.
Etymology — The species is named after the district Trongsa from where the new species was collected.
Discussion — In regard to the presence of an anteriorly broad and short Cx-I, an elongated genital field in the male (L/W ratio about 1.4), and a deep gnathosoma, with a short rostrum, the new species resembles T. retractipora (Lundblad, 1941) and T. siamis Pešić & Smit, 2009. The latter two species can be separated from T. trongsaensis sp. nov. by Cx-IV not extended posterior of the genital field, and a comparatively shorter and stouter gnathosomal rostrum. Torrenticola retractipora, known from a single male specimen collected in a stream at Kambaiti pass in Myanmar differs by the characteristic shape of the ejaculatory complex (similar to T. bhutana as illustrated in Figure 11G: proximal and distal arms short, proximal chamber large, proximal horns reduced, see Lundblad 1969, Figure 27f). Torrenticola siamis, a species originally described from Thailand, has a more elongated dorsal shield (L/W ratio 1.4 in ♂ holotype, data taken from Pešić and Smit 2009).
Distribution — Bhutan (this study).
Torrenticola dbgurungi sp. nov.
ZOOBANK: 6D1A6787-A6AC-4C70-9A01-F18E30530573 ![]()
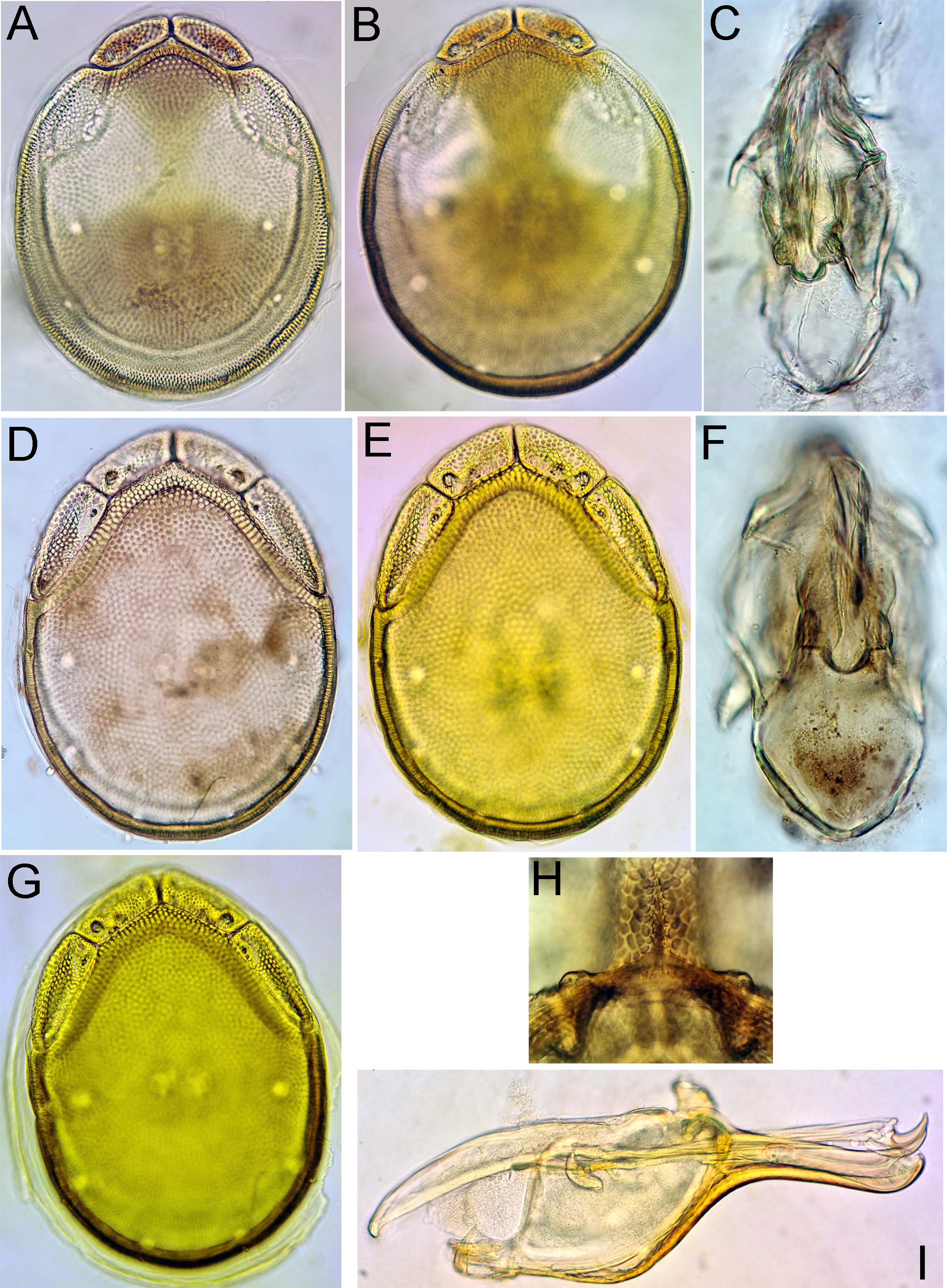
Figures 17, 18, 19A–G


Material examined — Holotype ♂ dissected and slide mounted, Bhutan, MG8 Dangdung Chhu, 27.33461°N, 90.59562°E, 1039 m asl, 30.x.2021. Paratypes: 2♂, 1♀, same data as the holotype, 1♂, 1♀ dissected and slide mounted; 1♂, MG8 Dangdung Chhu, 27.33461°N, 90.59562°E, 1039 m asl, 5.v.2021, dissected and slide mounted; 4 ♂ (2 of them juveniles), 1♀, MG9 Kartigang Chhu, 27.27896°N, 90.63088°E, 1456 m asl, 7.v.2021, 1♂, dissected and slide mounted; MG9 Kartigang Chhu, 27.27896°N, 90.63088°E, 1456 m asl, 1.xi.2021, 1♀, dissected and slide mounted.
Diagnosis — Medial suture line of Cx-II+III relatively short; area of primary sclerotization of the dorsal plate with four dorsoglandularia; Cx-IV extended posterior to the genital flaps in both sexes; ejaculatory complex with poorly developed anterior keel, proximal chamber relatively large.
Description — General features – Idiosoma oval; dorsal shield without a colour pattern (as photographed in Figures 19A, B, D); area of primary sclerotization of the dorsal plate with four dorsoglandularia; gnathosomal bay U-shaped, proximally rounded; Cxgl-4 subapical; medial suture line of Cx-II+III relatively short; suture lines of Cx-IV in both sexes distinctly extending posteriorly beyond posterior margin of genital field, laterally curved; excretory pore and Vgl-2 on the the line of primary sclerotization, excretory pore on the level of Vgl-2; gnathosomal ventral margin curved, rostrum almost equal to or less than depth of gnathosoma; P-2 longer than P-4; P-2 ventral margin straight, P-2 and P-3 ventrodistal protrusions bluntly pointed, P-4 with a ventral tubercle bearing one long and three shorter setae (Figure 17C). I-L-6 stout (Figure 17E). Male – Medial suture line of Cx-II+III relatively short; genital field with obtuse, but distinct anterior angles, posteriorly parabola-shaped; ejaculatory complex with poorly developed anterior keel, proximal chamber relatively large (Figures 19C, G). Female – Genital field wider than in male and slightly tapering posteriorly.
Measurements. Male (holotype) – Idiosoma (ventral view: Figure 17B) L 794, W 563; dorsal shield (Figure 17A) L 607, W 481, L/W ratio 1.26; dorsal plate L 563; shoulder plate L 203–206, W 77–81, L/W ratio 2.5–2.7; frontal plate L 134–138, W 69, L/W ratio 1.95–2.0; shoulder/frontal plate L 1.5. Gnathosomal bay L 147, Cx-I total L 328, Cx-I mL 178, Cx-II+III mL 78; ratio Cx-I L/Cx-II+III mL 4.2; Cx-I mL/Cx-II+III mL 2.3. Genital field L/W 163/128, ratio 1.28; distance genital field-excretory pore 150, genital field-caudal idiosoma margin 203. Ejaculatory complex L 244. Gnathosoma vL 281, chelicera L 322; palp total L 261, dL/H, dL/H ratio: P-1, 28/31, 0.9; P-2, 83/49, 1.7; P-3, 56/44, 1.3; P-4, 75/31, 2.4; P-5, 19/13, 1.5; L ratio P-2/P-4 1.1. dL of I-L-2–6: 79, 80, 97, 108, 111; I-L-6 H 52; dL/H I-L-6 ratio 2.15.
Female (paratype from Dangdung Chhu, n = 1) – Idiosoma (ventral view: Figure 18B) L 893, W 650; dorsal shield (Figure 18A) L 728, W 563, L/W ratio 1.29; dorsal plate L 675; shoulder plate L 228, W 84, L/W ratio 2.7; frontal plate L 159–163, W 75, L/W ratio 2.1–2.2; shoulder/frontal plate L 1.4–1.43. Gnathosomal bay L 175, Cx-I total L 359, Cx-I mL 181, Cx-II+III mL 45; ratio Cx-I L/Cx-II+III mL 8.0; Cx-I mL/Cx-II+III mL 4.0. Genital field L/W 191/163, ratio 1.17; distance genital field-excretory pore 206, genital field-caudal idiosoma margin 272. Gnathosoma vL 322; palp total L 292, dL/H, dL/H ratio: P-1, 31/33, 0.95; P-2, 95/53, 1.79; P-3, 66/50, 1.3; P-4, 83/34, 2.4; P-5, 17/13, 1.3; L ratio P-2/P-4 1.15. dL of I-L-4–6: 111, 112, 119; I-L-6 H 47; dL/H I-L-6 ratio 2.5.
Etymology — The species is named after Prof. Dhan Bdr Gurung (College of Natural Resources, Royal University of Bhutan) in appreciation of his work on the freshwater fishes of Bhutan.
Discussion — In regard to a short median suture line of Cx-II+III and Cx-IV extended posterior to the genital flaps in both sexes, the new species resembles Torrenticola bhutana Pešić & Smit, 2007. The latter species can easily be separated by the elongated gnathosomal rostrum and the characteristic shape of the ejaculatory complex (see Figure 1G).
Torrenticola dentipalpis Jin, 1997 a species described from a stream in Mengla, Yunnan province, China (Jin 1997) is similar in shape of the gnathosoma with a rostrum shorter than width of gnathosoma, and Cx-IV extended posterior to the genital flaps, but differs in I-L-6 not extended (see Figure 80H in Jin 1997) as in the new species from Bhutan, and the dorsal plate bears two dorsoglandularia (Figure 80E in Jin 1997) instead of four dorsoglandularia as in the new species from Bhutan.
Remarks — In one female specimen from Dangdung Chhu the palp has two setae on ventral projection of P-2 (Figure 18D), a character we consider as an individual aberration.
Distribution — Bhutan (this study).
Torrenticola himalayica sp. nov.
ZOOBANK: EE2A1CBD-893C-465B-8296-327EA645F191 ![]()
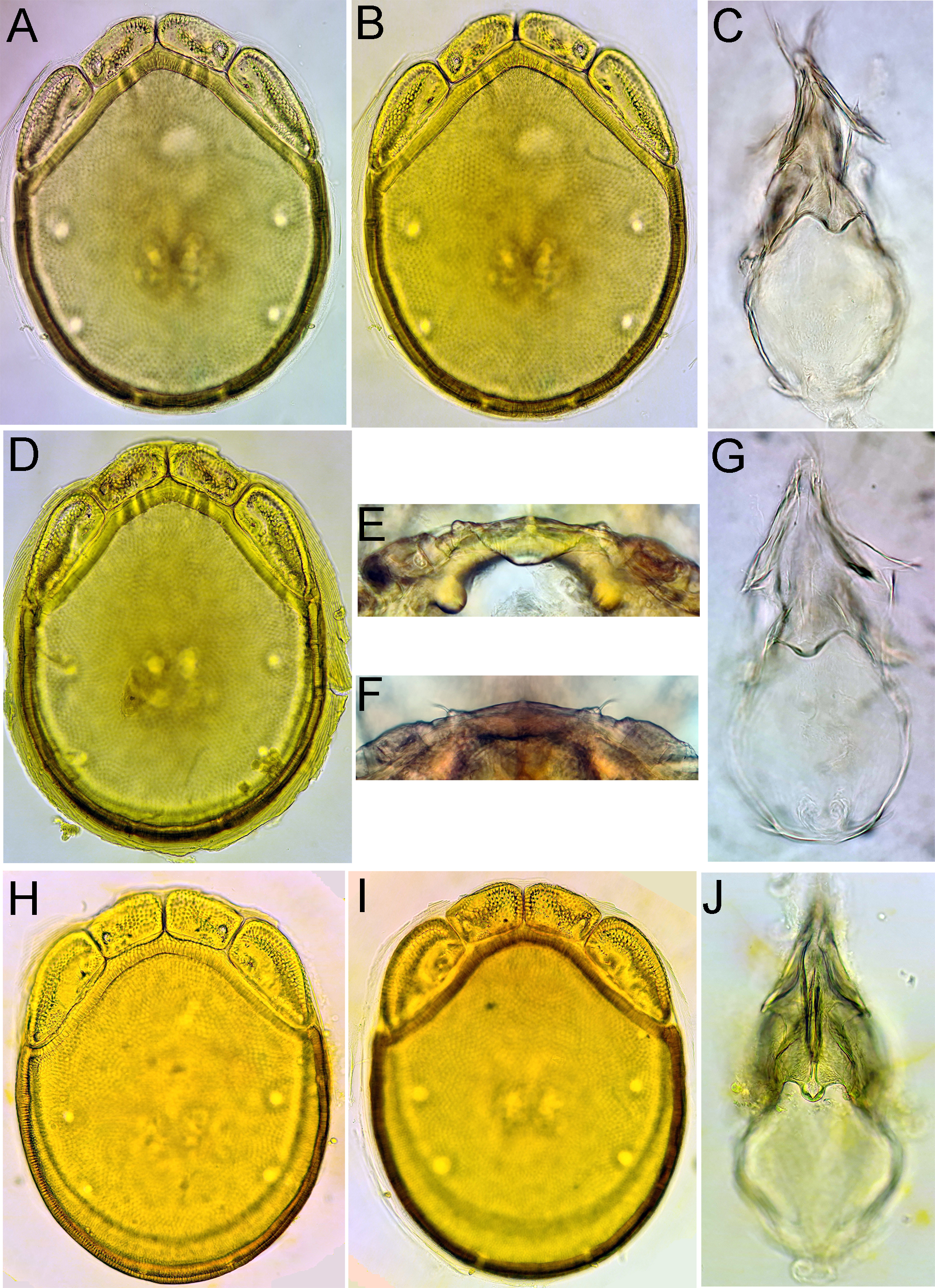
Figures 20, 21, 24C, D, G

Material examined — Holotype ♂, dissected and slide mounted, Bhutan, MG2 Berti Chhu, 27.16264°N, 90.66003°E, 590 m asl, 26.iv.2021. Paratype: 1♀, MG5 Dakpay Chhu, 27.14621°N, 90.69220°E, 539 m asl, 1.v.2021, dissected and slide mounted.
Diagnosis — Dorsal shield without a colour pattern; P-2 ventrally with a flat, blade-like hyaline anterior flange, and a short denticle-like seta, laterally at base of projection; P-3 with a short seta laterally at base of projection.
Description — General features – Idiosoma roundish; dorsal shield without a colour pattern (as photographed in Figures 24C-D); area of primary sclerotization of the dorsal plate with four dorsoglandularia; gnathosomal bay V-shaped; Cxgl-4 subapical; medial suture line of Cx-II+III moderately long; posterior suture lines of Cx-IV extending posteriorly beyond posterior margin of genital field, laterally curved; excretory and Vgl-2 close to the line of primary sclerotization pore; gnathosomal rostrum long and slender (Figure 21E); P-2 shorter than P-4, P-2 with a laterally compressed, longish (< 30% of ventral margin), anteriorly directed ventrodistal extension, and a short seta, laterally at base of projection; P-3 with a short, subrectangular, apically serrated ventrodistal projection, and a short seta laterally at base of projection; P-4 slender with a well-developed ventral tubercle ending in two tips separated by a concavity, bearing shorter setae (Figures 20D, 21C, D). Male – Genital field with obtuse, but distinct anterior angles, posteriorly parabola-shaped; male ejaculatory complex conventional in shape, with well-developed anterior keel and proximal arms (Figure 24G).
Measurements. Male (holotype) – Idiosoma (ventral view: Figure 20C) L 650, W 478; dorsal shield (Figure 20B) L 529, W 416, L/W ratio 1.27; dorsal plate L 488; shoulder plate L 172, W 63, L/W ratio 2.8; frontal plate L 125, W 56, L/W ratio 2.2; shoulder/frontal plate L 1.38. Gnathosomal bay L 106, Cx-I total L 234, Cx-I mL 125, Cx-II+III mL 122; ratio Cx-I L/Cx-II+III mL 1.9; Cx-I mL/Cx-II+III mL 1.03. Genital field L/W 112/94, ratio 1.19; distance genital field-excretory pore 147, genital field-caudal idiosoma margin 181. Palp total L 249, dL/H, dL/H ratio: P-1, 27/22, 1.2; P-2, 78/41, 1.89; P-3, 47/34, 1.36; P-4, 83/22, 3.8; P-5, 14/9, 1.5; L ratio P-2/P-4 0.94. dL of I-L-4–6: 77, 90, 84; I-L-6 H 20; dL/H I-L-6 ratio 4.1.
Female (paratype from Dakpay Chhu, n = 1) – Idiosoma (ventral view: Figure 21B) L 700, W 472; dorsal shield (Figure 21A) L 548, W 409, L/W ratio 1.34; dorsal plate L 500; shoulder plate L 169–172, W 66–67, L/W ratio 2.56–2.58; frontal plate L 125–128, W 58–61, L/W ratio 2.1–2.16; shoulder/frontal plate L 1.32–1.38. Gnathosomal bay L 119, Cx-I total L 255, Cx-I mL 136, Cx-II+III mL 91; ratio Cx-I L/Cx-II+III mL 2.8; Cx-I mL/Cx-II+III mL 1.5. Genital field L/W 134/122, ratio 1.1; distance genital field-excretory pore 159, genital field-caudal idiosoma margin 219. Gnathosoma vL 300, chelicera L 334; palp total L 256, dL/H, dL/H ratio: P-1, 27/23, 1.14; P-2, 81/48, 1.7; P-3, 45/41, 1.1; P-4, 89/22, 4.07; P-5, 14/9, 1.5 L ratio P-2/P-4 0.91.
Etymology — The species is named after the Himalayan mountain range from where the new species was collected.
Discussion — The new species is most similar to Torrenticola lamellifer (Lundblad, 1941). Both species have a long tapering blade-like flange extending to the middle of P-2. The new species from Bhutan differs from T. lamellifer in shape of the palp (P-2 with a small denticle-like ventral seta, a short ventral seta laterally at base of projection P-3, P-4 ventral setae shorter; compare Figures 6A and 21C, D) and by the lack of characteristic colour pattern (compare Figures 1E and 24C, D).
Distribution — Bhutan (this study).
Torrenticola ancylopalpis sp. nov.
ZOOBANK: BB8C2F60-A4B1-4E5D-9FDF-3AD4FA833E83 ![]()
Figures 22, 24E

Material examined — Holotype ♀, dissected and slide mounted, Bhutan, MG9 Kartigang Chhu, 27.27896°N, 90.63088°E, 1456 m asl, 7.v.2021.
Diagnosis (Male unknown) — Idiosoma elongated (dorsal shield L/W ratio 1.5); P-2 with a laterally compressed, blade-like anterior flange, ventral extensions on P-2 and P-3 apically irregularly serrated, P-4 curved, with a well-developed ventral tubercle, located in posterior half of the segment.
Description — Female – Idiosoma elongated; dorsal shield without a colour pattern (as photographed in Figure 24E); area of primary sclerotization of the dorsal plate with two dorsoglandularia; gnathosomal bay U-shaped, proximally pointed; Cxgl-4 subapical; posterior suture lines of Cx-IV strongly extending posteriorly beyond posterior margin of genital field; excretory and Vgl-2 on the same line, close to the line of primary sclerotization pore; gnathosomal rostrum long and slender (Figure 22F); P-2 with a laterally compressed, longish (< 30% of ventral margin), anteriorly directed and apically irregularly serrated ventrodistal extension, a short seta, laterally exceeding the tip of projection, at base of projection; P-3 with longish, apically irregularly serrated ventrodistal projection, and a short seta laterally at base of projection, exceeding the tip of projection; P-4 curved, with a well-developed ventral tubercle, located in posterior half of the segment, ending in two tips separated by a concavity, bearing three shorter setae and one slightly longer seta (Figures 22D, E).
Measurements – Idiosoma (ventral view: Figure 22C) L 681, W 450; dorsal shield (Figure 22B) L 547, W 371, L/W ratio 1.47; dorsal plate L 505; shoulder plate L 181–188, W 63–66, L/W ratio 2.8–3.0; frontal plate 122, W 48, L/W ratio 2.6; shoulder/frontal plate L 1.49–1.54. Gnathosomal bay L 116, Cx-I total L 244, Cx-I mL 128, Cx-II+III mL 84; ratio Cx-I L/Cx-II+III mL 2.89; Cx-I mL/Cx-II+III mL 1.5. Genital field L/W 138/116, ratio 1.19; distance genital field-excretory pore 144, genital field-caudal idiosoma margin 197. Gnathosoma vL 325, chelicera L 338; palp total L 289, dL/H, dL/H ratio: P-1, 33/24, 1.36; P-2, 88/44, 2.0; P-3, 63/38, 1.6; P-4, 89/27, 3.3; P-5, 16/13, 1.25; L ratio P-2/P-4 0.98. dL of I-L: 47, 64, 67, 78, 89, 83; I-L-6 H 29; dL/H I-L-6 ratio 2.8.
Male – Unknown.
Etymology — Named for its bent palp.
Discussion — In view of the long tapering blade-like flange extending to the middle of P-2, and a relatively short ventral seta on P-2 and P-3 the new species resembles T. himalayica sp. nov. (see above). From the latter species, the female of the new species differs in a more elongated dorsal shield, the ventrodistal extensions of P-2 and P-3 are apically irregularly serrated, and P-4 is curved with a ventral tubercle located in the posterior half of the segment (compare Figures 21C, D and 22D, E).
Distribution — Bhutan (this study).
Torrenticola megapalpis sp. nov.
ZOOBANK: 69C5D39A-0966-483C-93D0-E7F521F065BF ![]()
Figures 23, 24H–J

Material examined — Holotype ♀, dissected and slide mounted, Bhutan, MG9 Kartigang Chhu, 27.27896°N, 90.63088°E, 1456 m asl, 7.v.2021.
Diagnosis (Male unknown) — Gnathosomal bay shallow; gnathosoma long and narrow, rostrum long; palpal segments long and slender, P-2 ventrally with a flat, blade-like hyaline anterior flange, and a short denticle-like seta, laterally at base of projection.
Description — Female – Idiosoma oval; dorsal shield without a colour pattern (as photographed in Figure 24H); gnathosomal bay shallow; Cxgl-4 subapical; suture lines of Cx-IV distinctly extending posteriorly beyond posterior margin of genital field; excretory and Vgl-2 on the line of primary sclerotization pore; infracapitulum long and narrow, rostrum long (Figure 23F); P-2 elongated, with a laterally compressed, longish (< 30% of ventral margin), anteriorly directed ventrodistal extension, and a small, denticle-like seta, at base of projection; P-3 with longish, apically serrate, ventrodistal projection, and a small, denticle-like seta laterally at base of projection; P-4 curved, with a well-developed ventral tubercle, located in posterior half of the segment, ending in two tips separated by a concavity, bearing shorter setae (Figures 23D, E).
Measurements – Idiosoma (ventral view: Figure 23C) L 794, W 494; dorsal shield (Figure 23B) L 572, W 444, L/W ratio 1.29; dorsal plate L 513; shoulder plate L 213–216, W 81–84, L/W ratio 2.6; frontal plate L 159, W 69, L/W ratio 2.3; shoulder/frontal plate L 1.33–1.35. Gnathosomal bay L 72, Cx-I total L 306, Cx-I mL 233, Cx-II+III mL 101; ratio Cx-I L/Cx-II+III mL 3.03; Cx-I mL/Cx-II+III mL 2.3. Genital field L/W 153/131, ratio 1.17; distance genital field-excretory pore 150, genital field-caudal idiosoma margin 219. Gnathosoma vL 353, chelicera L 388; palp total 266, dL/H, dL/H ratio: P-1, 34/19, 1.8; P-2, 81/31, 2.6; P-3, 47/25, 1.9; P-4, 89/17, 5.2; P-5, 15/9, 1.6; L ratio P-2/P-4 0.92. dL of I-L: 42, 73, 63, 86, 98, 105; I-L-6 H 26; dL/H I-L-6 ratio 4.1.
Male – Unknown.
Etymology — The species is named for its long palp.
Discussion — Among the species with a flanged palp and a 4 + 1 arrangement of the dorsal plates, the new species is unique in the combination of long and slender palpal segments, a shallow gnathosomal bay and a long and narrow gnathosoma.
Distribution — Bhutan (this study).
Genus Neoatractides Lundblad, 1941
Subgenus Allotorrenticola Cook, 1967
Neoatractides (Allotorrenticola) tashiwangmoi sp. nov.
ZOOBANK: 8B40DA24-8864-4E00-86FB-E20587237305 ![]()
Figures 24L, 25

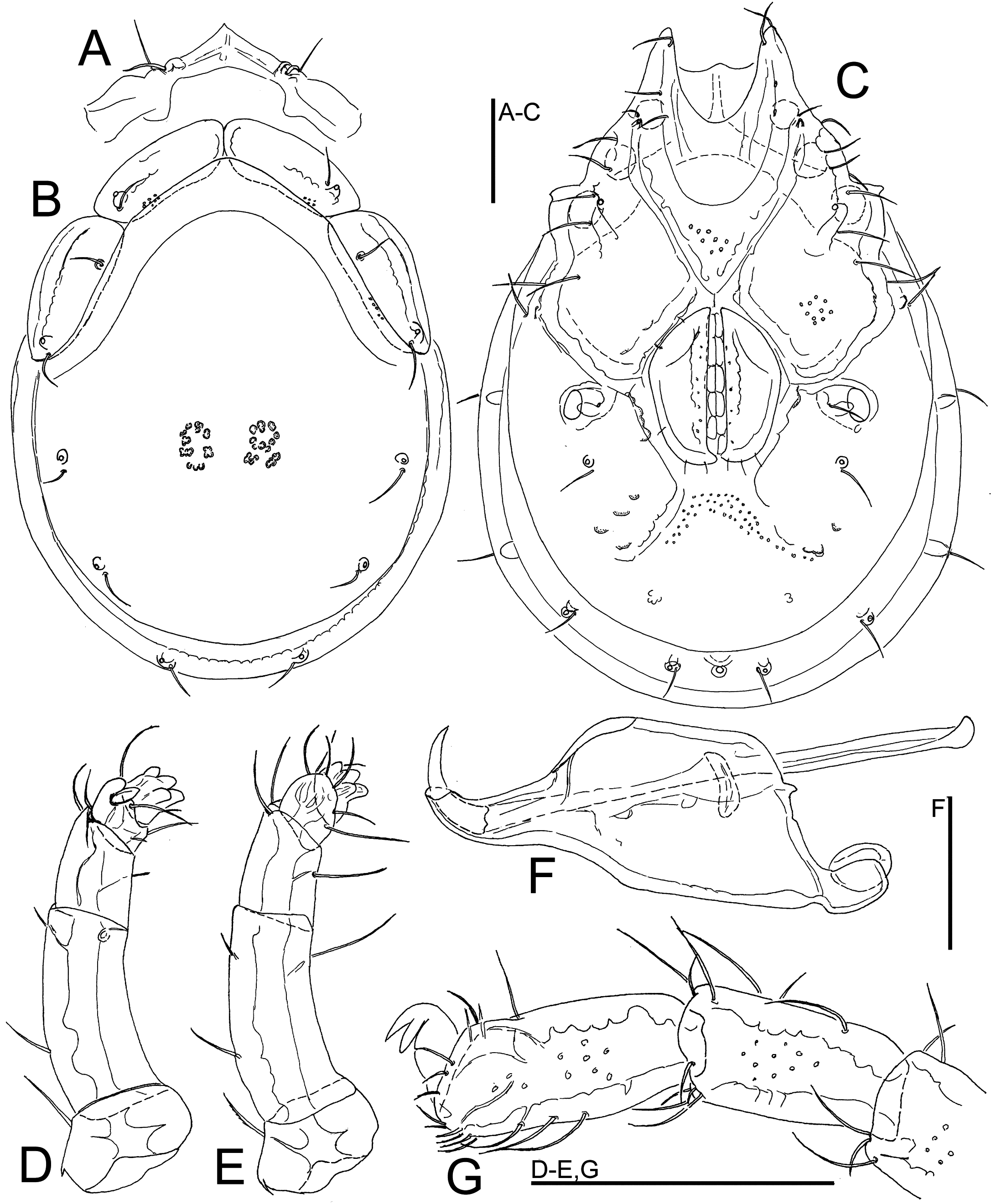
Material examined — Holotype ♀, dissected and slide mounted, Bhutan, MG5 Dakpay Chhu, 27.14621°N, 90.69220°E, 539 m asl, 27.x.2021.
Diagnosis (Male unknown) — Frontal and shoulder platelets relatively slender, and similar in dimensions; Cxgl-4 anterior to Cxgl-2 and located adjacent to II-Leg insertion; P-2 proportionally very long; ventral margin of P-2 and P-3 each with a moderately long seta, but without developed projections.
Description — Female – Idiosoma oval; dorsal shield without a colour pattern (as photographed in Figure 24K); area of primary sclerotization of the dorsal plate with four dorsoglandularia; shoulder platelets of similar dimensions as frontal platelets; gnathosomal bay shallow, U-shaped, proximally rounded; Cxgl-4 anterior to Cxgl-2 and located adjacent to II-Leg insertion; medial suture line of Cx-II+III short; genital field relatively large; suture lines of Cx-IV distinctly extending posteriorly beyond posterior margin of genital field; excretory pore and Vgl-2 away from the line of primary sclerotization, excretory pore on the level of Vgl-2; gnathosoma with a slightly curved ventral margin, rostrum relatively slender, cheliceral claw very large (Figure 25E); P-2 proportionally very long; ventral margin of P-2 and P-3 each with a moderately long seta, but without developed projections; P-4 and P-5 proportionally very short (Figure 25D).
Measurements – Idiosoma (ventral view: Figure 25C) L 677, W 478; dorsal shield (Figure 25B) L 544, W 425, L/W ratio 1.28; dorsal plate L 506; shoulder plate L 156, W 66, L/W ratio 2.4; frontal plate L 149–152, W 53–56, L/W ratio 2.7–2.8; shoulder/frontal plate L 1.03–1.05. Gnathosomal bay L 95, Cx-I total L 265, Cx-I mL 169, Cx-II+III mL 13; ratio Cx-I L/Cx-II+III mL 21.2; Cx-I mL/Cx-II+III mL 13.5. Genital field L/W 156/131, ratio 1.19; distance genital field-excretory pore 203, genital field-caudal idiosoma margin 242. Gnathosoma vL 309, chelicera L 363; palp total L 167, dL/H, dL/H ratio: P-1, 25/45, 0.55; P-2, 71/30, 2.4; P-3, 34/25, 1.36; P-4, 18/20, 0.9; P-5, 19/16, 1.15; L ratio P-2/P-4 3.9. dL of I-L-5-6: 75, 94; I-L-6 H 39; dL/H I-L-6 ratio 2.4.
Male – Unknown.
Etymology — The species is named after Tashi Wangmo (College of Natural Resources) for the long-lasting support of the junior author.
Discussion — The female of the new species resembles the female of Neoatractides suvarna (Cook, 1967), a species described from a small stream in Kerala State of India (Cook 1967). Females of both species share a P-2 separated from P-1, both P-2 and P-3 are without projections and the frontal plates are almost equal in size to the shoulder plates. The female of the latter species differs by a number of characters, i.e., shoulder and frontal plates broad, a colour pattern consisting of a posterior purple band and a smaller area of purple in the vicinity of the glandularia of the frontal platelets and the postocularia of shoulder platelets, palpal segments, especially P-2 are shortened, excretory pore located on a tubercle, and Cxgl-4 located close to Cxgl-2 (see Cook 1967).
Distribution — Bhutan (this study).
Acknowledgements
The fieldwork of this research was made possible by grants of the National Geographic Society (NGS-72271C-20) for a project titled ''Exploring the invertebrate diversity of the last virgin rivers of Bhutan, the Eastern Himalayas″ and a grant from the foundation Pro Acarologia Basiliensis (Basel). Vincent Kalkman (RMNH) coordinated the contact between Naturalis Biodiversity Center and institutions in Bhutan and assisted with the logistics. Moreover, we are indebted to Wim Klein and Oscar Vorst (RMNH) for collecting a number of water mites in Bhutan, and Mer Man Gurung would like to thank Cheten Dorji for his guidance and company during the collecting trips. We thank Joanna Mąkol (Wrocław), Hiroshi Abé (Fujisawa) and two anonymous reviewers, whose constructive comments greatly improved this work.
References
- Cook DR. 1967. Water mites from India. Mem. Amer. Entomol. Inst., 9: 1-411.
- Di Sabatino A., Gerecke R., Gledhill T., Smit H. 2010. Acari: Hydrachnidia II. In: Gerecke R, editor Chelicerata: Acari II. Süßwasserfauna von Mitteleuropa, 7, 2-2, 1-234. Elsevier Spektrum Akademischer Verlag, Heidelberg. https://doi.org/10.1007/978-3-8274-2266-8_1
- Gu X.-Y., Jin D.-C. & Guo J.-J. 2020a. Three new species and one new record of Torrenticolidae (Acari, Hydrachnidia) from Wuyishan with an updated key for Chinese fauna. Eur. J. Taxon., 625: 1-23. https://doi.org/10.5852/ejt.2020.625
- Gu X.-Y., Jin D.-C. & Guo J.-J. 2020b. Four new species of torrenticolid water mites with a newly recorded subgenus from Yunnan, China (Acari, Hydrachnidia, Torrenticolidae). Syst. Appl. Acarol., 25(8): 1495-1507. https://doi.org/10.11158/saa.25.8.11
- Lundblad, O. (1941) Neue Wassermilben. Vorläufige Mitteilung. Entomol. Tidskr., 62, 97-121.
- Lundblad O. 1956. Lundblad, O. (1956) Einige Wassermilben aus Java. Bollettino del Laboratorio di Entomologia Agraria "Filippo Silvestri", Portici, 33, 640-656.
- Lundblad O. 1967. Wassermilben von Hinterindien. Ark. Zool. (ser. 2), 19: 391-419.
- Lundblad O. 1969. Indische Wassermilben, hauptsächlich von Hinterindien. Ark. Zool., 22: 289-443.
- Jin D.C. 1997. Hydrachnellae-morphology systematics a primary study of Chinese fauna. Guizhou Science and Technology Publishing House, Guiyang, pp. 356. [in Chinese]
- Pesic V., Smit H. 2007. First records of water mites (Acari: Hydrachnidia) from Bhutan, with description of two new species. Zootaxa, 1613: 45-56. https://doi.org/10.11646/zootaxa.1613.1.3
- Pešić V., Smit H. 2014. Torrenticolid water mites (Acari: Hydrachnidia: Torrenticolidae) from Malaysian Borneo. Zootaxa, 3840 (1): 1-72. https://doi.org/10.11646/zootaxa.3840.1.1
- Pešić V., Kumar N., Kumar K. 2007. A new species of Monatractides (Acari: Hydrachnidia: Torrenticoli¬dae) and new records of other torrenticolid water mites from the Garhwal Himalayas (India). Syst. Appl. Acarol., 12(3-4), 205-212. https://doi.org/10.11158/saa.12.3.5
- Pešić V., Smit H. 2009. Water mites of the family Torrenticolidae Piersig, 1902 (Acari: Hydrachnidia) from Thailand, Part I. The genera Torrenticola Piersig, 1896, Neoatractides Lundblad, 1941 and Pseudotorrenticola Walter, 1906. Zootaxa, 1982: 38-62. https://doi.org/10.11646/zootaxa.1982.1.2
- Pešić V., Smit H. 2014. Torrenticolid water mites (Acari: Hydrachnidia: Torrenticolidae) from Malaysian Borneo. Zootaxa, 3840(1): 1-72. https://doi.org/10.11646/zootaxa.3840.1.1
- Pešić, V., Chatterjee, T., Das, M. & Bordoloi, S. (2012) Two rare water mite species (Acari, Hydrachnidia) from the streams of the Indian eastern Himalayan region. Syst. Appl. Acarol., 17(4), 458-464. https://doi.org/10.11158/saa.17.4.15
- Pešić V., Chatterjee T., Das M., Bordoloi S. 2013. A new species of water mite (Acari, Hydrachnidia) from Assam, India, found in the gut contents of the fish Botia dario (Botiidae). Zootaxa, 3746(3): 454-462. https://doi.org/10.11646/zootaxa.3746.3.4
- Pešić V., Smit H., Bahuguna P. 2019a. New records of water mites (Acari: Hydrachnidia) from the Western Himalaya with the description of four new species. Syst. Appl. Acarol., 24: 59-80. https://doi.org/10.11158/saa.24.1.5
- Pešić V., Smit H., Bahuguna P. 2019b. New records of water mites (Acari: Hydrachnidia) from the Western Himalaya and description of three new species from Asia. Syst. Appl. Acarol., 24: 1868-1880. https://doi.org/10.11158/saa.24.10.6
- Pešić V., Smit H., Negi, S., Bahuguna P. & Dobriyal, A.K. 2020. Torrenticolid water mites of India with description of three new species (Acari: Hydrachnidia, Torrenticolidae) Syst. Appl. Acarol., 25(2): 255-267. https://doi.org/10.11158/saa.25.2.7
- Sokolow I. 1926. Zwei neue Hydracarinenarten aus der Gattung Atractides C.L.Koch. Zool. Anz., 68: 72-76.
- Wangchuk J., Yoezer D., Wangdi, N., Wangdi K., Singye R., Dorji T. 2017. Macroinvertebrate and fish diversity in Mangdechhu Hydropower Plant, Trongsa Bhutan. NeBIO., 8(4): 335-342.
- Wiles P.R. 1991. Rheophilic watermites (Acari: Hydrachnidia) from mainland Malaysia. Acarologia, 32(1): 41-56.
- Wiles P.R. 1997. Asian and Oriental Torrenticolidae Piersig, 1902 (Acari: Hydrachnidia: Lebertioidea): a revision of the family and description of new species of Torrenticola Piersig and Pseudotorrenticola Walter, from Southeast Asia. J. Nat. Hist., 31: 191-236. https://doi.org/10.1080/00222939700770121



2022-06-01
Date accepted:
2022-08-24
Date published:
2022-09-01
Edited by:
Mąkol, Joanna

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Pešić, Vladimir; Smit, Harry and Gurung, Mer Man
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)