



Re-description of the female of Kampimodromus langei Wainstein & Arutunjan (Acari: Phytoseiidae) based on normal and abnormal specimens, with the first description of its male
Saglam, Damla  1
; Doker, Ismail
2
and Ozman-Sullivan, Sebahat K.
3
1
; Doker, Ismail
2
and Ozman-Sullivan, Sebahat K.
3
1Department of Plant Protection, Faculty of Agriculture, Ondokuz Mayis University, 55139, Samsun, Turkey.
2✉ Department of Plant Protection, Faculty of Agriculture, Cukurova University, 01330 Adana, Turkey.
3✉ Department of Plant Protection, Faculty of Agriculture, Ondokuz Mayis University, 55139, Samsun, Turkey.
2022 - Volume: 62 Issue: 2 pages: 446-453
https://doi.org/10.24349/d6cm-8rlyOriginal research
Keywords
Abstract
Introduction
The members of the mite genus Kampimodromus Nesbitt (Acari: Phytoseiidae) are considered to be Type III generalist predators living on pubescent leaves (McMurtry et al. 2013). Among them, K. aberrans (Oudemans) is an effective predator of phytophagous mites on a number of crops that include apples, grapes and hazelnuts (Kreiter et al. 2000; Kasap 2005; Ozman-Sullivan 2006; Duso et al. 2014). Natural populations of some other Kampimodromus species such as K. ericinus Ragusa di Chiara & Tsolakis, K. keae (Papadoulis & Emmanouel), K. langei Wainstein & Arutunjan and K. ragusai Swirski & Amitai have been reported from Quercus spp. trees in some European countries (Ragusa di Chiara and Tsolakis 1994; Tixier et al. 2008; Papadoulis et al. 2009; Cargnus et al. 2012; Doker et al. 2017, 2018; Faraji and Hoekstra 2021).
In this study, we report K. langei for the first time for Turkish fauna. It is re-described and illustrated, based on normal and abnormal female specimens. Also, the male is described for the first time in this study.
Material and methods
This study was conducted in 2021 in several remnant oak forests on the Kurupelit campus of Ondokuz Mayis University in Samsun Province, Turkey. Most of the surveys were done periodically in June, July, August, September and October. Leaves of oak trees were inspected with the aid of a 10X hand magnifying lens and those with phytoseiid mites were collected. The collected leaves were wrapped in paper, placed in a labeled plastic bag and stored in an ice-box until transferred to the laboratory. They were then refrigerated until checked for mites under a stereomicroscope. All of the collected mites were stored in 70% alcohol until cleared in lactophenol and mounted on slides in Hoyer's medium. The slides were then kept in an incubator at 55 °C for one week. The mites were identified and illustrated by the second author. The permanent slides were examined under an Olympus® CX-41 microscope. Illustrations were prepared by using a U-Da drawing attachment (Camera Lucida). The taxonomic system used followed Chant and McMurtry (2007), and the setal nomenclature used followed Lindquist and Evans (1965), as adapted by Rowell et al. (1978). The dorsal and ventral setal pattern used was that of Chant and Yoshida-Shaul (1989, 1991, 1992). The nomenclature for solenostomes for the dorsal idiosoma follows that of Athias-Henriot (1975). Leg chaetotaxy follows that of Evans (1963). Measurements are given in micrometers and presented as the mean, followed by the range in parentheses. The examined specimens are deposited in the mite collection of the Acarology Laboratory, Cukurova University, Adana, Turkey.
New record for Turkey
Kampimodromus langei Wainstein & Arutunjan
(Figures 1, 2)
Kampimodromus langei Wainstein & Arutunjan, 1973: 55, Fig. 2.
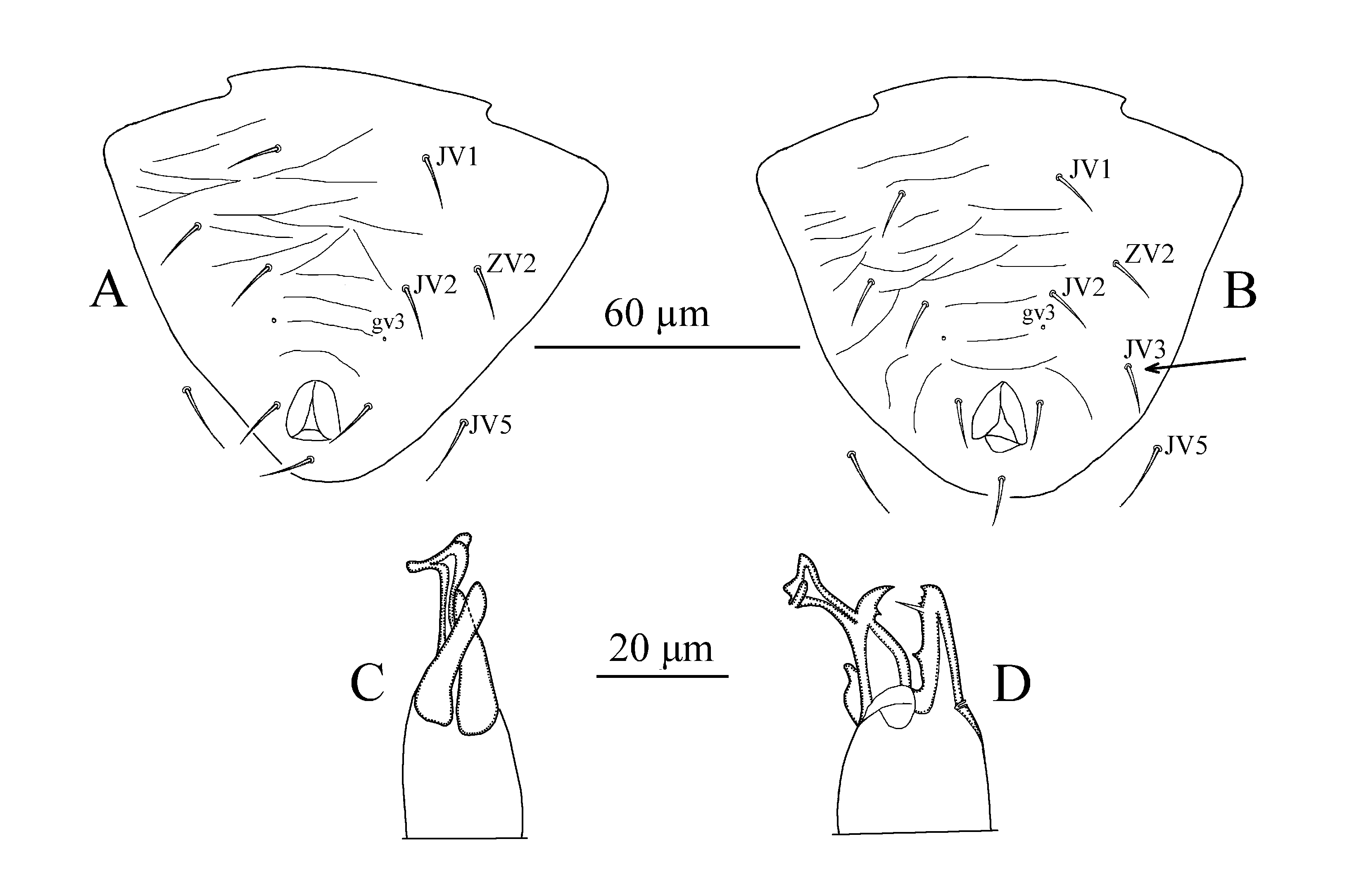
Diagnosis — Dorsal setal pattern 10A:8C, one female specimen with extra dorsal seta on both sides, dorsal shield striated, bearing five pairs of large solenostomes (gd1, gd2, gd6, gd8 and gd9). Dorsal setae serrated, except j4, j5, j6, J2, J5 and Z1 smooth, J2 and Z1 with few barbs in some specimens. Peritreme extending to level of seta z2. Sternal shield smooth with three pairs of setae, genital shield smooth. Ventrianal shield smooth with three pairs of preanal setae and one pair of rounded pores. Calyx of spermatheca cup-shaped; atrium small, nodular, attached to calyx without neck. Fixed digit with three teeth, and movable digit with one tooth. Genu II and Genu IV each with eight setae, and tibia IV seven setae. Macroseta StIV present and with blunt tip. Male dorsal shield reticulated and with setae r3 and R1. Male ventrianal shield striated, with three pairs of setae (JV1, JV2 and ZV2), and one specimen had unpaired seta JV3 on right side. Male spermatodactyl L-shaped with slightly developed toe.
Re-description
Female
(n=10)

Dorsum — (Figures 1A, B). Dorsal setal pattern 10A:8C (r3 and R1 off shield), one specimen with an extra dorsal seta on both sides (SS present, identical to location of S1). Dorsal shield striated, with waist at level of seta R1. A lateral cavity anterior to seta S5 on right side visible on the illustrated specimen. Bearing five pairs of solenostomes (gd1, gd2, gd6, gd8 and gd9). Muscle marks (sigillae) visible mostly on podosoma; length of dorsal shield 300 (275–310), width (at level of s4) 149 (145–153), width (at level of S2) 158 (155–160). Dorsal setae serrated, except j4, j5, j6, J2, J5 and Z1 smooth. Setae J2 and Z1 with few barbs in some specimens. Measurements of dorsal setae as follows: j1 19 (18–20), j3 27 (25–30), j4 12 (10–13), j5 11 (10–13), j6 13 (13–15), J2 20 (18–23), J5 5 (4–6), z2 27 (25–28), z4 29 (28–30), z5 11 (10–13), Z1 18 (15–20), Z4 41 (40–43), Z5 48 (45–50), s4 33 (30–35), SS 35 (only one specimen), S2 38 (35–40), S5 19 (18–20), r3 38 (35–40) and R1 27 (25–30). Peritreme extending to level of seta z2.
Venter — (Figure 1C). Ventral setal pattern 14: JV-3:ZV. Sternal shield smooth, lightly sclerotized; with irregular posterior margin, three pairs of setae (ST1, ST2 and ST3) and two pairs of pores (iv1 and iv2); length (ST1–ST3) 56 (55–58), width (ST2–ST2) 56 (55–58); metasternal setae ST4 and one pair of pores (iv3) on metasternal shields. Genital shield smooth, width at level of genital setae (ST5) 44 (43–45). Ventrianal shield elongate, with waist at level of seta JV2. Three pairs of pre-anal setae (JV1, JV2 and ZV2); and one pair of rounded pores (gv3) posteromedian to JV2, distance between pre-anal pores 22. One pair of para-anal seta (Pa) and unpaired post-anal seta (Pst). Length of ventrianal shield 91 (88–95), width at level of setae ZV2 (widest point) 51 (50–53), at level of anus (at level of muscle marks, widest point) 46 (45–48). Setae ZV1, ZV3, JV4 and JV5 and six pairs of poroids on integument surrounding ventrianal shield. Setae JV5 serrated, longer than other ventral setae, 27 (25–28) in length.
Chelicera — (Figure 1D). Fixed digit 23 (23–25) long with three apical teeth, and pilus dentilis; movable digit 23 (23–25) long, with one tooth.
Spermatheca — (Figure 1E). Calyx short, cup-shaped, flaring distally, 8 (8–10) in length. Atrium small, nodular; attached to calyx without neck. Major duct long, minor duct not visible.
Legs — (Figure 1F). Length of legs (base of coxae to base of claws): leg I 237 (230–240); leg II 191 (185–195); leg III 174 (168–175); leg IV 222 (215–225). Macrosetae StIV blunt tip 18 (18–20) in length. Chaetotaxy as follows: Leg I: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 2 3/1 2/2 2, genu 2 2/1 2/1 2, tibia 2 2/1 2/1 2. Leg II: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 2 3/1 2/1 1, genu 2 2/1 2/0 1, tibia 1 1/1 2/1 1. Leg III: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 1 2/1 1/0 1, genu 1 2/1 2/0 1, tibia 1 1/1 2/1 1. Leg IV: coxa 0 0/1 0/0 0, trochanter 1 0/1 0/2 1, femur 1 2/1 1/0 1, genu 1 2/1 2/0 2, tibia 1 1/1 2/0 2.
Male
(n=9)
Similar to female in many aspects.
Dorsum — Dorsal setal pattern 10A:8C (r3 and R1 on shield). Dorsal shield sclerotized, reticulated, bearing five pairs of solenostomes (gd1, gd2, gd6, gd8 and gd9); length of dorsal shield 220 (218–223), width (at level of s4) 116 (113–118). Serration of dorsal setae as in female. Measurements of dorsal setae as follows: j1 14 (13–15), j3 27 (25–30), j4 12 (10–13), j5 12 (10–13), j6 14 (13–15), J2 25 (23–26), J5 5 (4–6), z2 29 (28–30), z4 34 (33–35), z5 14 (13–15), Z1 19 (18–20), Z4 31 (30–33), Z5 33 (30–35), s4 41 (38–42), S2 32 (30–36), S5 11 (10–12), r3 31 (30–33) and R1 22 (20–25).
Peritreme extending to level of seta z2.
Venter — (Figure 2A, B). Sternogenital shield smooth, sclerotized, with five pairs of setae (ST1, ST2, ST3, ST4 and ST5). Distance between bases of ST1–ST5 98 (95–100). Ventrianal shield triangular, striated, bearing three pairs of pre-anal setae (JV1, JV2 and ZV2) (Figure 2A), (one specimen had an unpaired seta (JV3 on left side) (Figure 2B), a pair of para-anal (Pa) and unpaired post-anal seta (Pst), and with one pair of rounded pores (gv3) posteromedian to JV2, distance between pre-anal pores 19 (18–20). Length of ventrianal shield 95 (93–98), width (at anterior corners widest point) 105 (99–110). Setae JV5 slightly serrated, 15 (13–18) in length.
Chelicera — (Figure 2C, D). Fixed digit with three teeth, two apical and one well-behind pilus dentilis; movable digit with one tooth; spermatophoral process L-shaped, with toe slightly developed. The left and the right sides of the illustrated specimen, as well as some other specimens on both sides seemed to have different shape of spermatophoral process, with toe twisted in those cases.
Legs — Length of legs (base of coxae to base of claws): leg I 207 (205–210); leg II 191 (188–193); leg III 149 (148–150); leg IV 210 (205–213). Basitarsus IV with short macroseta, StIV 16 (15–18) in length. Leg chaetotaxy as in female.
Material examined
Ten females and 9 males were collected from Q. cerris var. cerris in remnants of oak forest on the Kurupelit campus Ondokuz Mayis University in Samsun Province, 41°21′58.9″N, 36°11′18.5″E: two females, 6 May 2021; one female, 27 May 2021; two males, 29 July 2021; two females, 18 August 2021; four females and six males, 28 August 2021; one female and one male, 18 September 2021.
World Distribution
Armenia (Wainstein and Arutunjan 1973), Russia (Meshkov 1999), Norway (Denmark and Edland 2002), Moldova (Kulikova 2011), Croatia, Italy (Cargnus et al. 2012), The Netherlands (Faraji and Hoekstra 2021) and Turkey (this study).
Remarks
Kampimodromus langei was described by Wainstein and Arutunjan (1973) based on the material collected from Quercus sp. (Fagaceae) in Stepanavan, Armenia. Our collection of this species in Turkey represents a new record for the country as previous studies of the genus Kampimodromus did not include K. langei (Ozman and Cobanoglu 2001; Cobanoglu and Ozman 2002; Kasap and Cobanoglu 2009; Yesilayer and Cobanoglu 2011; Doker et al. 2017, 2018). In addition, the male K. langei is described for the first time in this study. Morphological characters and measurements of the current specimens are similar to those of the original description and subsequent re-descriptions (Wainstein and Arutunjan 1973; Ragusa di Chiara and Tsolakis 1994; Tixier et al. 2008; Cargnus et al. 2012; Faraji and Hoekstra 2021). However, a single female specimen showed an abnormality in having an additional dorsal seta on both sides, with all other morphological characters and measurements of the specimen being almost identical to the other specimens examined in this study, as well as the earlier descriptions. To the best of our knowledge, the extra seta inserted at the location of S1 has previously been reported by Kolodochka (2006) for Typhlodromus (Typhlodromus) laurae Arutunjan, but is not known in any other Phytoseiidae species to date (Rowell et al. 1978; Chant and Yoshida-Shaul 1989; Chant and McMurtry 2007; Ferragut et al. 2010). However, it is not known whether the extra seta reported for T. (T.) laurae was paired or unpaired because Kolodochka (2006) illustrated only the right side of the specimen.
This extra seta could be considered seta z6 or s6 because both are already known for some members of the family Phytoseiidae (Chant and McMurtry 2007; Papadoulis et al. 2009). However, when those setae are present on other phytoseiid species their insertion is clearly anterior to seta R1 (Kreiter et al. 2002; Faraji et al. 2008; Ferragut et al. 2010; Demite et al. 2020). However, the extra setae described in this study were located posterior to R1. In addition, when the podosomal setae z6 and s6 are present in phytoseiids their insertion is clearly anterior to solenostome gd6 (Chant and McMurtry 2007; Papadoulis et al. 2009) which is not the case in the abnormal specimens reported by Kolodochka (2006) and in this study. Kolodochka (2006) named this seta, SS. Therefore, we have used the same notation and have described this seta as SS.
Instead of being called an abnormality, the presence of extra seta/e can also be considered to be a kind of variation that has been reported for some phytoseiid species such as Amblyseius palustris (Chant), Euseius finlandicus (Oudemans), Neoseiulus agrestis (Karg), N. longispinosus (Evans), Phytoseius echinus Wainstein & Arutunjan, Typhlodromus (Anthoseius) caudiglans Schuster, T. (A.) commenticus Livshitz & Kuznetsov, T. (T.) pyri Scheuten (Chant and Yoshida-Shaul 1989; Kolodochka 2006; Silva et al. 2020). In addition, Kolodochka (1985) reported a type specimen of T. (T.) rarus Wainstein without setae j3 and one each of S5 and Z4 but with an unpaired seta in the region of seta Z4. However, unpaired and unknown setae on the dorsal shield of phytoseiid mites can also represent a new taxonomic status as Demite et al. (2017) recently described a new genus and a new species, Amazoniaseius imparisetosus Demite, Cruz & McMurtry, with two unpaired setae.
In addition, an unpaired seta (JV3) was also observed on the ventrianal shield of a male specimen examined in this study. We consider the presence of this unpaired seta to be a variation because it has been already reported for A. nemorivagus Athias-Henriot and N. bicaudus (Wainstein) by Papadoulis et al. (2009).
Acknowledgements
This research is part of the first author's Master's thesis and was supported by the Scientific Research Foundation of Ondokuz Mayis University in Samsun, Turkey (Project number: PYO.ZRT.1904.21.014). The authors thank Prof. Dr. Hamdi G. Kutbay of Ondokuz Mayis University for his identification of the oak trees and Dr. Gregory T. Sullivan of The University of Queensland for comments and editing.
References
- Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini. II. Le relevé organotaxique de la facedorsale adulte (Gamasides, protoadéniques, Phytoseiidae). Acarologia, 17(1): 20-29.
- Cargnus E., Girolami V., Zandigiacomo P. 2012. Re-examination of specimens of three species of Kampimodromus Nesbitt (Acari: Phytoseiidae) from north-eastern Italy, with first report on Kampimodromus corylosus Kolodochka in Italy. Int. J. Acarol., 38: 583-594. https://doi.org/10.1080/01647954.2012.710247
- Chant D.A., McMurtry J.A. 2007. Illustrated keys and diagnoses for the genera and subgenera of the Phytoseiidae of the world (Acari: Mesostigmata). West Bloomfield: Indira Publishing House. pp. 219.
- Chant D.A., Yoshida-Shaul E. 1989. Adult dorsal setal patterns of the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol., 15: 219-233. https://doi.org/10.1080/01647958908683852
- Chant D.A., Yoshida-Shaul E. 1991. Adult ventral setal patterns in the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol., 17: 187-199. https://doi.org/10.1080/01647959108683906
- Chant D.A., Yoshida-Shaul E. 1992. Adult idiosomal setal patterns in the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol., 18: 177-193. https://doi.org/10.1080/01647959208683949
- Cobanoglu S., Ozman S.K. 2002. Beneficial mite species of hazelnut orchard ecosystems from the Black Sea Region of Turkey. Proceedings of the 2nd Meeting of WG 4: Prague 30-31st May 2002, Bio-control of Arthropod Pests in the Stored Products: 91-99.
- Demite P.R., Souza R.B., Cavalcante A.C.C. 2020. A new species of Phytoseius Ribaga (Acari: Mesostigmata: Phytoseiidae) from the Amazon Forest, Brazil, with a redefinition of the purseglovei species group Chant & Yoshida-Shaul. Syst. Appl. Acarol., 25: 1350-1358. https://doi.org/10.11158/saa.25.7.14
- Demite P.R., Cruz W.P. da, McMurtry J.A., Moraes G.J. de 2017. Amazoniaseius imparisetosus n.sp., n.g.: an unusual new phytoseiid mite (Acari: Phytoseiidae from the Amazon forest. Zootaxa, 4236: 302-310. https://doi.org/10.11646/zootaxa.4236.2.5
- Denmark H.A., Edland T. 2002. The subfamily Amblyseiinae Muma (Acari: Phytoseiidae) in Norway. Int. J. Acarol., 28: 195-220. https://doi.org/10.1080/01647950208684296
- Doker I., Kazak C., Karaca M.M., Karut K. 2017. Two new records of the genus Kampimodromus Nesbitt (Acari: Phytoseiidae) for Turkey with a revised key to the World species. Acarologia, 57: 355-363. https://doi.org/10.1051/acarologia/20164160
- Doker I., Karut K., Karaca M.M., Cargnus E., Kazak C. 2018. Internal Transcribed Spacer (ITS) sequences of some Kampimodromus (Acari: Phytoseiidae) species: Is Kampimodromus ragusai a valid species or a synonym of Kampimodromus aberrans? Syst. Appl. Acarol., 23: 2237-2243. https://doi.org/10.11158/saa.23.11.15
- Duso C., Ahmad S., Tirello P., Pozzebon A., Klaric V., Baldessari M., Malagnini V., Angeli G. 2014. The impact of insecticides applied in apple orchards on the predatory mite Kampimodromus aberrans (Acari: Phytoseiidae). Exp. Appl. Acarol., 62: 391-414. https://doi.org/10.1007/s10493-013-9741-3
- Evans, G.O. 1963. Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bull. Br. Mus. Nat. Hist. Zool., 10: 275-303. https://doi.org/10.5962/bhl.part.20528
- Faraji F., Hoekstra P.H. 2021. Some new species records of the predatory mite family Phytoseiidae (Acari: Mesostigmata) from The Netherlands. Soil Org., 93: 35-57. https://doi.org/10.25674/so93iss1pp35
- Faraji F., Sakenin-Chelav H., Kamali K., McMurtry J.A. 2008. Four new species records of Phytoseiidae (Acari: Mesostigmata) for Iran, and description of variability in the spermatheca of Typhlodromus bakeri. Syst. Appl. Acarol., 13: 123-132. https://doi.org/10.11158/saa.13.2.5
- Ferragut F., Pérez Moreno I., Iraola V., Escudero A. 2010. Ácaros depredadores em las plantas cultivadas. Família Phytoseiidae. Madrid: Ediciones Agrotécnicas. pp. 202.
- Kasap I. 2005. Life-history traits of the predaceous mite Kampimodromus aberrans (Oudemans) (Acarina: Phytoseiidae) on four different types of food. Biol. Control, 35: 40-45. https://doi.org/10.1016/j.biocontrol.2005.05.014
- Kasap I., Çobanoğlu S. 2009. Phytoseiid mites of Hakkâri province, with Typhlodromus (Anthoseius) tamaricis Kolodochka, 1982 (Acari: Phytoseiidae), a new record for the predatory mite fauna of Turkey. Turk. Zool. Derg., 33: 301-308. https://doi.org/10.3906/zoo-0805-9
- Kolodochka L.A. 1985. A redescription of Typhlodromus rarus (Parasitiformes, Phytoseiidae). Vestn. Zool., 6: 16-22. [in Russian].
- Kolodochka L.A. 2006. Phytoseiid mites of the Palaearctic Region (Parasitiformes, Phytoseiidae): faunistic, taxonomy, ecomorphology, evolution. Vestn. Zool., Suppl., 21: 1-250. [in Russian].
- Kreiter S., Ueckermann E.A., Quilici S. 2002. Seven new phytoseiid species, with a new generic assignement and a key to the species of La Reunion Island (Acari: Mesostigmata). Acarologia, 42: 335-350.
- Kreiter S., Tixier M.S., Auger P., Muckensturm N., Sentenac G., Doublet B., Weber M. 2000. Phytoseiid mites of vineyards in France (Acari: Phytoseiidae). Acarologia, 41: 77-96.
- Kulikova, L. 2011. Mites of fruit plantations of the Republic of Moldova. Muzeul Olteniei Craiova. Oltenia. Studii şi comunicări. Ştiinţele Naturii, 27: 55-62.
- Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Mem. Ent. Soc. Can., 47: 1-64. https://doi.org/10.4039/entm9747fv
- McMurtry J.A., Moraes, G.J. de, Famah Sourassou N. 2013. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol., 18: 297-320. https://doi.org/10.11158/saa.18.4.1
- Meshkov Yu.I. 1999. Contribution to phytoseiid fauna (Parasitiformes, Phytoseiidae) of Moscow District. Zool. Zhurnal, 78: 426-431 [in Russian].
- Ozman-Sullivan S.K. 2006. Life history of Kampimodromus aberrans as a predator of Phytoptus avellanae (Acari: Phytoseiidae, Phytoptidae). Exp. Appl. Acarol., 38: 15-23. https://doi.org/10.1007/s10493-005-5786-2
- Ozman S.K., Çobanoğlu S. 2001. Current status of hazelnut mites in Turkey. Acta Hortic., 56: 479-487. https://doi.org/10.17660/ActaHortic.2001.556.70
- Papadoulis G.Th., Emmanouel N.G., Kapaxidi, E.V. 2009. Phytoseiidae of Greece and Cyprus (Acari: Mesostigmata). West Bloomfield: Indira Publishing House. pp. 200.
- Ragusa di Chiara S., Tsolakis H. 1994. Revision of the genus Kampimodromus Nesbitt, 1951 (Parasitiformes, Phytoseiidae), with a description of a new species. Acarologia, 35: 305-322.
- Rowell H.L., Chant D.A., Hansell R.I.C. 1978. The determination of setal homologies and setal patterns on the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata). Can. Entomol., 110: 859-876. https://doi.org/10.4039/Ent110859-8
- Silva D.E., Nascimento J.M. do, Meira A. de A., Corrêa L.L.C., Johann L., Rodrigues R., Ferla N.J. 2020. Intraspecific variation of Typhlodromus (Typhlodromus) pyri Scheuten in vineyards of Northwest Portugal. Int. J. Acarol., 46: 60-62. https://doi.org/10.1080/01647954.2019.1695934
- Tixier M.-S., Kreiter S., Croft B.A., Cheval B. 2008. Kampimodromus aberrans (Acari: Phytoseiidae) from the USA: morphological and molecular assessment of its density. Bull. Entomol. Res., 98: 125-134. https://doi.org/10.1017/S0007485307005457
- Wainstein B.A., Arutunjan E.S. 1973. New species of predatory mites of the family Phytoseiidae (Parasitiformes) in Armenian SSR. Doklady Akademii Nauk Armyanskoy SSR, 56: 55-58. [in Russian].
- Yesilayer A., Çobanoğlu S. 2011. The distribution of predatory mite species (Acari: Phytoseiidae) on ornamental plants and parks of Istanbul, Turkey. Türk. Entomol. Bült., 1: 135-143.



2022-03-16
Date accepted:
2022-04-14
Date published:
2022-04-19
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Saglam, Damla; Doker, Ismail and Ozman-Sullivan, Sebahat K.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)