



Two new species of feather mites (Acarina: Psoroptidia) from the Oriental Magpie-Robin, Copsychus saularis (Passeriformes: Muscicapidae)
Constantinescu, Ioana Cristina1 ; Chişamera, Gabriel Bogdan2 ; Petrescu, Angela3 and Costică, Adam4
1✉ Grigore Antipa National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest, Romania.
2Grigore Antipa National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest, Romania.
3Grigore Antipa National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest, Romania.
4Grigore Antipa National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest, Romania.
2018 - Volume: 58 Issue: 2 pages: 313-331
https://doi.org/10.24349/acarologia/20184244ZooBank LSID: 4ED9857C-3E53-4396-80F8-289BBE6A9DD8
Keywords
Abstract
With over 100 known species, Indonesian archipelago is ranked 11th in the world with respect to recorded feather mite biodiversity (Doña et al. 2016). Studies on feather mites in this area began over 100 years ago, with the first papers realized by early acarologists on the feather mite fauna associated with parrots (Psittaciformes) (Trouessart 1884, 1885; Favette and Trouessart 1904). The fauna of parrots in this country, particularly varied and spectacular, with endemic or rare species, has continued to fascinate acarologists over time; so other feather mites species from these hosts have been described or reinvestigated in a series of papers published in the past forty years (Atyeo and Pérez 1982; Atyeo and Gaud 1991; Gaud and Atyeo, 1996; Mironov and Perez 2003; Mironov and Dabert 2010; Mironov et al. 2003, 2014). A comprehensive paper, dedicated to feather mites that live on birds from the family Megapodidae (Galliformes), includes 34 species of feather mites from Indonesia (Atyeo 1992). Descriptions of feather mite species from other hosts orders in Indonesia are scattered in various taxonomic papers (Bonnet 1924; Atyeo and Braasch 1966; Santana 1976; Peterson and Atyeo 1977; Pérez and Atyeo 1981; Gaud et al. 1985; Gaud and Atyeo 1986, 1987; Atyeo and Gaud 1992; Dabert and Ehrnsberger 1996; Mironov et al. 2002, Dabert et al. 2002; Mironov 2006; Dabert and Labrzycka 2009). Atyeo (in McClure and Ratanaworabhan 1973) reported 11 species of feather mites from this country, 9 of which are presumably new species. There are 1,748 species of birds in Indonesia (Clements et al. 2016), and 609 in Kalimantan (Indonesian Borneo) (Lepage 2017). Each is a potential host for several feather mite species, so it is evident that the investigation of this group in this country is far from being exhaustive. The genus Copsychus (Passeriformes: Muscicapidae) comprises 11 species of magpie-robins and shamas, distributed in Madagascar, South Asia, Indonesia and the Philippines (Clements et al. 2016). The Oriental Magpie Robin, Copsychus saularis (Linnaeus) is a small sized species distributed in the tropical area of southern Asia, being native to Bangladesh, Bhutan, Brunei Darussalam, Cambodgia, China, India, Indonesia, Laos People's Democratic Republic, Malaysia, Myanmar, Nepal, Pakistan, Singapore, Sri Lanka, Thailand, Vietnam, and was introduced to Taiwan (BirdLife International 2017). It is an anthropophilic bird, common in forests, parks, gardens and feeds on insects and other small invertebrates on the ground (MacKinnon and Phillipps, 1993).
The goal of the present paper is to describe two new feather mite species belonging to genera Trouessartia and Dolichodectes, collected from the Oriental Magpie-Robin, Copsychus saularis.
Mites were collected by the authors from museum specimens of birds preserved in ethanol, from the Ornithological Collection of “Grigore Antipa” National Museum of Natural History (Bucharest, Romania). The ethanol in which the birds were conserved was filtered through filter paper, and mite specimens were collected manually with a needle, and placed in tubes with 96% ethanol. Then mite specimens were cleared in 90 % lactic acid for 24 hours, and mounted on microscope slides in Hoyer’s medium. Drawings were made using an Olympus CX21 microscope, equipped with a camera lucida drawing device. The taxonomy of birds follows Clements et al. (2016). The body setation of mites follows that of Griffiths et al. (1990) with the modifications of Norton (1998) concerning coxal setae, while the leg setation follows Gaud and Atyeo (1996). In the descriptions, we used a set of standard diagnostic characters used for species of the family Trouesartiidae (Mironov and González-Acuña 2013; Hernandes 2014; Hernandes and Valim 2015) and Dolichodectes (Hernandes and Valim 2006, Mironov et al. 2010, 2012), respectively.
Measurements are given for the holotype (male) and followed by ranges for paratypes. All measurements are in micrometers (μm). The mite specimens studied herein are deposited in the Acari Collection of “Grigore Antipa” National Museum of Natural History, Bucharest, Romania (MGAB) and in the Acari Collection of the Department of Zoology of the Universidade Estadual Paulista, Rio Claro, São Paulo, Brazil (DZUnesp-RC).
Subfamily Pterodectinae Park et Atyeo, 1971
Genus Dolichodectes Park et Atyeo, 1971
The feather mite genus Dolichodectes Park and Atyeo, 1971 currently includes nine species; one of them is widespread throughout the Old Word, one is described from Europe, one is from Asia, and six other species are recorded from Africa (Trouessart 1885; Gaud and Mouchet 1957; Mironov and Fain 2003; Mironov et al. 2010, 2012, 2015). Atyeo (in: McClure and Ratanaworabhan 1973) reported 16 undetermined and presumably new species of this genus from Asia. To date, only one species of the genus Dolichodectes has been described from birds of the genus Copsychus: Dolichodectes furcilobus Mironov, Literák, Hung and Čapek, 2012 from the White-rumped Shama, Copsychus malabaricus (Scopoli) (Passeriformes: Muscicapidae) in Vietnam (Mironov et al. 2012). Of the nine species of the genus, three species were identified from birds of the family Muscicapidae, two species from birds of the family Acrocephalidae, and one from birds of the families Platysteiridae, Turdidae, Stenostiridae and Passeridae. Dolichodectes latilobus is the fourth species described from birds of the family Muscicapidae.
ZOOBANK: CF94906A-A01F-4B71-AE9E-6AA3599C88A7 ![]()
Type material — Holotype male and 16 paratypes (5 males and 11 females), from Copsychus saularis (Linnaeus) (Passeriformes: Muscicapidae), Indonesia, Kalimantan Island, Lhok Tuan, east of the Kutai National Park, 18 May 1991, bird inventory number 15719-47, no other data.
Type deposition — Holotype male (ANA 787), 4 male (ANA 788 – 791) and 9 female (ANA 792 – 800) paratypes in MGAB collection, 1 male and 2 female paratypes in DZUnesp-RC collection.
Etymology — The specific epithet refers to the widened posterior ends of the opisthosomal lobes in males (from Latin latus = wide, broad).
Description — MALE (holotype, range for 4 paratypes) — Length of idiosoma 436 (456 – 472), width 124 (112 – 120), length of hysterosoma 340 (328 – 342). Prodorsal shield: anterior margin with triangular rostral process, lateral margins without incisions around scapular setae, posterior margin concave, antero-lateral extensions widely connected to bases of epimerites Ia, length along midline excluding rostrum 120 (122 – 134), width at posterior margin 102 (82 – 112), posterior part with wavy transverse striae. Setae ve represented by alveoli. Scapular setae se separated by 52 (50 – 60). Humeral shields present, poorly developed, separated from epimerites III, not encompassing setae cp. Setae c2 situated on soft tegument, near anterior end of humeral shields. Setae c3 lanceolate, 20 (20 – 24) x 8 (7 – 8). Hysteronotal shield: greatest length from anterior margin to lobar apices 240 (240 – 245), width at anterior margin 96 (96 – 114), anterior margin almost straight, strongly narrowed between levels of setae d2 and e1, anterior half of this shield with wavy transverse striae, area from level of trochanters IV to bases of opisthosomal lobes with small ovate lacunae. Opisthosomal lobes widened in posterior half, posterior end of each lobe with a pair of narrow divergent extensions (outer extension rounded apically, inner extension acute), lateral margins at level of setae h2 convex (Fig. 1A). Terminal cleft a narrow parallel-sided slit, with lateral margins touching, length 56 (50 – 56). Supranal concavity long and narrow, setae f2 and ps2 situated at the same transverse level. Setae h1 situated approximately equidistant from levels of setae f2 and setae e2. Setae h3 lanceolate, with strongly narrowed posterior end and round tips, length 72 (72 – 80), greatest width 14 (11 – 13); setae ps2 74 (60 – 72) long, extending beyond to lobar apices; setae ps1 filiform, 10 (12 – 14) long. Distance between bases of dorsal setae: c2 – d2 110 (112 – 122), d2 – e2 86 (90 – 100), e2 – h2 66 (64 – 70), h2 – h3 36 (32 – 38), h2 – h2 24 (24 – 28), h3 – h3 20 (18 – 20), d1 – d2 46 (46 – 50), e1 – e2 36 (30 – 40), h1 – h2 44 (36 – 44), ps1 – h3 5 (4 – 6).




Epimerites I fused into a Y, sternum about ½ of total length of epimerites, posterior end of sternum with transverse extensions connected with medial part of epimerites II. Coxal fields I, II without large sclerotized areas. Coxal fields I – IV closed. Coxal fields IV with large sclerotized areas at bases of trochanters IV. Genital arch 18 (17 – 19) long, 22 (20 – 24) wide, basal sclerite of genital apparatus large, shaped as inverted trapezium. Aedeagus 61 (58 – 62) long, extending almost to posterior margin of the genital shield. Genital papillae well distinct, situated at midlevel of genital arch. Paragenital apodemes fused each other by their medial parts, anterior branches of these apodemes fused with inner margins of epimerites IIIa and posterior branches fused with epimerites IVa. Genital shield, posterior branches of paragenital apodemes and shield-like areas of coxal fields IV fused altogether to form closed sclerotized area surrounding genital apparatus (Fig. 2E). Setae 4b on anterior branches of paragenital apodemes, setae 4a on posterior branches of paragenital apodemes, setae g on inner margin of genital shield. Opisthoventral shields fused together immediately posterior to anal opening, forming entire shield covering posterior third of idiosoma. Anal shields absent, setae ps3 situated near anterior margin of opisthoventral shield. Anal suckers 11 (11 – 12) in diameter, corolla without indentations. Distance between ventral setae: 4b – 4a 52 (52 – 54), 4a – g 42 (36 – 40), g – ps3 54 (52 – 60), ps2 – ps2 36 (34 – 36), ps3 – ps3 32 (30 – 32), ps3 – h3 82 (84 – 88).
Legs I longer and thicker than legs II, tarsus I with dorsal and ventral longitudinal crest, tibia and genu I with dorsal longitudinal crest, femur I with dorso-lateral longitudinal crest, femur II with ventral longitudinal crest, genu IV with two ventral crests (Fig. 2A – D). Setae ra of tarsus I spiculiform, setae d of tarsus II half as long as corresponding seta f. Tarsus IV 22 (24 – 30) long, with small apical claw-like process and blunt-angular ventral extension; seta d hemispherical, situated in proximal part of this segment; setae e absent. Length of solenidia: ω1I 12 (12 – 16), ω1II 15 (16 – 21), φI 80 (81 – 86), φII 46 (44 – 54), φIII 26 (24 – 30), φIV 34 (40 – 41).
FEMALE (range for 5 paratypes) — Length of idiosoma (excluding terminal appendages) 414 – 432, width 116 – 124, length of hysterosoma 296 – 320. Prodorsal shield: antero-lateral extensions wide and connected to epimerites Ia, posterior margin slightly convex, 108 – 116 long, 90 – 112 wide, anterior part with ovate lacunae, posterior part with transverse dashes and lacunae (Fig. 3A). Setae ve represented by alveoli. Setae se separated by 50 – 60. Humeral shields represented by small longitudinal sclerite, setae c2 situated on soft tegument. Setae c3 lanceolate, 20 – 24 x 7 – 8. Anterior and lobar parts of hysteronotal shield separated by narrow transverse band of soft tegument. Hysteronotal shield with anterior margin almost straight, posterior margin with deep and narrow median invagination extending to level of e2 setae, length 240 – 245, width at anterior margin 96 – 114, anterior two thirds of this shield with transverse dashes, posterior one third with few ovate lacunae. Length of lobar region 58 – 70, width 68 – 72, median part of anterior margin slightly convex. Terminal cleft parallel-sided, narrow, with lateral margins almost touching, 50 – 56 long. Supranal concavity absent, setae f2 present, setae h1 on lobar shield, near its anterior margin. Setae h2 spindle-like, 48 – 52 long, 8 – 10 wide. Setae ps1 slightly closer to outer margins of opisthosomal lobes than inner margins, and close to lobar apices. Setae h3 filiform, 10 – 14 long. Distance between bases of dorsal setae: c2 – d2 108 – 114, d2 – e2 86 – 92, e2 – h2 36 – 42, h2 – h3 42 – 50, h2 – h2 42 – 50, h3 – h3 20 – 26, d1 – d2 40 – 50, e1 – e2 31 – 40, h1 – h2 14 – 18, h1 – h1 22 – 26, ps1 – h2 34 – 36.




Epimerites I fused into Y, sternum strongly narrowed posteriorly, about 1/5 from total length of epimerites (Fig. 3B). Epimerites II extending slightly beyond level of anterior margin of epigynum. Epimerites IVa present. Translobar apodemes of opisthosomal lobes wide, not fused to each other anterior to terminal cleft. Copulatory opening located ventraly, immediately posterior to anal opening. Primary spermaduct with punctuated enlargement in proximal part, secondary spermaducts 10 – 13 long (Fig. 4E). Distance between pseudanal setae: ps2 – ps2 31 – 35, ps3 – ps3 17 – 20, ps2 – ps3 20 – 22.
Legs I slightly longer than legs II, femur II with ventral crest; other segments of legs I, II without processes. Setae d of tarsi II – IV much shorter than corresponding setae f. Length of solenidia: ω1I 12 – 16, ω1II 7 – 10, φI 64 – 70, φII 50 – 52, φIII 22 – 26, φIV 6 – 8.
Differential diagnosis — The new species Dolichodectes latilobus n. sp. is most similar to D. furcilobus Mironov, Literák, Hung and Čapek, 2012 described from the White-rumped Shama, Copsychus malabaricus in Vietnam (Mironov et al. 2012). In both sexes of this species, the prodorsal and hysteronotal shields are very close to each other, and the ornamentation of these dorsal shields is similar, with transverse dashes and ovate lacunae; the prodorsal shield has the antero-lateral extensions widely connected to bases of epimerites Ia. In males, the prodorsal shield has the anterior margin with triangular rostral process and bilobate apices of opisthosomal lobes. In females, the shape of lobar region is similar, with median part of anterior margin slightly convex; the supranal concavity is absent; the terminal cleft is parallel-sided, narrow and with lateral margins almost touching. Dolichodectes latilobus n. sp. clearly differs from D. furcilobus by the following features: in males, the opisthosomal lobes are widened in the posterior half and the posterior end of each lobe has a pair of divergent extensions (outer extensions are round, inner extensions are pointed); the hysteronotal shield is strongly narrowed between the levels of setae d2 and e1; the genital shields are fused to each other at the midline of the body, and the opisthoventral shields are fused together forming a single shield that covers the posterior quarter of the body; the aedeagus does not reach the anterior margins of the anal suckers; legs I have dorsal crest-like processes on genu, tibia and tarsus, and setae ra of tarsi I are spiculiform. In females of the new species, the posterior margin of the hysteronotal shield has a deep median invagination extending up to the level of e2 setae and epimerites II extend to the level of the anterior margin of the epigynum. In males of D. furcilobus, the opisthosomal lobes are narrowed in the posterior half and the posterior end of each lobe has a pair of pointed and almost parallel extensions; the hysteronotal shield is moderately narrowed between the pairs of setae d2 and e1; the genital shields are separated and the opisthoventral shields are connected by only a wide transverse bridge; the aedeagus reaches the anterior margins of the anal suckers; legs I are devoid of any dorsal processes, and setae ra of tarsi I are filiform. In females of D. furcilobus, the posterior margin of hysteronotal shield is slightly concave and lacks the median invagination, and epimerites II are much shorter and do not extend to the level of the anterior margin of the epigynum.
ewpage
Genus Trouessartia Canestrini, 1899
The feather mite genus Trouessartia Canestrini, 1899 comprises about 118 species associated predominantly with birds from the order Passeriformes. A revision of this genus including 71 species was performed by Santana (1976); other species were described in the subsequent 40 years by various authors (Mauri and de Alzuet 1968; Černý and Lukoschus 1975; Gaud 1977; Černý 1979; Mironov 1983; Gaud and Atyeo 1986, 1987; Mironov and Kopij 1996, 2000; Mironov and Galloway 2002; OConnor et al. 2005; Carleton and Proctor 2010; Constantinescu et al. 2013, 2016a, 2016b, 2017; Mironov and González-Acuña 2013; Hernandes 2014; Hernandes and Valim 2015, Mironov and Overstreet 2016; Mironov and Palma 2016; Hernandes and OConnor 2017). Hernandes and Valim (2015) suggested that this genus could potentially include over 500 species. A number of undetermined Trouessartia species were reported in the following papers: 22 species from Colombia (Barreto et al. 2012), 15 species from Brazil (Silva et al. 2015) 162 species from Southeast Asia (Atyeo in: McClure and Ratanaworabhan 1973). To date, two Trouessartia species have been described from birds of the genus Copsychus: Trouessartia microfolia Gaud, 1952 from Copsychus albospecularis (Eydoux and Gervais) and T. sechellarum Mironov and Palma, 2016 from C. sechellarum Newton (Gaud 1952, Mironov and Palma, 2016). Three presumably new Trouessartia species were reported by Atyeo in the aforementioned paper from Copsychus saularis (Linnaeus), C. luzoniensis (Kittlitz) and C. malabaricus (Scopoli), but they have never been described.
ZOOBANK: DDA22F74-100B-4863-B579-69860FA8B68A ![]()
Type material — Holotype male and 19 paratypes (9 males and 10 females) from Copsychus saularis (Linnaeus) (Passeriformes: Muscicapidae), INDONESIA, Kalimantan Island, Lhok Tuan, east of the Kutai National Park, 18 May 1991, bird inventory number 15719-47, no other data.
Type deposition — Holotype male (ANA 822), 7 male (ANA 823 – 829) and 8 female (ANA 830 – 837) paratypes in MGAB collection, 2 male and 2 female paratypes in DZUnesp-RC collection.
Etymology — The species epithet is a noun in apposition given after the species name of the type host.
Description — MALE (holotype, range for 4 paratypes) — Length of idiosoma from anterior end to lobar apices (bases of setae h3) 392 (376 – 385), greatest width at level of humeral shields 166 (164 – 178). Length of hysterosoma from sejugal furrow to lobar apices 292 (276 – 292). Prodorsal shield: length along midline 128 (120 – 124), greatest width in posterior part 130 (130 – 132), lateral margins not fused with scapular shields, antero-lateral extensions short and widely rounded, not extending to body margins between bases of legs I, II, posterior margin slightly convex, surface without ornamentation (Fig. 5). Internal scapular setae si lanceolate, 24 (20 – 24) long, separated by 44 (38 – 44); external scapular setae se situated on prodorsal shield, separated by 80 (80 – 82). Vertical setae ve represented by alveoli. Humeral shields with setae c2 lanceolate, 44 (40 – 46) long. Setae c3 narrowly lanceolate, with acute apex, 24 (24 – 25) long. Dorsal hysterosoma with prohysteronotal shield and lobar shield connected, delimited from each other by lateral incisions immediately posterior to setae e2 and small desclerotised median area. Prohysteronotal shield length 160 (146 – 154), width at anterior margin 130 (122 – 130), lateral margins deeply incised at level of trochanters III, bottom of these incisions with C-shaped dark sclerotisation, dorsal hysterosomal apertures (DHA) absent, surface without ornamentation. Dorsal setae d1, d2, e2 present, minute. Lobar shield elongated and narrow, length of lobar shield excluding lamellae 100 (94 – 100). Apical parts of opisthosomal lobes approximate, separated by narrow terminal cleft; length of this cleft from anterior end to apices of lamellae 54 (50 – 56), width in anterior part 6 (6 – 8). Lamellae semi-ovate, distinctly attenuate apically, their inner margins almost touching at level of their midlength, outer margins convex, length from bases of setae h3 to lamellar apices 38 (32 – 40). Setae h1 anterior to setae h2. Distance between dorsal setae: c2 – d2 72 (72 – 80), d2 – e2 84 (80 – 82), e2 – h2 80 (72 – 76), h2 – h3 20 (18 – 22), h2 – h2 30 (30 – 32), h3 – h3 28 (28 – 32), d1 – d2 36 (36 – 40), e1 – e2 40 (42 – 47).


Epimerites I free. Rudimentary sclerites rEpIIa small, roughly ovoid. Genital apparatus situated between levels of trochanters III and IV, length 50 (48 – 52), greatest width 20 (18 – 21) (Fig. 6). Epiandrum present, setae g long and filiform, contiguous at bases, postgenital plaque absent. Adanal apodemes heavily sclerotised, with two narrow lateral membranes and without apophyses. Translobar apodeme present. Adanal shields small, teardrop-shaped, bearing setae ps3. Anal suckers 15 (14 – 16) in diameter. Anterior ends of epimerites IIIa with apices extending beyond level of setae 4b; epimerites IVa short. Setae 4b and 3a situated at the same transverse level, setae g situated posterior to level of setae 4a. Distance between ventral setae: 4b – 3a 31 (30 – 34), 4b – g 70 (64 – 72), g – ps3 38 (38 – 42), ps3 – h3 106 (100 – 105).

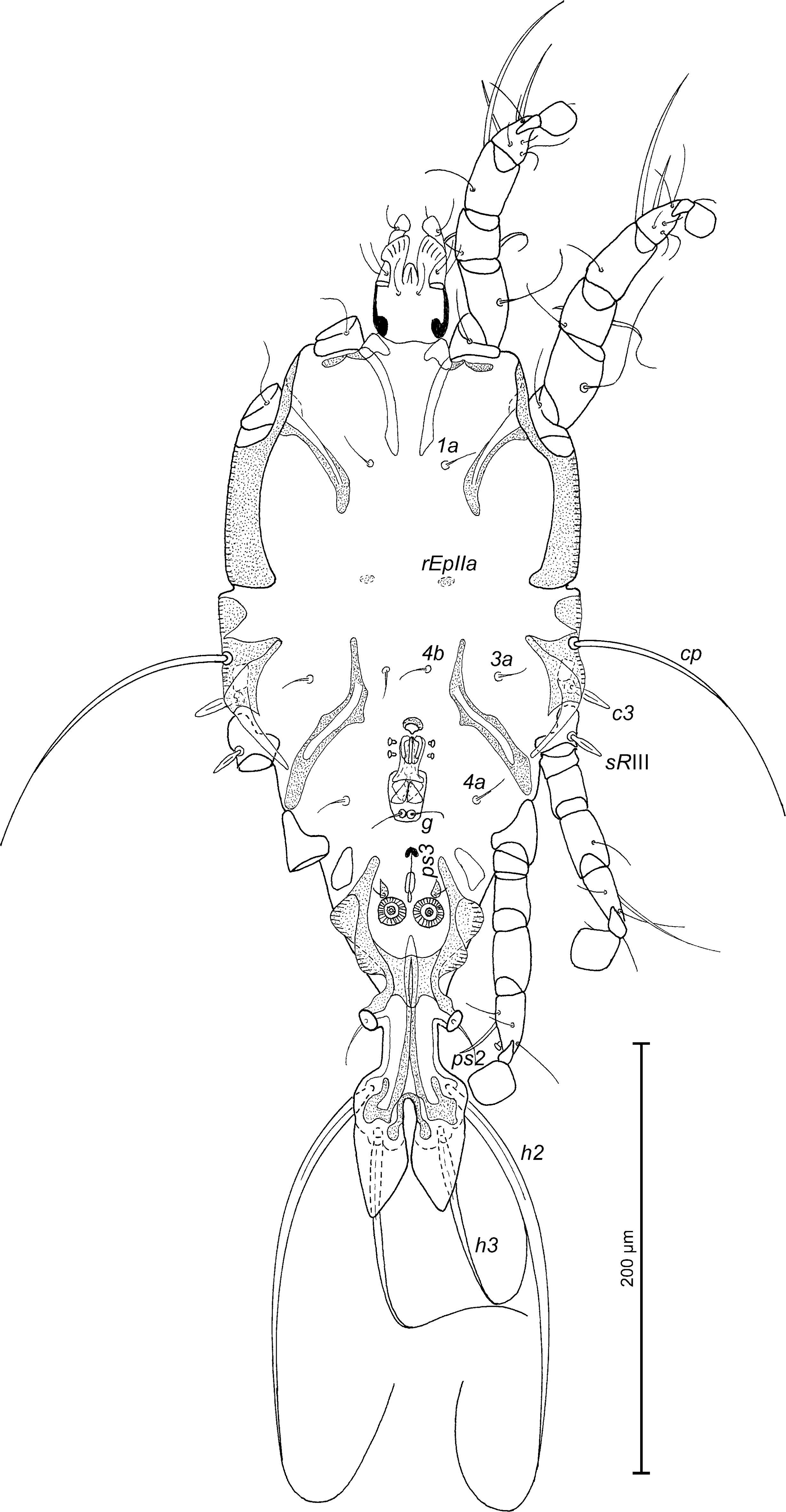
Setae sR of trochanters III narrowly lanceolate, with pointed apices 17 (17 – 20) long, setae cG and mG of genua I, II filiform. Tarsus IV 36 (30 – 34) long; seta d barrel-shaped, with discoid cap; seta e hemispheroid, without cap, situated subapically (Fig. 7D). Length of solenidia: σ1I 34 (30 – 34), σII 14 (14 – 18), σIII 18 (18 – 22), φIV 40 (35 – 40).


FEMALE (range for 5 paratypes) — Length of idiosoma from anterior end to apices of lamellar lobar processes 404 – 432, greatest width 162 – 184. Length of hysterosoma from sejugal furrow to apices of lamellar lobar processes 288 – 296. Prodorsal shield shaped as in male, 120 – 130 in length, 128 – 138 in width, surface without ornamentation. Setae si lanceolate, 20 – 24 long, separated by 40 – 44; external scapular setae se situated on prodorsal shield, separated by 80 – 86. Humeral shields with setae c2 lanceolate, 40 – 50 long. Setae c3 narrowly lanceolate, with pointed apices 25 – 26 in length. Hysteronotal shield length from anterior margin to bases of setae h3 248 – 256, width at anterior margin 114 – 120, lateral margins incised at level of trochanters III, bottom of these incisions with C-shaped dark sclerotisation, DHA absent, median area from level of setae cp to that of setae h1 with numerous ovate lacunae (Figure 8). Dorsal setae d1, d2, e2 present. Setae f2 absent. Setae h1 short spiculiform, surrounded by small ovoid area of unsclerotised tegument, 5 – 6 long, situated antero-mesal to bases of setae h2, 18 – 21 from corresponding lateral margins of hysteronotal shield. Setae ps1 positioned dorsally on opisthosomal lobes, equidistant from outer and inner margins of lobe, close to bases of setae h3. Distance from bases of setae h3 to membranous apices of lobes 24 – 34. Supranal concavity open posteriorly into terminal cleft. Length of terminal cleft together with supranal concavity 98 – 110, width of cleft at level of setae h3 20 – 24. Interlobar membrane occupying anterior 1/3 of terminal cleft, distance from free margin of interlobar membrane to membranous lobar apices 58 – 66. Spermatheca with primary spermaduct terminating ventrally on interlobar membrane, with long collar, bearing fringe of minute spines on posterior margin, length of secondary spermaducts 20 – 26 (Fig. 10E). Distance between dorsal setae: c2 – d2 68 – 73, d2 – e2 44 – 50, e2 – h2 94 – 100, h2 – h3 44 – 50, h2 – h2 54 – 60, h3 – h3 32 – 40, d1 – d2 32 – 40, e1 – e2 46 – 50, h1 – h2 14 – 16, h1 – h1 34 – 40, ps1 – h3 9 – 10.




Epimerites I free. Epigynum 42 – 48 in length, 70 – 80 in width (Figure 9). Epimerites IVa present. Anal opening with two small ovate adanal sclerites situated at level of its anterior end. Setae sR of trochanters III narrowly lanceolate, with pointed apices, 18 – 20 long, setae cG and mG of genua I, II filiform. Legs IV with ambulacral disc extending to midlevel between setae h2 and h3.


Differential diagnosis — Trouessartia saularis n. sp. morphologically is most similar to T. microfolia Gaud, 1952 described from Copsychus albospecularis from Madagascar (Gaud 1952). In both sexes of these species, the antero-lateral extensions of the prodorsal shield are not connected with bases of epimerites Ia, the lateral margins of the hysteronotal shields are without dorsal hysteronotal apertures and setae c2 are lanceolate and almost two times longer than setae c3 and sRIII. Males of both species have the hysteronotal shield entire and its prohysteronotal and lobar parts delimited from each other by small lateral incisions at the level of setae e2. In females, setae h1 are spiculiform and the collar of the spermathecal head has a fringe of minute spines. Trouessartia saularis differs from T. microfolia by the following features. In both sexes of T. saularis, the prodorsal shield lacks ornamentation; in males, the terminal lamellae are slightly attenuate apically, the inner ends of epimerites IVa do not extend to the level of setae g, the postgenital plaque is absent; in females, the terminal cleft width is smaller than the opisthosomal lobe width and the collar of the spermatheca covers 1/2 the length of the secondary spermaducts. In both sexes of T. microfolia, the prodorsal shield has an ornamentation of a faint network; in males, the terminal lamellae are bluntly rounded, the inner ends of epimerites IVa extend to the level of setae g, the postgenital plaque is present; in females, the terminal cleft is larger than the opisthosomal lobe width, and the collar of the spermatheca covers about 1/6 the length of the secondary spermaduct.
We are very grateful to the research team who collected birds specimens from some islands of the Indonesian archipelago in the expedition organized by “Grigore Antipa” National Museum of Natural History in 1991: Dr. Dumitru Murariu, the chief of expedition, Dr. Modest Guțu, Dr. Alexandru Marinescu and Dr. Corneliu Pârvu.
Atyeo W.T. 1992. The pterolichoid feather mites (Acarina, Astigmata) of the Megapodiidae (Aves, Galliformes). Zool. Scr., 21: 265-305. doi:10.1111/j.1463-6409.1992.tb00331.x ![]()
Atyeo W.T., Braasch L. 1966. The feather mite genus Proctophyllodes (Sarcoptiformes: Proctophyllodidae). Bull. Univ. Nebraska St. Mus., 5: 1-354.
Atyeo W.T., Gaud J. 1991. Lorilichus, a new genus of feather mites (Acarina, Pterolichidae) from lories and lorikeets (Aves, Psittacidae). Syst. Parasitol., 18: 139-146. doi:10.1007/BF00017667 ![]()
Atyeo W.T., Gaud J. 1992. The identity of Pterolichus obtusus Robin, 1877 with descriptions of new genera and species of feather mites (Acarina, Pterolichidae) from the Galliformes (Aves). Acarologia, 33: 193-206.
Atyeo W.T., Pérez T.M. 1982. New taxa of Psittacine feather mites (Acarina, Pterolichidae). Jour. Parasit., 68: 1158-1161. doi:10.2307/3281112 ![]()
Barreto M., Burbano M.E., Proctor H.C., Mironov S.V., Wauthy G. 2012. Feather mites (Acariformes: Psoroptidia) from Colombia: Preliminary list with new records. Zootaxa, 3516: 1-68.
BirdLife International 2017. Copsychus saularis. The IUCN Red List of Threatened Species 2017 [Internet]. [15 July 2017]. doi:10.2305/IUCN.UK.2017-1.RLTS.T103893432A111178145.en ![]()
Bonnet A. 1924. Révision des genres Megninia, Mesalges et genres voisins de la sous-famille des sarcoptides plumicoles (2e Partie). B. Soc. Zool. Fr., 49: 190-218.
Carleton R.E., Proctor H.C. 2010. Feather mites associated with Eastern Bluebirds (Sialia sialis L.) in Georgia, including the description of a new species of Trouessartia (Analgoidea: Trouessartiidae). Southeast. Nat., 9: 605-623. doi:10.1656/058.009.0317 ![]()
Černý V. 1979. Feather mites (Sarcoptiformes: Analgoidea) of some warblers from Czechoslovakia. Fol. Parasitol., 26: 81-84.
Černý V., Lukoschus F.S. 1975. Parasitic mites of Surinam XXXIII. Feather mites (Analgoidea). Stud. Fau. Surin. Guyan., 58: 184-203. doi:10.1007/978-94-017-7106-1_3 ![]()
Clements J.F., Schulenberg T.S., Iliff M.J., Roberson D., Fredericks T.A., Sullivan B.L., Wood C.L.. The eBird/Clements checklist of birds of the world: v2016 [Internet]. [15 July 2017]. Available from: http://www.birds.cornell.edu/clementschecklist/download/ ![]()
Constantinescu I.C., Chișamera G., Pocora V., Stanciu C., Adam C. 2013. Two new species of feather mites (Acarina: Analgoidea) from the Moustached Warbler, Acrocephalus melanopogon (Passeriformes, Acrocephalidae) in Romania. Zootaxa, 3709: 267-276. doi:10.11646/zootaxa.3709.3.5 ![]()
Constantinescu I.C., Cobzaru I., Mukhim D.K.B., Adam C. 2016a. Two new species of the genus Trouessartia (Acari, Trouessartiidae) from laughingthrushes (Passeriformes, Leiothrichidae). ZooKeys, 571: 59-79. doi:10.3897/zookeys.571.7724 ![]()
Constantinescu I.C., Cobzaru I., Mukhim D.K.B., Adam C. 2016b. Two new species of the feather mite genus Trouessartia (Acari: Trouessartiidae) from Asia. Zootaxa, 4137(3): 357-374. doi:10.11646/zootaxa.4137.3.4 ![]()
Constantinescu I.C., Cobzaru I., Geamana N.A., Mukhim D.K.B., Adam C. 2017. Two new species of feather mites (Acarina: Psoroptidia) from the blue-throated blue flycatcher, Cyornis rubeculoides (Passeriformes: Muscicapidae). J. Nat. Hist., 51: 277-297. doi:10.1080/00222933.2017.1280194 ![]()
Dabert J., Ehrnsberger R. 1996. Four new species of the feather mite genus Sokoloviana Dubinin, 1951 (Pterolichoidea; Ptiloxenidae) from waders (Charadriiformes; Charadrii). Osnabr. Natur. Mitteil., 22: 135-163.
Dabert J., Labrzycka A. 2009. Szeptyckiana, a new genus of the feather mite family Pterolichidae (Acari, Astigmata, Pterolichoidea) from the Black-thighed Falconet Microhierax fringillarius (Falconiformes, Falconidae). Acta Zool. cracov., 52: 61-72.
Dabert J., Mironov S.V., Ehrnsberger R. 2002. A revised diagnosis of the feather mite genus Magimelia Gaud, 1961 (Pterolichoidea: Pterolichidae: Magimeliinae) and the description of three new species. Syst. Parasitol., 53: 60-79. doi:10.1023/A:1019957822743 ![]()
Doña J., Proctor H., Mironov M., Serrano D., Jovani R. 2016. Global associations between birds and vane-dwelling feather mites. Ecology, 97: 3242. doi:10.1002/ecy.1528 ![]()
Favette J., Trouessart E.L. 1904. Monographie du genre Protolichus (Trs) et revision des Sarcoptides plumicoles (Analgesinae) qui vivent sur les perroquetes. Mém. Soc. zool. Fr., 17: 120-166.
Gaud J. 1952. Sarcoptides plumicoles des oiseaux de Madagascar. Mém. L'Inst. Scien. Madagascar, 7: 81-107.
Gaud J. 1977. La faune terrestre de l'Île de Sainte Hélène. 4.3. Acariens Sarcoptiformes Plumicoles parasites d'oiseaux. Ann. Mus. roy. Afr. centr., 220: 260-269.
Gaud J., Atyeo W.T. 1986. Les Trouessartia (Analgoidea, Trouessartiidae) parasites des hirondelles de l'Ancien Monde. I. Le Groupe appendiculata. Acarologia, 27: 263-274.
Gaud J., Atyeo W.T. 1987. Les Trouessartia (Analgoidea, Trouessartiidae) parasites des hirondelles de l'Ancien Monde. I. Le Groupe minutipes. Acarologia, 28: 367-379.
Gaud J., Atyeo W.T. 1996. Feather mites of the world (Acarina, Astigmata): The supraspecific taxa. Ann. Mus. roy. Afr. centr., 277:1-193 (Part 1, text), 1-436 (Part 2, illustrations).
Gaud J., Atyeo W.T., Barre N. 1985. Mites of the genus Megninia (Analgidae) parasitic upon Gallus gallus. Acarologia, 26: 171-182.
Gaud J., Mouchet J. 1957. Acariens plumicoles (Analgesoidea) des oiseaux du Cameroun. I. Proctophyllodidae. Ann. Parasitol. Hum. Comp., 32: 491-546.
Griffiths D.A., Atyeo W.T., Norton R.A., Lynch C.A. 1990. The idiosomal chaetotaxy of astigmatid mites. J. Zool., 220: 1-32. doi:10.1111/j.1469-7998.1990.tb04291.x ![]()
Hernandes F.A. 2014. Five new species of the feather mite genus Trouessartia Canestrini from South America (Acari: Trouessartiidae). Zootaxa, 3856: 50-72. doi:10.11646/zootaxa.3856.1.2 ![]()
Hernandes F.A., Valim M.P. 2006. Two new species of the feather mite subfamily Pterodectinae (Acari: Astigmata: Proctophyllodidae) from Brazil. Zootaxa, 1235: 49-61.
Hernandes F.A., Valim M.P. 2015. A new species of the genus Trouessartia Canestrini (Acari: Trouessartiidae) from Neotropical passerines (Aves: Tyrannidae). Int. J. Acarol., 41: 382-388. doi:10.1080/01647954.2015.1046921 ![]()
Hernandes F.A., OConnor B.M. 2017. Out of Africa: the mite community (Arachnida: Acariformes) of the common waxbill, Estrilda astrild (Linnaeus, 1978) (Passeriformes: Estrildidae) in Brazil. Parasite. Vector., 10: 299. doi:10.1186/s13071-017-2230-5 ![]()
Lepage D.. Avibase – Bird Checklists of the World [Internet]. [15 Julay 2017]. Available from: https://avibase.bsc-eoc.org/checklist.jsp?lang=EN ![]()
MacKinnon J., Phillipps K. 1993. A Field Guide to the Birds of Borneo, Sumatra, Java and Bali. The Greater Sunda Islans. Eds Oxford University Press. pp. 491.
Mauri R., De Alzuet A.B. 1968. Una nueva espécie de Trouessartia Canestrini, 1899 (Acarina: Proctophyllodidae). Rev. Mus. La Plata, 85: 169-172.
McClure H.E., Ratanaworabhan N. 1973. Some ectoparasites of the birds of Asia, Applied Scientific Research. Jintana Printing Ldt, Bangkok. pp. 219.
Mironov S.V. 1983. Feather mites of the genus Trouessartia of the USSR fauna and descriptions of new species (Analgoidea). Parazitologiya, 17: 361-369.
Mironov S.V. 2006. Feather mites of the genus Montesauria Oudemans (Astigmata: Proctophyllodidae) associated with starlings (Passeriformes: Sturnidae) in the Indo-Malayan region, with notes on the systematics of the genus. Acarina, 14: 21-40.
Mironov S.V., Dabert J. 2010. Systematic revision of the feather mite genus Protolichus Trouessart, 1884 (Astigmata, Pterolichidae). Zootaxa, 2526: 1-36. doi:10.11646/zootaxa.2526.1.1 ![]()
Mironov S.V., Fain A. 2003. New species of the feather mite subfamily Pterodectinae (Astigmata: Proctophyllodidae) from African passerines (Aves: Passeriformes). Bull. Soc. R. Belge Entomol., 139: 75-91.
Mironov S.V., González-Acuña D.A. 2013. A new feather mite species of the genus Trouessartia Canestrini, 1899 (Acariformes: Trouessartiidae) from the White-Crested Elaenia Elaenia albiceps (Orbigney et Lafresnaye) (Passeriformes: Tyrannidae) in Chile. Acarina, 21: 123-132.
Mironov S.V., Kopij G. 1996. New feather mite species (Acarina: Analgoidea) from some starlings (Passeriformes: Sturnidae) of South Africa. J. Afr. Zool., 110: 257-269.
Mironov S.V., Kopij G. 2000. New feather mites species of the genus Trouessartia (Acari: Analgoidea: Trouessartiidae) from South African passerines (Aves: Passeriformes). Mitt. hamb. zool. Mus. Instit., 97: 99-115.
Mironov S.V., Overstreet R.M. 2016. A new feather mite species of the genus Trouessartia Canestrini (Acariformes: Trouessartiidae) from the northern rough-winged swallow Stelgidopteryx serripennis (Passeriformes: Hirundinidae) in Pennsylvania. Acarina, 24: 3-9.
Mironov S.V., Palma R.L. 2016. A new feather mite of the genus Trouessartia Canestrini 1988 (Acariformes: Trouessartiidae) from the Seychelles magpie-robin, Copsychus sechellarum (Passeriformes: Muscicapidae). Acta Parasitol., 61: 629-635. doi:10.1515/ap-2016-0084 ![]()
Mironov S.V., Perez T.M. 2003. A review of feather mites of the Rhytidelasma generic group (Pterolichoidea Pterolichidae), specific parasites of parrots (Aves Psittaciformes). Bull. Inst. R. Sci. Nat. Belg. Entomol., 73: 135-176.
Mironov S.V., Dabert J., Ehrnsberger R. 2002. On systematics of the feather mite genus Triphyllochaeta Dubinin, 1956 (Astigmata: Pterolichidae). Mitt. hamb. zool. Mus. Instit.,14: 27-37.
Mironov S.V., Dabert J., Ehrnsberger R. 2003. A review of feather mites of the Psittophagus generic group (Astigmata, Pterolichidae) with descriptions of new taxa from parrots (Aves, Psittaciformes) of the Old World. Acta Parasitol., 48: 280-293.
Mironov S.V., Doña J., Jovani R. 2015. A new feather mite of the genus Dolichodectes (Astigmata: Proctophyllodidae) from Hippolais polyglotta (Passeriformes: Acrocephalidae) in Spain. Folia Parasitol., 62: 032. doi:10.14411/fp.2015.032 ![]()
Mironov S.V., Ehrnsberger R., Dabert J. 2014. New species of the feather mite genus Protolichus Trouessart, 1884 (Astigmata, Pterolichidae) from lories and lorikeets (Aves: Psittaciformes). Zootaxa, 3774: 131-151. doi:10.11646/zootaxa.3774.2.2 ![]()
Mironov S.V., Literák I., Čapek M., Koubek P. 2010. New species of the feather mite subfamily Pterodectinae (Astigmata, Proctophyllodidae) from passerines in Senegal. Acta Parasitol., 55: 399-413. doi:10.2478/s11686-010-0051-1 ![]()
Mironov S.V., Literák I., Hung M.H., Koubek P. 2012. New feather mites of the subfamily Pterodectinae (Acari: Proctophyllodidae) from passerines and woodpeckers (Aves: Passeriformes and Piciformes) in Vietnam. Zootaxa, 3440: 1-49.
Norton A.R. 1998. Morphological evidence for the evolutionary origin of Astigmata (Acari: Acariformes). Exp. Appl. Acarol., 22: 559-594. doi:10.1023/A:1006135509248 ![]()
OConnor B.M., Foufopoulos J., Lipton D., Lindström K. 2005. Mites associated with the small ground finch, Geospiza fuliginosa (Passeriformes: Emberizidae), from the Galápagos Islands. J. Parasitol., 91: 1304-1313. doi:10.1645/GE-581R.1 ![]()
Peterson P.C., Atyeo W.T. 1977. The feather mite family Alloptidae Gaud IV – A new genus of the Alloptinae (Acarina, Analgoidea). Acarologia, 19: 318-330.
Pérez T.M., Atyeo W.T. 1981. A new feather mite (Acarina: Pterolichidae) from a fruit pigeon (Aves: Columbidae). J. Parasitol., 67: 442-444. doi:10.2307/3280570 ![]()
Santana F.J. 1976. A review of the genus Trouessartia (Analgoidea: Alloptidae). J. Med. Entomol. Supp., 1: 1-128. doi:10.1093/jmedent/13.Suppl1.1 ![]()
Silva H.M., Hernandes F.A., Pichorim M. 2015. Feather mites (Acari, Astigmata) associated with birds in an Atlantic Forest fragment in Northeastern Brazil. Braz. J. Biol., 75: 726-735. doi:10.1590/1519-6984.23313 ![]()
Trouessart E. L. 1884. Les Sarcoptides plumicoles. Journal of Micrographie, 8: 63-70.
Trouessart E. L. 1885. Note sur le classification des Analgésines et diagnoses d'espèces et de genres nouveaux. Bull. Soc. Etud. Scient. Angers, 14: 46-89.



2017-08-01
Date accepted:
2017-10-10
Date published:
2018-02-22
Edited by:
Tixier, Marie-Stéphane

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Constantinescu, Ioana Cristina; Chişamera, Gabriel Bogdan; Petrescu, Angela and Costică, Adam
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)