



Molecular phylogenetic relationships of halacarid mites suggest the reevaluation of traditional subfamily classification
Meguro, Hiroko1 ; Iwasa, Masahiro A.2 ; Goka, Koichi3 and Abé, Hiroshi4
1Biological Laboratory, Graduate School of Bioresource Sciences, Nihon University, Kameino 1866, Fujisawa, Kanagawa Pref., Japan.
2✉ Laboratory of Wildlife Science, College of Bioresource Sciences, Nihon University, Kameino 1866, Fujisawa, Kanagawa Pref., Japan.
3National Institute for Environmental Studies, 16–2 Onogawa, Tsukuba, Ibaraki 305–0053, Japan.
4Biological Laboratory, Graduate School of Bioresource Sciences, Nihon University, Kameino 1866, Fujisawa, Kanagawa Pref., Japan.
2017 - Volume: 57 Issue: 3 pages: 633-641
https://doi.org/10.24349/acarologia/20174182Keywords
Abstract
The classification of mites has relayed mainly on morphological criteria, particularly exoskeletal characteristics (Krantz 1978; Walter and Proctor 1999). The classification was constructed traditionally, based only on the morphology-based relationships of mites. However, it is unclear whether such morphological characters reflect real evolutionary processes, partly because the traditional method inevitably includes arbitrary choices of the morphological characters used. In addition, it is sometimes difficult to find structural homology to be used for morphological comparisons between organisms. Recent molecular studies on mite taxonomy have raised conflicts between morphology- and molecular-based relationships challenging mite classifications (Cruickshank 2002; Pepato et al. 2010; Li et al. 2014; Matsuda et al. 2014), but have also highlighted their value in evaluating mite classifications on the basis of reliable phylogenetic relationships.
The vast number of Prostigmata mites, halacarid mites (Halacaridae) are aquatic taxa which mostly inhabit coastal sea areas (Abé 1990). Since the 1800s the classification of halacarid mites was performed by using morphological criteria (e.g., Bartsch 2015). So far approximately 1,300 species in 64 genera have been described worldwide and there still remain a large number of undescribed species. On the other hand, studies on the phylogenetic relationships among halacarid taxa are scarce. Phylogenetic studies based on morphological characters have been carried out only on Rhombognathinae mites which consist on of four genera: Rhombognathus, Rhombognathides, Isobactrus and Metarhombognathus (Abé 1998, 2001a, b; Abé and Bartsch 2007; Bartsch 2003, 2008, 2010). Only few studies reported on their classification (Abé 2001b; Bartsch 2015). Thereafter, halacarid mites have been used as partial information in the molecular phylogenetic analyses of acariform mites (Otto and Wilson 2001; Pepato et al. 2010; Pepato and Klimov 2015; Dabert et al. 2016).
In this study, we analyzed nuclear ribosomal gene (18S and 28S) sequences of several halacarid taxa: Copidognathinae (Copidognathus), Halacarinae (Halacarus, Agauopsis and Thalassarachna), Limnohalacarinae (Limnohalacarus), Halixodinae (Bradyagaue and Agaue) and Rhombognathinae (Rhombognathus, Metarhonbognathus and Rhombognathides) and re-evaluated the typological classification of the halacarid taxa by morphological criteria based on the current molecular findings.
We collected individual halacarid mites (Table 1). After sufficient starvation (over six months), each mite was stored in 70% EtOH at room temperature. To remove the EtOH of specimen storage, each mite was put into a microtube and air-dried at room temperature for a few hours. After removal of the EtOH, 15 µl of nuclei lysis solution (Wizard Genomic DNA Purification Kit, Promega) and 2 µl of 1% proteinase K solution (pK, Merck) were added into the microtube (Goka et al. 2001), and it was incubated at 50 °C for 120 min to soften the mite body. Previously, the hinge of a 1.5 ml microtube was cut to remove the cap. After heat treatment, the mite was placed inside the cap with all of the solution. Subsequently, in order to take out the inner contents of the mite’s body, we removed the gnathosoma from the body using needles and gently squeezed out inner contents from the body exoskeleton in the lysis – pK solution on the cap. The cap loaded with the mite body exoskeleton (idiosoma with legs), the gnathosoma, and the inner body contents was carefully placed back in the microtube with the solution, and the microtube, with the cap attached, was spun down at 12,000 rpm for a few seconds. Fresh 25 µl nuclei lysis solution and 5 µl of 1% pK solution were added into the microtube that contained the idiosoma with legs, the gnathosoma, and the inner contents with the primary solution, and all of the contents were incubated again at 50 °C for 120 min and then at 95 °C for 20 min. After incubation, the exoskeleton and the gnathosoma were picked up for specimen preparation. Finally, after incubation, the extract was diluted to 10% of its original concentration using TE buffer (0.001 M EDTA, 0.01 M Tris-HCl [pH 8.0]) following the method of Goka et al. (2001, 2009) and used as the source of the DNA template for PCR amplification.


Fragments of the nuclear 18S and 28S ribosomal RNA gene (rDNA) regions were amplified by the PCR method using the following primer sets: HAL 18S$_$2F (5’-GTG TCT GCC TTA TCA ACT TTC GAT GG-3’) for the forward direction and HAL 18S$_$2R (5’-GCC CCC GTC TGT CCC TCT TAA TC-3’) for the reverse direction for 18S rDNA, and 28S$_$V (28ee in Hillis and Dixon 1991) for the forward direction and 28S$_$VI (reverse of 28v in Hillis and Dixon 1991) for the reverse direction for 28S rDNA. PCR amplifications were conducted in accordance with the procedure of Goka et al. (2001), with 2 μL of each template DNA in a total reaction volume of 50 μL. The PCR reaction mix contained 0.2 mM of each dNTP, 2 mM MgCl2, 1.25 units of Taq DNA polymerase (AmpliTaq Gold), and 0.5 mM of each primer. All PCR reagents were purchased from Perkin Elmer Applied Biosystems. Conditions for the amplification were an initial denaturation for 9 min at 95 °C; 40 cycles of 30 s at 94 °C, 30 s at 44-54 °C, and 2 min at 72 °C; and a final extension for 7 min at 72 °C. After the removal of the primers with PEG (20% polyethylene glycol + 2.5M NaCl) precipitation, both strands of the PCR products were sequenced directly and automatically using the BigDye Terminator Cycle Sequencing Kit ver. 3.1 (ABI) and the 3730 Genetic Analyzer (ABI). All of the current sequences were deposited to the DNA databases with accession numbers LC171603–LC171640.
In the current analysis, we used newly produced sequences together with additional data from DNA databases: Acarothrix for 18S (KP276481) and 28S (KP276405), Halacarus for 18S (HM070350), Thalassarachna for 18S (AY692342), Rhombognathus for 18S (HM070351) and Rhombognathides for 18S (AY692341). All sequences were aligned automatically using the Clustal X program (Higgins et al. 1996) with outgroups: Allothrombium (KP276493, Pepato and Klimov 2015), Eupodidae sp. (GQ864273, Dabert et al. 2010) and Rhagidia sp. (GQ864272, Dabert et al. 2010) for 18S rDNA and Allothrombium (KP276417, Pepato and Klimov 2015), Eupodidae sp. (KM100954, Dabert et al. 2016) and Rhagidia sp. (KM100953, Dabert et al. 2016) for 28S rDNA. The sequences newly obtained were deposited in the DNA databases with accession numbers indicated in Table 1. Primarily, we estimated appropriate models for 18S, 28S and combined 18S + 28S sequences using PartitionFinder ver 1.1.1 (Lanfear et al. 2012). On the basis of the models chosen (Table 2), we constructed maximum likelihood trees for each of the three gene regions considering transitions and transversions at all positions with 1,000 replicate bootstrap analysis, using MEGA ver. 7 (Kumar et al. 2016).


PCR amplifications of the present halacarid mite samples are presented in Table 1. The amplifications of the 18S rDNA and 28S rDNA regions and the aligned sequences of 517-520 and 526-549 base pairs (bp) lengths, respectively, were compared by a homology search as BLAST of DDBJ. All the obtained sequences matched to the suborder Prostigmata by using BLAST excluding any possible experimental contamination. Phylogenetic trees for the current mite samples were subsequently constructed using these two gene sequences.
For the 18S rDNA sequences, the GTR+G+I model was chosen by lnL = –2258.46 and AICc 4635.45, which revealed the most appropriate ML tree (lnL = –2181.42, +G = 0.4947, +I= 52.81%) (Figure 1). The 18S rDNA tree showed that each of the subfamilies Halixodinae and Halacarinae consisted of an independent cluster. However, Rhombognathinae did not form a monophyletic group. For example, the genus Rhombognathus in Rhombognathinae consisted of a monophyletic cluster and was apparently different from the lineage Rhombognathides and Metarhombognathus in Rhombognathinae with the largest ML distance: 0.121. In addition, the subfamily Halacarinae was divided into three generic clusters, Thalassarachna, Agauopsis and Halacarus (Figure 1) and the ML distances were Thalassarachna vs. Agauopsis: 0.037, Thalassarachna vs. Halacarus: 0.107 and Agauopsis vs. Halacarus: 0.107. On the other hand, the subfamilies Halixodinae and Copidognathinae consisted of monophyletic clusters including two or three genera with ML distances 0.000 – 0.030. In Halixodinae, the clustering of Agaue and Bradyagaue was supported by the highest bootstrap value.
The model chosen for the 28S rDNA sequences was GTR+G by lnL = –3236.47 and AICc 6565.36, which provided the most appropriate ML tree (lnL = –3056.08, +G = 0.2834). The 28S tree revealed different topology compared to the 18S rDNA tree (Figure 1). In the 18S rDNA tree, the subfamily Halacarinae was divided into three clusters, while the 28S rDNA tree indicated the Halacarinae as monophyletic. On the other hand, the subfamily Rhombognathinae was divided into different clusters, as in the 18S rDNA tree, irrespective of the lower bootstrap values (Figure 1). In addition, only a Rhombognathus sp. (OSH0903$_$5) showing similarity to the Copidognathinae cluster showed a large distance (ranging from vs. KE0812$_$6: 0.357 to vs. KM100954: 0.583) from the other taxa in the 28S rDNA tree, and showed smaller distance (ranging from vs. OSH0903$_$37: 0.000 to vs. KM100954: 0.179) in the 18S rDNA tree. The subfamilies Halixodinae and Copidognathinae also appeared each as monophyletic clusters. In Halixodinae, the clustering of Agaue and Bradyagaue was supported by the highest bootstrap value as in the 18S rDNA tree.
For the combined 18S and 28S rDNA sequences (18S + 28S), the model estimation chose the GTR+G+I model by lnL = –5176.94 and AICc 10448.09. The tree (lnL = –4930.22, +G = 0.4525, +I: 38.32%) by the combined 18S and 28S rDNAs strongly indicated the separation of the Holixadinae–Halacarinae–Rhombognathinae (Metarhombognathus) cluster to the Copidognathinae –Rhombognathinae (Rhombognathus) cluster with high bootstrap value (87%) (Figure 2). Thus, Rhombognathinae was divided into two different lineages while the other subfamilies appeared as monophyletic.
The phylogenetic trees based on sequences of the nuclear ribosomal (18S and 28S) genes revealed some incongruence between tree clustering and the current classification of the subfamilies Halacarinae and Rhombognathinae (Figure 1). By contrast, Halixodinae and Copidognathinae appeared phylogenetically robust and in line with the classifications based on morphological criteria (Abé 1998; Bartsch 1993, 2006, 2008, 2010).
In the 18S tree, the three genera of Halacarinae were divided into three different clusters. In particular, Halacarus was apparently differentiated from the other halacarine genera, Agauopsis and Thalassarachna (Figure 1). In addition, the affinity between Agauopsis and Thalassarachna was recognized in the 28S tree and the 18S + 28S tree (Figures 1 and 2). These results suggest that the taxonomic assignment of Halacarus in the higher classification should be reevaluated. Halacarus is the type genus of the subfamily Halacarinae. The diagnostic characters of the subfamily were sometimes mentioned in past literature (Newell 1947, Green and Macquitty, 1987, Bartsch, 1983, 1989, 1993): antero dorsal plate present; other dorsal plates sometimes absent; epimeral and genitoanal plates present and sometimes fused; idiosoma generally furnished with six pairs of dorsal setae and four or five pairs of gland pores; three or four segmented palpi inserted laterally on the gnathosoma; first leg often stouter than the other legs; solenidion on the first tarsus dorsolateral and that on the second tarsus dorsomedial in position; all tarsal ends furnished with two large lateral claws and one small median claw; carpite absent. In the diagnosis mentioned above, however, there is no unique characteristic by which Halacarinae is distinguishable from the other subfamilies. As mentioned in Bartsch (2015), Halacarinae is a bulky heterogeneous subfamily consisting of various rambling genera that do not share synapomorphies. The present results clearly showed molecular evidence for the polyphyletic nature of Halacarinae. Bartsch (2015) suggested an appropriate solution to address the incongruence between the current classification and phylogenetic grouping by including only the type genus Halacarus in the subfamily Halacarinae. While the results here presented partly support Bartsch’s proposition, how to classify the rest of the current halacarine genera in the taxonomic system of halacarid mites remains unclear.
A second incongruence between current and molecular based classifications highlighted in this study refers to the Rhombognathinae genera, as revealed by the 18S, 28S and 18S + 28S trees (Figures 1 and 2). The genera Rhombognathides and Metarhombognathus were clearly differentiated from Rhombognathus and close to Halacarinae and Halixodinae taxa in the 18S tree. In addition, a large differentiation between Rhombognathus and Metarhombognathus was also observed in the 28S and 18S + 28S trees. Moreover, Metarhombognathus was closer to Copidognathinae in the 28S tree and was also closer to the Halixodinae–Halacarinae cluster in the 18S + 28S tree, with a low bootstrap in both trees. The current results indicate that the phylogenetic position of Metarhombognathus remains unstable. According to Bartsch (2003), Rhombognathus is distinctive from the other three genera of Rhombognathinae (Isobactrus, Rhombognathides and Metarhombognathus) based on the developmental characteristics. This finding is line with the current results showing the large differentiations of Rhombognathus in the subfamily Rhombognathinae.

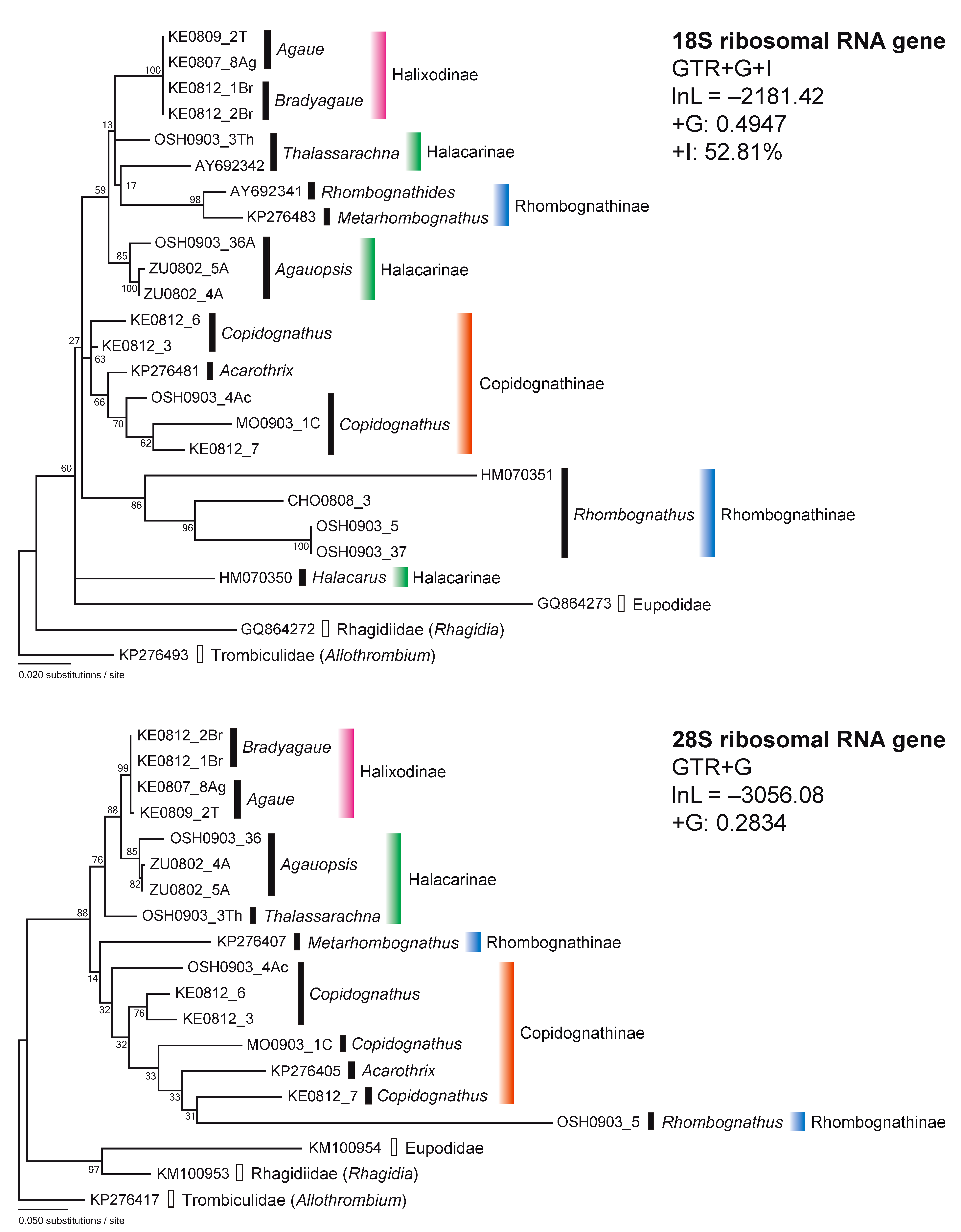

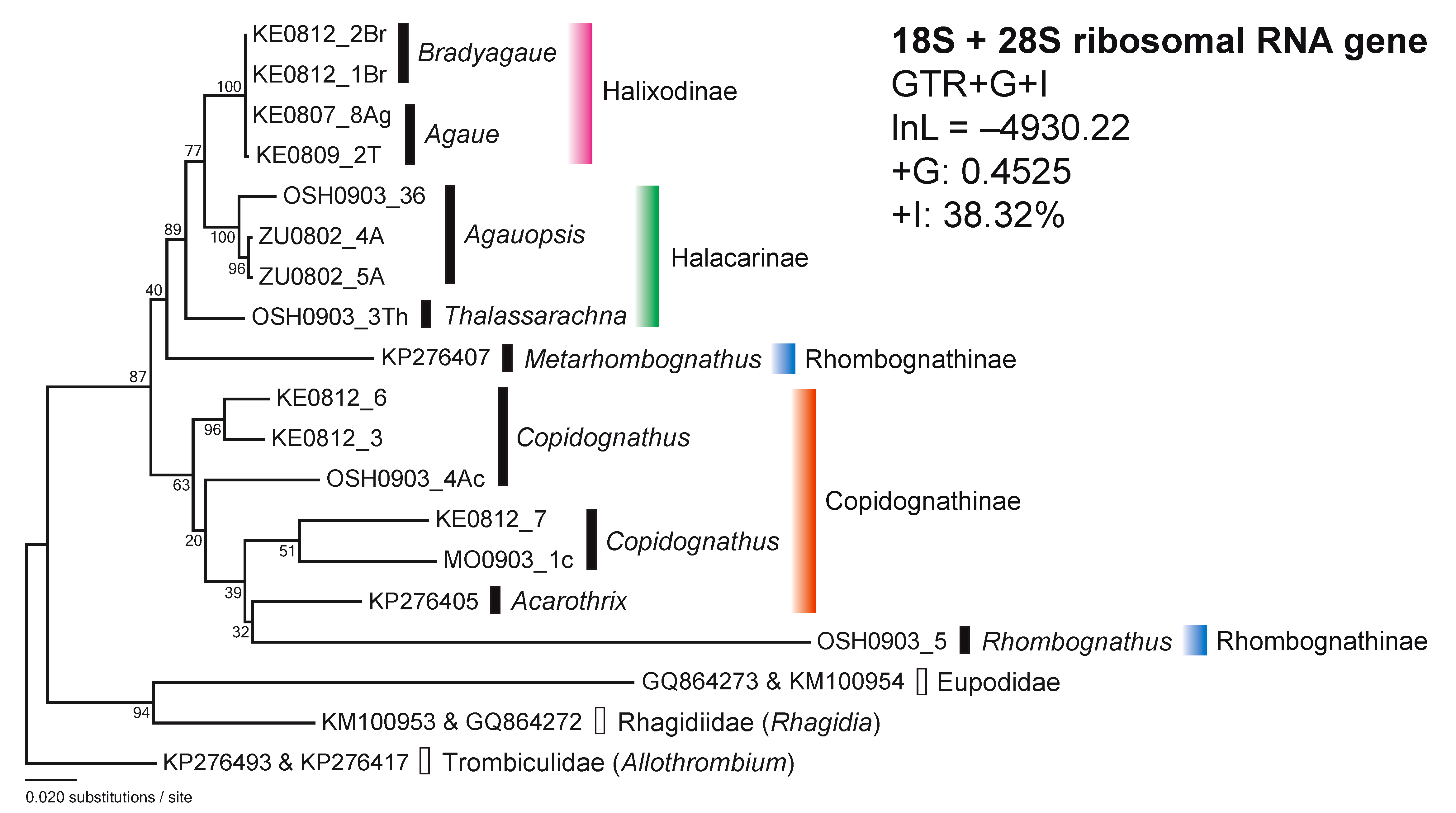
Based on the here presented phylogenetic trees, the unstable position of Rhombognathinae was confirmed and Rhombognathinae, particularly Rhombognathus, was closer to the copidognathine mites (Figures 1 and 2). According to Bartsch (1993), Copidognathinae is morphologically discernible from Rhombognathinae based on the leg characteristics, the gnathosoma together with the developmental stages which differ between the two taxa. Carpites are absent in the legs of Copidognathinae and present in those of Rhombognathinae. In addition, the gnathosoma is more globular in Rhombognathinae than in Copidognathinae. Likewise, Copidognathinae has a single nymphal stage (protonymph) throughout the development from larva to adult, whereas Rhombognathinae principally has three nymphal stages (protonymph, deutonymph and tritonymph). Although the morphological criteria are important for the current classification of halacarid mites (Bartsch 1993, 2006, 2008, 2010), it would be needed to reevaluate how much these three characters contribute to the phylogenetic relationships and evolutionary processes in halacarid mites, particularly for the Rhombognathinae. The conflict between the morphology-based taxonomic status and the molecular phylogenetic status here revealed, suggests that the halacarid taxonomy needs to be entirely reconstructed by using both molecular and morphological information for a revision of the family Halacaridae.
Abé H. 1990 — Method in the studies of halacarid mites. A guide to collection, preparation and identification — Seibutsu Kyouzai, 25: 21-45 (in Japanese).
Abé H. 1998 — Rhombognathine mites: taxonomy, phylogeny, and biogeography — Hokkaido University Press, Sapporo.
Abé H. 2001a — Phylogeny and character evolution of the marine mite genus Isobactrus (Acari: Halacaridae) — J. Nat. Hist., 35: 617-625. doi:10.1080/00222930151098279 ![]()
Abé H. 2001b — Phylogenetic taxonomy of the marine mite genus Rhombognathides (Acari: Halacaridae: Rhombognathinae) — Hydrobiologia, 464: 79-88. doi:10.1023/A:1013947110052 ![]()
Abé H., Bartsch I. 2007 — Historical biogeography of the halacarid mite genus Isobactrus (Prostigmata, Halacaridae). — In: Morares-Malacara J. B., Behan-Pelletier V., Ueckermann E., Pérez T. M., Estrada-Venegas E. G., Badii M. (Eds). Acarology XI: Proceedings of the International Congress. Mexico: Instituto de Biología and Facultad de Ciencias, Universidad Nacional Autónoma de México Sociedad Latinoamericana de Acarología, p. 119-123.
Bartsch I. 1983 — Vorschlag zur Neugliederung des Systems der Halacaridae (Acari). — Zool. Jb. Syst., 110: 179-200.
Bartsch I. 1989 — Süßwasser bewohnende Halacariden und ihre Einordnung in das System der Halacaroidea (Acari) — Acarologia, 30: 217-239.
Bartsch I. 1993 — Synopsis of the Antarctic Halacaroidea (Acari) — Koenigstein: Koeltz Scientific Books. pp. 176.
Bartsch I. 2003 — The subfamily Rhombognathinae: developmental pattern and re-evaluation of the phylogeny (Arachnida, Acari, Halacaridae) — Senckenberg. Biol., 82: 15–57.
Bartsch I. 2006 — Halacaroidea (Acari): a guide to marine genera — Organ Div. Evol. 6, Electronic Suppl. 6: 1–104.
Bartsch I. 2008 — Global diversity of halacarid mites (Halacaridae: Acari: Arachnida) in freshwater. Hydrobiologia, 595:317–322. doi:10.1007/s10750-007-9026-0 ![]()
Bartsch I. 2010 — Reflections on distribution and origin of the halacarid genus Isobactrus (Acari: Halacaridae) — Mar. Biodiv., 40:17-32. doi:10.1007/s12526-009-0030-7 ![]()
Bartsch I. 2015 — The genital area of Halacaridae (Acari), life stages and development of morphological characters and implication on the classification — Zootaxa, 3919: 201-259.
Brower A.V.Z., DeSalle R., Vogler A. 1996 — Gene trees, species trees, and systematics: A cladistic Perspective — Ann. Rev. Ecol. Syst., 27: 423–450. doi:10.1146/annurev.ecolsys.27.1.423 ![]()
Cruickshank, R.H. 2002 — Molecular markers for the phylogenetics of mites and ticks — System. Appl. Acarol., 7: 3-14.
Dabert M., Witalinski W., Kazmierski A., Olszanowski Z. and Dabert J. 2010 — Molecular phylogeny of acariform mites (Acari, Arachnida): strong conflict between phylogenetic signal and long-branch attraction artifacts — Mol. Phylogenet. Evol. 56: 222-241. doi:10.1016/j.ympev.2009.12.020 ![]()
Dabert M., Proctor H., Dabert J. 2016 — Higher level molecular phylogeny of the water mites (Acariformes: Prostigmata: Parasitengonina: Hydrachnidiae) — Mol. Phylogenet. Evol., 101: 75-90. doi:10.1016/j.ympev.2016.05.004 ![]()
Folmer O., Black M., Hoeh W., Lutz R., Vrijenhoek R. 1994 — DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates — Mol. Mar. Biol. Biotech., 3: 294-299.
Goka K., Okabe K., Yoneda M., Niwa S. 2001 — Bumblebee commercialization will cause worldwide migration of parasitic mites — Mol. Ecol., 10: 2095-2099. doi:10.1046/j.0962-1083.2001.01323.x ![]()
Goka K., Une Y., Kuroki T., Suzuki K., Nakahara M., Kobayashi A., Yokoyama J., Mizutani T., Hyatt A. D. 2009 — Amphibian chytridiomycosis in Japan: distribution, haplotypes, and possible entry into Japan — Mol. Ecol., 18: 4757–4774. doi:10.1111/j.1365-294X.2009.04384.x ![]()
Green J., Macquitty M. 1987 — Halacarid Mites (Synopses of the British Fauna No. 36 — London: The Bath Press. pp. 178. Higgins D.G., Thompson J.D., Gibson T.J. 1996 — Using CLUSTAL for multiple sequence alignments — Meth. Enzymol., 266: 383-402.
Hillis D.M., Dixon M.T. 1991 — Ribosomal DNA: Molecular evolution and phylogenetic inference — Qua. Rev. Biol., 66: 411-453. doi:10.1086/417338 ![]()
Krantz G.W. 1978 — A manual of Acarology — Corvallis: Oregon State University Bookstores.
Kumar S., Stecher G., Tamura K. 2016 — MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets — Mol. Biol. Evol. doi:10.1093/molbev/msw054 ![]()
Lanfear R., Calcott B., Ho S.Y.W., Guindon S. 2012 — PartitionFinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses — Mol. Biol. Evol., 29: 1695-1701. doi:10.1093/molbev/mss020 ![]()
Li H.-S., Xue X.-F., Hong X.-Y. 2014 — Homoplastic evolution and host association of Eriophyoidea (Acari, Prostigmata) conflict with the morphological-based taxonomic system — Mol. Phylogenet. Evol. 78: 185-198. doi:10.1016/j.ympev.2014.05.014 ![]()
Matsuda T., Morishita M., Hinomoto N., Gotoh T. 2014 — Phylogenetic analysis of the spider mite sub-family Tetranychinae (Acari: Tetranychidae) based on the mitochondrial COI gene and the 18S and the 5′ end of the 28S rRNA genes indicates that several genera are polyphyletic — PLoS ONE 9: e108672. doi:10.1371/journal.pone.0108672 ![]()
Mitani H., Takahashi M., Masuyama M., Fukunaga M. 2007 — Ixodes philipi (Acari: Ixodidae): phylogenetic status inferred from mitochondrial cytochrome oxidase subunit I gene sequence comparison — J. Parasitol. 93: 719-722. doi:10.1645/GE-953R.1 ![]()
Moore W.S. 1995 — Inferring phylogenies from mtDNA variation: mitochondrial-gene trees versus nuclear-gene trees — Evolution, 49: 718-726.
Navajas M., Boursot P. 2003 — Nuclear ribosomal DNA monophyly versus mitochondrial DNA polyphyly in two closely related mite species: the influence of life history and molecular drive — Proc. R. Soc. Lond. B (Suppl.), 270: S124-S127. doi:10.1098/rsbl.2003.0034 ![]()
Newell, I. M. 1947 — A systematic and ecological study of the Halacaridae of Eastern North America — Bull. Bingham. Oceanogr. Coll. New Haven, 10: 1-232.
Otto J.C., Wilson K.J. 2001 — Assessment of the usefulness of ribosomal 18S and mitochondrial COI sequences in Prostigmata phylogeny — In: Halliday B., Walter E., Proctor C., Norton A., Colloff M. J. (Eds). Acarology: Proceedings of the 10th International Congress. Melbourne: CSIRO Publishing, p. 100-109.
Pepato A.R., da Rocha C.E.F., Dunlop J.A. 2010 — Phylogenetic position of the acariform mites: sensitivity to homology assessment under total evidence — BMC Evol. Biol., 10: 235. doi:10.1186/1471-2148-10-235 ![]()
Pepato A.R. Klimov P.B. 2015 — Origin and higher-level diversification of acariform mites – evidence from nuclear ribosomal genes, extensive taxon sampling, and secondary structure alignment — BMC Evol. Biol., 15: 178. doi:10.1186/s12862-015-0458-2 ![]()
Söller R., Wohltmann A., Witte H., Blohm D. 2001 — Phylogenetic relationships within terrestrial mites (Acari: Prostigmata, Parasitengona) inferred from comparative DNA sequence analysis of the mitochondrial cytochrome oxidase subunit I gene — Mol. Phylogenet. Evol., 18: 47-53. doi:10.1006/mpev.2000.0855 ![]()
Swofford D.L., Olsen G.J., Waddell P.J., Hillis D.M. 1996 — Phylogenetic Inference — In: Molecular Systematics, 2nd edition. Sunderland: Sinauer and Associates. p. 407–514.
Tamura K., Nei M. 1993 — Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees — Mol. Biol. Evol., 10: 512-526.
Walter D.E., Proctor H.C. 1999 — Mites: Ecology, Evolution and Behaviour — Wallingford: CABI Publishing.



2016-08-04
Date accepted:
2016-01-15
Date published:
2017-05-16
Edited by:
Navajas, Maria

This work is licensed under a Creative Commons Attribution 4.0 International License
2017 Meguro, Hiroko; Iwasa, Masahiro A.; Goka, Koichi and Abé, Hiroshi
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)