



The mesostigmatid mite most commonly associated with stingless bees in southern Brazil: complementary description and generic reassignment to Bisternalis (Mesostigmata: Laelapidae)
Godoy, Lucas Lorena  1
; Da-Costa, Tairis
2
and Castilho, Raphael de Campos
3
1
; Da-Costa, Tairis
2
and Castilho, Raphael de Campos
3
1Departamento de Entomologia e Acarologia, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 13418–900 Piracicaba, SP, Brazil.
2Laboratório de Acarologia, Tecnovates, Universidade do Vale do Taquari – Univates, Lajeado, Rio Grande do Sul 95913-528, Brazil.
3Departamento de Entomologia e Acarologia, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, 13418–900 Piracicaba, SP, Brazil.
2026 - Volume: 66 Issue: 2 pages: 612-630
https://doi.org/10.24349/j9vz-z6j9ZooBank LSID: 5F2EB50D-5182-49E9-8585-220C7EF33FD6
Original research
Keywords
Abstract
Introduction
Nests of eusocial hymenopterans constitute biodiversity-rich microhabitats that frequently harbor diverse assemblages of mites, including Mesostigmata. However, the diversity and biology of many of these associates remain poorly understood (Eickwort 1990; Klompen and Campbell 2025).
The mite family Laelapidae (Mesostigmata) comprises species that occupy a wide range of habitats; many of them are associated with insects (Moraes et al. 2022). Among these, several genera include species associated with bees (Hymenoptera) of the family Apidae, such as Androlaelaps Berlese, 1903, Bisternalis Hunter, 1963, Dinogamasus Kramer, 1898, Eumellitiphis Turk, 1948, Hunteria Delfinado-Baker, Baker & Flechtmann, 1984, Meliponaspis Vitzthum, 1930, Melittiphis Berlese, 1918, Melittiphisoides Delfinado-Baker, Baker & Flechtmann, 1984, Neohypoaspis Delfinado-Baker, Baker & Roubik, 1983, Pneumolaelaps Berlese, 1920, Stevelus Hunter, 1963, Stigmatolaelaps Krantz, 1998, Suracarus Flechtmann, 2005, Tropilaelaps Delfinado & Baker, 1961, Urozercon Berlese, 1901 and Xylocolaelaps Royce & Krantz, 2003. Laelaspoides Eickwort, 1966 and Raymentia Womersley, 1956 have been reported in association with bees of the family Halictidae. All of these mites may inhabit nests or occur on their hosts' bodies. The ecological roles and impacts of these mites on insect colonies remain poorly understood. The well-known Varroa Oudemans, 1904 is not included among those genera for being currently placed in its own family Varroidae (Lindquist et al. 2009).
In 2022, a group of dark brown mesostigmatid mites was collected from nests of a stingless bee species (Apidae: Meliponini) in the state of Santa Catarina, southern Brazil. In 2025, additional specimens were collected in association with stingless bees in the state of São Paulo, southeastern Brazil (relatively close to northern Santa Catarina). These specimens were subsequently sent to the Acarology Laboratory, Department of Entomology and Acarology, Escola Superior de Agricultura ''Luiz de Queiroz'' (ESALQ), Universidade de São Paulo (USP), Piracicaba, state of São Paulo, Brazil, for identification. Preliminary examination of slide-mounted specimens indicated that they belonged to Laelapidae.
Additionally, in a recent survey of the acarofauna associated with stingless bees, Da-Costa et al. (2021) examined colonies of Melipona quadrifasciata quadrifasciata Lepeletier, Scaptotrigona bipunctata Lepeletier, and Tetragonisca fiebrigi Schwarz across seven localities in the state of Rio Grande do Sul, southern Brazil (a neighboring state of Santa Catarina). Among the mites recorded, laelapids were particularly prominent. One taxon, initially identified as Hypoaspis alphabeticus (Berlese, 1904), was by far the most abundant species, accounting for 773 of the 938 individuals collected, all of which were associated with M. quadrifasciata quadrifasciata. Da-Costa et al. (2021) interpreted the high abundance and exclusive occurrence of H. alphabeticus in Melipona colonies (among the stingless bee hosts sampled) as evidence of a host-restricted association, reinforcing earlier suggestions that certain laelapid genera may maintain specialized relations with eusocial Hymenoptera.
This finding led us to consider that the species under study from Santa Catarina and São Paulo could correspond to the predominant taxon reported by Da-Costa et al. (2021). In connection with these records, L. (H.) alphabeticus was originally described from Brazil, with specimens collected from a nest of Melipona mandacaia Smith (cited as Meliponae mandaçaiae).
These specimens seemed to be most closely related to Bisternalis, but differed in some diagnostic characteristics, as detailed in the Results section. This raised a taxonomic dilemma: whether to describe a new genus or to revise the concept of Bisternalis to accommodate the specimens at hand. The present study was conducted to test the hypothesis that the specimens from the three populations belonged to a single laelapid species, through a detailed morphological characterization of the taxon and an evaluation of its current generic placement.
Material and methods
The specimens from the three populations examined in this study were collected from bee nests in the states of Rio Grande do Sul, Santa Catarina, and São Paulo, Brazil. The mites were mounted on slides in Hoyer's medium and subsequently examined and measured under differential interference contrast microscopy (Zeiss Axio Imager A2). Photographs were taken with a Zeiss AxioCam HRc camera attached to the microscope, and illustrations were prepared using Adobe Illustrator 2020.
In the complementary descriptions provided in this publication, shield lengths were taken along the midline from the anterior to the posterior margins, and widths, at the widest level, except where otherwise specified. For the sternal shield, length was considered from the level of insertion of setae st1 to the posterior margin of the shield; the length of the epigynal shield included the hyaline flap. The length of the corniculus was taken along its median longitudinal line, and the width, at the widest level; leg lengths were measured from the base of the coxa to the apex of the tarsus, not including the pretarsus. The notation of the idiosomal setae is based on Lindquist and Evans (1965); leg chaetotaxy, on Evans (1963a); palp chaetotaxy, on Evans (1963b); and notation of pore or pore-like structures, on Athias-Henriot (1971, 1975).
To determine the genus of the specimens examined, a list of morphologically similar laelapid genera associated with Hymenoptera was compiled from the literature. A dichotomous key was then prepared to separate them. A key to the currently recognized species of Bisternalis was also constructed, based on specimens collected and on literature information.
Results and discussion
Examination of the available specimens led to the conclusion that they all belonged to a single species, originally described as Laelaps (Hypoaspis) alphabeticus Berlese, despite slight differences in a few characters, as subsequently discussed. In recent publications, this species has been assigned to Hypoaspis, but Moraes et al. (2022) considered it as of uncertain generic placement. Details of its nomenclature and morphology are provided below:
Bisternalis alphabeticus (Berlese, 1904) n. comb.
Laelaps (Hypoaspis) alphabeticus Berlese, 1904: 17.
Hypoaspis alphabeticus.— Buitendijk, 1945: 295; Turk, 1948: 113; Castagnoli and Pegazzano, 1985: 10; Moraes et al. 2022: 404.
alphabeticus non-Laelaps.— Tipton, 1960: 286.
Hypoaspis alphabetica.— Klimov et al. 2016: internet page.
Hypoaspis alfabetica [sic].— Da-Costa et al. 2021: 4.
Material examined
All specimens were collected from nests of Melipona quadrifasciata quadrifasciata (Hymenoptera: Apidae) in Brazil. Rio Grande do Sul State: one adult female at Porto Alegre (30°4′S, 51°10′W), three adult females and one adult male at Bom Princípio (29°30′S, 51°22′W), and six adult females and two adult males at Rolante (29°39′S, 50°34′W); all collected by Tairis Da-Costa, between September 2018 and February 2019. Santa Catarina State: 10 adult females and one adult male at Florianópolis (27°59′09″S, 48°29′30″W); all collected by Bernardo Telles de Souza, in January 2022. São Paulo State: six adult females at Jaguariúna (22°43′54″S, 47°01′06″W); all collected by Denise de Araujo Alves, in October 2025.
Diagnosis
Adult female — Dorsal shield with 39 pairs of setae plus 4-7 postero-median unpaired setae. Sternal shield with posterior margin slightly concave. Seta st1 anteriad a distinct lateral incision of the sternal shield. Epigynal shield snake-head-shaped, posteriorly blunt, with U-shaped striae, and not extending beyond the middle of coxa IV. Anal shield subtriangular, wider than long, with anterior margin concave, and with a pair of lateral incisions between levels of para-anal and post-anal setae. Parapodal plates crescent-shaped and narrow.
Adult male — Similar to adult female in dorsal morphology. Venter with holoventral shield bearing ten pairs of setae (st1–st5, JV1–JV3, ZV1, ZV2), besides the circum-anal setae. Unsclerotized opisthogastric cuticle with four pairs of setae. Femur II with a short, distally blunt conical seta (pv2).
Complementary descriptions
Adult female — (10 specimens from Rio Grande do Sul, 10 from Santa Catarina, and 6 from São Paulo) – (Figures 1–3; Table 1).
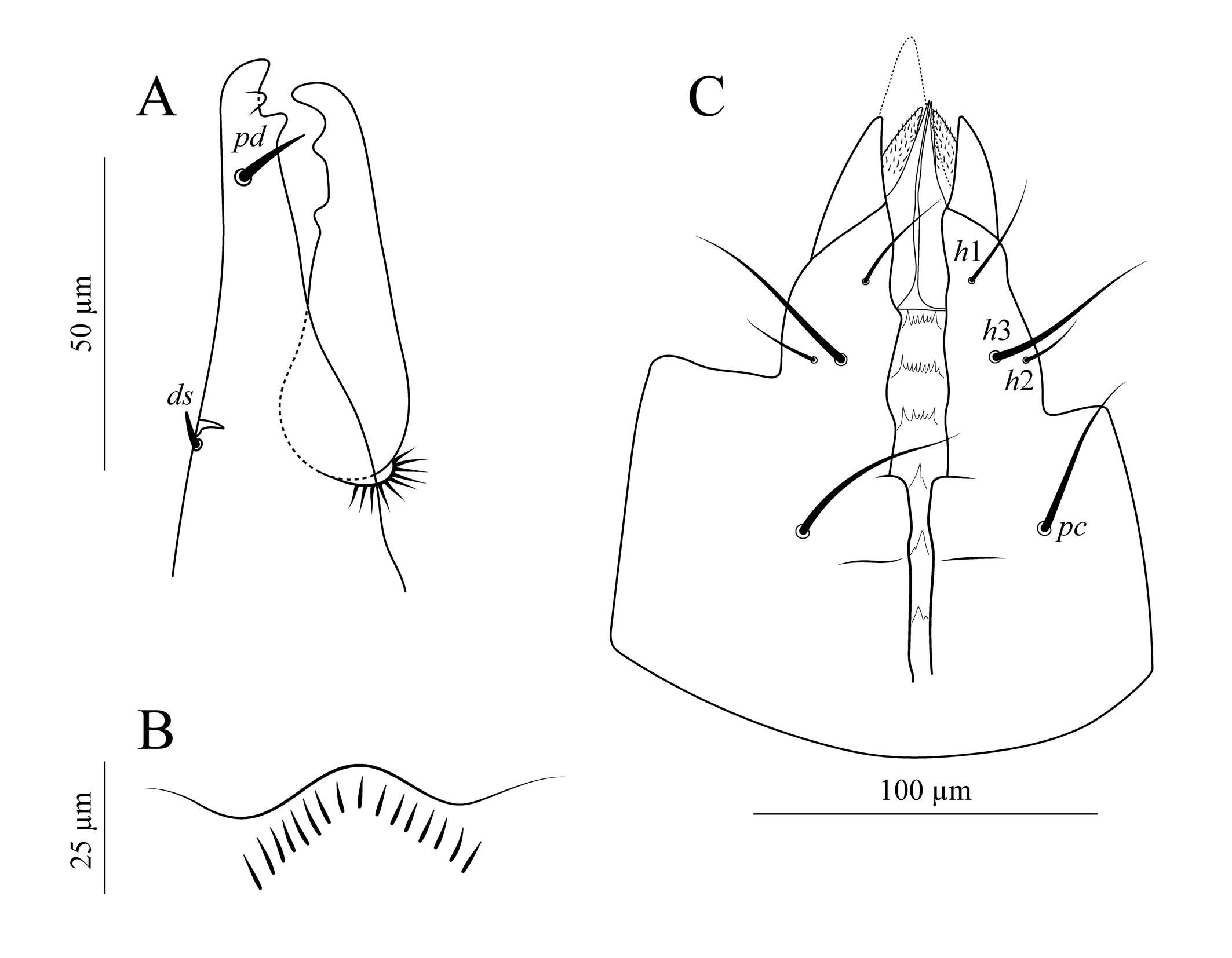
Gnathosoma. Fixed cheliceral digit (Figure 1A) with two subapical teeth not aligned with the apical tooth and a setiform pilus dentilis; movable digit with two teeth in addition to the apical tooth, all aligned; dorsal cheliceral seta small and prostrate forward; dorsal lyrifissure distinct; arthrodial membrane with distinct transversely aligned ciliate structures. Number of setae on palp trochanter–tibia normal (2-5-6-14; tarsus not counted); setae acicular and smooth, except two slightly stout lateral setae on palp genu; apotele 2-tined. Epistome (Figure 1B) weakly sclerotized, with smooth anterior margin surrounded by a series of short, nearly parallel internal lines perpendicular to the margin. Deutosternum (Figure 1C) delimited by lateral lines, about twice as wide in the anterior as in the posterior half; with seven transverse lines, the anterior margin smooth, followed by three lines each bearing 6–9 denticles, these followed by the remaining lines with 1–2 denticles each. Internal mala reaching slightly past the level of the tip of the corniculus; inner margin smooth and outer margin fringed. Corniculus horn-like. Seta h3 posterolaterad h1 and transversely aligned with h2, all setiform and smooth. Labrum reaching well beyond the tip of the malae, tapering distally and ciliate. Measurements in Table 1.
Download as
Characteristics
Female
Male
RS (n=10)
SC (n=10)
SP (n=6)
Average
RS (n=3)
SC (n=1)
Average
FCD
73 (69–77)
66 (60–71)
70 (68–72)
69 (60–77)
51 (49–55)
40
46 (40–55)
MCD
65 (60–70)
63 (55–67)
65 (65–65)
64 (55–70)
44 (42–48)
48
46 (42–48)
PD
10 (10–11)
10 (8–11)
9 (8–9)
9 (8–11)
-
-
-
CL
48 (44–52)
51 (44–57)
42 (40–44)
47 (40–57)
47 (46–53)
49
48 (46–53)
CW
14 (11–16)
17 (14–21)
14 (13–15)
15 (11–21)
14 (13–15)
15
15 (13–15)
h1
41 (33–51)
38 (30–50)
40 (36–44)
39 (30–51)
47 (45–51)
45
46 (45–51)
h2
21 (20–24)
23 (21–29)
24 (23–25)
23 (20–29)
21 (20–24)
23
22 (20–24)
h3
76 (72–80)
74 (67–81)
75 (72–78)
75 (67–81)
76 (74–78)
70
73 (74–78)
Pc
53 (52–55)
54 (45–60)
55 (53–56)
54 (45–60)
54 (51–55)
54
54 (51–55)
IL
914 (867–999)
919 (871–947)
944 (939–948)
925 (867–999)
858 (790–950)
768
813 (768–950)
IW
613 (552–658)
555 (518–582)
615 (607–623)
594 (518–658)
567 (501–637)
530
548 (501–637)
DSL
871 (807–964)
885 (845–940)
910 (899–921)
889 (807–964)
835 (765–933)
760
798 (760–933)
DSW
578 (542–634)
549 (520–637)
587 (583–590)
571 (520–637)
528 (477–591)
480
504 (477–591)
j1
50 (50–61)
62 (55–72)
61 (54–68)
57 (51–72)
60 (52–72)
59
60 (52–72)
j2
30 (26–31)
52 (48–56)
50 (41–59)
44 (26–59)
53 (49–56)
60
56 (49–60)
j3
41 (35–46)
46 (42–50)
48 (45–52)
45 (35–52)
47 (44–49)
38
42 (38–49)
j4
47 (42–52)
44 (37–50)
47 (46–48)
46 (37–52)
46 (44–50)
49
48 (44–50)
j5
43 (37–46)
36 (30–44)
43 (39–46)
41 (30–46)
36 (30–40)
47
42 (30–47)
j6
42 (38–46)
41 (36–47)
42 (39–46)
42 (36–47)
40 (39–42)
40
40 (39–42)
z1
30 (24–36)
28 (24–32)
27 (25–30)
28 (24–36)
24 (21–28)
25
25 (21–28)
z2
41 (38–45)
49 (44–53)
51 (48–53)
47 (38–53)
50 (47–52)
51
50 (47–52)
z3
53 (50–55)
51 (46–56)
54 (50–56)
53 (46–56)
52 (49–56)
58
55 (49–58)
z4
53 (48–58)
50 (48–53)
54 (51–57)
52 (48–57)
52 (49–55)
55
53 (49–55)
z5
45 (39–49)
44 (40–49)
44 (41–45)
44 (39–49)
43 (41–47)
46
45 (41–47)
z6
46 (42–49)
42 (36–47)
44 (42–46)
44 (36–49)
41 (39–42)
48
45 (39–48)
s1
46 (41–53)
60 (56–65)
64 (61–65)
57 (41–65)
50 (45–55)
68
59 (45–68)
s2
52 (48–54)
56 (52–62)
61 (57–63)
56 (48–63)
54 (52–59)
64
59 (52–64)
s3
53 (47–58)
62 (59–69)
65 (63–67)
60 (47–69)
59 (56–62)
68
64 (56–68)
s4
54 (51–58)
54 (50–61)
57 (56–58)
55 (50–61)
61 (53–69)
58
60 (53–69)
s5
55 (52–59)
57 (53–63)
57 (57–58)
57 (52–63)
60 (55–65)
64
62 (55–65)
s6
63 (57–68)
70 (64–80)
76 (72–81)
70 (57–81)
65 (58–68)
82
73 (58–82)
r2
61 (58–66)
66 (58–75)
64 (61–67)
64 (58–75)
57 (52–66)
85
71 (52–85)
r3
60 (54–66)
70 (65–76)
67 (66–67)
65 (54–76)
53 (50–57)
75
64 (50–75)
r4
61 (51–73)
67 (62–72)
68 (67–69)
65 (51–73)
58 (53–62)
72
65 (53–72)
r5
66 (62–70)
70 (66–77)
67 (65–69)
68 (62–77)
62 (60–64)
80
71 (60–80)
r6
71 (60–78)
77 (68–84)
78 (74–81)
75 (60–84)
59 (58–61)
91
75 (58–91)
J1
38 (33–41)
34 (32–38)
40 (37–45)
38 (32–45)
37 (33–40)
38
38 (33–40)
J2
38 (34–42)
35 (32–40)
43 (39–45)
39 (32–45)
41 (38–45)
37
39 (37–45)
J3
39 (32–41)
34 (27–41)
40 (39–42)
38 (27–42)
37 (31–40)
33
35 (31–40)
J4
47 (43–50)
45 (43–47)
49 (46–51)
47 (43–51)
45 (44–47)
59
52 (44–59)
J5
60 (57–63)
57 (51–61)
58 (53–62)
58 (51–63)
55 (53–58)
61
58 (53–61)
Z1
50 (45–54)
52 (48–56)
58 (55–63)
53 (45–63)
47 (43–49)
53
50 (43–53)
Z2
60 (55–65)
58 (54–65)
57 (50–61)
58 (50–65)
48 (45–50)
58
53 (45–58)
X1–7
35 (31–39)
33 (31–35)
35 (32–39)
35 (31–39)
35 (33–36)
33
34 (33–36)
Zx2
40 (34–49)
45 (40–50)
54 (53–55)
46 (34–55)
40 (36–44)
46
43 (36–46)
Z3
61 (54–65)
61 (55–69)
69 (67–71)
64 (54–71)
52 (49–55)
63
58 (49–63)
Zx3
43 (40–48)
44 (39–48)
53 (49–56)
46 (39–56)
38 (36–40)
45
41 (36–45)
Z4
73 (70–77)
71 (66–81)
76 (73–78)
73 (66–81)
77 (66–99)
74
76 (66–79)
Z5
82 (79–85)
81 (76–89)
75 (67–82)
79 (67–89)
86 (75–100)
92
89 (75–100)
S1
70 (68–71)
67 (61–75)
74 (70–77)
70 (61–77)
62 (58–66)
75
69 (58–75)
S2
76 (72–84)
74 (70–86)
79 (75–82)
76 (70–86)
66 (63–69)
84
75 (63–84)
S3
77 (73–83)
77 (72–86)
80 (78–82)
78 (72–86)
72 (70–74)
84
78 (70–84)
S4
83 (80–88)
80 (72–91)
86 (85–86)
83 (72–91)
76 (74–79)
91
84 (74–91)
S5
83 (80–85)
84 (80–92)
88 (87–88)
85 (80–92)
65 (63–68)
93
79 (63–93)
R1
71 (62–78)
71 (65–78)
77 (70–77)
73 (62–78)
70 (66–75)
74
72 (66–75)
R2
73 (67–78)
71 (66–81)
77 (72–81)
74 (66–81)
72 (65–89)
82
77 (65–89)
R3
77 (70–84)
78 (74–81)
76 (70–82)
77 (70–84)
73 (72–75)
84
79 (72–84)
R4
80 (75–84)
78 (70–83)
79 (78–80)
79 (70–84)
76 (70–79)
88
82 (70–88)
R5
79 (77–80)
79 (74–92)
77 (74–80)
78 (74–92)
83 (80–88)
94
88 (80–94)
R6
84 (80–89)
86 (72–92)
87 (83–90)
85 (72–92)
83 (81–85)
99
91 (81–99)
Ant. UR
68 (63–71)
73 (67–78)
75 (70–79)
72 (63–79)
72 (67–75)
84
78 (67–84)
Post. UR
79 (72–85)
75 (70–81)
78 (76–80)
77 (70–85)
70 (68–71)
80
75 (68–80)
BTL
17 (15–20)
15 (13–17)
19 (15–20)
17 (13–20)
17 (15–18)
14
16 (15–18)
BTW
24 (21–25)
24 (21–25)
19 (17–21)
22 (17–25)
24 (22–25)
23
24 (22–25)
Lacinia
140 (132–148)
141 (135–148)
133 (129–146)
138 (129–148)
144 (139–148)
138
141 (139–148)
Dist. st1-st3
146 (135–155)
126 (117–135)
135 (130–140)
135 (117–155)
-
-
-
Dist. st2-st2
175 (170–182)
176 (171–181)
175 (170–180)
175 (170–182)
-
-
-
Dist. st3-st3
229 (222–241)
225 (213–234)
222 (214–230)
225 (213–241)
-
-
-
ESL
360 (338–385)
404 (349–431)
421 (419–423)
395 (338–431)
-
-
-
ESW
167 (156–176)
201 (188–214)
209 (195–222)
192 (156–222)
-
-
-
ASL
99 (85–113)
112 (101–121)
109 (108–110)
107 (85–121)
-
-
-
ASW
215 (210–225)
215 (204–228)
235 (230–240)
222 (204–240)
-
-
-
MS1
22 (20–25)
28 (21–36)
29 (27–32)
26 (20–36)
-
-
MS2
19 (16–21)
22 (18–26)
21 (19–22)
21 (16–26)
-
-
MS3
45 (41–51)
46 (40–53)
42 (41–48)
44 (40–53)
-
-
HSL
-
-
-
-
675 (610–771)
650
663 (610–771)
HSW
-
-
-
-
350 (325–390)
325
338 (325–390)
st1
69 (63–74)
65 (61–69)
50 (50–52)
61 (50–69)
33 (26–39)
38
36 (26–39)
st2
72 (68–77)
66 (54–76)
61 (59–62)
66 (54–76)
34 (32–37)
39
36 (32–39)
st3
59 (53–63)
52 (47–59)
54 (52–56)
55 (47–63)
32 (30–33)
35
33 (30–35)
st4
63 (57–67)
54 (47–57)
52 (50–54)
56 (47–67)
38 (34–44)
48
43 (34–48)
st5
59 (52–64)
53 (47–60)
51 (46–55)
54 (46–64)
44 (42–45)
49
46 (42–49)
JV1
44 (40–47)
40 (34–48)
37 (35–39)
40 (34–48)
36 (33–38)
38
37 (33–38)
JV2
47 (32–52)
45 (41–49)
41 (34–47)
44 (32–52)
37 (36–38)
42
40 (36–42)
JV3
52 (47–56)
50 (43–55)
42 (41–42)
48 (41–56)
41 (40–42)
47
44 (40–47)
JV4
76 (74–78)
76 (70–84)
79 (78–80)
77 (70–84)
79 (77–81)
85
82 (77–85)
JV5
88 (83–91)
85 (80–90)
89 (86–92)
87 (80–92)
85 (76–89)
86
86 (76–89)
ZV1
42 (39–47)
39 (34–43)
39 (38–40)
40 (34–47)
37 (35–38)
39
38 (35–39)
ZV2
48 (46–50)
50 (43–57)
43 (41–44)
47 (41–57)
39 (37–42)
44
42 (37–42)
ZV3
67 (63–70)
67 (60–75)
68 (65–71)
67 (60–75)
55 (45–60)
74
64 (45–74)
ZV4
79 (72–84)
82 (77–87)
81 (76–86)
81 (72–87)
70 (60–78)
80
75 (60–80)
PaA
56 (52–58)
53 (43–58)
49 (48–50)
52 (43–58)
55 (52–60)
54
55 (52–60)
PoA
83 (71–95)
83 (77–89)
79 (73–84)
82 (71–95)
85 (81–95)
78
82 (81–95)
Leg I
731 (687–790)
691 (621–723)
755 (735–775)
726 (621–790)
681 (568–800)
605
643 (568–800)
Leg II
590 (536–637)
575 (489–670)
601 (561–640)
588 (489–670)
522 (461–574)
504
513 (461–574)
Leg III
638 (602–697)
584 (527–675)
659 (635–683)
627 (527–697)
596 (538–686)
500
548 (500–686)
Leg IV
746 (691–791)
724 (637–773)
789 (780–816)
756 (637–816)
683 (596–759)
625
654 (596–759)
Idiosoma. Oval; unsclerotized cuticle with fingerprint-like pattern.
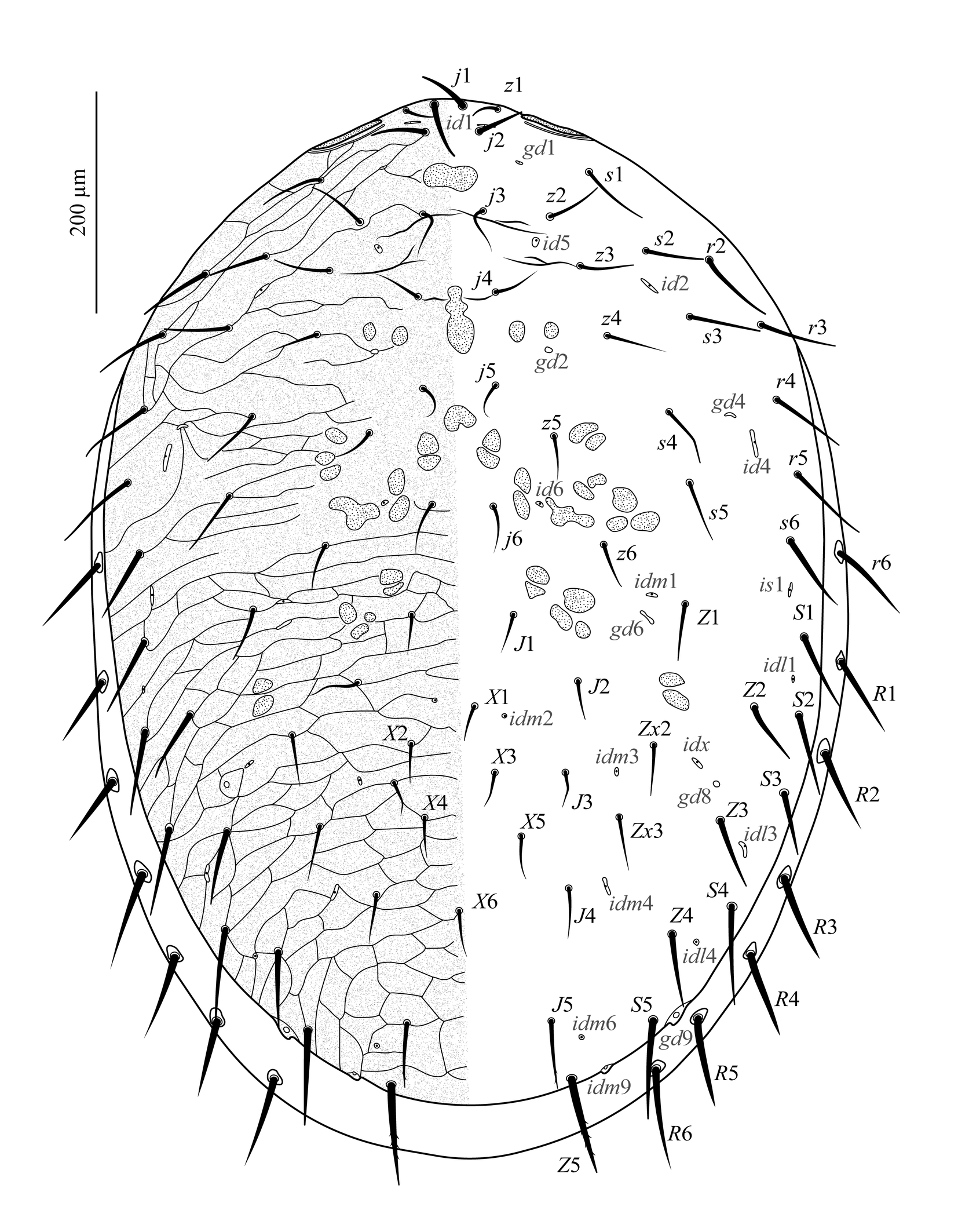
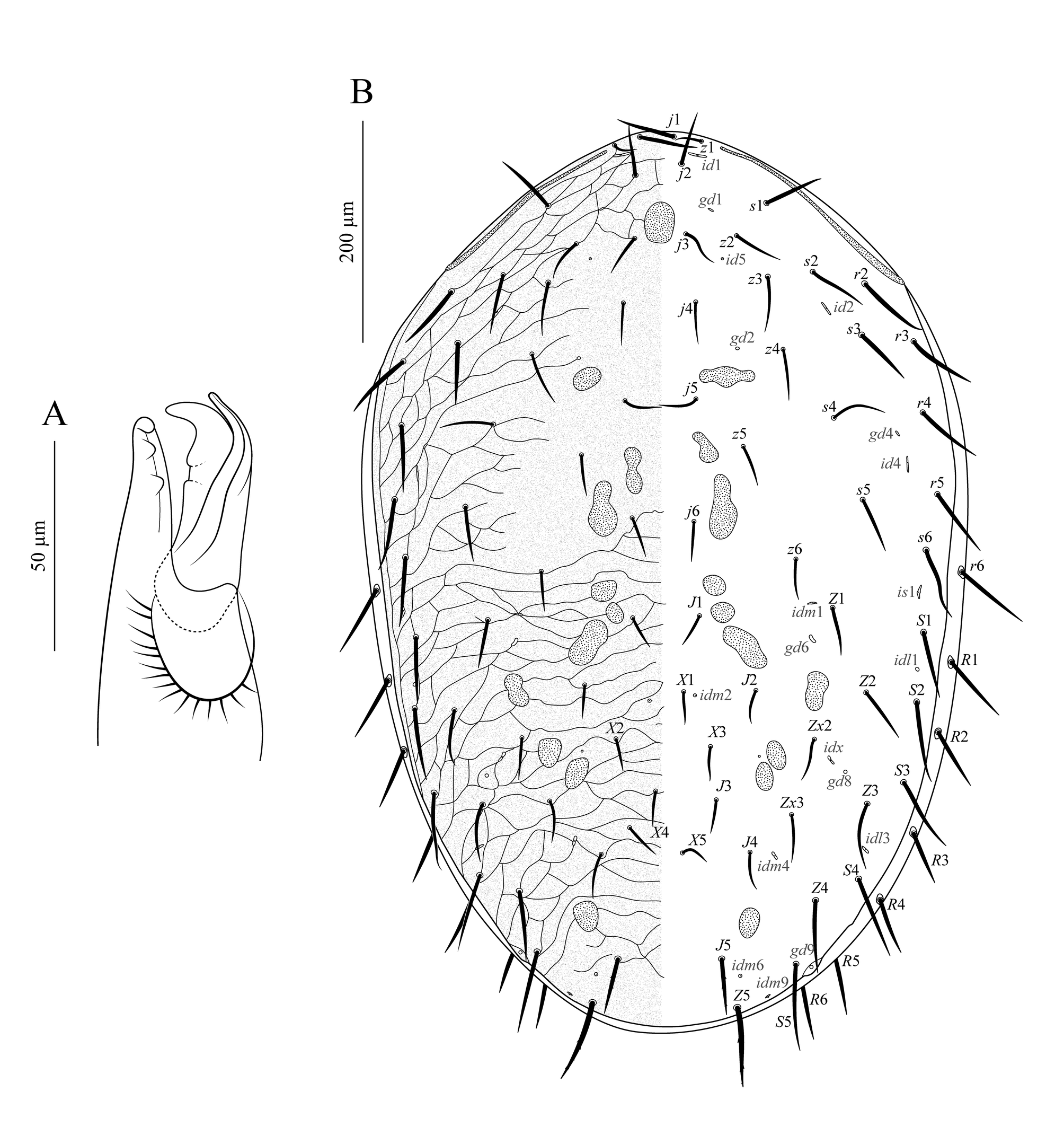
Dorsum of idiosoma (Figure 2). Dorsal shield entire, finely punctate throughout, covering about 90% of the idiosoma, leaving a thin band of unsclerotized cuticle laterally and posteriorly. Podonotal region reticulate, except for a smooth medio-longitudinal band that widens up between j4 and j6; with 22 pairs of setae (j1–j6, z1–z6, s1–s6, r2–r5), five pairs of distinguishable lyrifissures (id1, id2, id4, id5, id6), and three pairs of distinguishable gland pores (gd1, gd2, gd4). Unsclerotized cuticle along lateral margins of the podonotal region with one pair of setae (r6), inserted in a tiny, rounded sclerite. Opisthonotal region reticulate over all of its extent; with 17 pairs of setae (J1–J5, Z1–Z5, S1–S5, Zx2, Zx3), 1–7 unpaired setae between levels of J2 and J4 (from Rio Grande do Sul: six specimens with four, two with three, one with two and one with one; from Santa Catarina: three with seven, four with six, two with five, and one with four; from São Paulo: two with three, two with two, two with one), 11 pairs of distinguishable lyrifissures (is1, idl1, idl3, idl4, idm1, idm2, idm3, idm4, idm6, idm9, idx), and three pairs of distinguishable gland pores (gd6, gd8, gd9). Unsclerotized cuticle along lateral margins of the opisthonotal region with one gland pore (gd3) and eight pairs of setae (R1–R6 and two UR), each inserted in a tiny, rounded sclerite; lyrifissures Rp indistinguishable; R5, R6, UR, and gd3 most often visible in ventral view. Setae setiform and smooth, except J5 and Z5, slightly barbed. Measurements in Table 1.
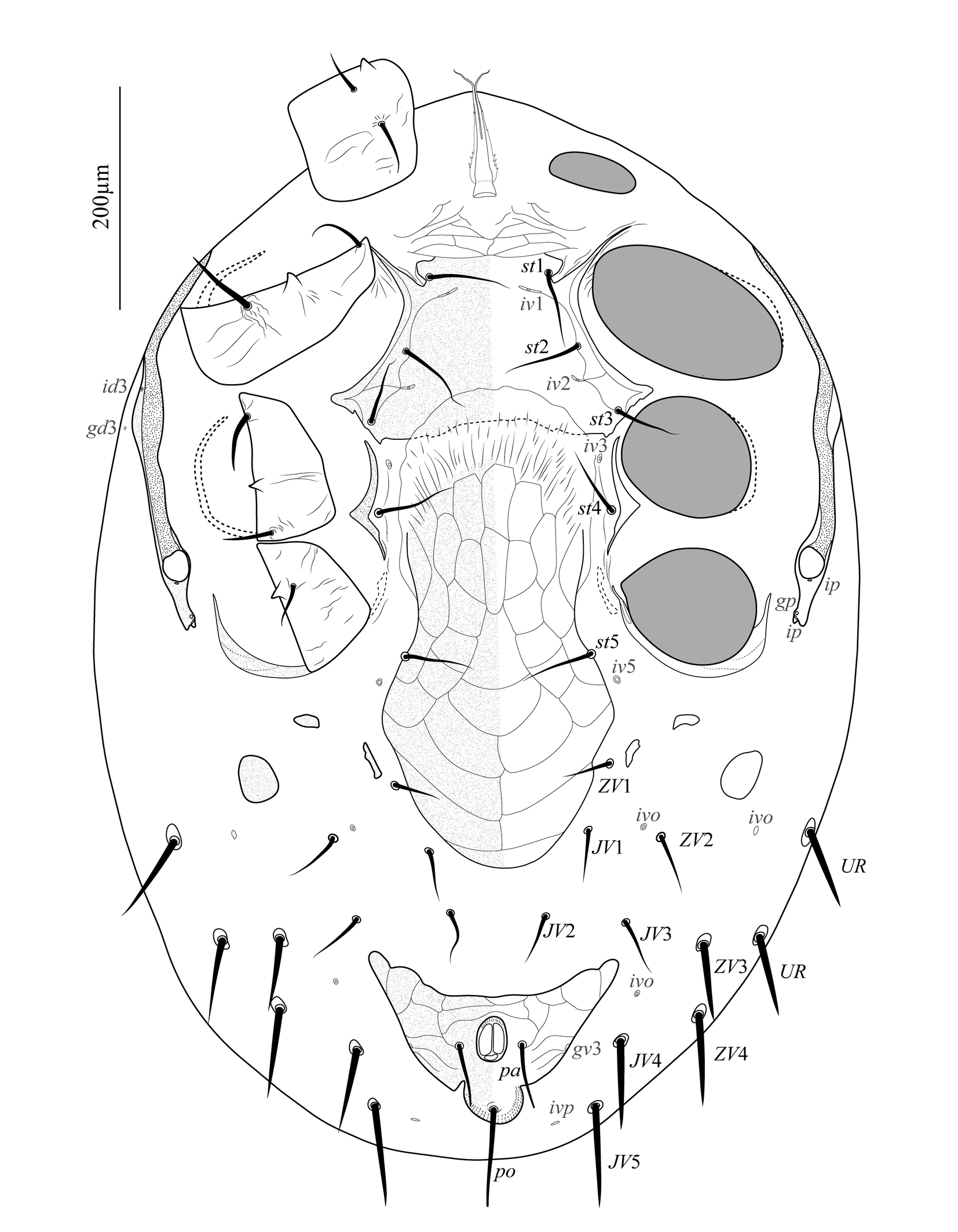
Venter of idiosoma (Figure 3). Base of tritosternum wider than long, with line of fusion with laciniae barely distinguishable; laciniae separated for about 60% of their total length, with a slender fimbriate membrane-like margin. Presternal area lightly sclerotized, reticulate, with anterior and lateral margins indistinct and posteriorly connected to the main section of the sternal shield by a narrow and smooth transverse band delineated by an incision posteriad each st1. The remaining section of the sternal shield smooth, except for a few striae along the lateral margins; with setae st2 and st3, and two pairs of lyrifissures (iv1 and iv2); posterior margin slightly concave. Metasternal plates absent. Seta st4 and lyrifissure iv3 on unsclerotized cuticle. Section of endopodal plate laterad coxae III–IV represented by a triradiate fragment; genital apodemes barely distinguishable. Epigynal shield snake-head-shaped, posteriorly blunt, ornamented with large reticles; with a single pair of setae (st5). Lyrifissure iv5 on unsclerotized cuticle, posterolaterad st5. Three pairs of metapodal platelets, the outermost distinctly larger than the others and oval; others elongate to rounded. Anal shield subtriangular, with anterior margin concave (medially straight in some specimens); reticulate, with a pair of lateral incisions between para-anal and post-anal setae; cribrum composed of three curved and parallel lines of denticles along posterior margin of the shield behind incisions. Unsclerotized opisthogastric cuticle with nine pairs of setae (JV1–JV5, ZV1–ZV4), and four pairs of distinguishable lyrifissures (three pairs of ivo and one pair of ivp); JV4, JV5, ZV3, and ZV4 each inserted in a tiny, rounded sclerite, and others in the unsclerotized cuticle. Parapodal plates crescent-shaped, narrow; exopodal plates indistinguishable. All setae setiform and smooth. All ventral shields and platelets finely punctate. Measurements in Table 1.
Peritreme and peritrematic plate. Peritreme extending anteriorly to the level between j2 and z1. Peritrematic plate narrow, fused to dorsal shield at level of j2; posteriorly, extending to about mid-level of coxa IV, with posterior end discreetly bifurcate; with lyrifissure id3 at the level of coxa II; with two lyrifissures ip and a gland pore gp behind each stigma.
Spermathecal apparatus (Figure 4). Well visible in some specimens. With a pair of tubuli of about uniform diameter in most of their extension, flaring slightly towards both ends. With distinct constriction immediately before reaching the ramus.
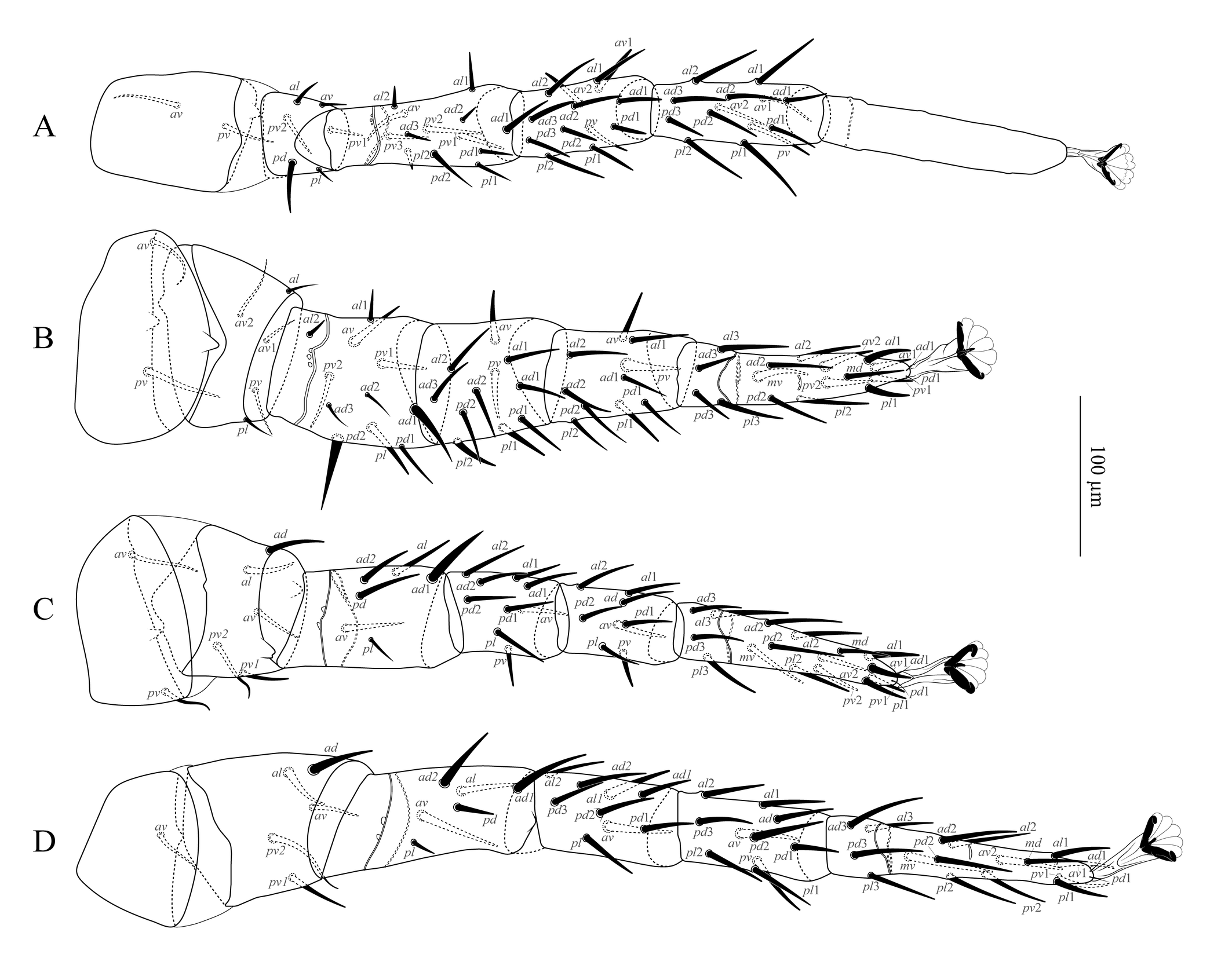
Legs (Figure 5). Chaetotaxy: coxae: I: 0-0/1, 0/1-0; II: 0-0/1, 0/1-0; III: 0-0/1, 0/1-0; IV: 0-0/1, 0/0-0; trochanters: I: 1-0/1, 1/2-1; II: 1-0/2, 0/1-1; III: 1-1/1, 0/2-0; IV: 1-1/1, 0/2-0; femora: I: 2-3/1, 2/3-2; II: 2-3/1, 2/2-1; III: 1-2/1, 1/0-1; IV: 1-2/1, 1/0-1; genua: I: 2-3/2, 3/1-2; II: 2-3/1, 2/1-2; III: 2-2/1, 2/1-1; IV: 2-2/1, 3/0-1; tibiae: I: 2-3/2, 3/1-2; II: 2-2/1, 2/1-2; III: 2-1/1, 2/1-1; IV: 2-1/1, 3/1-2; tarsi: I: not counted; II–IV: 18 setae each (3-3/2, 3/2-3 + mv, md). All setae setiform and smooth. Seta ad1 of femur of all legs, pd of trochanter I, av and pd2 of trochanter II, pv of tibia II, and ad1 and ad2 of femur IV, slightly stouter than other leg setae. Each leg with a pretarsus, including a pair of claws and a pulvillus with a rounded median lobe. Measurements in Table 1.
Adult male (3 specimens from Rio Grande do Sul, 1 specimen from Santa Catarina) – (Figures 6–7)
Gnathosoma. Fixed cheliceral digit (Figure 6A) with two distal teeth in addition to an apical tooth and a setiform pilus dentilis; movable digit with two teeth in addition to an apical tooth, and with a slightly longer spermatodactyl, directed forward, tapered distally, and curved upward. Epistome, apotele, corniculus, deutosternum, gnathosomal setae, and labrum as in the adult female. Measurements in Table 1.
Idiosoma. Oval; unsclerotized cuticle with fingerprint-like pattern.
Dorsum of idiosoma (Figure 6B). Dorsal shield finely punctate throughout, covering about 95% of the idiosoma, with ornamentation similar to that of the adult female. Podonotal region with the same setae as in adult female, four pairs of distinguishable lyrifissures (id1, id2, id4, id5), and three pairs of distinguishable gland pores (gd1, gd2, gd4). Unsclerotized cuticle along lateral margins of the podonotal region with one pair of setae (r6), inserted in a tiny, rounded sclerite. Opisthonotal region with 17 pairs of setae (J1–J5, Z1–Z5, S1–S5, Zx2, Zx3), 2–5 unpaired setae between levels of J2 and J4 (from Rio Grande do Sul: two with three, one with two; from Santa Catarina: one with five), nine pairs of distinguishable lyrifissures (is1, idl1, idl3, idm1, idm2, idm4, idm6, idm9, idx), and three pairs of distinguishable gland pores (gd6, gd8, gd9). Unsclerotized cuticle along lateral margins of the opisthonotal region with seven pairs of setae (R1–R5 and two UR), each inserted in a rounded sclerite; lyrifissures Rp indistinguishable; R2–R5 and UR visible in ventral view. Setae setiform and smooth, except J5 and Z5, lightly barbed. Measurements in Table 1.
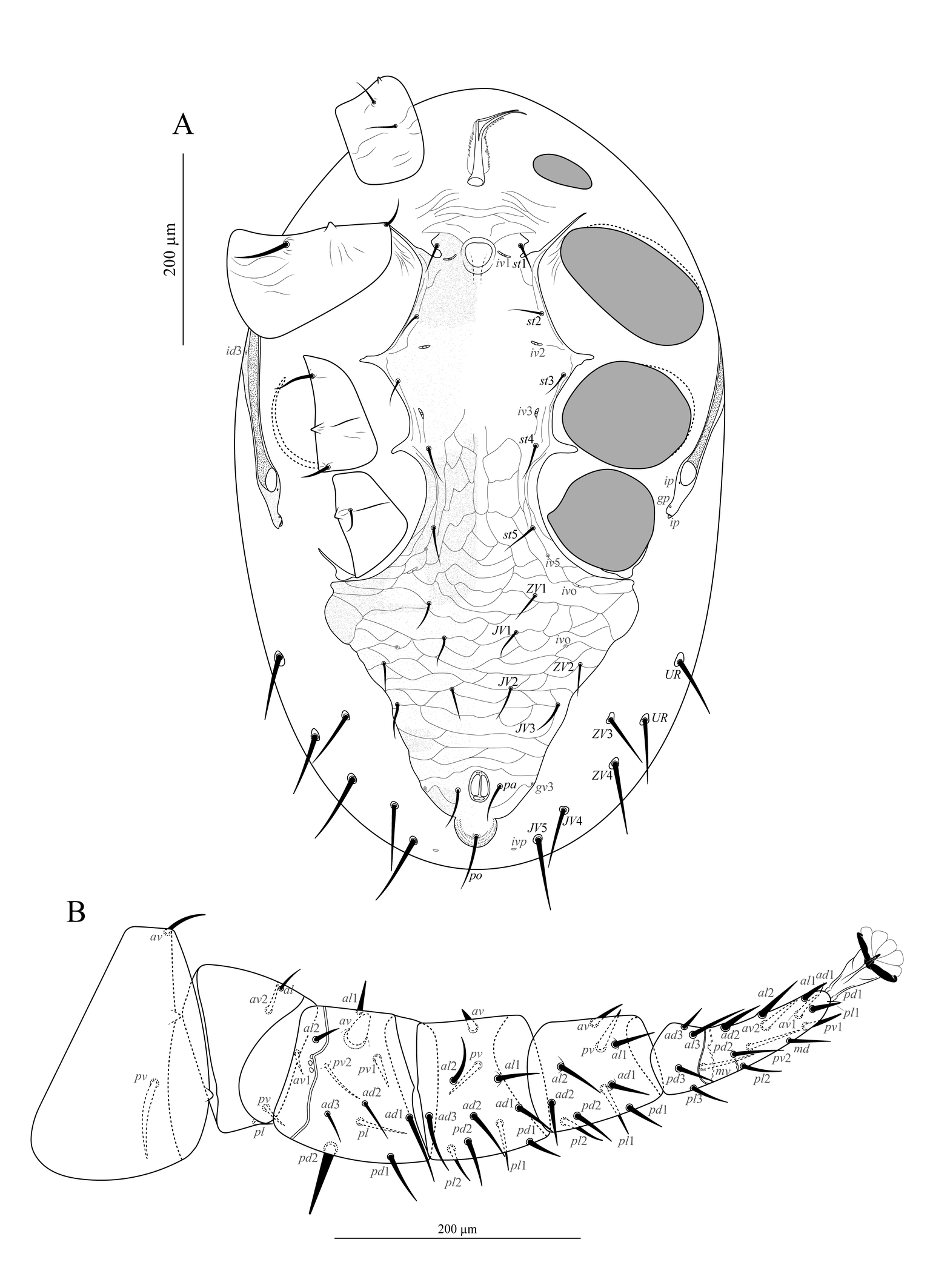
Venter of idiosoma (Figure 7A). Tritosternum and presternal area as in the adult female. Sternal, endopodal, epigynal, and anal shields fused into a holoventral shield; section behind coxa IV subtriangular, with a pair of lateral incisions at mid-level between para-anal and post-anal setae; with ten pairs of setae (st1–st5, JV1–JV3, ZV1, ZV2) in addition to circum-anal setae, six pairs of lyrifissures (iv1, iv2, iv3, iv5, and two ivo), and one pair of pores (gv3); reticulate and lightly punctate, except between st1 and st3, smooth, with scant lateral striae. All setae setiform and smooth. Measurements in Table 1.
Peritreme and peritrematic plate. As in the adult female.
Legs. Chaetotaxy as in the adult female. All setae smooth. Most setae setiform, but av of femur (especially), av and pv of genu and of tibia II, distinctly stouter than other leg setae (Figure 7B); the first distally blunt and others sharp-tipped. Pretarsus as in the adult female. Measurements in Table 1.
Remarks

Small differences between populations were observed in morphometric measurements, the number of unpaired setae on the dorsal shield, the degree of sclerotization of the presternal area, and the degree of concavity of the anterior margin of the anal shield. These differences are considered minor and likely represent intraspecific variation.

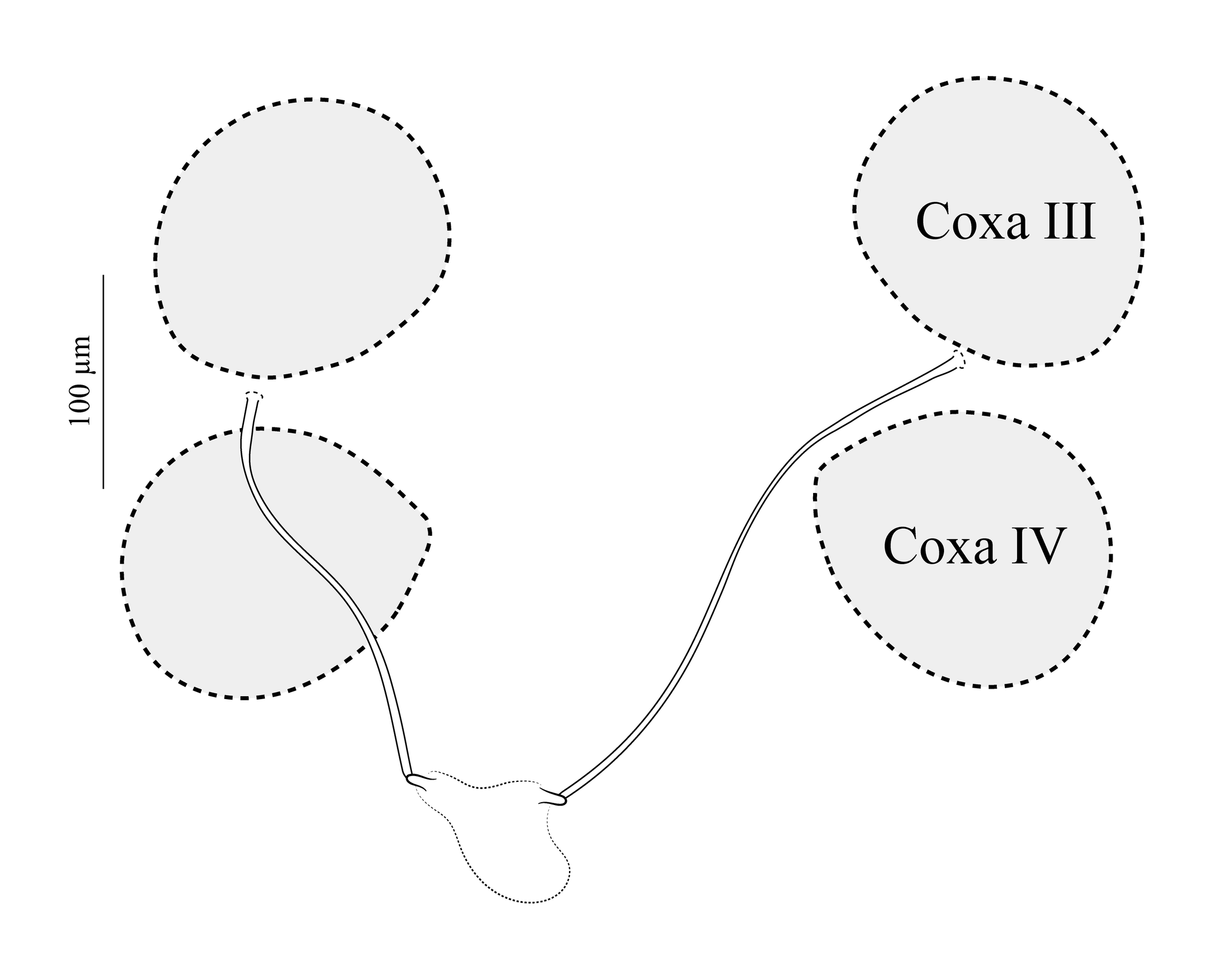
In the morphometric measurements, partial overlap was observed among the ranges of the different populations for several characters (Table 1). The number of unpaired setae on the dorsal shield of females from Santa Catarina varied from four to seven, whereas in females from Rio Grande do Sul and São Paulo, it varied from one to four (Figure 8). The anterior margin of the sternal shield is more strongly sclerotized in females from Rio Grande do Sul and Santa Catarina (with the insertion of st1 clearly visible in the sternal shield), whereas in specimens from São Paulo, this margin was weakly sclerotized (with st1 appearing to be inserted in the unsclerotized cuticle in some specimens) (Figure 9). The shape of the anal shield of specimens from Rio Grande do Sul and São Paulo was similar, with the anterior margin nearly straight medially, whereas in two specimens from Santa Catarina, this margin was more concave (Figure 10). Several female specimens from the three populations were observed to contain fully developed larvae, which suggests that at least some individuals give birth to well-formed post-embryonic progeny that develop within the eggshell.

An effort was made to examine the type specimen of L. (H.) alphabeticus, which is deposited at the Istituto Sperimentale per la Zoologia Agraria, Florence, Italy. At our request, Dr. Sauro Simoni examined the specimen, but due to the long time since mounting, the condition of the preparation did not allow reliable conclusions regarding its morphology.
Apparently, a single specimen of H. alphabeticus was also collected by F. Koenike from a locality reported only as ''southern Brazil″. The slide-mounted specimen belongs to the Oudemans collection, which was subsequently donated to the Rijksmuseum van Natuurlijke Historie in Leiden, the Netherlands (a predecessor of the present-day Naturalis Biodiversity Center). We were able to locate this specimen (RMNH.ACA.P.3535); however, as in the case of the type material, its poor state of preservation precluded a reliable morphological assessment and prevented any conclusions.
Given the relatively informative original description of L. (H.) alphabeticus by Berlese (1904), although brief and apparently based on a single specimen, and its fairly detailed accompanying illustration, together with the morphological uniformity observed among the specimens collected in the three different (albeit neighboring) states in southern Brazil, as well as the apparent dominance of this species in this part of the country in association with stingless bees, it is concluded that all specimens examined belong to the species described by Berlese over 120 years ago as L. (H.) alphabeticus.
Update of the generic placement
This species does not conform to the concept of Hypoaspis presented by Joharchi and Halliday (2011) and Moraes et al. (2022), as it lacks whip-like dorsal setae, bears 2–7 unpaired dorsal setae between J2 and J4, and has setae r2 and r3 positioned on the dorsal shield. Most Hypoaspis sensu stricto species have been described from specimens collected on beetles of the family Scarabaeidae (Coleoptera) (Moraes et al. 2022).
Based on the key and the generic characterization, both provided by Moraes et al. (2022), the species re-characterized in this study was determined to be most similar to species of the following laelapid genera that include species commonly reported in association with Neotropical eusocial Hymenoptera (bees and ants): Bisternalis, Holostaspis Kolenati, 1858, Hunteria, Melittiphisoides and Myrmozercon Berlese, 1902. They share the following morphological characteristics:
Also, based on Moraes et al. (2022), the following key can be used to separate these genera:
1. Sternal shield unusually broad anteriorly, expanding between coxae I and II, and bearing three pairs of setae (st1–st3); arthrodial process at the base of the movable digit unusually large and setiform
...... Hunteria
— Sternal shield not expanded between coxae I and II, and either bearing three pairs of setae (st1–st3), or with st1 inserted on a platelet partly or entirely separate from the shield, or st1 inserted in the presternal unsclerotized area; arthrodial process not particularly developed
...... 2
2. Epigynal shield with a distinctive wedge-shaped median section, with posterior margin nearly truncate, and abutting the anal shield
...... Melittiphisoides
— Epigynal shield snake-head-shaped, typically acuminate posteriorly (sometimes blunt or bilobed), either reaching the anal shield or not
...... 3
3. Sternal shield bearing st1–st3, with a well-sclerotized anterior margin; peritreme extending anteriorly at most to the mid-level of coxa II
...... Myrmozercon
— Sternal shield bearing st1–st3, or with st1 inserted on a platelet partly or entirely separate from the shield, or st1 inserted in the presternal unsclerotized area; peritreme extending anteriorly beyond coxa II
...... 4
4. Cheliceral digits short and robust (17–30 µm long); movable cheliceral digit usually edentate (rarely bearing 1–2 small teeth); anal shield longer than wide
...... Holostaspis
— Cheliceral digits elongate (at least 50 µm long); movable cheliceral digit bearing two teeth; anal shield wider than long
...... Bisternalis
Based on the key provided above, the species re-characterized in this study was determined to belong to Bisternalis. The diagnosis of this genus provided by Moraes et al. (2022) seems rather appropriate to accommodate L. (H.) alphabeticus. Specimens of this species exhibit key diagnostic features consistent with those attributed to that genus, including the partial detachment of setae st1 from the sternal shield and anal shield with a distinctly concave anterior margin. We therefore transfer this species to Bisternalis as Bisternalis alphabeticus (Berlese) n. comb. and revise the generic characterization as subsequently detailed.
While maintaining the taxonomic decisions regarding species originally assigned to Bisternalis, Moraes et al. (2022) summarized the diagnostic characteristics of this genus. In that account, the genus was already considered to accommodate certain morphological variations, primarily concerning: (a) the insertion of st1 relative to the sternal shield, (b) the shape of the posterior margin of the sternal shield; (c) the shape of the posterior end of the epigynal shield; and (d) the shape of the anterior margin of the anal shield. Among these variations, the subdivision of the sternal shield, reflected in the generic name (Bisternalis), is not consistently expressed across all species currently included in this genus, notably B. camargoi Baker, Flechtmann & Delfinado-Baker, 1984 and B. mexicanus Baker, Delfinado-Baker & Ordaz, 1983. The posterior margin of that shield is deeply concave in most species, but not in B. alphabeticus and B. camargoi.
As for the epigynal shield, although it consistently exhibits a snake-head shape, the configuration of its posterior margin varies from blunt in B. alphabeticus and B. hunteri Baker, Delfinado-Baker & Ordaz, 1983; bilobed in B. rettenmeyeri Hunter, 1963; to acuminate in B. camargoi, B. formosus Baker, Flechtmann & Delfinado-Baker, 1984, B. mexicanus and B. trigonarum Baker, Flechtmann & Delfinado-Baker, 1984. The posterior extent of the epigynal shield is rather variable, ranging from nearly reaching the anal shield to being considerably shorter (in B. alphabeticus and B. camargoi).
Bisternalis alphabeticus differs from other species of this genus in usually bearing more than two unpaired dorsal setae, in having the posterior margin of the sternal shield slightly concave (also observed in B. camargoi), and in having the parapodal plate not particularly enlarged. Nevertheless, a few specimens from the Rio Grande do Sul and São Paulo populations were observed to bear only one unpaired dorsal seta, indicating some degree of intraspecific variation in this character.
Unlike B. camargoi, in which st1 is inserted in the unsclerotized area anteriad sternal shield (presternal region), B. alphabeticus bears these setae on the anterior, transversely elongate platelet that is partially separate from the sternal shield by a pair of lateral incisions. This feature also contrasts with B. formosus and B. rettenmeyeri, in which both st1 setae are located on a platelet completely separate from the sternal shield.
Key to the world species of Bisternalis based on adult females
The following key can be used to separate the species now placed in Bisternalis:
1. Setae st1 in presternal unsclerotized area anteriad sternal shield
...... B. camargoi – Brazil, in nest of Lestrimellita limao (Hymenoptera: Apidae)
— Setae st1 in a platelet, partly or entirely separate from the sternal shield
...... 2
2. Setae st1 in a platelet entirely separate from the sternal shield
...... 3
— Setae st1 in a platelet, partly separate from the sternal shield by a pair of incisions in the lateral margins of the sternal shield
...... 4
3. Epigynal shield bilobed posteriorly; anal shield crescentic
...... B. rettenmeyeri – Panama, on Trigona sp. (Hymenoptera: Apidae) and its nests
— Epigynal shield acuminate posteriorly; anal shield bowl-shaped
...... B. formosus –Brazil, in nest of Trigona fulviventris (Hymenoptera: Apidae)
4. Posterior section of epigynal shield extending laterally beyond the middle of coxa IV
...... 5
— Posterior section of epigynal shield not extending beyond the middle of coxa IV
...... 6
5. Epigynal shield blunt posteriorly; unsclerotized opisthogastric cuticle with setae about 25–35 µm long
...... B. hunteri – Panama, in wasp (Hymenoptera) nest
— Epigynal shield acuminate posteriorly; unsclerotized opisthogastric cuticle with setae about 90–100 µm long
...... B. mexicanus – Mexico, in nest of Melipona sp. (Hymenoptera: Apidae)
6. Sternal shield with posterior margin strongly concave; epigynal shield acuminate posteriorly, with V-shaped striae; parapodal plates enlarged
...... B. trigonarum – Brazil, in nest of Trigona chanchamayoensis (Hymenoptera: Apidae)
— Sternal shield with posterior margin slightly concave; epigynal shield blunt posteriorly, with U-shaped striae; parapodal plates narrow
...... B. alphabeticus – Brazil, in nest of Melipona mandacaia (Hymenoptera: Apidae)
Other incertae sedis species originally assigned to Hypoaspis
Other species also associated with stingless bees, originally placed in Hypoaspis and treated as incertae sedis by Moraes et al. (2022) are subsequently indicated.
Hypoaspis meliponarum Vitzthum, 1930 was described from Colombia, collected in the nest of Melipona interrupta Latreille (Hymenoptera: Apidae). Published information about the morphology of this species, provided in the original description (Vitzthum, 1930) and in the redescription of Baker et al. (1983: 10), does not allow its reliable distinction from B. alphabeticus. The apparent absence of certain setae on the posterior section of the podonotal region could be due to the loss of setae in the mounting process. Discrepancies in the literature regarding the insertion of st1, reported as occurring in an unsclerotized surface in the original description and in a sclerotized surface by Baker et al. (1983), may represent intraspecific variation.
The description of H. meliponarum was apparently based on two specimens exhibiting very similar morphological characteristics. According to the original description, the only discernible difference between them was the size of the idiosoma. One specimen (from Sevilla), with idiosoma approximately 910 µm long and 590 wide, was comparable (in relation to size) to that reported for B. alphabeticus in the original description (and in the present study), whereas the other specimen (from Rio Frio), measuring about 620 µm in length and 460 in width, was considerably smaller.
Contacts were established with the Zoologische Staatssammlung München and Senckenberg Museum für Naturkunde Görlitz, Germany, considered the most probable depositories for the H. meliponarum types, with the assistance of Dr. Diana Rueda, in an effort to locate the type specimens. Unfortunately, these could not be found. Additional attempts were made to obtain fresh material from northern Colombia, with the support of local collaborators, but these efforts were also unsuccessful. Thus, although available evidence suggests that the two nominal species may be conspecific, we refrain from formally synonymizing them until specimens can be collected from the type locality for examination.
Hypoaspis hoffmannae Smiley, Baker & Delfinado-Baker, 1996 was described from specimens collected in nests of Trigona iridipennis (Smith) (Hymenoptera: Apidae) in Malaysia. This species resembles Bisternalis in having a wide, concave anal shield, a snake-head-shaped epigynal shield, and one unpaired dorsal seta. As in B. camargoi, it was described as having the st1 setae in the unsclerotized cuticle anteriad the sternal shield, whose posterior margin is not deeply concave, although this was illustrated but not explicitly mentioned in the description. However, it apparently lacks expanded parapodal plates, its st3 setae are apparently barbed, and it seems to have an edentate movable cheliceral digit; these features are not typical of Bisternalis.
Hypoaspis favosus Turk, 1948 was described from Trinidad in association with the stingless bee Melipona favosa favosa (Fabricius) (Apidae). Both st1 setae of this species are inserted in a single platelet anteriad the sternal shield, as in B. formosus and B. rettenmeyeri. However, other similarities to Bisternalis are scarce; its epigynal shield is flask-shaped and large, abutting the subtriangular anal shield, which is neither wide nor provided with a concave anterior margin.
Klimov et al. (2016) report an undescribed species that they provisionally assigned to Hypoaspis. As in Bisternalis, this species has a wide anal shield with a deeply concave anterior margin, and apparently two unpaired dorsal setae. However, its st1 setae are inserted in the sternal shield, its epigynal shield is narrow and tongue-shaped, and the parapodal plate is discreet. Both H. hoffmannae and the undescribed species have large pilus dentilis and a single tooth in each row of the deutosternal groove. Based on these features, Klimov et al. (2016) suggested that it might refer to a new genus.
Accordingly, we consider it more prudent to postpone any decision regarding the possible placement of these species within Bisternalis or the establishment of a new genus or genera for the time being.
Acknowledgements
To Gilberto J. de Moraes, for his invaluable cooperation during the conduction of the whole work. To Bernardo Telles de Souza, Prof. Alex Sandro Poltronieri, Prof. Denise Araujo Alves, Prof. Noeli Juarez Ferla, and Dr. Cristiano Menezes for providing the specimens studied in this work. To Dr. Sauro Simoni for locating and photographing the type material of H. alphabeticus. To Mónica Guimarães Cruz for locating and photographing Oudemans' specimen. To Dr. Diana Marcela Rueda-Ramírez for the attempt to locate the type material of H. meliponarum. This study was financed, in part, by the São Paulo Research Foundation (FAPESP), Brazil, Process Number 2024/21481-1 (scholarship provided to Lucas L. Godoy) and Process Number 2017/12004-1 (part of BIOTA-FAPESP program), and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Brazil (Process Number 309433/2025-8 – CNPq researcher of the third author).
References
- Athias-Henriot C. 1971. La divergence néotaxique des Gamasides (Arachnides). Bull. Sci. Bourgogne, 28: 93-106.
- Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini. II-Le relevé organotaxique de la face dorsale adulte (Gamasides Protoadeniques, Phytoseiidae). Acarologia, 17: 20-29.
- Baker E.W., Delfinado-Baker M., Reyes Ordaz F. 1983. Some laelapid mites (Laelapidae: Mesostigmata) found in nests of wasps and stingless bees. Int. J. Acarol., 9: 3-10. doi:10.1080/01647958308683305 https://doi.org/10.1080/01647958308683305
- Baker E.W., Flechtmann C.H.W., Delfinado-Baker M. 1984. Acari domum meliponinarum brasiliensium habitantes. VI. New species of Bisternalis Hunter (Laelapidae: Acari). Int. J. Acarol., 10: 181-189. doi:10.1080/01647958408683373 https://doi.org/10.1080/01647958408683373
- Berlese A. 1904. Acari nuovi. Manipulus III. Redia, 2: 10-32 + 2 plates.
- Buitendijk A.M. 1945. Voorloopige catalogus van de Acari in de collectie-Oudemans. Zool. Meded., 24: 281-391.
- Castagnoli M., Pegazzano F. 1985. Catalogue of the Berlese Acaroteca. Istituto Sperimentale per la Zoologia Agraria, Firenze, 490 pp.
- Da-Costa T., Santos C.F., Rodighero L.F., Ferla N.J., Blochtein B. 2021. Mite diversity is determined by the stingless bee host species. Apidologie, 52: 950-959. doi:10.1007/s13592-021-00878-2 https://doi.org/10.1007/s13592-021-00878-2
- Eickwort G.C. 1990. Associations of mites with social insects. Annu. Rev. Entomol., 35: 469-488. doi:10.1146/annurev.en.35.010190.002345 https://doi.org/10.1146/annurev.en.35.010190.002345
- Evans G.O. 1963a. Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bull. Br. Mus. Nat. Hist. Zool., 10: 275-303. https://doi.org/10.5962/bhl.part.20528
- Evans G.O. 1963b. Some observations on the chaetotaxy of the pedipalps in the Mesostigmata (Acari). Ann. Mag. Nat. Hist. Ser. 13, 6: 513-527. doi:10.1080/00222936308651393 https://doi.org/10.1080/00222936308651393
- Hunter P.E. 1963. Two new genera of mites associated with stingless bees (Acarina, Laelaptidae). Acarologia, 5: 5-12.
- Joharchi O., Halliday B. 2011. New species and new records of mites of the family Laelapidae (Acari: Mesostigmata) associated with Coleoptera in Iran. Zootaxa, 2883: 23-38. doi:10.11646/zootaxa.2883.1.2 https://doi.org/10.11646/zootaxa.2883.1.2
- Klimov P.B., OConnor B., Ochoa R., Bauchan G.R., Redford A.J., Scher J. 2016. Bee Mite ID: Bee-Associated Mite Genera of the World [Internet]. Fort Collins: USDA APHIS Identification Technology Program (ITP); [27 March 2026]. Available from: https://idtools.org/id/mites/beemites
- Klompen H., Campbell K.U. 2025. Myrmecophily among mesostigmatid mites. Insectes Soc., 72: 285-302. doi:10.1007/s00040-024-01003-z https://doi.org/10.1007/s00040-024-01003-z
- Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Mem. Entomol. Soc. Can., 47: 5-66. doi:10.4039/entm9747fv https://doi.org/10.4039/entm9747fv
- Lindquist E.E., Krantz G.W., Walter D.E. 2009. Order Mesostigmata. In: Krantz G.W., Walter D.E. (Eds.). A Manual of Acarology. Third Edition. Texas Tech University Press, Lubbock, pp. 124-232.
- Moraes G.J., Moreira G.F., Freire R.A.P., Beaulieu F., Klompen H., Halliday B. 2022. Catalogue of the free-living and arthropod-associated Laelapidae Canestrini (Acari: Mesostigmata), with revised generic concepts and a key to genera. Zootaxa, 5184: 1-509. doi:10.11646/zootaxa.5184.1.1 https://doi.org/10.11646/zootaxa.5184.1.1
- Smiley R.L., Baker E.W., Delfinado-Baker M. 1996. New species of Hypoaspis (Acari: Mesostigmata: Laelapidae) from the nest of a stingless bee in Malaysia (Hymenoptera: Meliponinae: Apidae). An. Inst. Biol. Univ. Nac. Autón. México Ser. Zool., 67: 197-203.
- Tipton V.J. 1960. The genus Laelaps with a review of the Laelaptinae and a new subfamily Alphalaelaptinae (Acarina: Laelaptidae). Univ. Calif. Publ. Entomol., 16: 233-356 + 26 plates.
- Turk F.A. 1948. Insecticolous Acari from Trinidad, B.W.I. Proc. Zool. Soc. Lond., 118: 82-125. doi:10.1111/j.1096-3642.1948.tb00367.x https://doi.org/10.1111/j.1096-3642.1948.tb00367.x
- Vitzthum H.V. 1930. Milben als Pestträger? Ein Beitrag zu den Untersuchungen der mandschurischen Peststudienkommission in Harbin. Der `Acarologischen Beobachtungen'. 16. Reihe. Zool. Jahrb. Abt. Syst. Geogr. Biol. Tiere, 60: 381-428.



2026-04-24
Date accepted:
2026-06-22
Date published:
2026-06-26
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Godoy, Lucas Lorena; Da-Costa, Tairis and Castilho, Raphael de Campos
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)