



Trachycilliba dominicana n. sp., the first confirmed representative of genus Trachycilliba in the Neotropical realm (Acari: Mesostigmata: Uropodidae)
Kontschán, Jenő  1
and Ermilov, Sergey G.
2
1
and Ermilov, Sergey G.
2
1✉ Plant Protection Institute, HUN-REN Centre for Agricultural Research, H-1525 Budapest, P.O. Box 102, Hungary & Department of Plant Sciences, Albert Kázmér Faculty of Mosonmagyaróvár, Széchenyi István University, Vár square 2., H-9200 Mosonmagyaróvár, Hungary.
2Institute of Environmental and Agricultural Biology (X-BIO), University of Tyumen, Lenina str. 25, 625000 Tyumen, Russia.
2026 - Volume: 66 Issue: 2 pages: 579-587
https://doi.org/10.24349/8puf-bftwZooBank LSID: 8082B03B-2851-45D9-978E-48CB03489431
Original research
Keywords
Abstract
Introduction
The Dominican Republic shared territory with Haiti on the island of Hispaniola in the Caribbean. The majority of the countries of the Caribbean Islands are scarcely investigated regions of the Neotropics, with very few species described and named (see Wiśniewski 1993). Only five rotundabaloghiid mites have been described from the Dominican Republic (Kontschán 2005), so this country is considered unknown from the Uropodina point of view.
The genus Trachycilliba was erected by Berlese (1903) for the species Uropoda splendida Kramer, 1882. Afterward, many researchers treated species of this genus under another genus name (such as Neodiscopoma (Vitzthum, 1942) or Uropoda (Wiśniewski & Hirschmann, 1993)) or as a species group (such as the Uropoda splendida-group (Wiśniewski & Hirschmann, 1993)). Kazemi and Kontschán (2025) redefined this genus and characterised by an elevated central area on the dorsal shield with strongly sclerotized margins and lateral incisions, and with eight post-dorsal platelets (Kazemi and Kontschán 2025).
The majority of known Trachycilliba species are known from East Asia (19 species in Japan and South Korea); seven are from the Oriental (Malaysia, Indonesia, Vietnam) and Australasian (New Guinea) realms. Six species are listed from Europe and the Middle East, two species from Africa and one from North America (Kazemi and Kontschán 2025). Up to date, there are no representatives of this genus in the Neotropical region.
Neodiscopoma, the junior synonym of Trachycilliba (Kazemi and Kontschán 2025), has caused several confusions in the identification of their species. Earlier, Marais and Theron (1986) presented some species from South Africa under the name Neodiscopoma. These species differed from the Trachycilliba species in morphology; therefore, Wiśniewski and Hirschmann (1993) placed them in a different species group (Uropoda difoveolata-species group) and earlier, erected a new genus for these species (Foveolaturopoda Hirschmann, 1979), but only the type species were transferred to this new genus. Therefore, later, Kontschán (2024) clarified this genus and transferred the other species into it.
The genus Foveolaturopoda is widely distributed across the Neotropical realm, with many species known from Cuba to Chile (Kontschán 2024). So, we need to assume that these Neodiscopoma species, which were presented without species identification and with no illustration or description in Salazar Martinez (1996) and Deleva et al. (2023), belong to the genus Foveolaturopoda and not Trachycilliba.
During the investigation of Uropodina mite specimens from the Natural History Museum, Geneva (Switzerland), we found an unusual Trachycilliba species in soil samples from the Dominican Republic, which is the first conformed representative of this genus in the Neotropical realm.
Material and methods
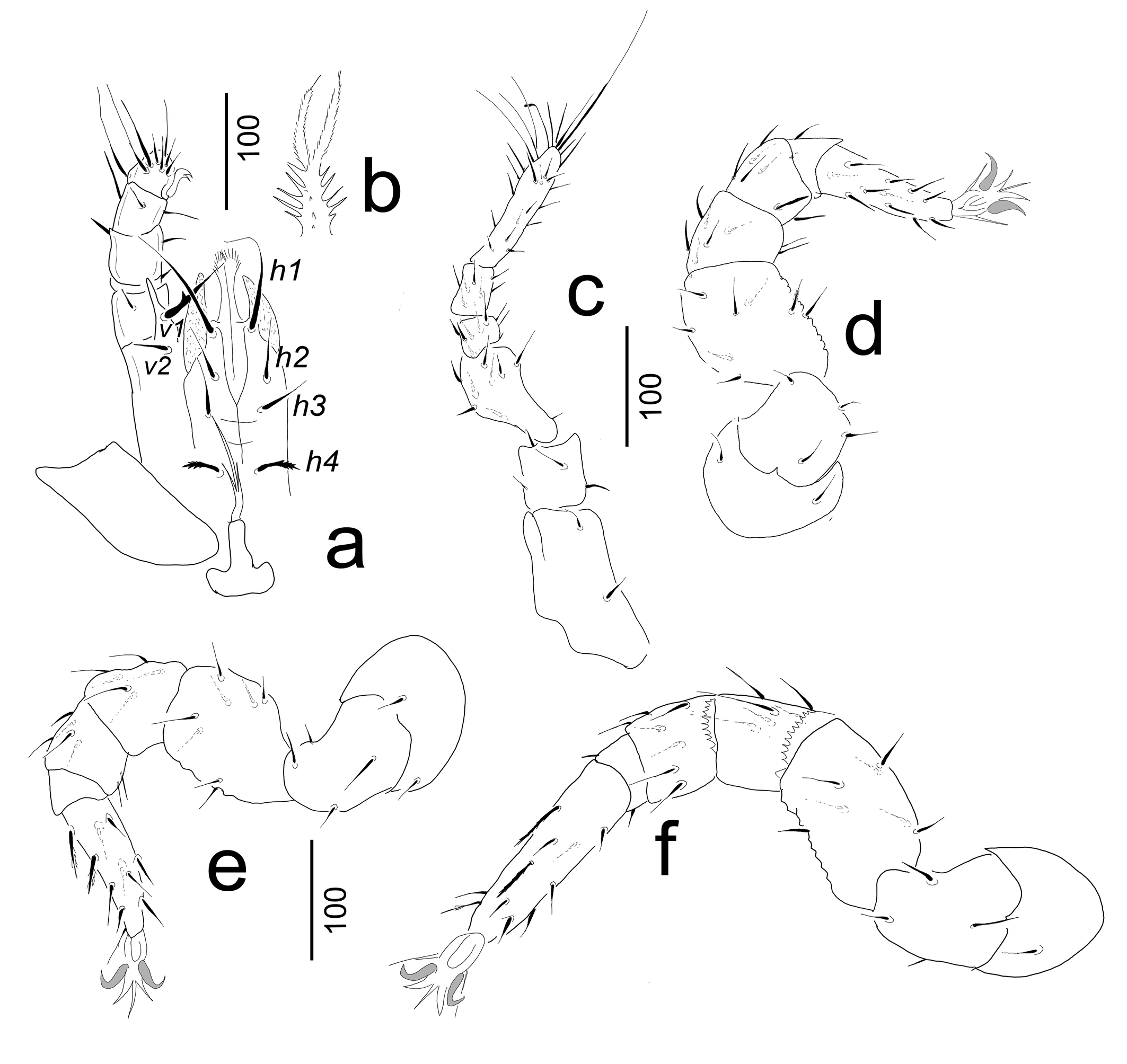
Specimens examined were cleared in lactic acid for a week and afterwards were investigated in a half-covered cavity slide with a Leica 1000 scientific microscope with a drawing tube. The pencil drawings were clarified with black ink and after they were scanned by a Canon scanner. Photographs were taken with a Keyence 5000 digital microscope. Specimens examined are stored in 70% ethanol and deposited in the Natural History Museum, Geneva (NHMG). Measurements are given in micrometers (μm). Abbreviations: h = hypostomal setae, st = sternal setae, lf = lyriform fissure, p = pores, ad = adanal setae.
Taxonomy
Family Uropodidae Kramer, 1881
Genus Trachycilliba Berlese, 1903
Trachycilliba dominicana n. sp.
ZOOBANK: 03394C3E-2923-42C6-82AF-48A0E398C632 ![]()
(Figures 1–7)
Diagnosis
Elevated part of dorsal shield divided into central part with a transversal furrow. One pair of ring-like strongly sclerotized structures situated between strongly sclerotized margins of anterior and posterior part of elevated dorsal areas. Eight post-dorsal platelets with eight post-dorsal setae absent.
Material examined
Holotype — Female. Dominican Republic, Colonia Cordillera Central, 100 m 1. May 1974, J. and S. Klappenich coll. Paratypes. Six females, eight males and eight deutonymph, with same collection data as those for the holotype.
Description
Female — (n=7). Idiosoma reddish-brown, 1135–1140 long and 885–894 wide, its shape oval with rounded posterior margin

Dorsal idiosoma (Figures 1, 7a–b). Dorsal and marginal shields completely separated.
Central area of dorsal shield elevated from neighbouring areas. Margins of elevated area strongly sclerotized. Elevated area divided into a larger anterior (460–470 long) part and a smaller posterior (250–255 long) part with a transversal furrow on central part. One pair of ring-like (130–135 long and 142–147 wide) strongly sclerotized structure connect anterior and posterior parts of elevated area on their lateral parts. One pair of lateral incision (80–85 long) situated on outer margins of ring-like structures. Seven pairs of smooth and needle-like setae (ca 60–65 long) situated on margins of dorsal shield and 14 pairs of long (ca 100–108) and smooth setae on central area of dorsal shield. One pair of pilose setae (ca 65–70 in length) inserted close to caudal margin of dorsal shield. Majority of surface of dorsal shield smooth, with some oval pits (ca 13–14 long and 14–15 wide) on caudal area and some muscle scars present. One pair of lyriform fissure situated on caudal part of elevated area.
Marginal shield wide (145–150), its surface with irregular pits and bearing seven-eight pairs of ca 92–105 long and needle-like setae. Margins of idiosoma with several short (ca 55–60) and curved setae. Caudal part of dorsal idiosoma with membranous cuticle without any platelets or setae.
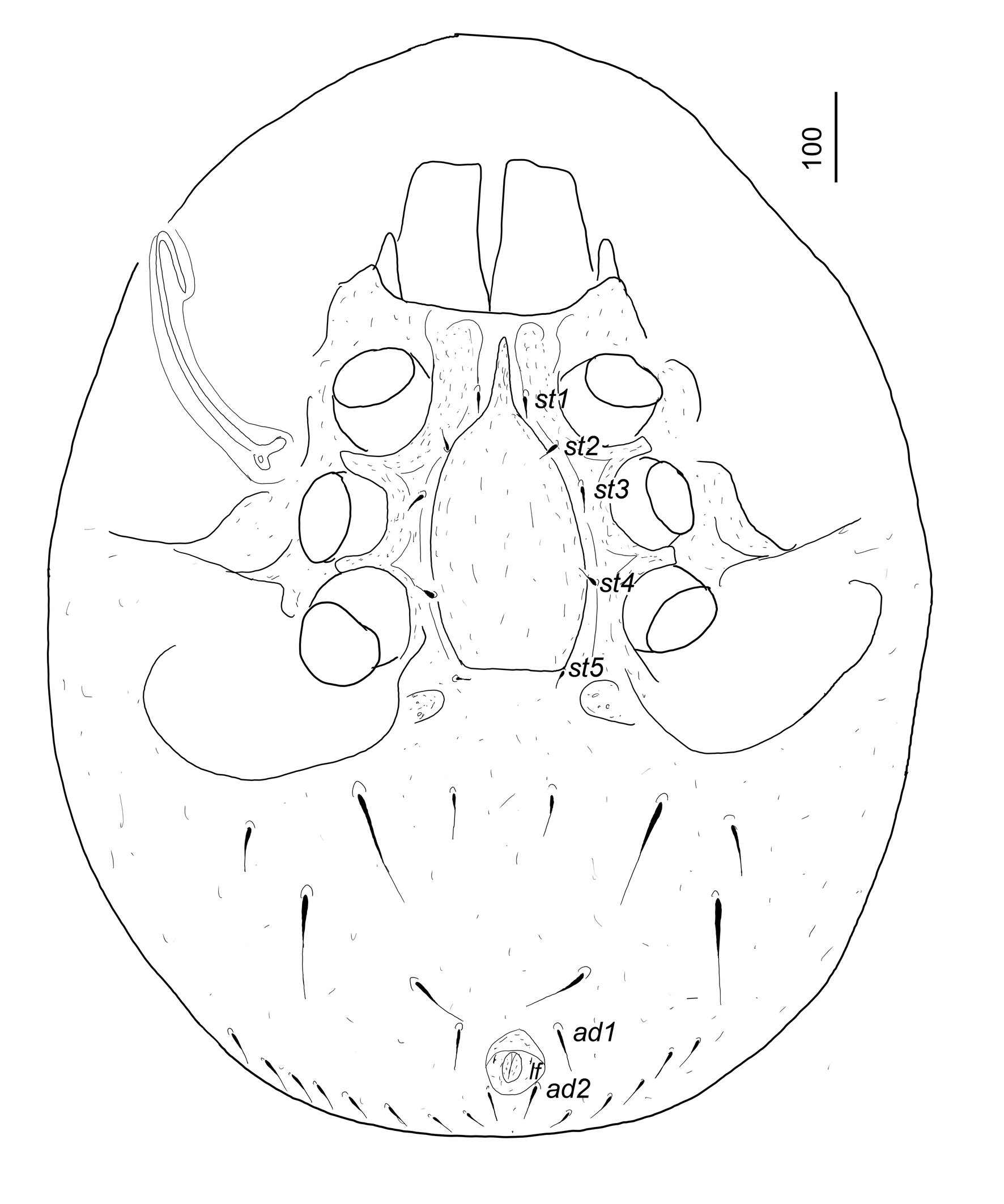
Ventral idiosoma (Figures 2, 3a and 7c). Surface of sternal shield without sculptural pattern. Five pairs of sternal setae smooth, needle-like and ca 23–30 long. Setae st1 situated at level of midcoxae II, st2 at level of posterior margin of coxae II, st3 at level of anterior margin of coxae III, st4 at level of anterior margin of coxae IV, st5 close to basal edges of genital shield. Surface of ventral shield without sculptural pattern and bearing two pairs of smooth and long (ca 122–127) and three pairs of smooth and short (ca 54–72) ventral setae anterior to anal opening. Adanal setae smooth and needle-like (ca 45–55 long), anal opening small (ca 30–32 long and ca 23–25 wide). Several short and needle-like setae situated close to posterior margin of ventral shield.
Genital shield large (ca 289–295×170–172), linguliform with a long (ca 75–79) apical prolongation. Surface without sculptural pattern. Stigmata situated between coxae II and III; prestigmatic part of peritremes long and hook-shaped, poststigmatic part short and straight. Leg grooves relatively deeper, with smooth surface; separated furrows for tarsi I absent. Base of tritosternum ca 62–64 long, basally wide (ca 52–54) and apically narrow (ca 14–16). Tritosternal laciniae trifurcated, central branch ca 92–97 long, lateral branches shorter (ca 45–47) (Figure 3a).
Gnathosoma (Figure 3a–b). Corniculi horn-like, internal malae apically pilose and longer than corniculi. Hypostomal setae h1–h3 smooth, h4 with serrate margin. Setae h1 very long (ca 100–103 in length) other setae shorter (ca 35–50). Palp trochanter with a short v2 (ca 28–30) and long (ca 67–70) v1 setae. Setae v1 situated on a large prolongation and bearing a lateral tooth. Other setae on palp smooth and needle-like. Base of epistome serrate, apically divided into two pilose branches (Figure 3b). Chelicerae not visible.
Legs (Figures 3c–f). Legs length: I 515–522, II 442–448, III 478–484, IV 600–615. Legs I without tarsal claws. Leg setae smooth and needle-like.
Male — (n=8). Body 1127–1136 long and 860–882 wide.

Dorsal idiosoma. As in adult female.
Ventral idiosoma (Figure 4a). Intercoxal area, with sternal setae and genital opening, as in Figure 4a. All sternal setae smooth and needle-like (ca 17–20 long), its position presented on Figure 4a. Surface of sternal shield without sculptural pattern, one long horizontal furrow (ca 153–156 long and 22–26 wide) situated on central part. Genital opening oval (ca 92–98 long and 77–80 wide), situated between coxae IV and bearing one pair of long (ca 85–88) and smooth eugenital setae.
Legs. Femora of leg II with a large spine-like seta (Figure 4b). Other characters as in adult female.
Deutonymph female — (n=8). Idiosoma yellowish-brown, 960–965 long and 766–772 wide, its shape oval with rounded posterior margin

Dorsal idiosoma (Figures 5, 7d–e). Dorsal and marginal shields completely separated. Dorsal shield (ca 682–690 long and ca 770–774 wide) bearing more than 30 pairs of smooth and needle-like setae (ca 37–40 in length) and one pair of pilose caudal setae (ca 46–48). Surface of dorsal shield smooth, only some muscle scars present. Marginal shield wide (ca 102–107), without sculptural pattern. Setae on margins smooth, curved and short (ca 12–15 long).

Ventral idiosoma (Figures 6 and 7f). Majority of Surface of sternal shield without sculptural pattern, only some oval pits situated between coxae IV. Five pairs of sternal setae smooth, needle-like and ca 21–26 long. Setae st1 situated at level of midcoxae II, st2 at level of posterior margin of coxae II, st3 at level of midcoxae III, st4 at level of anterior margin of coxae IV, st5 posteromesad coxae IV. Surface of ventral shield without sculptural pattern and bearing two pairs of smooth and longer (ca 73–76) and three pairs of smooth and shorter (ca 44–53) ventral setae. Anal opening wider (44–46) than long (20–24).
Stigmata situated at level of coxae II. Peritremes straight, prestigmatic part three times longer than poststigmatic part, short and straight. Leg grooves relatively deeper, with smooth surface; separated furrows for tarsi I absent.

Protonymph and larvae — Unknown.
Etymology
The name of the new species refers to the country where type material of the new species were collected.
Remarks
The new species differs from the congers in the following characters: specifically shaped, elevated central area on the dorsal shield and the absence of the eight post-dorsal platelets associated with the caudal setae, which are unique within the genus Trachycilliba. Only one species has been reported from American continents, T. permagna (Hiramatsu and Hirschmann, 1979). The new species differs from T. permagna in the latter mentioned characters and in presence of the long anterior process of female genital shield, which are observed in the new species, but it is missing in the known one.
The aforementioned character, the absence of the eight post-dorsal platelets associated with the caudal setae, is unique within the genus Trachycilliba but is one of its characteristics. However, it is missing in the new species; we are sure, based on the dorsal sclerotization, that this species belongs to the genus Trachycilliba.
Zoogeographical note
Overall, Trachycilliba species are reported from Africa, Europe, Asia and North America. Till today, no species has been recorded from Central and South America. Based on the morphology of the new species, we hypothesized that it belongs to a distinct lineage within the genus that evolved independently of the African and Eurasian lineages.
Acknowledgements
We are very grateful to Dr. Peter Schwendinger (MHNG) for his kind hospitality during the first author's visit to Geneva.
References
- Berlese A. 1903. Acari nuovi. Manipulus I.us. Redia, 1: 235-252.
- Deleva S., Ulloa A., Oliveira H.F.M., Simov N., Didonna F., Chaverri G. 2023. Cave-dwelling fauna of Costa Rica: current state of knowledge and future research perspectives. Subterr. Biol., 47: 29-62. https://doi.org/10.3897/subtbiol.47.113219
- Hirschmann W. 1979. Stadiensystematik der Parasitiformes. Teil 1. Stadienfamilien und Stadiengattungen der Atrichopygidiina, erstellt im Vergleich zum Gangsystem Hirschmann 1979. Acarologie., 26: 57-70.
- Hiramatsu N., Hirschmann W. 1979. Gangsystematik der Parasitiformes. Teil 307. Teilgänge, Stadien von 10 neuen Uropoda (Phaulodinychus)-Arten aus Kalifornien, Ekuador, Peru, Mexiko und Panama (Uropodini, Uropodinae). Acarologie., 25: 16-26.
- Kazemi S., Kontschán J. 2025. Redescription of Trachycilliba persica (Kazemi & Kontschán), notes on the genus Trachycilliba Berlese (Acari: Mesostigmata: Uropodidae), and a key to world species of the genus. Syst. Appl. Acarol., 30(4): 734-753. https://doi.org/10.11158/saa.30.4.7
- Kontschán J. 2005. New Rotundabaloghia Hirschmann, 1975 species (Acari: Mesostigmata: Uropodina) from Dominican Republic (Caribbean Islands). Annls. hist-nat. Mus. natn. Hung., 97: 241-249.
- Kontschán J. 2024. Uropodina genera of the World. Budapest: HUN-REN Agrártudományi Kutatóközpont, p. 94-95.
- Kramer P. 1881. Ueber die Prinzipien der Classification bei den Gamasiden. Zeitschr Ges Naturwiss., 54: 638-642.
- Kramer P. 1882. Ueber, Gamasiden. Arch Naturgesch., 48: 374-434.
- Marais J.F., Theron P.D. 1986. Four new species of Neodiscopoma Vitzthum, 1941 (Acari, Mesostigmata) from Southern-Africa. Acarologia, 27(3): 211-222.
- Salaraz Martinez A. 1996. Acarofauna asociada a la Hijarasca monoespecifica de Puemo y Bold. Rev. Soc. Entomol. Arg., 55(1-4): 85-93.
- Vitzthum H. 1940-1943. Acarina. In: Bronn H.G. (Ed). Klassen und Ordnungen des Terreichs. 5. Band: Arthropoda. IV. Abteilung: Arachnoidea. 5. Buch. Acarina. Leipzig: Akademische Verlagsgesellschaft Becker & Erler Kom.-Ges., pp. 1011.
- Wiśniewski J. 1993. Gangsystematik der Parasitiformes 549. Der Uropodiden der Erde nach Zoogeographischen Regionen und Subregionen geordnet (Mit Angabe der Länder). Katalog der Ganggattungen, Untergattungen, Gruppe und Arten der Uropodiden der Erde. Acarologie., 40: 221-291.
- Wiśniewski J., Hirschmann W. 1993. Gangsystematik der Parasitiformes 548.katalog der Gattungen, Untergattungen, Gruppen und Arten der Uropodiden der Erde. Acarologie., 40: 1-220.



2026-04-20
Date accepted:
2026-06-09
Date published:
2026-06-15
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Kontschán, Jenő and Ermilov, Sergey G.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)