



A new feather mite Microgruolichus armogaster gen. n., sp. n. (Acariformes: Pterolichidae) from the Common Crane Grus grus (Gruiformes: Gruidae)
Mironov, Sergey V.  1
1
1✉ Zoological Institute, Russian Academy of Sciences, Universitetskaya Embankment 1, 199034, Saint Petersburg, Russia.
2026 - Volume: 66 Issue: 2 pages: 588-600
https://doi.org/10.24349/nbx7-y9tmZooBank LSID: F549AC61-BDEE-48C4-A18F-6DAD5851D0F0
Original research
Keywords
Abstract
Introduction
Feather mites (Astigmata: Analgoidea and Pterolichoidea) are highly specialized ectosymbionts (commensals and parasites) permanently living on birds (Gaud and Atyeo 1982, 1996; Dabert and Mironov 1999; Proctor and Owens 2000; Proctor 2003; OConnor 2009; Mironov 2016). Most of these mites occupy variety of microhabitats in the plumage of their avian hosts although representatives of some taxa inhabit the skin and the nasal cavities of birds. To date, feather mites have been reported from representatives of all currently recognized avian orders, although the primary origin of these mites collected from some orders of these hosts remains disputable.
The extant orders of birds have been studied rather unevenly with respect to the diversity of feather mite associated with them (Gaud and Atyeo 1996; Proctor 2003; Mironov 2016). The order Gruiformes currently includes about 145 species arranged in six families (Gill et al. 2025), but only the feather mites associated with the family of rails (Rallidae) — mainly with bird species distributed in Europe and Africa — have been examined to meaningful extent (Dubinin 1956; Gaud and Mouchet 1959a, 1959b, 1963; Gaud 1960, 1968; Peterson and Atyeo 1968; Peterson 1971; Atyeo and Peterson 1976; Gaud and Atyeo 1981; Mironov 1981; Mironov and Galloway 2002; Mironov et al. 2024; Waki et al. 2024). Diversity of feather mites distributed on birds from the remaining families of gruiforms (Aramidae, Gruidae, Heliornithidae, Psophiidae, and Sarothruridae) is known quite incompletely (Gaud and Mouchet 1963; Gaud 1968; Peterson and Atyeo 1968; Atyeo and Gaud 1977, 1980, 1984; Gaud and Atyeo, 1996).
In the family of cranes (Gruidae) including 15 extant species, feather mites are currently known only from four cranes: the Common Crane Grus grus (Linnaeus), Sandhill Crane Antigone canadensis (Linnaeus), Black Crowned Crane Balearica pavonina (Linnaeus), and Grey Crowned Crane B. regulorum (Bennett). Feather mites recorded on these hosts belong to the families Alloptidae, Analgidae (Analgoidea), Kramerellidae and Pterolichidae (Pterolichoidea) (Gaud 1968; Atyeo and Windingstad 1979; Atyeo and Gaud 1977, 1980; Mironov and Malovichko 2023; Mironov 2025). Each of these hosts bears a complex of 3–5 monoxenous or stenoxenous feather mite species. Among mite genera recorded from cranes, all pterolichid genera, Apatelacarus Atyeo & Gaud, 1980, Doleracarus Atyeo & Gaud, 1980, Geranolichus Gaud, 1968 and Gruolichus Atyeo & Windingstad 1979 are specific to this avian family, the genus Megniniella Gaud, 1958 (Analgidae) is restricted to Gruiformes, while the genera Brephosceles Hull, 1934 (Alloptidae) and Pseudogabucinia Černý, 1961 (Kramerellidae) are distributed on various orders of aquatic birds.
The present work provides description of new feather mite, which was found on the Common Crane in European Russia and recognized as a new species and new genus of the family Pterolichidae. Additionally, a brief review of feather mites known from birds of the family Gruidae is given.
Material and methods
The specimens of Common Cranes examined in the present study were found deceased in the field in the Stavropol Krai, Russia, in November 2022. The wings and parts of the body plumage were removed from the bird carcasses and stored in plastic bags in a freezer. In the laboratory conditions, the remiges and covert feathers were examined for the presence of feather mites with a Leica stereomicroscope M60 (Leica Microsystems, Inc.). Dead mites detected on feathers were collected manually with a preparation needle and fixed in vials with 95% ethanol. Additionally, all feathers and parts of wings were subjected to a washing collection processing (Mironov and Galloway 2002; Galloway and Lamb 2014). After washing, the water was passed through a filter, and mites were collected under a stereomicrosope and placed in vials with ethanol. Further, the collected mites were softened and cleaned in lactic acid (10%) during one day at room temperature and following the standard technique for small mites (Krantz and Walter 2009) were mounted on microscope slides in Hoyer's medium and dried at 60 °C for 5 days. The examination of mites, preparing the line drawings and taking of measurements were made with a Leica microscope DM2500 (Leica Microsystems, Inc.), equipped with differential interference contrast optics (DIC), a camera lucida, and an ocular micrometer.
Among numerous mites specimens of Geranolichus gruis (Trouessart, 1884), Pseudogabucinia malovichkoae Mironov, 2025 and Megniniella gruophila Mironov & Malovichko, 2023 found on examined feathers and being common to the Common Crane, were detected few specimens of small pterolichid mites, which were recognized as representatives of a new genus and species. The descriptions and diagnoses of new mite taxa are provided according to the formats and measuring techniques used for corresponding taxa of the family Pterolichidae (Dabert and Labrzicka 2009; Mironov et al. 2015; Hernandes and Mironov 2015; Waki et al. 2023; Mironov 2024, 2025). General morphological terms and chaetotaxy follow Gaud and Atyeo (1996), with minor modifications by Norton (1998). All measurements are in micrometers (μm). Scientific names of birds and their supraspecific classification follow the IOC World Bird List, v 15.1 (Gill et al. 2025). Mite specimens are deposited in the Zoological Institute of the Russian Academy of Sciences (Saint Petersburg, Russia).
Systematics
Family Pterolichidae Trouessart and Mégnin, 1884
Subfamily Pterolichinae Trouessart and Mégnin, 1884
Genus Microgruolichus gen. nov.
ZOOBANK: 812125DC-64A9-4941-9F35-52399F2FA2B3 ![]()
Type species — Microgruolichus armogaster sp. n.
Description
Both sexes — Small-sized pterolichines, idiosoma noticeably elongated, with almost parallel lateral margins. Subcapitulum nearly quadrangular; pseudolrutellar projections short and rounded, without lateral extension. Prodorsal shield entire, occupying almost entire dorsal surface of prodorsum, strongly widened posteriorly (roughly triangular in shape). Vertical seta vi unpaired, filiform; setae ve absent. Scapular macrosetae se short, less than half-width of idiosoma; setae si small filiform. Sejugal area with fine transverse striation. Hysteronotal shield covering most part of hysterosoma. Lateral hysteronotal bands not separated from the main body of hysteronotal shield. Humeral shields present, ventrally fused with epimerites III. Setae cp and c3 on sclerotized areas at bases of epimerites III. Lateral hysteronotal setae c2, d2, e2, f2, short filiform. Setae c3 shorter than half-width of idiosoma. Setae h1 present. Hysteronotal gland openings gl distinct. Cupules ia indistinct, cupules im present. Supranal concavity present. Epimerites I-IV simple, shaped as slightly curved sticks. Epimerites I fused into a V. Epimerites I, II without basal inflation. Coxal fields I-IV without extensive sclerotized areas. All segments of legs I, II simple, without processes and expansions. All setae of tarsus, tibia and genu I, II simple filiform. Seta ba anterior to base of corresponding solenidion ω1 on tarsi I, II. Genu I with one solenidion σ situated at distal margin of segment. Solenidia σ on genua II, III present. Trochanteral setae sRIII short filiform, not extending to distal margins of femora III. Distal margin of ambulacral disc concave, with pair of minute spines. Leg chaetotaxy formula (tarsus–trochanter, proral setae not included, solenidia in brackets): I 8(2)–1(1)–2(1)–1–1, II–8(1)–1(1)–2(1)–1–1, III 6–1(1)–(1)–0–1, IV 6–1(1)–0–0.
Male — Idiosoma elongated, over 2 times longer than wide. Opisthosomal lobes present, roughly triangular in shape, moderately elongated, approximately as wide as long at base. Terminal cleft U-shaped. Lateral margins of terminal cleft with narrow interlobar membranes. Bases of setae f2 and ps2 close to each other and situated laterally on posterolateral margins of opisthosoma. Setae h2 represented by macrosetae, situated at level of opisthosomal lobe bases; setae h3 long needle-like; setae ps1 small lanceolate; bases of setae h3 and ps1 on or near lobar apex. Genital apparatus situated at level of trochanters IV. Genital papillae at level of genital arch. Setae 4b posterior to level of setae 3a. Setae g at level of genital arch. Epiandrum and paragenital apodemes absent. Genital shields paired, large, bearing setae 4a. Adanal shields absent. Opisthoventral shields large, flanking anal area anterolaterally. Adanal suckers large, longitudinally ovate, surrounded by wide membrane with radial striation, corolla without denticles. Setae ps3 anterior to opisthoventral shield. Legs IV slightly thicker than legs III, with tarsus extending to bases of opisthosomal lobes. Tarsus IV cylindrical similar in length to tibia IV, without subapical claw- or spine-like projections; modified setae dIV, eIV button-like.
Female — Posterior end of opisthosoma widely rounded. Hysteronotal shield entire, covering most area of hysterosoma from humeral shields to posterior margin of opisthosoma; lateral areas of this shield spread onto ventral surface of hysterosoma forming peripheral opisthoventral shield from trochanters III to bases of setae h2. Setae d1, d2, e1, e2, and h1 on hysteronotal shield. Setae e1, e2, and h1 moved closely to posterior end of opisthosoma. Bases of setae f2 and ps2 close to each other. Setae h3 much shorter than marcosetae h2. Epigynum small bow-shaped, situated at level of sejugal area. Oviporus at level of epimerites III and humeral shields. Genital papillae anterior to level of setae 3a. Genital setae g posterior to level of setae 3a and oviporus. Legs IV not extending to posterior margin of opisthosoma
Remarks
The new genus Microgruolichus gen. n. and four previously known pterolichine genera, Apatelacarus Atyeo and Gaud, 1980, Doleracarus Atyeo and Gaud, 1980, Geranolichus Atyeo and Windingstad, 1979, and Gruolichus Atyeo and Windingstad, 1979 associated with cranes, constitute the Gruolichus generic group. This group can be characterized by the following set of characters. In both sexes, idiosoma is realtively short, about 300 in males and 400 in females, the lateral bands are not separated from the main body of hysteronotal shield, epimerites I are fused in a V or Y, setae si are setiform and close to corresponding setae se, most setae of hysterosoma are short setiform, setae ba are anterior from solenidia ω1 in tarsi I, II, and solenidion σ on genu III is present. In males, the adanal suckers lack denticles, legs IV are slightly thicker than legs III, tarsus IV with or without short spine, and pretarsus IV is present. In females, the hysteronotal shield is entire and covers most surface of hysterosoma, setae e1 and h1 are close to each other, arranged in a rectangle or trapezoid, and usually strongly moved to the posterior part of opisthosoma.
Download as Remark: (?)—diagnosis of the genus does not give unambiguous information that the spine is absent.
Characters
Apatelacarus
Doleracarus
Geranolichus
Gruolichus
Microgruolichus gen. n.
Both sexes
Setae kTIII
Present
Present
Present
Absent
Present
Number of setae vi
2
2
2
1
1
Setae cGI, II
Bifurcate
Bifurcate
Setiform
Setiform
Setiform
Setae c3
Setiform
Spine
Setiform
Setiform
Setiform
Male
Shape of epimerites I
V
Y
Y
Y
V
Genital app./ trochanters
Between III and IV
At trochanters III
Between III and IV
Between III and IV
At trochanters IV
Genital papillae position
Apex of arch
Midlevel of arch
Midlevel of arch
Posterior to arch
Midlevel of arch
Setae ps1
Triangular
Setiform
Lanceolate
Setiform
Lanceolate
Setae ps2
Setiform
Setiform
Lanceolate
Setiform
Setiform
Spine on tarsus IV
Present
Absent
Present
Absent (?)
Absent
Legs IV/lobar apices
Beyond
Extending to
Beyond
Beyond
Not extending
Female
Shape of epimerites I
V
Y
Y
Y
V
Legs IV/opisthosoma
Not extending
Beyond
Beyond
Not extending
Not extending
The new genus, Microgruolichus gen. n. is most close to the genus Gruolichus in sharing the following features: in both sexes, seta vi is unpaired, setae c3 are short setiform, and genual setae cGI, II are simple setiform; in females, the idiosoma is noticeably parallel-sided, legs IV are short and do not extend to the posterior margin of opisthosoma (Table 1). The genus Microgruolichus differs from the latter in the following features: in both sexes, tibial setae kTIV are present, epimerites I are fused into a V; in males, setae ps1 are lanceolate, the genital papillae and setae g are situated at the level of genital arch apex, the adanal suckers are longitudinally ovate, the genital and opisthoventral shields are present, and legs IV do not extend to the level of lobar apices; in females, the hysteronotal shield margins are curved onto the ventral side of the body forming wide peripheral opisthoventral shield from trochanters III to bases of setae h2, and setae h3 are much shorter than marcrosetae h2. In both sexes of the genus Gruolichus, the tibial setae kTIV are absent, epimerites I are fused into a Y; in males, setae ps1 are short filiform, the genital papillae are posterior to the tips of genital arch, while setae g are anterior from this arch, the adanal suckers are circular, the genital and opisthoventral shields are absent, and legs IV with tarsi extend beyond the level of lobar apices; in females, the posterolateral areas of ventral hysterosoma are not sclerotized, and setae h3 are about 3/4 the length of macrosetae h2.
Etymology
The generic name refers to the close affinity to the genus Gruolichus and relatively smaller size of mites of the new genus.
Microgruolichus armogaster sp. n.
ZOOBANK: 8AF7104F-D56B-470D-BC6B-7F8EFFF6EA2A ![]()
(Figures 1–3)
Type material
Male holotype and 2 female paratypes (ZISP 24216) from Grus grus (Linnaeus, 1758) (Gruiformes: Gruidae), Russia, Stavropol Krai, Turkmen District, near Poperechnyi, 45°25′22″N 43°15′28″E, 10 November 2022, bird coll. L.V. Malovichko.
Description

Male — (holotype) (Figs. 1, 3 A–F). Idiosoma narrowly ovate, length × width, 275 × 125; length of hysterosoma 200. Subcapitulum nearly rectangular, length including palps 35, width at base 32. Prodorsal shield: roughly triangular, occupying almost entire surface of prodorsum, anterolateral extensions not developed, lateral margins with small incisions posterior to level of trochanters II, setae se on border of these incisions, posterolateral corners rounded, posterior margin straight, length along midline 75, greatest width of posterior part 95. Seta vi minute filiform, about 10 long. Distance between scapular setae se 63; setae si short filiform, about 15 long, separated by 45. Scapular shields narrow, with poorly distinct margins. Humeral shields not developed dorsally, fused ventrally with outer margin of epimerites III. Setae c2 small filiform, about 5 long, situated on striated tegument. Subhumeral setae c3 filiform, 15 long, situated near bases of corresponding humeral setae cp on sclerotized area of epimerites III. Lateral margins of hysterosoma from level of trochanters IV to bases of setae h2 flattened and membranous. Hysteronotal shield: covering almost entire surface of dorsal hysterosoma; anterior margin slightly concave, anterior corners rounded, lateral bands not separated from the main body of this shield; most surface uniformly punctured, anterior ¼ with several poorly outlined ovate lacunae, posterolateral borders heavily sclerotized, greatest length from anterior margin to lobar apices 185, width at anterior margin 105. Hysteronotal gland openings gl on hysteronotal shield near lateral margins, approximately at midlength between trochanters III and IV. Cupules im situated submarginally at level of trochanters IV. Setae d2 anterior to openings gl. Opisthosomal lobes roughly triangular, slightly shorter than wide at base, with narrow rounded apices. Terminal cleft U-shaped, inner margins of lobes with interlobar membranes. Incision outlined by free margins of membranes shaped as a narrowly U, length of incision 23, width between free margins 14. Supranal concavity poorly outlined, represented by several longitudinal folds anterior from terminal cleft. Setae f2, ps2 and macrosetae h2 situated on posterolateral margin of opisthosomal lobe, approximately at level of lobar bases. Setae h3 rod-like and acute apically, 35 long, situated on lobar apices. Setae ps1 lanceolate, 18 × 5, situated subapically on inner margins of opisthosomal lobes. Setae f2 short filiform. Setae ps2 long filiform, extending beyond lobar apices. Setae h1 minute filiform, situated at level of supranal concavity. Distances between dorsal setae and openings: c2:d2 76, d2:e2 64, d2:gl 10, e2:h3 52, d1:d2 20, e1:e2 28, h2:h2 59, h3:h3 35, ps1:ps1 30, ps2:ps2 43.
Epimerites I fused in a V. Epimerites I, II without extensively sclerotized areas. Epimerites III, IV with narrow triangular sclerotized areas flanking bases of corresponding trochanters. Epimerites IVa small, connected with sclerotized margins of hysteronotal shield curved to ventral side of hysterosoma. Genital apparatus at level of trochanters IV (in holotype, the arch is turned back). Genital arch narrow, 11 × 7; aedeagus with curved tip, slightly extending beyond arch apex. Genital papillae situated at level of anterior part of genital arch. Genital shields large, pear-shaped, situated posterior to level of trochanters IV and orientated transversely, with setae 4a on anterior margins. Setae 4b far posterior to level of setae 3a, approximately at level of epimerites IIIa. Setae g anterior to level of genital arch apex, at midlength of epimerites IV. Cupules ih absent. Opisthoventral shields large triangular, flanking anal area anterolaterally. Pseudanal setae ps3 anterior from inner ends of opisthosventral shields. Adanal suckers large, longitudinally ovate, 25 × 12, corolla without denticles, surrounding membrane with sparse striae. Distances between ventral setae: 3a:4b 17, 4b:g 16, g:4a 32, 4a:ps3 30, ps3:h3 55, ps3:ps3 40.
Ambulacral discs of all legs with slightly concave distal margin and with a pair of minute spines. Legs I–III without apophyses and enlargements. Legs IV slightly thicker than legs III, with tarsus not extending to level of setae h2 on margins of opisthosomal lobes. Tarsus IV without spine-like process, modified setae d, e button-like. Tibial solenidion φIV extends beyond distal margin of ambulacral disc. Lengths of tarsi: I 20, II 22, III 18, IV 20. Lengths of solenidia: ω1I 10, ω1II 21, σI 35, σII 4, σIII 9, φIII 32, φIV 50.
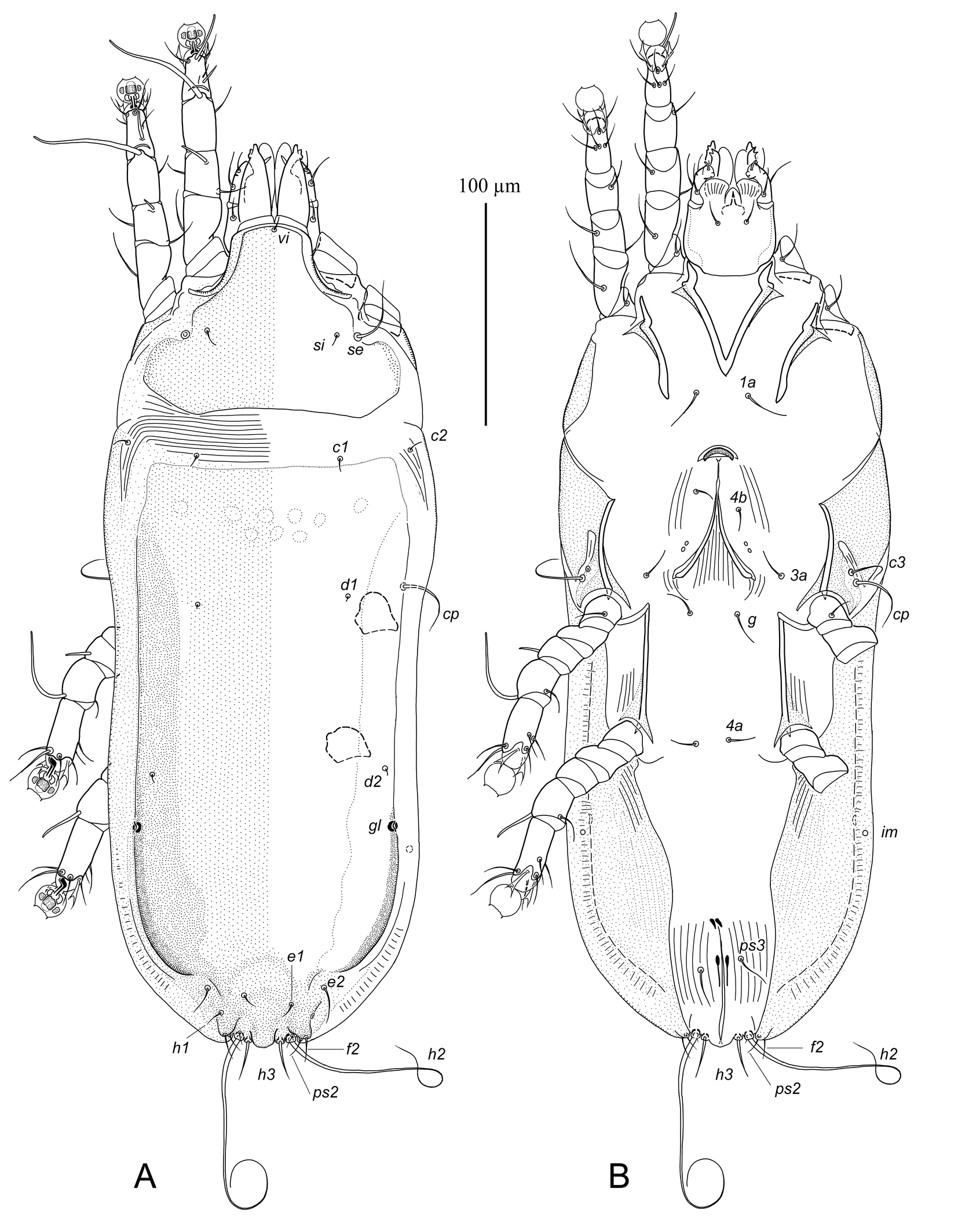

Female — (range for 2 paratypes) (Figures 2, 3G–I). Idiosoma elongated, with almost parallel lateral margins, length × width, 360–375 × 150–155; length of hysterosoma 270–280. Gnathosoma as in the male, 47–48 × 43–45. Prodorsal shield shaped as in the male, except posterior margin slightly concave, length along midline 82–85, and width of posterior part 116–120. Seta vi filiform, about 10 long. Distance between scapular setae se 75–80. Setae si thin filiform, about 15 long, separated by 60–65. Scapular shields narrow, barely developed dorsally. Humeral shields: border with hysteronotal shield indistinct, ventral part fused with epimerites III. Setae c2 short filiform, about 10 long, situated near anterior end of humeral shields. Subhumeral setae c3 filiform, 12–15 long, situated near bases of setae cp on sclerotized areas of epimerites III. Hysteronotal shield: covering almost entire dorsal surface of hysterosoma, anterior margin straight or slightly concave, lateral bands not separated from the main body, lateral parts spread to ventral side of hysterosoma forming wide opisthoventral shields from level of trochanters III to bases of setae h2, surface punctured, lateral areas near margins sclerotized much stronger than median area, length 255–265, and width at anterior margin 120–125. Lateral areas of opisthosoma from level of trochanters III to bases h2 strongly flattened, looking like narrow and strongly sclerotized membranes. Opisthosoma widely rounded posteriorly, with smoothed median extension between bases of setae h3. Supranal concavity absent. Hysteronotal gland openings gl near lateral margins of hysteronotal shield, significantly posterior to trochanters IV. Cupules im situated submarginally, posterior to openings gl. Setae c1, d1, d2 minute filiform; setae c1 on soft tegument or on anterior margin of hysteronotal shield. Setae e1, e2 and h1 moved to posterior 1/8 of hysterosoma; setae e1 and h1 short filiform, about 10 long, arranged in a low trapezoid and situated posterior to level of setae e2. (Figure 3H). Setae h3 17–22 long, much shorter than macrosetae h2. Setae ps1 minute, barely distinct, situated at bases of macrosetae h2. Distances between setae and openings: c2:d2 140–145, d2:e2 95–100, d2:gl 22–25, e2:h3 22–25, d1:d2 70–78, e2:e1 6–10, e1:e1 18–22, e2:e2 53–55, h2:h2 24–25, h3:h3 15–16.
Epimerites I fused in a V. Epimerites I, II without sclerotized areas. Epimerites IVa absent. Opisthoventral shields formed by lateral parts of hysteronotal shields posterior to trochanters IV strongly widened, poorly sclerotized median area approximately as wide as 1/3 opisthosoma width. Epigynum small, nearly crescent-shaped, 7–8 long and 20–22 wide. Setae 4b approximately at midlength of oviporus. Setae g posterior to level of setae 3a and tips of ovoporal apodemes. Setae 4a at level of trochanters IV. Genital papillae at level of posterior 1/3 of oviporus. Copulatory opening terminal, on median convexity of opisthosoma. Distal part of spermatheca narrow, poorly sclerotized; secondary spermaducts 10–12 long (Figure 3I). Distances between ventral setae: 4b:3a 28–30, 3a:g 10–18, g:4a 55–62.
Legs I–II as in the male. Legs IV with ambulacral discs not extending to posterior margin of opisthosoma. Lengths of tarsi: I 22–23, II 24–25, III 25–26, IV 27–29. Lengths of solenidia: ω1I 8–10, ω1II 12–13, σI 26–28, σII 3–4, σIII 10–11, φIII 38–40, φIV 12–14.
Remark
Differential characters as for the genus.
Etymology
The specific epithet is a combination of arma (L., armor) and gaster (Gr. stomach, belly) referring to the extensive peripheral slerotisation of the opisthosoma in females.
Discussion
The feather mite fauna associated with cranes (Gruiformes: Gruidae) is currently known only from four avian species belonging to three genera (Table 2). This fauna is represented by mites from four families: Alloptidae, Analgidae (Analgoidea), Kramerellidae and Pterolichidae (Pterolichoidea) (Gaud 1968; Atyeo and Windingstad 1979; Atyeo and Gaud 1980; Mironov and Malovichko 2023; Mironov 2025). Mites of the three families, Alloptidae, Kramerellidae and Pterolichidae, live on the wing feathers of these hosts. Pterolichid mites inhabit the vanes of primaries and secondaries, kramerellids are located on the same feathers, but in the very proximal parts of vanes and near the rachis; and alloptids commonly occur on the vanes of the lower coverts of the wings. These observations were made in the process of manual collecting of mites from wings of Grus grus. Mites of the genus Megniniella, as for most analgids, apparently occupy downy and covert feathers of the wing and body.
Download as
Host species
Mite species
Mite family
Locality
Reference
Antigone canadensis (Linnaeus)
Brephosceles petersoni Atyeo & Windingstad, 1979
Alloptidae
USA: WI
Atyeo and Windingstad 1979
Geranolichus canadensis Atyeo & Windingstad, 1979
Pterolichidae
USA: IN, WI; Russia: Chukotka
Dubinin 1956; Atyeo and Windingstad 1979; Atyeo and Gaud 1980
Gruolichus wadashae Atyeo & Windingstad, 1979
Pterolichidae
USA: IN, WI
Atyeo and Windingstad 1979; Atyeo and Gaud 1980
Pseudogabucinia reticulata Atyeo & Windingstad, 1979
Kramerellidae
USA: IN
Atyeo and Windingstad 1979
Balearica pavonina (Linnaeus)
Apatelacarus brachychaetus (Gaud, 1968)
Pterolichidae
Cameroon
Gaud 1968; Atyeo and Gaud 1980
Doleracarus tetrachaetus (Gaud, 1968)
Pterolichidae
Kenya
Atyeo and Gaud 1980
Pseudogabucinia moucheti Gaud, 1968
Kramerellidae
Cameroon
Gaud 1968
Balearica regulorum (Bennett)
Brephosceles geranoxenus Peterson, 1968
Alloptidae
Malawi, Zimbabwe
Gaud 1968; Peterson 1971
Apatelacarus brachychaetus (Gaud, 1968)
Pterolichidae
Kenya
Atyeo and Gaud 1980
Doleracarus tetrachaetus (Gaud, 1968)
Pterolichidae
Kenya, Malawi
Gaud 1968; Atyeo and Gaud 1980
Grus grus (Linnaeus)
Brephosceles discurus (Trouessart, 1885)
Alloptidae
Europe; Russia: European pt.
Trouessart 1885; Peterson and Atyeo 1968; Mironov 2025
Geranolichus gruis (Trouessart, 1884)
Pterolichidae
Europe; Russia: European pt., Transcaucasia, Chita oblast
Trouessart 1884; Dubinin 1956; Mironov 2025
Microgruolichus armogaster gen. n., sp. n.
Pterolichidae
Russia: European pt.
Present study
Megniniella gruophila Mironov & Malovichko, 2023
Analgidae
Russia: European pt.
Mironov and Malovichko 2023; Mironov 2025
Pseudogabucinia malovichkoae Mironov, 2025
Kramerellidae
Russia: European pt.
Mironov 2025
It is interesting to note that on each of these cranes, the family Pterolichidae is represented by a pair of cohabiting pterolichid species. The genus Microgruolichus described herein fills the gap in a pair of pterolichid genera on the Common Crane. Mites in these pairs are noticeably different in size that probably allows them to live in different areas of the vanes of wing feathers and avoid competition. On Antigone canadensis and Grus grus, the larger pterolichids are represented by monoxenous species of the genus Geranolichus, while small-sized ones on these hosts are represented by mites of the different genera, Gruolichus and Microgruolichus, respectively. On both species of crowned cranes of the genus Balearica, the larger and smaller pterolichids are represented by the species Doleracarus tetrachaetus and Apatelacarus brachychaetus, respectively. Similar pattern, the occurrence of two pterolichid species on one host species, is observed on many species of rails (Gruiformes: Rallidae) (Gaud and Mouchet 1963; Gaud 1968), on which larger and smaller pterolichines are represented by the genera Grallolichus Gaud, 1960 and Grallobia Hill, 1934. More complicate cases of cohabitation are presented by trumpeters (Gruiformes: Psophiidae), each species of these birds bears three pterolichid species belonging to different monotypic genera (Atyeo and Gaud 1984). Of them, Ceratothrix corniger (Trouessart & Neumann, 1888) is a large pterolichid species, while Atrichotibia fistella Atyeo & Gaud, 1984 and Mastigodiscus bucolis Atyeo & Gaud, 1984 are small-sized pterolichids.
Mites of the genus Pseudogabucinia (Kramerellidae) were found on Antigone canadensis, Grus grus and on Balearica pavonina. On each of these hosts, the genus Pseudogabucinia is represented by a specific mite species. Similarly, mites of the genus Brephosceles (Alloptidae) were recorded on the same species of large cranes, A. canadensis and G. grus, but also on another species of crowned crane, Balearica regulorum. As in the case of the genus Pseudogabucinia, each of these avian hosts bore a specific species of the genus Brephosceles. Taking in attention that the pterolichid mites Apatelacarus brachychaetus and Doleracarus tetrachaetus occur on both species of crowned cranes – Balearica spp. – (Atyeo and Gaud 1980), it is reasonable to suggest that Brephosceles geranoxenus and Pseudogabucinia moucheti, each found so far on only one Balearica species, apparently could inhabit both species of crowned cranes.
Megniniella gruophila (Analgidae) described from the Common Crane is the only representative of this genus recorded so far from cranes (Mironov and Malovichko 2023). Since all the remaining species of the genus Megniniella are widely distributed on rails (Rallidae) and one species is also known from the Limpkin, Aramus guarauna (Aramidae), it is reasonable to expect that some species of this genus could also be found on other cranes.
Regarding the taxonomic content of feather mite complexes of feather mites associated with cranes, it is interesting to note that feather mites of the family Xolalgidae (Analgoidea), which members are quite diverse on closely related rails (Rallidae) and also occur on other gruiforms such as trumpeters (Psophiidae) and finfoots (Heliornithidae), have never been recorded on cranes (Gaud 1968; Atyeo and Gaud 1984; Atyeo and Peterson 1976; Gaud and Atyeo 1981).
Acknowledgements
The author thanks Aleksandr V. Matyukhin (A.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences, Moscow, Russia) for the delivery wing feather of crane to the Zoological Institute RAS. This study was supported by the Ministry of Science and Higher Education of the Russian Federation (project No. 125013001089-0).
References
- Atyeo W.T., Gaud J. 1977. Gruiformes, a new host group for pterodectine feather mites (Acarina: Analgoidea). Journal of Parasitology, 63: 141-144. https://doi.org/10.2307/3280121
- Atyeo W.T., Gaud, J. (1979) 1980. Geranolichus Gaud and related genera (Acarina, Analgoidea, Pterolichidae). Acarologia, 21: 487-495.
- Atyeo W.T., Gaud J. 1984. The feather mites (Acarina) of the Psophiidae (Aves). Acarologia, 25: 365-375.
- Atyeo W.T., Peterson P.C. 1976. The feather mite genera Zumptia Gaud & Mouchet and Parazumptia gen. nov. (Acarina, Analgoidea). Tijdschrift voor Entomologie, 119: 327-335.
- Atyeo W.T., Windingstad R.M. 1979. Feather mites of the greater sandhill crane. Journal of Parasitology, 65: 650-658. https://doi.org/10.2307/3280336
- Dabert J., Labrzicka A. 2009. Szeptyckiana, a new genus of the feather mite family Pterolichidae (Acari, Astigmata, Pterolichoidea) from the Black-thighed Falconet Microhierax fringillarius (Falconiformes, Falconidae). Acta Zoologica Cracoviensia, 52B (1-2): 61-72. https://doi.org/10.3409/azc.52b_1-2.61-72
- Dabert J., Mironov S.V. 1999. Origin and evolution of feather mites (Astigmata). Experimental and Applied Acarology, 23: 437-454. https://doi.org/10.1023/A:1006180705101
- Dubinin V.B. 1956. Feather mites (Analgesoidea). Part III. Family Pterolichidae. Fauna of the USSR, Paukoobraznye, Vol. 6, Fasc. 7, Publisher: AN SSSR, Moscow-Leningrad, 813 pp. [In Russian]
- Galloway T.D., Lamb R.J. 2014. Abundance and stability are species traits for four chewing lice (Phthiraptera: Menoponidae, Philopteridae) on feral pigeons, Columba livia Gmelin (Aves: Columbiformes: Columbidae). The Canadian Entomologist, 146 (4): 444-456. https://doi.org/10.4039/tce.2013.86
- Gaud J. 1960. Quelques Sarcoptiformes plumicoles du Congo belge (Analgesoïdea). Revue de Zoologie et de Botanique Africaines, 61: 133-159.
- Gaud J. 1968. Acariens sarcoptiformes plumicoles (Analgoidea) parasites sur les oiseaux Ralliformes et Gruiformes d'Afrique. Annales du Musée Royale de l′Afrique Centrale, Sér. in-8°, Sciences Zoologiques, 164: 1-101.
- Gaud J., Atyeo W.T. 1981. La famille Xolalgidae Dubinin (Acariens plumicoles, Analgoidea). II. Sous-familles Xolalginae et Zumptiinae, n. sub-fam. Acarologia, 22: 313-324.
- Gaud J., Atyeo W.T. 1982. Spécificité parasitaire chez les acariens Sarcoptiformes plumicoles. Mémoires du Muséum National d'Histoire Naturelle, N. S., Sér. A, Zoologie, 123: 247-254.
- Gaud J., Atyeo W.T. 1996. Feather mites of the World (Acarina, Astigmata): the supraspecific taxa. Musée Royal de l′Afrique Centrale, Annales, Sciences Zoologiques, 277 (Pt. 1): 1-193 (text) and (Pt. 2): 1-436 (illustrations).
- Gaud J., Mouchet, J. 1959a. Acariens plumicoles (Analgesoidea) parasites des oiseaux du Cameroun. II. Analgesidae. Annales de Parasitologie Humaine et Comparée, 34: 149-208. https://doi.org/10.1051/parasite/1959341149
- Gaud J., Mouchet J. 1959b. Acariens plumicoles des oiseaux du Cameroun. V. Pterolichidae. (1re Partie). Annales de Parasitologie Humaine et Comparée, 34: 493-545. https://doi.org/10.1051/parasite/1959344452
- Gaud J., Mouchet J. 1963. Révision des genres Grallobia Hull et Grallolichus Gaud (Pterolichidae). Acarologia, 5: 628-643.
- Gill F., Donsker D., Rasmussen P. (Eds.). 2025. IOC World Bird List (v 15.2). Available from: http://www.worldbirdnames.org/ (accessed: 15 May 2026). https://doi.org/10.14344/IOC.ML.15.2.
- Hernandes F.A., Mironov S.V. 2015. The feather mites of the hoatzin Opisthocomus hoazin (Müller) (Aves: Opisthocomiformes), with description of two new genera and six new species (Acari: Analgoidea, Pterolichoidea). Zootaxa, 4034 (3): 401-444. https://doi.org/10.11646/zootaxa.4034.3.1
- Krantz G.W., Walter D.E. (Eds.). 2009. A Manual of Acarology. 3rd Edition. Texas Tech University Press, Lubbock, TX, USA, p. 565-657.
- Mironov S.V. 1981. Feather mites of the genus Metanalges (Sarcoptiformes, Analgoidea) of the USSR fauna. Parazitologiya, 15 (5): 459-468. [In Russian with English summary]
- Mironov S.V. 2016. Chapter V. Host-parasite relations of mites of the parvorder Psoroptidia (Acariformes: Astigmata) with birds. In: Galaktionov K.V. (Ed.), Coevolution of Parasites and Hosts. Proceedings of the Zoological Institute of the Russian Academy of Sciences, 320, Supplement 4, p. 264-348. [In Russian]
- Mironov S.V. 2024. A new feather mite genus of the family Pterolichidae (Acariformes) from the common pheasant, Phasianus colchicus, with a checklist of pterolichines associated with galliform birds (Aves: Galliformes). Acarina, 32 (2): 107-128. https://doi.org/10.21684/0132-8077-2024-32-2-107-128
- Mironov S.V. 2025. Feather mites (Acariformes: Psoroptidia) associated with the Common Crane Grus grus (Gruifomes: Gruidae). Acarina, 33 (1): 17-36. https://doi.org/10.21684/0132-8077-2025-33-1-17-36
- Mironov S.V., Galloway T.D. 2002. Four new feather mite species (Acari: Analgoidea) from some birds in Canada. The Canadian Entomologist, 134: 605-618. https://doi.org/10.4039/Ent134605-5
- Mironov S.V., Haarder S., Galloway T.D. 2024. A new feather mite species of the genus Temnalges (Acariformes: Psoroptoididae) from coots, Fulica spp. (Gruiformes: Rallidae). Acarologia, 64 (4): 1106-1116. https://doi.org/10.24349/i1su-eqk9
- Mironov S.V., Hernandes F.A., Pedroso L.G. 2015. New feather mites of the genera Aniacarus and Aniibius (Acariformes: Pterolichidae) from two cuckoo species (Cuculiformes: Cuculidae) in Brazil. Zootaxa, 3937 (1): 103-126. https://doi.org/10.11646/zootaxa.3937.1.5
- Mironov S.V., Malovichko L.V. 2023. A new feather mite of the genus Megniniella (Acariformes: Analgidae) from the Common Crane Grus grus (Gruiformes: Gruidae). Systematic and Applied Acarology, 28(9): 1553-1564. https://doi.org/10.11158/saa.28.9.8
- Norton R. 1998. Morphological evidence for the evolutionary origin of Astigmata (Acari: Acariformes). Experimental and Applied Acarology, 22: 559-594. https://doi.org/10.1023/A:1006135509248
- OConnor B.M. 2009. Cohort Astigmatina. In: Krantz G.W. and Walter D.E. (Eds.). A Manual of Acarology. 3rd Edition. Texas Tech University Press, Lubbock, TX, USA, p. 565-657.
- Peterson P.C. 1971. A revision of the feather mite genus Brephosceles (Proctophyllodidae: Alloptinae). Bulletin of the University of Nebraska State Museum, 9: 89-172.
- Peterson P.C., Atyeo W.T 1968. New genera related to the genus Brephosceles Hull, 1934 (Acarina: Proctophyllodidae). Bulletin of the University of Nebraska State Museum, 8: 217-236.
- Proctor H.C. 2003. Feather mites (Acari: Astigmata): ecology, behavior and evolution. Annual Review of Entomology, 48: 185-209. https://doi.org/10.1146/annurev.ento.48.091801.112725
- Proctor H.C., Owens I. 2000. Mites and birds: diversity, parasitism and coevolution. Trends in Ecology and Evolution, 15: 358-364. https://doi.org/10.1016/S0169-5347(00)01924-8
- Trouessart E.L. 1884. Les Sarcoptides plumicoles. Journal de Micrographie, 8: 527-532, 572-579.
- Trouessart E.L. (1884) 1885. Note sur le classification des Analgésiens et diagnoses d'espèces et de genres nouveaux. Bulletin de la Société d'Etudes Scientifiques d'Angers, 14: 46-89.
- Waki T., Mironov S.V, Nakajima A., Shimano S. 2023. A new feather mite of the genus Pelargolichus (Acariformes: Pterolichidae) from the Oriental White Stork Ciconia boyciana (Ciconiiformes: Ciconiidae) in Japan. Systematic and Applied Acarology, 28(3): 471-482. https://doi.org/10.11158/saa.28.3.5
- Waki T., Mironov S., Nakaya Y., Nagamine T., Shimano S. 2024. A new feather mite species of genus Metanalges (Acariformes: Analgidae) from the Okinawa Rail, Hypotaenidia okinawae (Gruiformes: Rallidae), in Okinawa Island, Japan. Species Diversity, 29: 1-9. https://doi.org/10.12782/specdiv.29.161



2026-04-13
Date accepted:
2026-06-09
Date published:
2026-06-18
Edited by:
Akashi Hernandes, Fabio

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Mironov, Sergey V.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)