



Functional response of the predatory mite Amblydromalus limonicus to Tetranychus urticae (Acari: Phytoseiidae, Tetranychidae)
Santana, Rita  1
; Pérez Salinas, Ruth
2
; Mangui, José
3
; Ortiz-Tirado, Paúl
4
; Telenchana, Norma
5
and López-Villacís, Cristina
6
1
; Pérez Salinas, Ruth
2
; Mangui, José
3
; Ortiz-Tirado, Paúl
4
; Telenchana, Norma
5
and López-Villacís, Cristina
6
1✉ Escuela de Posgrados. Universidad Nacional de Trujillo, Peru & Facultad de Ciencias Agropecuarias, Universidad Técnica de Ambato, Ecuador.
2Facultad de Ciencias Agropecuarias, Universidad Técnica de Ambato, Ecuador.
3Facultad de Ciencias Agropecuarias, Universidad Técnica de Ambato, Ecuador.
4Facultad de Ciencias Agropecuarias, Universidad Técnica de Ambato, Ecuador.
5Agrícola La Bota, Ecuador.
6Facultad de Ciencias Agropecuarias, Universidad Técnica de Ambato, Ecuador.
2026 - Volume: 66 Issue: 2 pages: 543-551
https://doi.org/10.24349/y3eo-3v5mOriginal research
Keywords
Abstract
Introduction
In Ecuador, ornamental plant production is one of the most important horticultural activities, reaching about 9,316 hectares of flowers in 2019, of which 77.39% corresponds to Hypericum, Gypsophila, Aster, and rose cultivation. The latter represents 62.09% of the total flowers cultivated, with a national production of 3,346 million cut stems (Rivero Guerra 2021). Rose cultivars are highly susceptible to infestations by the two-spotted spider mite (Tetranychus urticae Koch), whose feeding causes substantial economic losses (Lee et al. 2022). Conventional management of T. urticae still depends largely on chemical acaricides; however, increasing attention is being directed towards biological control strategies as concerns over resistance, environmental impact and residue regulations continue to mount.
The two-spotted spider mite is a highly polyphagous and destructive pest affecting a wide range of economically important crops, including ornamentals such as roses. Its rapid life cycle, high reproductive potential, and ability to develop resistance to acaricides have made its management increasingly challenging, driving the need for sustainable control strategies such as biological control using predatory mites of the family Phytoseiidae (Fathipour and Maleknia 2016; Van Leeuwen et al. 2015; Lemaire et al. 2025). Among these, specialist species such as Phytoseiulus persimilis Athias-Henriot and Neoseiulus californicus (McGregor) are widely used due to their high efficiency in suppressing T. urticae (Tiftikçi et al. 2020; Urbaneja-Bernat and Jaques 2025; Xiao et al. 2013).
In contrast, generalist predators may provide complementary advantages, particularly under low prey densities or early crop stages, due to their ability to persist on alternative food sources. Amblydromalus limonicus (Garman & McGregor) is a generalist phytoseiid commonly found in crops across the Ecuadorian Andes. This predatory mite is widely used in biological control programmes targeting thrips and whiteflies (Lee and Zhang 2018; Schoeller et al. 2022). However, its role in the control of T. urticae remains unclear and potentially limited. Previous studies indicate that generalist predators may face constraints when preying on spider mites, particularly due to dense webbing that reduces prey accessibility and predator mobility (McMurtry and Croft 1997; Wang et al. 2024). Furthermore, A. limonicus may not complete development when feeding exclusively on T. urticae under certain conditions (Wang et al. 2024), suggesting that its effectiveness as a standalone control agent is restricted.
Despite these limitations, evaluating the functional response of A. limonicus remains essential to quantify its predation capacity and to better define its potential role within integrated pest management (IPM) strategies, particularly under Andean production conditions where locally adapted natural enemies may offer practical advantages. Functional response analysis provides key parameters, such as attack rate and handling time, that allow assessment of predator efficiency across prey densities and contribute to predicting its performance as a complementary biological control agent.
Therefore, this study aimed to evaluate the functional response of A. limonicus when preying on eggs and mobile stages of T. urticae under controlled laboratory conditions.
Material and methods
Study site and biological material
The study was conducted in the Entomology Laboratory of the Faculty of Agricultural Sciences, Universidad Técnica de Ambato (Ecuador), under controlled conditions (25 ± 2 °C, 65 ± 5% RH, 16:8 h L:D photoperiod). Tetranychus urticae and A. limonicus were collected from naturally infested rose (Rosa sp.) plants grown at the Querochaca Experimental Farm.
Both species were reared following standard phytoseiid rearing protocols (Pazmiño et al. 2018). T. urticae colonies were maintained on fresh rose leaves replaced every 48 h. A. limonicus colonies were reared on T. urticae-infested leaves, and adult females aged 3–5 days were isolated and starved for 24 h prior to experimentation to standardise hunger level.
Experimental arenas and prey densities
Functional response assays were performed on 2-cm diameter rose-leaf discs placed over moistened cotton inside Petri dishes to maintain leaf turgor and prevent prey escape. Two separate experiments were conducted: eggs: 5, 10, 20, 30, and 40 eggs per predator and mobile stages (larvae + protonymphs): 1, 2, 4, 8, and 16 individuals per predator. Each density treatment had five independent replicates, and the whole experiment was repeated on three independent days, treated as temporal blocks. Control arenas without predators were used to correct for natural prey mortality. After 24 h, the number of preys consumed was recorded under a stereomicroscope.
Identification of functional response type
To determine the functional response type, logistic regression of the proportion of prey consumed (Na/N0) as a function of initial density (N0) was fitted using a polynomial model (Juliano 2001). A significant negative linear coefficient (P1 < 0) was interpreted as evidence of a type II functional response.
Because preys were confined to leaf discs and not replaced during the 24-h period, prey depletion was expected. Therefore, attack rate (a) and handling time (Th) were estimated using Rogers' Random Predator Equation (RRE) (Rogers 1972):
\[N_a=N_0\left[1-\exp \left(-a\left(T-T_h N_{a)}\right)\right]\right.\]
where Nₐ is the number of preys consumed, N₀ the number initially offered, a the instantaneous attack rate (h⁻¹), Tₕ the handling time (h prey⁻¹), and T the total experimental period (24 h), using maximum likelihood estimation implemented in R with the frair package (frair_fit function). The Quasi-Newton algorithm was applied for optimisation. Parameter uncertainty was assessed through 1,000 bootstrap iterations, yielding 95% confidence intervals.
The choice of RRE is supported by empirical evidence indicating its superior accuracy under finite-prey conditions typical of acarological assays (Ferreira et al. 2022; Fu et al. 2021; Sentis et al. 2012).
Statistical analysis
As prey consumption data are discrete counts and typically exhibit overdispersion, analyses were conducted using generalised linear mixed models (GLMMs) with a negative binomial distribution and log-link function. This modelling approach represents a robust and appropriate framework for handling overdispersed count data, improving parameter estimation and inference reliability compared to traditional linear models.
For each prey stage, the GLMM included:
\[\text { Consumption }_{i j} \sim N B\left(\mu_{i j}, \quad \log \left(\mu_{i j}\right)=\beta_0+\beta_1 \text { Density }_j+(1 / \text { Day })\right.\]
where ''Day'' is a random effect accounting for temporal replication. Model significance was assessed via likelihood-ratio tests. When density had a significant effect, post hoc comparisons were performed using estimated marginal means (EMMeans) with Tukey correction (α = 0.05). All statistical analyses were carried out in R 4.3.0. (R Core Team 2023).
Results
Functional response of A. limonicus to T. urticae eggs
Download as
Prey density (eggs/arena)
Mean number of eggs consumed (eggs/female/day)
5
2.8 ± 0.47 b
10
4.4 ± 0.99 b
20
8.4 ± 0.39 a
30
9.8 ± 0.16 a
40
11.2 ± 0.07 a
The number of T. urticae eggs consumed by adult females of A. limonicus increased significantly with prey density (GLM, χ² = 56.42, P < 0.001). Mean consumption ranged from 2.8 ± 0.65 eggs/female/day at the lowest density (5 eggs) to 11.2 ± 0.78 eggs/female/day at the highest density (40 eggs) (Table 1). Logistic regression analysis of the proportion of prey consumed as a function of initial density yielded significant negative first-order terms for both eggs and mobile stages. This confirms a Holling Type II functional response for A. limonicus, characterized by a consumption rate that decelerates as prey density increases.
Parameter estimation using Rogers' Random Predator Equation (RRE) yielded a robust representation of predator–prey dynamics under finite-prey conditions. The estimated attack rate (a) and handling time (Tₕ) were 1.52 h⁻¹ (95% CI: 1.21–1.89) and 0.050 h (95% CI: 0.042–0.061), respectively, corresponding to a theoretical maximum consumption rate of approximately 20 eggs per day (24 h/Tₕ). Residual diagnostics indicated an adequate model fit (Shapiro–Wilk, P = 0.37; R² = 0.97), supporting the suitability of the RRE model for describing the predator's functional response.
The predation rate increased sharply with prey density up to 20 eggs/arena, after which it reached a plateau, indicating predator satiation (Fig. 1).

The increasing consumption rate with prey density followed by a plateau suggests a satiation effect, consistent with type II functional responses previously reported for other phytoseiid species such as Amblyseius swirskii (Athias-Henriot) fed on Eotetranychus frosti (McGregor) (Tetranychidae) and Cenopalpus irani Dosse (Tenuipalpidae) (Bazgir et al. 2020), and P. persimilis (Sulek et al. 2023) and N. californicus on T. urticae (Nouri-Miri et al. 2025).
Functional response of A. limonicus to mobile stages of T. urticae
Download as
Prey density (mobile stages/arena)
Mean number of prey consumed (individuals/female/day)
1
1.0 ± 0.28 c
2
1.6 ± 0.25 c
4
2.4 ± 0.54 bc
8
3.2 ± 0.37 b
16
5.2 ± 0.54 a
A similar trend was observed when A. limonicus was offered mobile stages (larval and protonymph) of T. urticae. Consumption of larvae and protonymphs also increased significantly with density (GLMM: χ² = 61.23, P < 0.001), ranging from 1.6 ± 0.28 to 5.2 ± 0.40 individuals/female/day at densities of 2 and 16 preys, respectively, forming three categories: low: 2 preys; intermediate: 4–8 preys; and high: 16 preys (Table 2).
Model fitting using the RRE indicated an attack rate (a) of 0.80 h⁻¹ (95% CI: 0.63–0.99) and a handling time (Tₕ) of 0.215 h (95% CI: 0.178–0.259), resulting in a predicted maximum consumption of 4.7 mobile prey/day (Fig. 2). Residual diagnostics confirmed an adequate model fit (R² = 0.98).
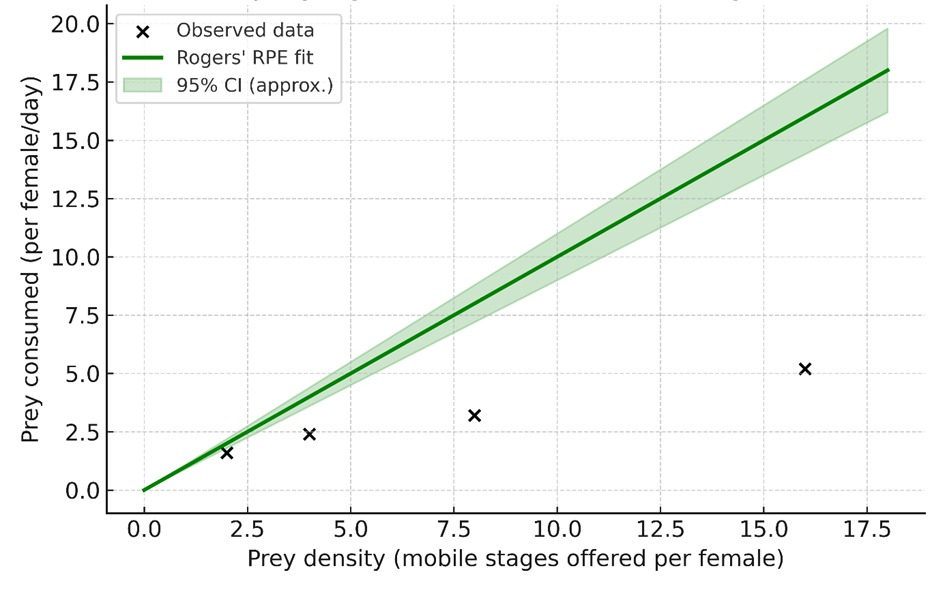
The relationship between prey density and prey consumed was also best described by a quadratic model (R² = 0.988), consistent with a type II functional response (Figure 2). The markedly higher Tₕ and lower a relative to the egg stage suggest that prey mobility reduces capture efficiency and increases manipulation time. Similar constraints associated with motile prey have been observed for N. californicus and P. persimilis feeding on T. urticae larvae and protonymphs (Fathipour et al. 2018; Zheng et al. 2017).
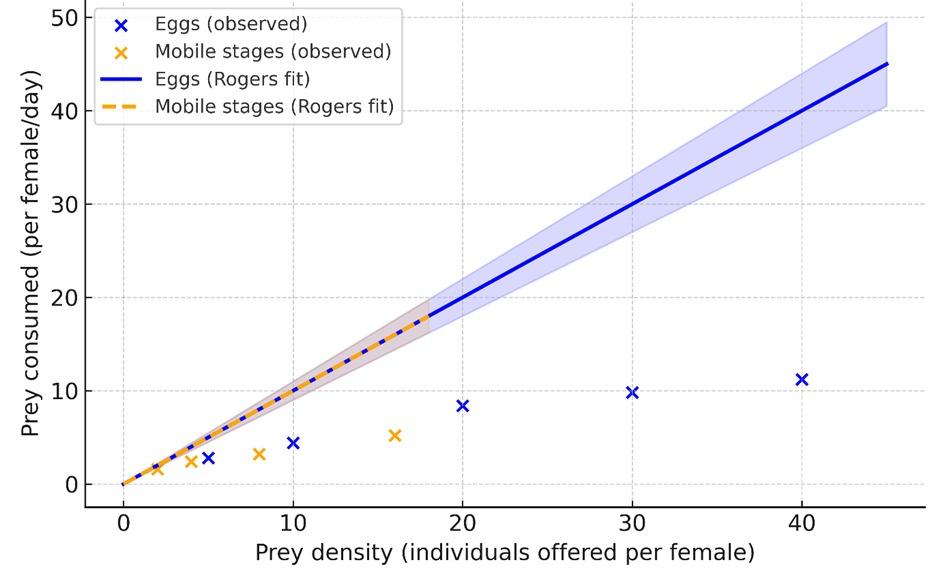
The predator exhibited markedly higher efficiency on eggs than on mobile stages, with a significantly higher a and lower Th (Table 3). Thus, females of A. limonicus exhibited a significantly higher attack rate and shorter handling time when preying on eggs than on mobile stages, resulting in a maximum theoretical consumption approximately four times greater for eggs (20.0 vs. 4.7 prey/day).
Download as
Prey stage
Attack rate (a) (h⁻¹)
95 % CI
Handling time (Tₕ) (h)
95 % CI
Maximum consumption (T/Tₕ) (prey/day)
R²
Functional response type
Eggs
1.52
1.21 – 1.89
0.05
0.042 – 0.061
20
0.97
Type II
Mobile stages
0.8
0.63 – 0.99
0.215
0.178 – 0.259
4.7
0.98
Type II
This indicates a higher predation and greater foraging efficiency for immobile stages, likely due to lower escape potential and reduced handling complexity. The functional response curves for both prey stages displayed a rapid increase in consumption at low prey densities, followed by an asymptotic plateau typical of type II predators (Fig. 3). These results corroborate the saturation behaviour predicted by the RRE model, wherein prey consumption increases rapidly at low densities but reaches an upper asymptote at higher densities due to the time required for handling and digestion.
Discussion
The functional response observed for A. limonicus corresponds to the Type II pattern commonly reported for phytoseiid mites, indicating a rapid increase in consumption at low prey densities followed by saturation due to handling constraints (Fantinou et al. 2012; Sulek et al. 2023). This response suggests that the predator can exert measurable predation pressure under controlled conditions, although its regulatory capacity is inherently limited at higher prey densities (Holling 1959).
Re-estimating the parameters with Rogers' Random Predator Equation (RRE) provided a more realistic representation of predation dynamics under conditions of finite prey availability. Unlike Holling's disc equation, which assumes a constant prey supply, the RRE explicitly accounts for prey depletion during the experiment. Using this model and incorporating bootstrap confidence intervals, the estimated parameters better reflect the underlying biological processes governing the interaction (Papanikolaou et al. 2021; Rogers 1972).
The comparison between prey stages showed that the attack rate on eggs was nearly twice that observed for mobile forms, whereas the handling time for eggs was more than four times shorter. These findings highlight the strong influence of prey mobility and behaviour on predator efficiency. Immobile eggs are readily located and manipulated, requiring minimal pursuit, while mobile instars can evade predation attempts and demand considerably longer handling periods. Similar stage-dependent differences have been reported for A. swirskii and N. californicus preying on T. urticae (Bazgir et al. 2020; Elmoghazy et al. 2024; Zheng et al. 2017), supporting the general rule that the energy expenditure of predators increases with prey motility.
The higher predation rate on eggs also has important ecological and applied implications. Egg predation interrupts the pest's reproductive cycle, reducing the emergence of mobile stages and thus constraining population growth. This pre-emptive feeding behaviour is consistent with the regulatory mechanisms observed in other predatory mites, where early-stage predation has a disproportionate effect on lowering the pest's intrinsic rate of increase (rₘ) (Fathipour et al. 2018). By targeting immobile stages, A. limonicus effectively limits generational turnover, an advantage particularly relevant in controlled environments such as greenhouses where T. urticae develops rapidly under optimal microclimatic conditions.
From a comparative perspective, the predation capacity observed in this study remains lower than that reported for specialist predators such as P. persimilis, which can consume approximately 20–30 spider mite eggs per day under comparable laboratory conditions (Fathipour et al. 2018; Sulek et al. 2023). This difference reflects fundamental ecological strategies exhibited by some specialist predators highly adapted to exploit web-producing spider mites and can penetrate dense webbing, whereas generalist predators such as A. limonicus typically exhibit broader diets but reduced efficiency on this prey type. Consequently, the consumption rate showed by A. limonicus should be interpreted as moderate predation capacity rather than strong suppression potential, particularly under field conditions where prey accessibility is further constrained.
Environmental conditions, however, can modulate functional parameters. Temperature and humidity directly influence predator metabolic activity, mobility, and digestion rate (Davidson et al. 2021). In A. swirskii, attack rates typically increase with temperature up to an optimum range (26–28 °C) before declining at higher temperatures due to thermal stress and associated reductions in predator efficiency (Elmoghazy et al. 2024). Given that the present study was conducted under controlled laboratory conditions (25 ± 2 °C; 65 ± 5% RH), the estimated parameters may differ under the fluctuating thermal and humidity regimes characteristic of field environments. Future research should therefore incorporate temperature and moisture gradients to develop temperature-dependent functional response models, which would enhance the predictive accuracy of biological control performance under real-world conditions.
Besides, plant architecture also plays a decisive role in predator efficiency. The smooth rose leaves used here could have provided minimal physical barriers, facilitating predator movement and prey detection. In natural systems, however, leaf pubescence, trichome density, and wax layers can significantly reduce prey accessibility. For instance, Typhlodromalus aripo De Leon on cassava exhibits lower encounter rates on pubescent leaves compared to glabrous varieties due to interference from trichomes and exudates (Famah Sourassou et al. 2025). Therefore, extrapolation of laboratory-derived parameters to field or greenhouse systems must be approached cautiously, recognising that habitat complexity and microtopography can alter search and handling dynamics.
While the results confirm a predatory capacity, they represent only the functional component of the predator's regulatory potential. Long-term pest suppression depends equally on the numerical response, defined as the predator's ability to increase in population size through reproduction as prey abundance rises (Fathipour et al. 2020). Integrating such life-table parameters with functional and numerical response models would provide a more comprehensive assessment of its population-level impact and long-term regulatory capacity (Ferreira et al. 2022; Rondoni et al. 2024).
From an applied perspective, A. limonicus should not be considered a standalone biological control agent for T. urticae. Instead, its ecological role is better understood within a complementary framework, as this generalist predator can establish early in the crop, suppressing low-density populations and acting synergistically with specialist predators to achieve effective control during pest outbreak (Yang and Zhang 2025; Yang et al. 2025). However, large-scale implementation will require evaluating compatibility with selective acaricides, interactions with other natural enemies, and persistence within commercial cropping systems across diverse environmental conditions.
Conclusion
The present study provides experimental evidence that Amblydromalus limonicus exhibits a Holling type II functional response towards both eggs and mobile stages of Tetranychus urticae, as accurately described by Rogers' Random Predator Equation. The higher attack rate and shorter handling time on eggs demonstrate that this predator is particularly effective against the immobile stage of the pest, thereby disrupting its reproductive cycle and reducing potential population outbreaks. These results underline the species' capacity for density-dependent regulation and its potential integration as a complementary and preventive agent within integrated biological control programmes, especially in protected crops such as roses. By refining the estimation of functional parameters under finite-prey conditions, this research strengthens the predictive basis for modelling predator–prey dynamics in acarology. Future studies linking these functional parameters to numerical response and environmental variability will be essential to validate A. limonicus as a reliable component of integrated mite management strategies.
References
- Bazgir, F., Shakarami, J., Jafari, S. 2020. Functional response of the predatory mite Amblyseius swirskii (Acari: Phytoseiidae) to Eotetranychus frosti (Tetranychidae) and Cenopalpus irani (Tenuipalpidae). Acarologia. 60 (1): 30-39. https://doi.org/10.24349/acarologia/20204359
- Davidson, A.T., Hamman, E.A., McCoy, M.W., Vonesh, J.R. 2021. Asymmetrical effects of temperature on stage-structured predator-prey interactions. Funct. Ecol. 35 (5): 1041-1054. https://doi.org/10.1111/1365-2435.13777
- Elmoghazy, M.M.E., Elsherbini, D.M.A., Mashlawi, A.M., Ibrahim, A.M., El-Mansi, A.A., El-Sherbiny, M. 2024. Implications of temperature and prey density on predatory mite Amblyseius swirskii (Acari: Phytoseiidae) functional responses. Insects. 15: 444. https://doi.org/10.3390/insects15060444
- Famah Sourassou, N., Hanna, R., Agboka, K., Negloh, K., Tchabi, A. 2025. Beyond field success: insights into functional and numerical response of the predatory mite Typhlodromalus aripo and the role of cassava exudate in predatory performance. Int. J. Adv. Res. 13 (8): 1252-1264 https://doi.org/10.21474/IJAR01/21615
- Fantinou, A.A., Baxevani, A., Drizou, F., Labropoulos, P., Perdikis, D., Papadoulis, G. 2012. Consumption rate, functional response and preference of the predaceous mite Iphiseius degenerans to Tetranychus urticae and Eutetranychus orientalis. Exp. Appl. Acarol.58 (2): 133-144. https://doi.org/10.1007/s10493-012-9557-6
- Fathipour, Y., Karimi, M., Farazmand, A., Talebi, A.A. 2018. Age-specific functional response and predation capacity of Phytoseiulus persimilis (Phytoseiidae) on the two-spotted spider mite. Acarologia. 58 (1): 31-40. https://doi.org/10.24349/acarologia/20184225
- Fathipour, Y., Maleknia, B. 2016. Mite Predators. In: Gavkare, O. (Ed.). Ecofriendly Pest Management for Food Security. Oxford: Academic Press. p. 329-366. https://doi.org/10.1016/B978-0-12-803265-7.00011-7
- Fathipour, Y., Maleknia, B., Bagheri, A., Soufbaf, M., Reddy, G.V.P. 2020. Functional and numerical responses, mutual interference, and resource switching of Amblyseius swirskii on two-spotted spider mite. Biol. Control. 146: 104266. https://doi.org/10.1016/j.biocontrol.2020.104266
- Ferreira, C.T., Noronha, A.C. da S., Souza Neto, E.P., De Oliveira, R.P., Lins, P.M.P., Batista, T.F.V. 2022. Functional and numerical responses of the predatory mite Amblyseius aerialis (Acari: Phytoseiidae) to Aceria guerreronis (Acari: Eriophyidae). Acarologia. 62 (1): 27-35. https://doi.org/10.24349/w600-25ar
- Fu, X., Liu, Q., Liu, J., Meng, R. 2021. Functional response of Amblyseius andersoni and Neoseiulus neoreticuloides (Acari: Phytoseiidae) to adults of the wolfberry gall mite Aceria pallida (Acari: Eriophyoidae). Syst. Appl. Acarol. 26 (4): 809-817. https://doi.org/10.11158/saa.26.4.11
- Holling, C.S., 1959. The components of predation as revealed by a study of small-mammal predation of the European pine sawfly. Can. Entomol. 91 (5): 293-320. https://doi.org/10.2351/1.5058368
- Juliano, S.A. 2001. Nonlinear curve fitting: Predation and Functional Response curves. In: Schneider, S.S., Gurevitch, J. (Eds.). Design and Analysis of Ecological Experiments. Oxford: Oxford University Press. p. 178-196. https://doi.org/10.1093/oso/9780195131871.003.0010
- Lee, M.H., Zhang, Z.Q. 2018. Assessing the augmentation of Amblydromalus limonicus with the supplementation of pollen, thread, and substrates to combat greenhouse whitefly populations. Sci. Rep 8: 12189. https://doi.org/10.1038/s41598-018-30018-3
- Lee, S.Y., Shin, J.Y., Kwon, D.H., Xu, J., Kim, J.H., Ahn, C.H., Jang, S., Kwon, O.H., Lee, H.J., Kim, W.H. 2022. Preventing scattering of Tetranychus urticae in Rosa hybrida through dsCOPB2 expression. Sci. Hort. 301: 111113. https://doi.org/10.1016/j.scienta.2022.111113
- Lemaire, É., McCune, F., Roy, M., Bourgeois, G., Fournier, V. 2025. Efficacy of releases of phytoseiid mites (Acari: Phytoseiidae) on spider mite control on raspberry in high tunnel production, and implication on control strategy through a population model of Tetranychus mcdanieli (Acari: Tetranychidae). Biol. Control. 206: 105799. https://doi.org/10.1016/j.biocontrol.2025.105799
- McMurtry, J.A., Croft, B.A. 1997. Life-styles of phytoseiid mites and their roles in biological control. Annu. Rev. Entomol. 42: 291-321. https://doi.org/10.1103/PhysRevE.65.066410
- Nouri-Miri, M., Kheradmand, K., Saboori, A., Fathipour, Y. 2025. Age-gender-specific functional and age-specific numerical responses of Neoseiulus californicus (Acari: Phytoseiidae) on two-spotted spider mite. Exp. Appl. Acarol. 94: 5. https://doi.org/10.1007/s10493-024-00969-6
- Papanikolaou, N.E., Kypraios, T., Moffat, H., Fantinou, A., Perdikis, D.P., Drovandi, C. 2021. Predators' functional response: statistical inference, experimental design, and biological interpretation of the handling time. Front. Ecol. Evol. 9: 740848. https://doi.org/10.3389/fevo.2021.740848
- Pazmiño, P., Lema, G., Mendoza, D., Velástegui, G., Vásquez, C. 2018. Parámetros biológicos de Tetranychus urticae Koch (Acari: Tetranychidae) alimentados sobre dos cultivares de fresa en Ecuador. Bioagro. 30 (3): 229-234
- R Core Team. 2023. R: A language and environment for statistical computing [Computer software]. R Foundation for Statistical Computing, 2023. [8 Oct 2025]. Available on: https://www.r-project.org/.
- Rivero Guerra, A.O. 2021. Caracterización de los viveros de Pelileo, Tunguragua, Ecuador. RIIARn. 8 (1): 62-82 https://doi.org/10.53287/tpjc7586ek97g
- Rogers, D. 1972. Random search and insect population models. J. Anim. Ecol. 41 (2): 369-383. https://doi.org/10.2307/3474
- Rondoni, G., Collatz, J., Jonsson, M., Rennstam Rubbmark, O., Riddick, E.W., Schmidt, J.M., Brodeur, J. 2024. Recent advances in characterizing trophic connections in biological control. Biol. Control. 199: 105656. https://doi.org/10.1016/j.biocontrol.2024.105656
- Schoeller, E.N., McKenzie, C.L., Osborne, L.S. 2022. Chilli thrips rose management using an Amblyseius swirskii or Amblydromalus limonicus (Acari: Phytoseiidae) pepper banker plant. J. Appl. Entomol. 146: 1281-1292. https://doi.org/10.1111/jen.13066
- Sentis, A., Hemptinne, J.L., Brodeur, J. 2012. Using functional response modeling to investigate the effect of temperature on predator feeding rate and energetic efficiency. Oecologia. 169 (4): 1117-1125. https://doi.org/10.1007/s00442-012-2255-6
- Sulek, N., Döker, I., Saboori, A., Cakmak, I. 2023. Prey consumption capacity and functional response of Phytoseiulus persimilis (Acari: Phytoseiidae) feeding on Tetranychus urticae (Acari: Tetranychidae) on different cotton Varieties. Acarologia. 63 (3): 665-675. https://doi.org/10.24349/o7gh-1c6y
- Tiftikçi, P., Kök, Ş., Kasap, İ. 2020. Biological control of twospotted spider mites [Tetranychus urticae Koch (Acari: Tetranychidae)] using Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseidae) at different ratios of release on field-grown tomatos. Biol. Control. 151: 104404. https://doi.org/10.1016/j.biocontrol.2020.104404
- Urbaneja-Bernat, P., Jaques, J.A. 2025. The impact of global change on the intraguild predation among three predatory mites of Tetranychus urticae in citrus orchards. J. Pest Sci. 98: 2363-2374. https://doi.org/10.1007/s10340-025-01943-w
- Van Leeuwen, T., Tirry, L., Yamamoto, A., Nauen, R., Dermauw, W. 2015. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pest. Biochem. Physiol. 121: 12-21. https://doi.org/10.1016/j.pestbp.2014.12.009
- Wang, J., Zhang, K., Li, L., Zhang, Z. 2024. Development and reproduction of four predatory mites (Parasitiformes: Phytoseiidae) feeding on the spider mites Tetranychus evansi and T. urticae (Trombidiformes: Tetranychidae) and the dried fruit mite Carpoglyphus lactis (Sarcoptiformes: Carpoglyphidae). Syst. Appl. Acarol. 29 (2): 269-284. https://doi.org/10.11158/saa.29.2.7
- Xiao, Y., Osborne, L.S., Chen, J., McKenzie, C.L. 2013. Functional responses and prey-stage preferences of a predatory gall midge and two predacious mites with twospotted spider mites, Tetranychus urticae, as host. J. Insect Sci. 13: 8. https://doi.org/10.1673/031.013.0801
- Yang, Y., Zhang, K., Zhang, Z.Q. 2025. Predatory mites Amblydromalus limonicus and Amblyseius herbicolus (Acari: Phytoseiidae) as potential biocontrol agents of Eotetranychus sexmaculatus (Acari: Tetranychidae) in avocado: examining predation on different prey life stages. J. Econ. Entomol. 118 (3): 1335-1343. https://doi.org/10.1093/jee/toaf036
- Yang, Y., Zhang, Z.-Q. 2025. Preliminary shade-house evaluations of Amblydromalus limonicus and Amblyseius herbicolus (Acari: Phytoseiidae) as biological control agents against Eotetranychus sexmaculatus (Acari: Tetranychidae) on avocado leaves. Syst. Appl. Acarol. 30 (6): 1085-1088. https://doi.org/10.11158/saa.30.6.11
- Zheng, Y., Clercq, P. De, Song, Z.W., Li, D.S., Zhang, B.X. 2017. Functional response of two Neoseiulus species preying on Tetranychus urticae Koch. Syst. Appl. Acarol. 22 (7): 1059-1068. https://doi.org/10.11158/saa.22.7.13



2025-11-20
Date accepted:
2026-04-18
Date published:
2026-05-11
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Santana, Rita; Pérez Salinas, Ruth; Mangui, José; Ortiz-Tirado, Paúl; Telenchana, Norma and López-Villacís, Cristina
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)