



The water mite species Piona conglobata (Koch, 1837), Piona punctata (Neuman, 1875), and Piona lacustricola K.Viets, 1949 (Acari, Hydrachnidia, Pionidae) in Norway
Gerecke, Reinhard  1
and Stur, Elisabeth
2
1
and Stur, Elisabeth
2
1University of Tübingen, Faculty of Science, Department of Evolution and Ecology, DE-72074 Tübingen, Germany; Arachnologie, Senckenberg Forschungsinstitut, Mertonstr. 17-21, DE-60325 Frankfurt, Germany.
2Department of Natural History, NTNU University Museum, Norwegian University of Science and Technology, NO-7491 Trondheim, Norway.
2026 - Volume: 66 Issue: 2 pages: 455-468
https://doi.org/10.24349/sro5-jv62Original research
Keywords
Abstract
Introduction
Species of the highly diversified water mite genus Piona Koch, 1842 are present worldwide except Antarctica. All Piona species inhabit standing waters, and most of them prefer large water bodies where they are found mostly in the macrophyte belt (Pieczinsky 1976), although some specialists extend to the profundal (K. Viets 1931) or, in large populations, into the planktonic zone (Schwoerbel 1972). Under favourable conditions, Piona species may undergo spectacular mass developments as reported by Punčochař and Hrbáček (1991).
Scandinavia is an important area for diversity studies in the genus. Of the 18 species so far reported from Norway (Gerecke et al. 2022), more than 50% were originally described from that area. Molecular studies (Stålstedt et al. 2013) have revealed high levels of hidden diversity within the genus. According to Gerecke et al. (2025), the Norwegian nominate species comprise at least seven additional cryptic species. The aim of this paper is to re-establish two of these taxa by revising the species of the Piona conglobata aggregate.
In his definition of a Piona conglobata species group, Smith (1976) restricted his treatment to specimens and species of the New World and did not address questions concerning separation and definition of Old World species – besides P. conglobata, the only Palaearctic species listed there is P. discrepans Koenike, 1895. As discussed below ₋ and as also suggested by sequence data available in BOLD ₋ North American populations published under the name Piona conglobata most probably represent a distinct species, Piona wisconsinensis Marshall, 1935. The aim of the present study is not to discuss the Piona conglobata group as defined by Smith (1976) but has in focus the West-Palaearctic P. conglobata-like species. We consider this assemblage ₋ hereafter referred to as the ''Piona conglobata aggr.'' ₋ to represent a monophyletic group, defined in adults by the following diagnostic features: Both sexes: Integument lineated; dorsum widely membranous, with at least one pair of platelets in anterior part; I-II-legs often with a more or less distinct sexual dimorphism, swimming setation: I-leg-5, 2; II-leg-4/5, 2/4; III-leg-4/5, 3-4/5-6; IV-leg-5/6, 3-4/5-7; genital field with 10-30 pairs of Ac; palp rather stout, P-4 with two to five pointed, seta-bearing ventral projections. Males: III-leg-6 with a short, curved claw; IV-leg-4 with a central incision flanked by strong and stout setae, further similar setae at distal segment margin; genital field broadly fused to Cx-IV, gonopore flanked by a pair of Ac, in the area of fusion between genital field and Cx-IV, posterior to gonopore no groove, a narrow step delimitating a flattened medial area. Females: Gonopore length 130-320, acetabula scattered in bowed lines in the membranous area flanking gonopore; anterior- and posteriormost Ac on setae-bearing platelets, these Ac, occasionally also others, fused with basal sclerites of neighbouring Ac, but never lying on continuous, crescent-shaped plates.
Material and methods
The mites examined in this study were collected in 2022 and 2023 from standing waters using hand nets, as part of the NTNU University Museum project ''Calcareous mites and midges» (Gerecke et al. 2025 – see there for details concerning methods applied for molecular analysis). Barcode sequences and associated metadata are publicly accessible in the dataset ''DS-CMMHY Water mites in calcareous water bodies» (Stur et al. 2025), available at dx.doi.org/10.5883/DSCMMHY. All examined material is housed in the scientific collections of the NTNU University Museum (NTNU-VM).
Specimens selected for morphological investigation were dissected and mounted on slides in Hoyer's fluid or glycerine jelly following the protocol outlined in Gerecke et al. (2016 – see there also for the morphological terminology used here). All measurements are presented in µm. Unless otherwise specified, length of appendage segments is given as dorsal length.
Systematics
Family Pionidae
Genus Piona Koch, 1842
Piona conglobata (Koch, 1836)
No holotype defined. Type locality: Ponds near Regensburg, Germany.
Material examined – Norway
Viken, Ullensaker, Elstad landskapsvernområde, lake Transjøen, N60.21119°, E11.13815°, 172 m a.s.l., 5 June 2022, leg. Gaute Kjærstad (GK) & Reinhard Gerecke (RG), 1 ♀ on slide, NTNU-VM 291073. Trøndelag, Nærøysund, lake Staverengvatnet, N64.82626° E11.30524°, 6 m a.s.l., 11 Aug. 2022, leg. GK & Karstein Hårsaker (KH), 1♂ on slide, NTNU-VM 291068, BOLD Specimen id HYDCA858. Trøndelag, Nærøysund, lake Sørvatnet, N64.91816°, E11.29508°, 1 m a.s.l., 12 Aug. 2022, leg. GK & KH, 2♀♀ on slide, NTNU-VM 291070 (from this site, 1♂ was successfully barcoded [BOLD specimen id HYDCA873], voucher lost). Nordland, Vega, lake Langklubbvalen, N65.698°, E11.85402°, 1 m a.s.l., 17 July 2023, leg. GK & KH, 1 ♀ on slide, NTNU-VM 291071, BOLD Specimen HYDCA975.
The specimens for which a DNA barcode is available are assigned to BIN BOLD:ACS0245. The latter BIN includes sequences from Norway, Russia, and the Netherlands available in the BOLD database.

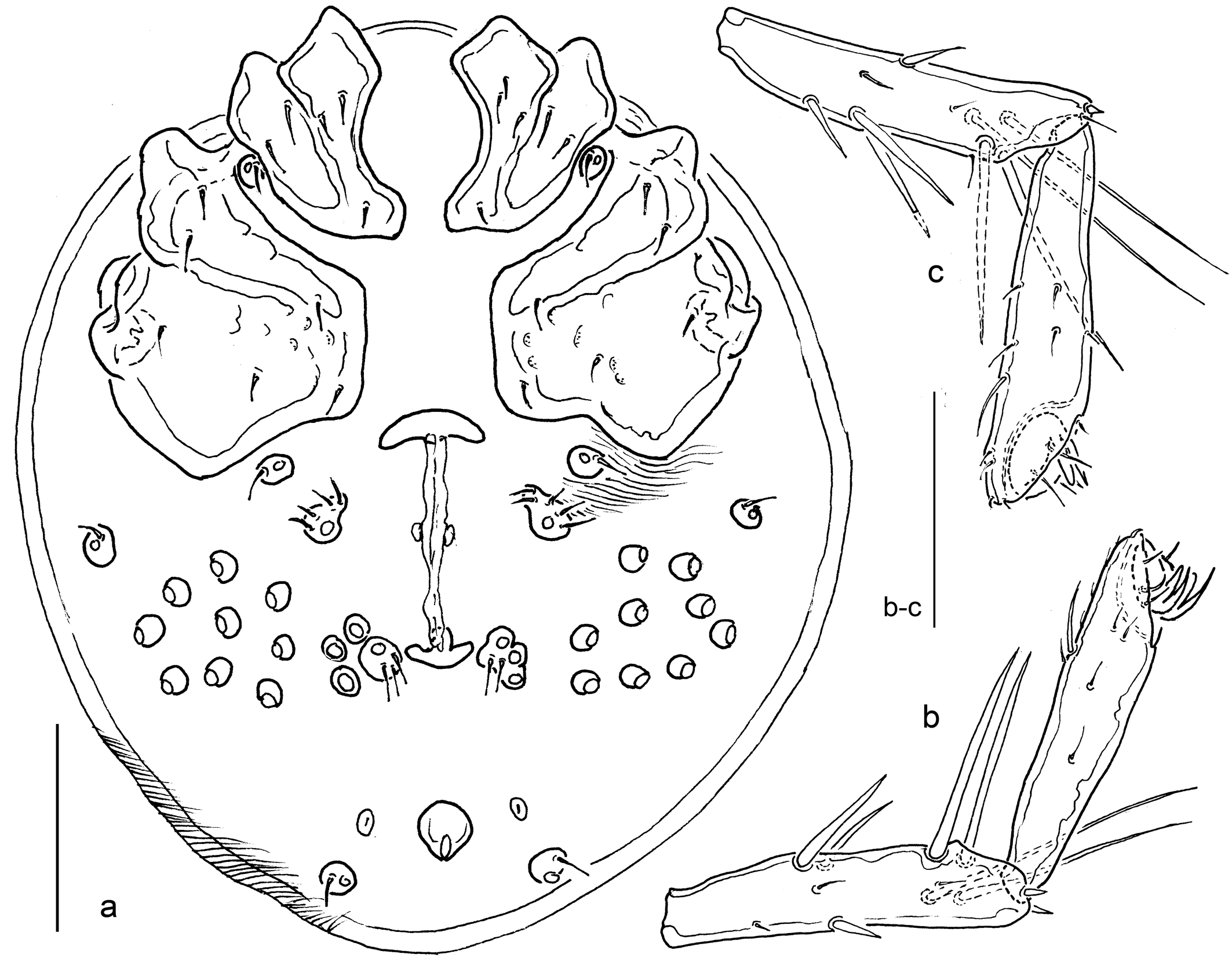
Diagnosis
Both sexes — Ac small (diameter 14-15); P-4, in addition to the two pointed ventral seta-bearing extensions, typically with two further projections. I-/II-legs rather slender, without remarkable sexual dimorphism.
Males — III-leg-6 relatively long (L/H ratio III-leg-5/6, 1.2); setal pores flanking excretory pore fused to genital field.
Females — Genital field with anteriormost Ac on a setae-bearing platelet, posteromedial Ac on a setae-bearing platelet occasionally including 1-2 neighbouring Ac; all remaining Ac isolated from each other.
Description
Both sexes — Colour of the investigated material not documented. Following the original description (Koch 1837) idiosoma and appendages green, body contents reddish-olive brown, coxal area reddish, the narrow Y-shaped excretory organ (''Gabelstreif'') intense yellow. Later authors gave slightly different data: occasionally yellowish – probably referring to immature specimens (Piersig 1900, Soar & Williamson 1929, K.Viets 1936); terminal segments of I-II-L brownish-red (K. Viets 1936). Integument lineated. Genital field with 22-27 Ac (on one side 11-14). Leg segments 2-5 with scattered long stiff setae (probably all involved in swimming); longer ''swimming setae'' (n in parentheses) arranged distally in lines on the posterior segment surface of I-leg-5 (2), II-leg-4/5 (2/4), III-leg-4/5 (3-4/5-6), IV-leg-5/6 (3-4/5-7). Palp: P-2 thick, dorsal margin strongly bowed, ventral margin nearly straight; P-3 short, ventral margin concave, dorsal and ventral margin converging; P-4 stout, dorsal and ventral margin converging, three to four ventral setae on pointed, distoventrally directed projections standing close together – the distalmost more medially, the most proximal one more laterally and one further projection on the lateral surface, distomedial peg seta rather strong, on the acute-angled distoventral segment edge; P-5 slender and long, with three claw-like apical setae. Chelicera basal segment in central part thick and strongly bowed, proximal end narrowed and pointed, L/H ratio 2.9-3.0, basal segment/claw ratio 2.3-2.6.
Males — (Fig. 1). Idiosoma L/W 510/440. Dorsum with muscle attachment sclerites posterior to the posterior antenniform setae fused to a pair of outwards-directed L-shaped platelets, and one further pair of round muscle attachments in posterior part. Ventral sclerotization consisting of three distinct parts: paired Cx-I+II mL 170, lat L 110, separated medially from each other and posteriorly from Cx-III+IV by membranous integument, Cx-I posteromedial margin bluntly pointed, posterolateral margin extending into posterolaterally directed apodemes. Cx-III+IV medially fused, elongate, L/W 320/230 (ratio 1.4), suture line Cx-III-IV oblique, medially obliterated; triangular projections covering IV-leg insertions. Genital field L (incl. excretory pore)/W 145/340, fused to medial part of posterior margin of Cx-III+IV, with gonopore (L 35) in anterior position at the level of fusion genital plate-Cx-IV, flanked by one pair of Ac and posteriorly followed by a flat depression; the remaining Ac scattered over the transverse, laterally rounded genital plates, total n 22 (right/left 11/11), maximum Ac diameter 15; excretory pore and flanking setal pores fused to the posterior sclerotization of genital field, the flanking glandularia free in the integument. Legs: I-/II-legs-6 rather slender, basal H about 12, dorsal margin in proximal part slightly convex, ventral margin in basal part very slightly concave; III-leg-6 slender and rather long, maximum H in the centre, claw short and rounded; grasping organ of IV-leg-4 with a deep groove in central part, surrounded by strong setae (long in ventral part, gradually shorter versus dorsal part), distal segment end prolonged and ending in a rounded tip, with a further group of strong setae, IV-leg-6 slender, maximum H in proximal part, claw short as in III-leg, but in distal part straight. Leg segment measurements (in parentheses, L/H ratio): I-leg-4, 145 (3.6); I-leg-5, 160 (3.8); I-leg-6, 120 (3.4) – I-leg-6 L max 170 (4.9); II-leg-4, 170 (4.5); II-leg-5, 190 (5.1); II-leg-6, 145 (4.5) – II-leg-6 L max 185 (5.7); III-leg-4, 180 (4.7); III-leg-5, 210 (5.3); III-leg-6, 160 (5.3) – III-leg-6 L max 165 (5.5); ratio III-leg-5 L/III-leg-6 L max 1.3; IV-leg-4, 160 (3.6); IV-leg-5, 190 (5.4); IV-leg-6, 160 (6.7) – IV-leg-6 L max 175 (7.3); L ratio IV-leg-4/5, 0.84; IV-leg-4/6, 1.00; IV-leg-4/6 L max 0.91; IV-leg-5/6, 1.19; IV-leg-5/6 L max 1.09. Mouthparts: Gnathosoma L 160; palp total L 345-333, segments L (L/H ratio, relative L): P-1, 33 (0.62, 9%); P-2, 123 (1.58, 33%); P-3, 63 (1.19, 17%); P-4 (ventral extensions included in H measurements) 103 (2.05, 27%); P-5, 55 (2.20, 15%); L ratio P-2/P-3, 1.96; P-2/P-4, 1.20; P-3/P-4, 0.61.
Females — (Fig. 2). Idiosoma L/W 520-1000/500-900. Dorsum with one pair of elongate platelets posterior to the posterior antenniform setae, and a pair of round muscle attachments in posterior part. Ventral sclerotization consisting of four distinct parts: paired Cx-I+II, mL 180-210, lat L 130-160; Cx-III+IV L 290-330, W 230-265 (L/W ratio 1.2-1.3), suture Cx-III/-IV reaching medial plate margin; genital field L (pregenital sclerite-excretory pore)/W 290-440/320-580, gonopore L 130-250, most Ac lying free in the membranous integument, scattered over paired crescent-shaped areas extending posterolaterally to gonopore, the anteriormost Ac on each side on a triangular platelet together with 4-5 fine setae, on the level of the postgenitale, one Ac on a platelet with 2-3 fine setae, occasionally fused with the basal sclerites of 1-2 neighbouring Ac; total Ac n 23-27 (right/left 11-13/11-14), maximum Ac diameter 14-15. Legs: I-/II-legs-6 slender, basal H about 17, dorsal margin very slightly convex, ventral margin very slightly concave; III-leg-6 and IV-leg-4 without the modifications described for males: III-leg-6 maximum H at the claw groove, claw fine and long as in other legs; IV-leg-4 with subparallel dorsal and ventral margins, dorsodistally not prolonged, IV-leg-6 slender, maximum H near claw groove, claw shorter than in I-III-legs; segment measurements (in parentheses, L/H ratio): I-leg-4, 135-140 (3.4-3.5); I-leg-5, 145-155 (4.1-4.4); I-leg-6, 120-125 (3.6-4.0) – I-leg-6 L max 160 (4.6-5.3); II-leg-4, 160-165 (4.0-4.4); II-leg-5, 175-180 (5.0-5.1); II-leg-6, 130-143 (3.7-4.1) – II-leg-6 L max 165-180 (4.7-5.1); III-leg-4, 170-175 (4.3-5.0); III-leg-5, 185-203 (5.3-5.8); III-leg-6, 135-150 (4.5-5.5) – III-leg-6 L max 170-188 (5.7-6.8); ratio III-leg-5 L/III-leg-6 L max 1.1; IV-leg-4, 200-235 (5,2-5.3); IV-leg-5, 205-245 (5.9-6.3); IV-leg-6, 170 (7.6-8.5) – IV-leg-6 L max 180-185 (8.0-9.3); L ratio IV-leg-4/5, 0.95-0.98; IV-leg-4/6, 1.18-1.24; IV-leg-4/6 L max 1.11-1.14; IV-leg-5/6, 1.21-1.29; IV-leg-5/6 L max 1.14-1.19. Mouth parts: Gnathosoma L 185; palp total L 343-365, segments L (L/H ratio, relative L): P-1, 28-35 (0.58-0.63, 8%); P-2, 120-140 (1.63-1.66, 33-36%); P-3, 48-73 (0.9-1.1, 14-17%); P-4, 95-100 (2.1-2.2, 27-28%); P-5, 50-53 (1.8-2.3, 14-15%); L ratio P-2/P-3, 1.9-2.6; P-2/P-4, 1.2-1.2; P-3/P-4, 0.5-0.6.
Discussion
The tiny figure of a male in the original description of Piona conglobata s. str. (Koch, 1837, Fig. 9.7) and the short text give a detailed description of the coloration of the species (see above). Furthermore, a thickening of I-/II-leg-6 and rather large lateral eyes are mentioned, but no details are given concerning the shape of palps and genital field. However, an important detail also observed in the specimens reported here from Norway can be deduced from the Koch's figure: segments III-leg-6 are only slightly shorter than the preceding III-leg-5. In addition, both sexes are characterized by rather small Ac, and the presence of additional ventral projections on P-4. Notwithstanding the lack of type material, we follow the consensus developed, e.g., by Koenike (1907), K.Viets (1936) and Lundblad (1962) to consider the presence of more than two ventral projections on P-4 as typical for Piona conglobata. This perspective is further supported by the distinct character state of the elongate male III-leg-6 described here. Under these aspects, the species is quite well defined and does not require a neotype designation.
In his comprehensive revision of Swedish Piona species, Lundblad (1962) considered the two species reinstated below – P. punctata Neuman, 1875, first described from Gotland (Neuman 1875) and P. conglobata lacustricola K.Viets, 1949, first described by Halbert (1944) from Ireland under the preoccupied name P. lacustris, as ''unsignificant forms''. The diagnostic features of Piona conglobata wisconsinensis Marshall, 1935, a further subspecies with elevated Ac n (> 50), and in addition characterized by an ''upturned margin where the male genital plates meet the posterior angle of the fourth epimera'' (Marshall 1935), were considered by Lundblad (1962) as unsignificant deviations not valuable for subspecies discrimination. Instead, Ac numbers appear rather stable in our examined material and the American taxon, possibly a separate species, merits to be reconsidered. Another subspecies, P. conglobata rubra Musselius, 1913 with elevated Ac number (ca. 50), described from the Saint Petersburg area (Russia) (Musselius 1913), was completely disregarded by Lundblad (1962). Little is known about its other morphological features. Since no type material is available, this taxon will remain a subspecies incerta. It certainly does not correspond to any of the taxa treated in this study.
Piona punctata (Neuman, 1875)
Piona stuxbergi (Piersig, 1897); P. distermina (Koenike, 1907); P. conjugula (Koenike, 1909): Lundblad 1962
Material examined – Norway
Viken, Drammen, lake Hagatjern near Dalevika, N59.72547°, E10.05901°, 255 m a.s.l., 4 June 2022, leg. GK & RG, 1♂ on slide, NTNU-VM 291049, BOLD Specimen id HYDCA641. Viken, Ullensaker, Nordbytjern landskapsvernområde, lake Nordbytjernet, N60.15588°, E11.15848°, 188 m a.s.l., 5 June 2022, leg. GK & RG, 1 ♀ on slide, NTNU-VM 291050, BOLD Specimen id HYDCA663. Viken, Ullensaker. Elstad landskapsvernområde, lake Hersjøen, N60.21444°, E11.1522°, 158 m a.s.l., 5 June 2022, leg. GK & RG, 6♀♀ in tube, NTNU-VM 291072. Viken, Ringerike, Nordre Tyrifjorden og Storelva naturreservat, Juveren, oxbow, N60.12606°, E10.25533°, 63 m a.s.l., 6 June 2022, leg. GK & RG, 1♂ on slide, NTNU-VM 291051, BOLD Specimen id HYDCA690. Viken, Asker, lake Gjellumvannet, N59.78781°, E10.44327°, 97 m a.s.l., 7 June 2022, leg. GK & RG, 7♀♀ in tube, NTNU-VM 291074. Viken, Asker, lake Finnsrudvannet, N59.8303°, E10.38719°, 176 m a.s.l., 7 June 2022, leg. GK & RG, 6♀♀ in tube, NTNU-VM 291075. Viken, Asker, lake Padderudvannet, N59.82216°, E10.36345°, 186 m a.s.l., 7 June 2022, leg. GK & RG, 3♀♀ in tube, NTNU-VM 291052, 1♀ on slide, NTNU-VM 291053, BOLD Specimen id HYDCA724, 1♀ on slide, NTNU-VM 291060. Viken, Lunner, lake Kalvsjøtjernet, N60.28771°, E10. 56247°, 359 m a.s.l., 8 June 2022, leg. GK & RG, 4♀♀ in tube, NTNU-VM 291076. Viken, Lunner, Galtedalstjerna naturreservat, lake Galtedalstjerna, N60.28883°, E10.47007°, 296 m a.s.l., 9 June 2022, leg. GK & RG, 1♀ on slide, NTNU-VM 291054, BOLD Specimen id HYDCA746, 1♀ on slide, NTNU-VM 291055, BOLD Specimen id HYDCA747, 16♀♀ in tube, NTNU-VM 291056, 1♀ in tube, NTNU-VM 291057, 2♂♂ in tube, NTNU-VM 291214. Viken, Lunner, lake Markatjernet, N60.29092°, E10.4799°, 313 m a.s.l., 9 June 2022, leg. GK & RG, 1♂ on slide, NTNU-VM 291058, BOLD Specimen id HYDCA753, 13♀♀ in tube, NTNU-VM 291061. Innlandet, Gran, lake Jarenvannet, N60.39123°, E10.54421°, 200 m a.s.l., 9 June 2022, leg. GK & RG, 1♀ on slide, NTNU-VM 291059, BOLD Specimen id HYDCA763, 2♀♀ in tube, NTNU-VM 291063. Trøndelag, Frøya, Kalvdalen, pond north, N63.68941°, E8.58363°, 22 m a.s.l., 29 June 2022, leg. GK, 2♀♀ in tube, NTNU-VM 291077. Trøndelag, Frøya, Kalvdalen, pond south, N63.68849°, E8.58485°, 22 m a.s.l., 29 June 2022, leg. GK, 6♀♀ in tube, NTNU-VM 291078. Trøndelag, Nærøysund, lake Staverengvatnet, N64.82626°, E11.30524°, 6 m a.s.l., 11 Aug. 2022, leg. GK & RG, 1♀ in tube, NTNU-VM 291069. Nordland, Evenes, lake Nautåvatnet, N 68.49489°, E16.70889°, 17 m a.s.l., 12 July 2023, leg. GK, 2♀♀ in tube, NTNU-VM 291079. Nordland, Brønnøy, lake Aunvatnet, N65.40108°, E12.60014°, 5 m a.s.l., 16 July 2023, leg. GK & KH, 1♀ in tube, NTNU-VM 291080. Nordland, Vega, lake Olhåksåvatnet, N65.69632°, E11,8888°, 6 m a.s.l., 17 July 2023, leg. GK & KH, 7♀♀ in tube, NTNU-VM 291067. Nordland, Brønnøy, lake Tilremsvatnet, N65.51337°, E12.26493°, 5 m a.s.l., 18 July 2023, leg. GK & KH, 1♀ in tube, NTNU-VM 291082.
Piona distermina type series: SMNH female type 896 = NHRS-GULI000106537; male type 900 = NHRS-GULI000106538, both partly decipherable, ''Seeland [...] Teich, 26.7.95 Koen. leg.'' (type locality in Koenike's list: ''Seeland, Stor Skov b. Frederiksdal, Teich, 29.7.95 Koen. coll.'').
The specimens with available DNA barcodes are assigned to the following BINs: BOLD:AFA2526, BOLD:AFA2527, BOLD:ACS0324.
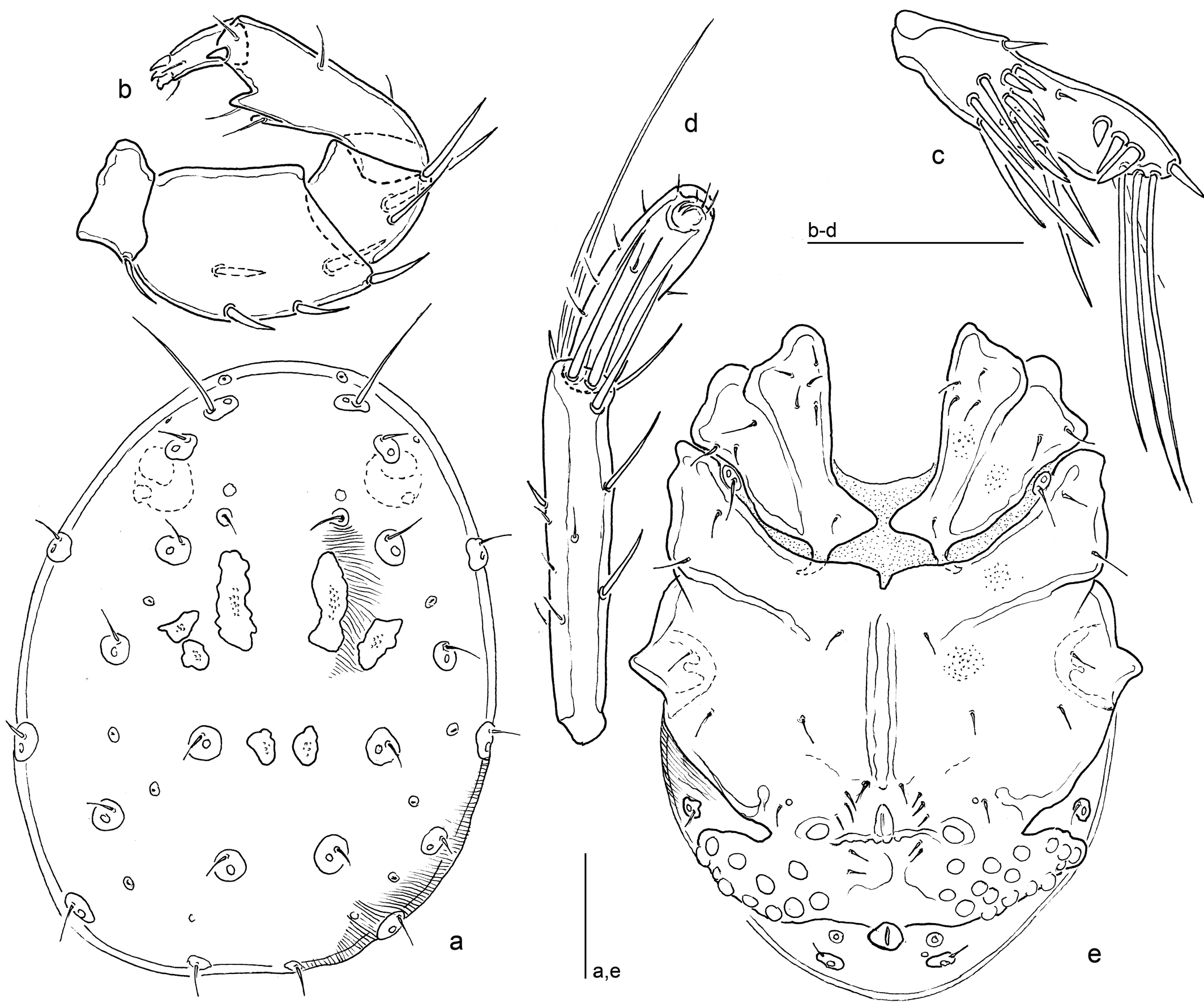

Diagnosis
Both sexes — Ac rather large (diameter 18-25); P-4, in addition to the two-pointed ventral seta-bearing extensions, without further projections.
Males — III-leg-6 strongly shortened (L/H III-leg-5/6, 1.5-1.7); setal pores flanking excretory pore not fused to genital field.
Females — Genital field with anteriormost Ac on a setae-bearing platelet, 3-4 posteromedial Ac fused to a setae-bearing platelet.
Description
Both sexes — Colour of the investigated material not documented; following Neuman (1880) in dorsal view with brown dots separated by extended areas of whitish colour. Integument lineated. Genital field with 24-39 Ac (on one side 11-20). Legs: I-/II-legs-6 thickened, with sexual dimorphism in shape; III-leg-4 and IV-leg-5-6 as described for P. conglobata. Palp: P-2 thick, dorsal margin strongly bowed, ventral margin nearly straight; P-3 short, ventral margin concave, dorsal and ventral margin converging; P-4 stout, dorsal and ventral margin strongly converging, two ventral setae on pointed, distoventrally directed projections – the distal one shifted to the medial surface, the proximal one to the lateral surface – in general, no further projections on this segment, distomedial peg seta rather strong, on the acute-angled distoventral segment edge; P-5 slender, and long, with four claw-like apical setae. Chelicera as described for P. conglobata.
Males — (Fig. 3). Idiosoma L/W 500-540/400-450. Dorsum, in addition to the paired elongate platelets posterior to the posterior antenniform setae with three further pairs of muscle attachment platelets – two pairs located posterolaterally to the elongate platelets and occasionally fused to each other, and one in posterior part of dorsum. Ventral sclerotization widely as described for P. conglobata; Cx-I+II mL 160-180, lat. L 120-125, integument, Cx-I apodemes directed posterolaterally; Cx-III+IV L/W 270/ 200 (ratio 1.4). Genital field L (incl. excretory pore)/W 120-130/300-350, broadly fused to posterior margin of Cx-III+IV, area of gonopore (L 30-35) and Ac as described for P. conglobata, total n Ac 28-39 (right/left 14-20/13-19), maximum Ac diameter 18-23; excretory pore may be isolated or (in aged specimens) fused to the posterior sclerotization of genital field, the flanking glandularia are free in the integument. Legs: I-/II-legs-6 thickened, club-shaped: at base strongly narrowed (basal H about 8), dorsal and ventral margins in distal part strongly convex; leg segment measurements (in parentheses, L/H ratio): I-leg-4, 125-130 (2.9-3.1); I-leg-5, 140-145 (3.4-3.6); I-leg-6, 100-110 (2.5-2.8) – I-leg-6 L max 145-155 (3.6-3.9); II-leg-4, 140-145 (3.5-3.7); II-leg-5, 150-160 (4.0-4.3); II-leg-6, 100-115 (2.4-2.6) – II-leg-6 L max 145-170 (3.4-3.8); III-leg-4, 150-155 (3.8-3.9); III-leg-5, 180-190 (4.8-5.3); III-leg-6, 100-110 (3.2-3.8) – III-leg-6 L max 113-120 (3.5-4.1); L ratio III-leg-5/6 (max) 1.5-1.7; IV-leg-4, 150-155 (3.8-3.9); IV-leg-5, 170-175 (4.7-5.0); IV-leg-6, 140-155 (6.4-7.8) – IV-leg-6 L max 155-170 (7.2-8.5); L ratio IV-leg-4/5, 0.86-0.91; IV-leg-4/6, 0.97-1.11; IV-leg-4/6 L max 0.88-1.00; IV-leg-5/6, 1.13-1.21; IV-leg-5/6 L max 1.03-1.10. Mouthparts: Gnathosoma L 120-130; palp total L 345-333, segments L (L/H ratio, relative L): P-1, 28-30 (0.56-0.63, 8%); P-2, 110-115 (1.66-1.80, 32%); P-3, 58-60 (1.18-1.23, 16-17%); P-4 (ventral extensions (included in H measurements) 100-105 (2.33-2.50, 29-30%); P-5, 46-48 (2.11-2.24, 13-14%); L ratio P-2/P-3, 1.83-2.00; P-2/P-4, 1.07-1.10; P-3/P-4, 0.55-0.60.
Females — (Fig. 4). Idiosoma L/W 550-950/480-700; dorsum with one pair of elongate platelets posterior to the posterior antenniform setae, without further muscle attachment platelets. Ventral sclerotization consisting of four distinct parts: paired Cx-I+II, mL 180-220, lat L 135-159; Cx-III+IV L 270-315, W 215-260 (L/W ratio 1.2-1.3), suture Cx-III/-IV reaching medial plate margin; genital field L (pregenital sclerite – excretory pore)/W 220-320/320-520, gonopore L 150-230, most Ac lying free in the membranous integument, scattered over paired crescent-shaped areas extending posterolaterally to gonopore, the anteriormost Ac on each side on a triangular platelet together with 4-5 fine setae, on the level of the postgenitale, basal sclerites of 2-3 Ac fused together to form a platelet bearing 4 fine setae; total Ac n 24-31 (right/left 11-15/13-16), maximum Ac diameter 23-24. Legs: I-/II-legs-6 rather slender, basal H about 14, dorsal and ventral margin nearly straight, equally diverging from base to tip; leg segment measurements (in parentheses, L/H ratio): I-leg-4, 145-165 (3.3-3.5); I-leg-5, 155-180 (4.2-4.4); I-leg-6, 115-125 (3.1) – I-leg-6 L max 165-185 (4.4-4.6); II-leg-4, 165-190 (4.0-4.1); II-leg-5, 180-200 (4.7-5.3); II-leg-6, 125 (3.1-3.6) – II-leg-6 L max 180 (4.5-5.1); III-leg-4, 175-190 (4.2-4.7); III-leg-5, 195-220 (5.2-5.6); III-leg-6, 135-165 (3.9-4.1) – III-leg-6 L max 185-205 (5.1-5.3); ratio III-leg-5 L/III-leg-6 L max 1.1; IV-leg-4, 195-215 (4.8-5.0); IV-leg-5, 200-230 (5.8-6.2); IV-leg-6, 165-180 (6.2-6.4) – IV-leg-6 L max 190-205 (7.1-7.2); L ratio IV-leg-4/5, 0.93-0.98; IV-leg-4/6, 1.11-1.21; IV-leg-4/6 L max 1.00-1.05; IV-leg-5/6, 1.14-1.28; IV-leg-5/6 L max 1.03-1.12. Mouth parts: Gnathosoma L 160-180; palp total L 380-415, segments L (L/H ratio, relative L): P-1, 35-40 (0.67-0.76, 9-11%); P-2, 120-138 (1.53-1.57, 32-33%); P-3, 63-65 (0.93-1.24, 16-17%); P-4, 105-120 (2.53-2.80, 28-29%); P-5, 48-55 (2.20-2.63, 13-14%); in one case of asymmetry, with an additional projection on P-4 of one of the two palps; L ratio P-2/P-3, 1.85-2.12; P-2/P-4, 1.12-1.19; P-3/P-4, 0.54-0.60.
Discussion
The name-giving coloration is probably due to the expanded whitish excretory organ, reducing the basic brown colour to isolated dots as it is found in aged specimens. Both sexes of Piona punctata differ from P. conglobata in having rather large Ac and only two ventral projections on P-4. This latter character state has also been observed in P. coacta (Koenike, 1895), a species incerta apparently described from an abnormal specimen (K. Viets 1956, Lundblad 1962). Concerning the synonymy of Piona stuxbergi, P. distermina and P. conjugula, we follow Lundblad (1962) – he had the occasion to compare the type material of the latter two and P. punctata. The female of P. distermina, described from Sweden and conservated in Stockholm, agrees with the description of P. punctata from all points of view.
Piona lacustricola K.Viets, 1949 nov stat.
Piona conglobata lacustricola K.Viets, 1949 – nom. nov. pro Piona conglobata lacustris Halbert, 1944 [praeocc.: Koch, 1836])
Material examined – Norway
Trøndelag, Frosta, pond Beitemarksdammen, N63.58126°, E10.68968°, 105 m a.s.l., 8 Aug. 2022, leg. GK, 1♂ on slide, NTNU-VM 291064, BOLD Specimen id HYDCA826. Nordland, Brønnøy, pond Hallarauntjønna, N65.40398°, E12.46316°, 23 m a.s.l., 16 July 2023, leg. GK & KH, 4♀♀ in tube, NTNU-VM 291081. Nordland, Vega, lake Olhåksåvatnet, N65.69632°, E11,8888°, 6 m a.s.l., 17 July 2023, leg. GK & KH, 1♀ on slide, NTNU-VM 291065, BOLD Specimen id HYDCA979, 1♀ in tube, NTNU-VM 291066.
The specimens of Piona lacustricola for which the DNA barcode has been obtained are placed in BIN BOLD:AEC4987. The p-distance between the latter BIN and its nearest neighbour, BOLD:AFA2527, which includes specimens of Piona punctata from Norway, was estimated at 9.33% indicating a long genetic separation between these two species.


Diagnosis
Both sexes — Ac diameter 13-16; P-4, in addition to the two-pointed ventral seta-bearing extensions, without further projections.
Males — III-leg-6 strongly shortened (L/H III-leg-5/6, 1.6); setal pores and glandularia flanking excretory pore fused to genital field.
Females — Genital field with 2-3 anteriormost Ac on a common setae-bearing platelet, 3-7 posteromedial Ac fused to a setae-bearing platelet, most other Ac not isolated, but fused with their basal sclerites with 1-2 neighbours.
Description
Both sexes – Colour unknown (not reported in the original description, not documented in the investigated material). Integument lineated. Genital field with 25-26 Ac (on one side 11-14). I-/II-legs-6 thickened, with sexual dimorphism in shape; III-leg-4 and IV-leg-5-6 and chelicera as described for P. conglobata.
Males — (Fig. 5). Idiosoma L/W 480/390. Dorsum with the paired elongate platelets posterior to the posterior antenniform fused with neighbouring muscle attachment sclerites to form laterally directed ''L-shaped'' platelets, and one pair of round muscle attachment sclerites in posterior part of dorsum. Ventral sclerotization as described for P. conglobata; Cx-I+II mL 170, lat L 110, Cx-III+IV L/W 260/185 (ratio 1.4). Genital field L (incl. excretory pore)/W 130/280, broadly fused to posterior margin of Cx-III+IV, area of gonopore (L 33) and Ac as described for P. conglobata, total n Ac 25 (right/left 14/11), maximum Ac diameter 13; excretory pore and flanking setal pores and glandularia fused to genital field. Legs: I-/II-legs-6 thickened, club-shaped: at base strongly narrowed (basal H about 10), dorsal and ventral margins in distal part strongly convex; leg segment measurements (in parentheses, L/H ratio): I-leg-4, 120 (3.4); I-leg-5, 130 (3.7); I-leg-6, 95 (2.7) – I-leg-6 L max 145 (4.1); II-leg-4, 130 (3.7); II-leg-5, 143 (4.1); II-leg-6, 105 (3.0) – II-leg-6 L max 150 (4.3); III-leg-4, 145 (4.5); III-leg-5, 170 (5.7); III-leg-6, 95 (3.2) – L max 100 (3.3) ; L ratio III-leg-5/6 (max) 1.7; IV-leg-4, 145 (4.1); IV-leg-5, 150 (4.8); IV-leg-6, 130 (5.8) – IV-leg-6 L max 145 (6.4); L ratio IV-leg-4/5, 0.97; IV-leg-4/6, 1.12; IV-leg-4/6 L max 1.00; IV-leg-5/6, 1.15; IV-leg-5/6 L max 1.03. Mouthparts: Gnathosoma L 140; palp total L 335, segments L (L/H ratio, relative L): P-1, 28 (0.61, 8%); P-2, 108 (1.69, 32%); P-3, 55 (1.16, 16%); P-4 (ventral extensions included in H measurement) 100 (2.50, 30%); P-5, 45 (2.12, 13%); L ratio P-2/P-3, 1.95; P-2/P-4, 1.08; P-3/P-4, 0.55.
Females — (Fig. 6). Idiosoma L/W 900/770; dorsum and ventral sclerotization as in P. conglobata; Cx-I+II, mL 250, lat L 150; Cx-III+IV L 310, W 230 (L/W ratio 1.4); genital field L (pregenital sclerite-excretory pore)/W 375/410, gonopore L 225, Ac tending to fuse with their neighbours in groups of 2-7 units, the anteriormost Ac on each side on a triangular platelet including 1-2 neighbouring Ac and 3-4 fine setae, on the level of the postgenitale, basal sclerites of 3-7 Ac fused together to form a platelet bearing 2 fine setae; total Ac n 26 (right/left 13/13), max Ac diameter 6. Legs: I-/II-legs-6 rather slender, basal H about 15, dorsal and ventral margin nearly straight, equally diverging from base to tip; leg segment measurements (in parentheses, L/H ratio): I-leg-4, 155 (3.9); I-leg-5, 170 (4.9); I-leg-6, 170 (4.9) – I-leg-6 L max 180 (5.1); II-leg-4, 190 (4.8); II-leg-5, 210 (6.0); II-leg-6 lost; III-leg-4, 205 (5.1); III-leg-5, 215 (5.2); III-leg-6, 160 (6.4) – III-leg-6 L max 185 (7.4); L ratio III-leg-5/6 (max) 1.2; IV-leg-4, 183 (4.6); IV-leg-5, 195 (5.2); IV-leg-6, 180 (5.1) – IV-leg-6 L max 190 (5.4); L ratio IV-leg-4/5, 0.94; IV-leg-4/6, 1.0; IV-leg-4/6 L max 0.96; IV-leg-5/6, 1.08; IV-leg-5/6 L max 1.03. Mouth parts: Gnathosoma L 160.
Discussion
The original description of this species, initially presented as a subspecies of P. conglobata and thus far known only from females, is extremely brief. It comprises just a few lines noting the sclerotization of the genital field and the structure of P-4. Although an illustration of the genital field is provided, no measurements are included. It is one of the taxa which Lundblad (1962) proposed to lump under the stem species. Females from our collections that are here attributed to P. lacustricola correspond closely with Halbert's (1944) Fig. 47. They differ from the other species of the P. conglobata aggr. in that most Ac are fused with basal sclerites of 1-2 neighbours. In males, which are here described for the first time, the setal pores and glandularia flanking the excretory pore are fused to the genital field. Both sexes agree with P. conglobata in having small Ac, but differ from that species in the more slender P-4 lacking additional extensions and the stouter IV-leg segments; males additionally differ in the club-shaped I-/II-leg-6 and the markedly shortened III-leg-6 (L ratio III-leg-5/-6 in P. conglobata < 1.5); in these character states they resemble P. punctata.
Towards the identification of the species of the Piona conglobata aggregate
Males
1. P-4 with additional ventral projections; III-leg-6 relatively long (L ratio III-leg-5/6 < 1.5); I-/II-legs not particularly thickened, ventrally straight; Ac small (diameter < 17)
...... P. conglobata
— P-4 without additional ventral projections; III-leg-6 relatively short (L ratio III-leg-5/6 > 1.5); I-/II-legs-6 more or less inflated, with convex ventral margin; Ac various in size
...... 2
2 (1). Ac small (diameter < 16); I-/II-legs 6 slightly thickened; dorsal muscle attachments widely fused; genital field broadly fused to Cx-IV, excretory pore, as well as flanking glandularia and setal pores, fused to genital field
...... P. lacustricola
— Ac larger (diameter > 17); I-/II-legs strongly inflated, club-shaped; dorsal muscle attachments separate; fusion of genital field to Cx-IV less extended, at least glandularia flanking excretory pore separate from genital field
...... P. punctata
Females
1. P-4, in addition to the distomedial peg seta, with more than two setae-bearing projections. Ac small (diameter < 17)
...... P. conglobata
— P-4, in addition to the distomedial peg seta, with not more than two setae-bearing ventral projections. Ac may be larger
...... 2
2 (1). Idiosoma sclerotization strong (basal sclerites of neighbouring Ac fused in many cases); Ac small as in P. conglobata
...... P. lacustricola
— Idiosoma sclerotization less developed (most Ac not fused to basal sclerites of neighbours); Ac larger (diameter > 17)
...... P. punctata
Acknowledgements
The fresh material was collected within the Artsprosjekts 17-22 ''Calcareous mites and midges'' and 13-18 ''Water Mites and Midges in southern Norway'', both funded by the Norwegian Biodiversity Information Center. A great thank you to Gaute Kjærstad, Karstein Hårsaker and Torbjørn Ekrem for collecting water mites during field work within these projects. For information about the types of Piona distermina and providing high-quality photographs, we are thankful to Victor Paranha and Gunvi Lindberg (Museum of Natural History, Stockholm). We are grateful to Mirela Cîmpean and Vladimir Pešić for their critically reading of a former draft of this paper, and to Joanna Mąkol for her patient editing.
References
- Gerecke R., Gledhill T., Pešić V., Smit H. 2016. Chelicerata: Acari III. In: Gerecke R, ed. Süßwasserfauna von Mitteleuropa, Bd. 7/2-3. Springer-Verlag Berlin, Heidelberg, pp. 1-429. https://doi.org/10.1007/978-3-8274-2689-5
- Gerecke R., Kjærstad G., Ekrem T., Stur E. 2022. A faunistic study of water mites (Hydrachnidia and Halacaridae) from southern Norway. Norwegian Journal of Entomology, 69: 243-268.
- Gerecke R., Kjærstad G., Ekrem T., Stur E. 2025. Exploring water mites (Hydrachnidia) in calcareous freshwater habitats of Norway. Norwegian Journal of Entomology, 72: 184-223. https://doi.org/10.61698/nje.v72n2.7
- Halbert J.N. 1944. List of Irish fresh-water mites (Hydracarina). Proceedings of the Royal Irish Academy, 50, B (4): 39-104.
- Koch C.L. 1837. Deutschlands Crustaceen, Myriapoden und Arachniden 11: 20-21.
- Koenike F. 1907. Zwei unbekannte Sperchoniden und eine Curvipes-Spezies. Abhandlungen herausgegeben vom naturwissenchaftlichen Verein zu Bremen, 19 (1): 133-138. https://doi.org/10.5962/bhl.part.16693
- Lundblad O. 1962. Die Hydracarinen Schwedens. II. Arkiv för Zoologi (2), 14 (1): 1-635.
- Marshall R. 1935. Preliminary list of the Hydracarina of Wisconsin. Part. IV. - Transactions of the Wisconsin Academy of Science, 29: 273-297.
- Musselius A.A. 1913. On the knowledge of the Hydrachnid fauna of Russia [in Russ.]. Arbeiten aus der hydrobiologischen Station Glubokoje, 5 (1): 136-157.
- Neuman C.J. 1875. Gottlands och Ölands spindlar oich vattenkvalster. Öfvers. Vetenskaps-Akademien Förhandlingar, Stockholm, 2: 91-104.
- Neuman C.J. 1880. Om Sveriges Hydrachnider - Svenska Vetenskaps-Akademiens Handlingar, Stockholm (N.F.), 17 (3): 1-123. https://doi.org/10.5962/bhl.title.9705
- Piersig R. 1896-1899. Deutschlands Hydrachniden. Zoologica, 1896-1899, Stuttgart (Nägele), Bd. 19, H. 22, pp. 7+1-601.
- Punčochař P., Hrbáček J., 1991. Water mites in the plankton of Hubenov reservoir and their relations to fish stock composition. In: Modern Acarology, Vol 1: 449-457. Dusbábek F. & Bukva V. (eds.), SPB acad. Publ. bv, The Hague.
- Schwoerbel J. 1972. VII. Hydrachnellae, Wassermilben, Chelicerata, Acari. in: Das Zooplankton der Binnengewässer. Binnengewässer, 26 (1): 281-285.
- Soar C.D., Williamson W. 1929. The British Hydracarina. Vol.III. London, Ray Society, 115: 1-232.
- Stur E., Gerecke R., Kjærstad G., Ekrem T. 2025. Dataset - DS-CMMHY Exploring water mites (Hydrachnidia) in Calcareous Freshwater Habitats of Norway. https://doi.org/10.61698/nje.v72n2.7
- Stålstedt J., Bergsten J., Ronquist F. 2013. ''Forms» of water mites (Acari: Hydrachnidia): intraspecific variation or valid species? Ecology and Evolution, 3: 3415-3435. https://doi.org/10.1002/ece3.704
- Viets K. 1931. Tiefenverteilung einiger Hydracarinen in norddeutschen Seen. Verhandlungen der Internationalen Vereinigung für Limnologie, 5: 276-281. https://doi.org/10.1080/03680770.1931.11898461
- Viets K. 1936. Wassermilben oder Hydracarina (Hydrachnellae und Halacaridae). - in: Dahl, F.: Tierwelt Deutschlands, Jena (G. Fischer), 31: 10+1-288., 32: 289-574.
- Viets K. 1949. Nomenklatorische und taxonomische Bemerkungen zur Kenntnis der Wassermilben (Hydrachnellae, Acari). I-X. Abhandlungen herausgegeben vom naturwissenschaftlichen Verein zu Bremen, 32 (2): 292-327.
- Viets K. 1956. Die Milben des Süßwassers und des Meeres. Hydrachnellae et Halacaridae (Acari). II. und III. Teil: Katalog und Nomenklator. 870 pp. G. Fischer, Jena.



2026-03-13
Date accepted:
2026-04-24
Date published:
2026-05-18
Edited by:
Mąkol, Joanna

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Gerecke, Reinhard and Stur, Elisabeth
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)