



Variations, fuzzy boundaries and normality in Tydeidae (Acariformes: Tydeoidea)
André, Henri M.  1
1
1✉ Retired, currently, 22, route de la Côte, CH-1615 Bossonnens, Switzerland & Musée royal de l’Afrique centrale, B-3080 Tervuren, Belgium & Université Catholique de Louvain, B-1348 Louvain-la-Neuve, Belgium.
2026 - Volume: 66 Issue: 2 pages: 414-430
https://doi.org/10.24349/46un-n697ZooBank LSID: 243AD056-B2D1-4CB0-BD8A-985EF16881DF
Original research
Keywords
Abstract
It at once struck me that under these circumstances favourable variations
would tend to be preserved, and unfavourable ones to be destroyed.
Darwin (1958: 20), Autobiography.
Introduction
I have three objectives in the present work. First of all, I amend and supplement my article ''Sixty years after ′A review of the genera of the family Tydeidae′ (Acariformes: Tydeoidea: Tydeidae)'' (André, 2025).
Second, I analyze tydeid genera and their variants. Although morphometric variations support the distinction of cryptic species (e.g. Bukva (1990) on Demodicidae, Navia et al. (2015) on Phytoptidae, Marini et al. (2021) on Eriophyidae...), the importance of meristic variations in evolutionary processes is emphasized even if they are rarely reported in Tydeidae. A few additional examples concerning other prostigmatid mites will be given.
My third objective arises from these observed variations and concerns fuzzy boundaries and a probabilistic definition of taxa even if they are designated within a language system of discrete and abstract elements (Carignan et al., 2022). The systematics of Tydeidae illustrates the perennial debate between discreteness and continuity (Langacker, 2006).
Material and Methods
This article treats extant morphospecies (i.e. species based on morphological criteria) described up to 31 December 2024. Fossil specimens would not exist as concluded by Klimov et al. (2025: 10). As in André (2025), data are based mainly on personal data and bibliographical references (books, articles, review papers) devoted to the taxonomy of Tydeoidea and on the database in the Wikispecies platform database (André, 2021). Character-state polarity follows the cladistic analyses found in Kaźmierski (1998a) and André and Fain (2000).
From idionymous elements to meristic traits
Idionymy was formally defined, in French, by Grandjean (1949: 801) and reiterated in English by Hammen (1976; 1981: 6), André (1979: 207), and in the Online Dictionary of Maggenti et al. (2008: 183). Another definition of idionymy is given in the glossary of acarine terms published by Walter (2006) and is not followed here. Idionymy and the resulting notation system is not applicable to Tydeidae except in a few cases such as ft′, k″, ip... (André, 1981a, b). In other words, ft′ in Tydeus spathulatus is homologous to ft′ in Brachytydeus (Eotydeus) mirabilis. In contrast, genital setae are listed from anterior to posterior and ge1 is simply the first of the alignment; ge1 in Tydeus spathulatus is not necessarily homologous to ge1 in B. (E.) mirabilis.
Therefore, a designation system based on the location is used (for example, the number of setae on a podomere) and only meristic traits are considered, i.e. counts of discrete serially homologous structures (Lawing et al. 2008). In this article, counts concerns only setae, solenidia and lyrifissures. The numbers of genital acetabula, genital discs, podomeres and eye-spots, are excluded from analysis. However, the two last elements are discussed even though they do not appear in the figures and calculations.
Meristic traits and morphospace
The total meristic value, MVtot, is therefore the summation of all (n) meristic characters considered:
\[\mathrm{MV}_{\mathrm{tot} }=\sum_{i=1}^n m c_i\]
where mci = 1 if the character i is present and mci = 0 if it is absent. As explained hereafter, setae ep, eI, sc1, sc2, a1, a2 are not taken into account.
The total value can be broken down into 3 partial values: MVsol, i.e. the meristic value restricted to appendage solenidia (2, 3 or 4); MVapp, i.e. the meristic value restricted to appendage chaetotaxies and MVbod, i.e. the meristic value restricted to body chaetotaxies (from mouth to anus, prodorsum, opisthosoma, epimeral area, genital region...).
With these three partial values it is therefore possible to construct a three-dimensional character-based morphospace (Budd, 2021), even a three-dimensional meristic (morpho)space. The meristic unit is then the minimum spatial distance (González-González, 1998) in a three-dimensional meristic space such as those shown in figs 3. The possible combinations of states represent the vertices of a cube, and each genus sits at one of these vertices. The differences between each genus can then be estimated by the so-called ′Hamming distance′, which is simply the number of meristic differences in a particular pairwise comparison (Budd, 2021: 1183), or by the Euclidian distance as in figure 3b.
Fig. 4 is similar to fig. 3, but the Z coordinates, MVsol in fig. 3, are replaced by the number of species in the genus.
Terminology of variations
In addition to anomalies, Grandjean (1949: 802) distinguished four categories of individual variations: mutations, ''écarts'' later renamed vertitions (Grandjean, 1952: 2), numerical differences with collective significance and continuous fluctuations.
Anomalies are individual and exceptional variations of any kind, almost always unilateral, to which, based on our current knowledge, no precise evolutionary significance can be attributed (Grandjean, 1972: 455). In this work, they are part of the fluctuating asymmetry (FA) discussed by Van Valen (1962) but concern only idionymous organs (setae, solenidia, lyrifissures) and their presence/absence whereas FA is frequently assessed for continuously varying features (e.g., length of a structure) (Palmer and Strobeck, 1986).
Vertitions are also asymmetrical (Grandjean, 1952: 2) and constitute the current manifestation of evolution (Grandjean, 1952: 3). They are variations in the presence or absence of an idionymous organ observed in individuals of the same species at the same stase, when these variations are essentially unilateral and our general knowledge allows us to attribute evolutionary significance to them (Grandjean, 1972: 455). The evolutionary significance of vertitions is also outlined by Maggenti et al. (2008: 372) in their Dictionary of Invertebrate Zoology.
This leaves us with the task of interpreting asymmetrical drawings such as that of Lorryia superba published by Oudemans (1928: 230). Do the asymmetries reflect genuine anomalies or do they indicate an imperfect reproduction of the observed traits?
Sometimes, the disappearance of a setae is accompanied by the movement of the remaining element of the pair on the sagittal plane: it is the bisynthesis defined by André (1981a: 32). This variation seems to be close to the fusion of genital discs reported by André and Fain (2000: fig. 3A).
Results
The tydeid genera
Instead of the usual 6 pairs of genital setae (ge), there are at least two species of Tydeus with only 5 pairs. Firstly, there is Tydeus (Pertydeus) schusteri described by André and Naudo (1965). Next, there is Tydeus tuttlei described by Baker (1965: 100-101) in his seminal paper. These species were grouped together in the same subgenus, Pertydeus (Baker, 1970: 164) although, according to the descriptions, they have different leg chaetotaxies. They should therefore be assigned to different genera, which had not been done previously.
Another problem arises due to the ''oversight'' of Brachytydeus. The genus was synonymized with Tydeus without any explanation by Baker and Wharton (1952: 191) and only restored in 2005 (André, 2005: 995) (Fig. 1). The species described in the meantime were therefore assigned to Tydeus even though they had a different leg chaetotaxy. This was precisely the case with the mite described by André and Naudo (1965); which actually belongs to the genus Brachytydeus and not to Tydeus. Again, correct assignments were not made in André (2025).
Consequently, table 1 presented by André (2025) is corrected by Fig. 1 in this work (Acanthotydides instead of Acanthodides, tibia I of Novzelorryia and Ueckermannia) and completed. Moreover, table 1 presented by André (2025) did not consider the variations, particularly anomalies and vertitions, which are detailed below.
The nature and variations of eyespots
The term ''eye'' poses three problems. It can refer to a lens-like structure, an eyespot formed by silvery granules that is easily visible on living mites (André and N′Dri, 2013: fig. 11A), or a structure that is mistakenly considered to function for sensing light. Secondly, on slide-mounted mites, pigments are frequently decolored. Finally, the absence of pigments does not imply the absence of a physical structure underlying the integument or lens: some media for acrylics are milky white when wet but colorless and transparent when dry.
There is no lens-like structure in Tydeidae but such a structure is visible in several tydeoid Ereynetidae, for instance the genera Astrida, Speleognathus.
In Tydeidae, a first clear interpretation was offered by Berlese: he stated that the sensory organs comprised the eyes: very rarely none, usually two (Berlese, 1882: 13). He added that the two eyes (one on each side) were conspicuous on (what he called) the cephalothorax, always shimmering silvery, extremely bright and, unique to this family, endowed with iridescent pigment [''Oculi duo (utrinque unus), ad latera cephalithoracis conspicui, semper argenteo micantes, lucidissimi et, quod in hac tantum est familia, pigmento iridescenti praediti.''] (Berlese,1882: 20). This view was supported by André and Naudo (1965: 674) who wrote that the propodosoma may or may not have a pair of pigment spots (eyes) [''Le propodosoma peut porter une paire de taches pigmentaires (yeux) ou non.''] and Kaźmierski (1998b: 286) who asserted that ''a pair of primitive eyes (oculi) (oc) commonly occur laterally''. Baker (1965: 97) adopts a similar point of view. The spot was considered a derived character, resulting from the disappearance of the lens (Fain, 1956: 644; 1963: 51), but, after cladistic analysis, the median pigment spot was considered a plesiomorphic character by André and Fain (2000).
Another interpretation was given by Sig Thor who described Tydeidae as colorless or grayish-white animals with darker spots, which originate mainly from stomach contents or excretory organs [''...farblose oder grauweiße Tiere mit dunklere Flecken, die namentlich von dem Mageninhalt oder von Exkretionsorgan herrühren''] (Thor, 1933: 4). The caption of his figure 1, right next to the quoted text, was ''Tydeus subalpinus [...] Dorsal outline of the body, with eyes, sensory hairs...'' [''Dorsaler Umriß des Körper, mit Augen, Sinneshaaren...{ }'']. In German as in English and French, the term ''Auge[n]'' (eye[s], oeil [yeux]) may refer to something whose shape resembles an eye (such as the bud on a potato, Duden online) and not necessarily to sensory organs of animals. There is no visible lens and the eye is composed of many small pigment corpuscles (Thor, 1933: 33). A similar interpretation — an internal origin, precisely glandular globules — was proposed by A. Fain in André and Fain (2000: 416, photos of Fig. 1) and concerns not only Ereynetidae but all the Tydeoidea. The proximity of the posterior end of the podocephalic canal (cpc) was a key point. The connection between a gland and the cpc had already been described in Brachytydeus (B.) viviparus by Grandjean (1938, dg in Figure 1).

Secondly, on slides, pigments are frequently decolored (Thor, 1933: 6; Baker, 1965: 97; Wood, 1965: 672; Baker, 1968a: 987; André, 1981a: 32), but not always (Kaźmierski, 1998b: 301). Decoloration or discoloration are not all-or-non characters, they can be partial, the spot becomes indistinct or is not clearly demarcated This has an impact on the representation through drawings or photos. The photograph of Pseudotydeus lebruni (André and Ducarme, 2003: Fig. 1) suggests the presence of two eyespots/coxal glands as discussed in the text (André and Ducarme, 2003: 377). The glands of Hanriccardoella faini are barely discernible (André et al. 2004, Fig. 3A) and would probably not have been sketched if a line drawing had been chosen as the representation.
Lastly, the famous aphorism ''absence of evidence is not evidence of absence'' applies to eyespots. Baker (1965: 97) concluded that eyes were not used for either generic or specific identification but also noted (page 110) that three eyespots were visible in mounted specimens of Triophtydeus. Kaźmierski (1998b: 301-302) reached a similar conclusion: ''However, it is perfectly certain that this feature cannot be accepted as a decisive one in the construction of a key.'' However, the genus Triophtydeus (currently in Triophtydeidae, formerly in Tydeidae) was defined by the presence of three eyes, and the genus Opseyrenetes (Ereynetidae) was based on the presence of two eyespots (Thor, 1932). Several tydeid genera lack them: Acanthotydides, Afridiolorryia, Edlorryia, Kenlorryia, Momenia, Quasitydeus, Ueckermannia. The problem becomes crucial when considering the genus Australotydaeus and the monospecific subfamily Australotydaeinae. Eyespots were not described in the original description (Spain, 1969), or in subsequent articles (André, 1980: 111; 1981a: 32) and it is still unclear whether or not Australotydaeus has eyespots.
′Eyespots' are not primitive sensory organs and are probably part of the podocephalic gland complex (Krantz, 2009: 25) as suspected by Thor (1933). They are internal structure covered by striae of the integument while eyes photographed by Khaustov (2015: Fig. 13C) locally modify the striation pattern. Consequently, this trait is not listed in the table of genera presented in Figure 1 and is not used in the morphospace of Figures 3 and 4. A unique variant is documented by André (1985: 192; André and Fain, 2000: 412) who reported a Sicilian population of Tydeus with 3 eyespots. The species was not precised.
Variations of poroidotaxy
There are four pairs of lyrifissures in Australotydaeus: (ia), (im), (ip) and (ih) (André, 1980: 111; 1981: fig. 2A). The lyrifissure ip seems to be missing in most other Tydeidae. The poroidotaxy is unknown in several genera: Afridiolorryia, Edlorryia, Novzelorryia (''Lyrifissures very poorly visible, hidden between the elements of ornamentation'', Kaźmierski, 1996: 192), Quasitydeus, Ueckermannia... Only 2 lyrifissures, ia and im, are reported in Acanthotydides, Kenlorryia and Momenia. Consequently, poroidotaxy is not listed in the table of genera presented in Figure 1 and is not used in the morphospaces of Figures 3 and 4. No variation is reported.
Variations of palp phanerotaxy
In Tydeidae, the number of palpomeres does not vary as it does in Ereynetidae, the palp is ''four segmented and typically shaped'' as noted by Baker (1965: 96).
A key character of Lorryia superba is the weak armor of the tibia-tarsus of the palp [''de povere bewapening van den tibia-tarsus van den palp.''] (final remark by Oudemans (1928: 233)). The usual chaetotaxy, (6-2-2), or more precisely (6-2-2-0), seems to be plesiomorphic state in Tydeidae; it becomes (6-1-2) in Afridiolorryia, Andrelorryia, Apolorryia, Idiolorryia, Krantzlorryia, Metalorryia and Neoapolorryia and (6-2-1) in Acanthotydides and Kuznetsovtydides.
An opposite trend, a rich chaetotaxy of ultimate segment, is described by Kaźmierski (1998: 304). The unique specimen of B. globulipalpus has 7 setae on the right tarsus and 8 setae on the left ultimate segment. Such an unusual chaetotaxy is reminiscent of that of Edbackerella (Triophtydeinae). However, the palp of B. globulipalpus drawn by Kaźmierski and Sikora (2008: 85, fig. 2) shows double setae, l'' and d, and a simple terminal eupathidium which has nothing to do with the three terminal eupathidia observed on the triophtydeid species (André, 1980: Fig. 9J).
The palp solenidion, ωp, is always present on the tarsus; no variations are reported.
Variations of apotele chaetotaxy
The apotele is the terminal tridactyl podomere of legs with two lateral claws, ol′ and ol″, and one median empodium, om. Together with tarsus I, apotele I is the seat of the palpian evolution in Tydeoidea (André, 1981: 166; André, 2025: fig. 5). The so-called putative step was described in Idiolina augustae (André, 1980: fig. 2C; André and Fain, 2000: fig. 4D).
In Lorryia superba, a claw, ol′ or ol″, is missing on all legs, only a scar is visible. However, there is an intermediate state: a lateral claw is also absent on the 4 front legs of a specimen of Brachytydeus (B.) reticulatus (Kaźmierski and Sikora, 2008: 88). Apotele IV is missing on a side of Brachytydeus (B.) exiguelitteratus (Kaźmierski and Sikora, 2008: 85, fig. 4). No other variations of apotele are reported.
Variations of leg chaetotaxy
Numerous anomalies were observed on other podomeres by Momen and Lundqvist (1993) and Kaźmierski and Sikora (2008: 85). They are shown at the bottom of figure 1 (upwards arrows) and are reminiscent of the anomalies studied in Tetranychidae by Leponce et al. (2001). A special case is offered by seta k'' in Tydeoidea: ''We could not see seta k, but this seta is always difficult to observe'' (Momen and Lundqvist, 1993:139, caption table 2). It is sometimes not mentioned in the original descriptions, even though it was subsequently observed on the holotype (André, 1980: 106 [A. setsukae], 106 [N. kenyensis], 107 [A. congoensis]... ; Kaźmierski, 1996: 190 [U. grewiae], 216 [M. longichela]). This small seta is important as it is a key component of ereynetal organ of the Ereynetidae.
Near the podocephalic canal are the two supracoxal setae, two short spines often overlooked in descriptions (e.g. Nozvelorryia, Pretydeus, Ueckermannia). They seem to be constant (André, 1980: 104); they are not counted in MVapp and do not appear in figure 1.
Similarly, asymmetry of the leg chaetotaxy has been observed in Ereynetidae (Booth, 1983: 10), Camerobiidae (Paredes-León et al. 2019: 11), Halacaridae (Abé, 2024: table 1), Penthalodidae (Brunetti et al. 2021: 123), Tuckerellidae (Sidorchuk and Khaustov, 2018: 111) and Tetranychidae (Wauthy et al., 1998; Leponce et al., 2001) as well as variations in Iolinidae (De Vis et al., 2025: 351, 362).
Variations of leg solenidiotaxy
On legs, solenidion φI is observed on tibia I in most Pretydeinae (in Novzelorryia, Pretydeus, Ueckermannia but not in Prelorryia). ωII is missing in Andrelorryia, Idiolorryia and Orfareptydeus. The total number of solenidia in the legs varies from 2 to 4.
Individual variations are rarely reported. Kaźmierski and Sikora (2008: 85) observed a specimen of Brachytydeus tragelaphus with a tarsus I quite nude and Momen and Lundqvist (1993: 139) observed a specimen of Brachytydeus with no ωII.
In Tetranychus urticae, numerous variations (frequency of uni- or bilateral absence of solenidia up to 10%) were observed on leg podomeres by Wauthy et al. (1998: table 2). In Brevipalpus (Tenuipalpidae), asymmetry in the number of solenidia on tarsi II was reported by Kitajima et al. (2011).
Variations of dorsal setae (prodorsum and opisthosoma)
The prodorsum usually has four pairs of setae in Tydeidae. However, the anomaly of p1 was observed in Neohomeotydeus formosus (André, 1984: 121, fig. 2B), and the anomaly of v2 was drawn by Khaustov et al. (2024: fig. 2). The reduction to 6 prodorsal setae was reported in the tydeoid families Iolinidae (André,1981a: 32), and Ereynetidae (André and Fain, 2000: 415). Sometimes, a seta is disappearing as in N. formosus; sometimes, the disappearance of a setae is accompanied by the movement of the remaining element of the pair on the sagittal plane (bisynthesis in the iolinid Proctotydaeus schistocercae observed by André (1981a: 32) and in the ereynetid Metaboydaia poffei observed by Skoracki et al. (2018, fig. 4A).
Counting dorsal opisthosomal setae is sometimes difficult, as demonstrated by Knop and Hoy (1983) which leads to confusion between the iolinid genera Pronematus and Homeopronematus: Baker (1943, 1968b) did not report a terminal opisthosomal setae (D5 or h1 depending on the nomenclature). Tydeidae generally have 9 or 10 dorsal opisthosomal setae. Australotydaeus is an exception, with 11 pairs of opisthosomal setae. Anomalies concern setae d1 or d2, which may be missing in tydeoid Ereynetidae (André and Ducarme, 2003: 376).
In some Caeculidae, dorsal chaetotaxy is highly variable, even within the same species as shown by Coineau (1974: fig. 54) and Per et al. (2017: table 2). Asymmetry of prodorsal setae is also reported in Syringophilidae by Głowska and Skoracki (2009: table VII).
Variations of subcapitular setae
''There are always two subcapitular setae, sc1 and sc2'' (André, 1981: 42). ''Usually, two pairs of adoral setae (ad1) and (ad2), emerge near the apex of the subcapitulum. These are difficult to see because they are small, recumbent and probably eupathidial'' (André, 1981a: 42). These four setae are visible in the scanning electron microscope (SEM) micrographs published by Lorençon et al. (2016).
Subcapitular setae appear to be a constant feature in Tydeidae, even if they are neglected in many descriptions (e.g. Nozvelorryia, Pretydeus, Ueckermannia). Consequently, they are not counted in MVapp and do not appear in figure 1. However, in the closely allied family, Ereynetidae, a subcapitular seta is missing in Lawrencarinae and in some Speleognathinae (Fain, 1963: 14), for example in Boydaia (B.) aratingae (Fain, 1963: fig. 38).
Variations of epimeral setae
The new table in figure 1 displays epimeral formulae (related to epimeral or coxisternal setae listed from anterior to posterior). The most frequent is (3-1-4-2). In Krantzlorryia, most Pretydeinae and Australotydaeinae, the formula is (3-1-4-3), and it is (3-1-3-2) in Orfareptydeus. It does not allow differentiation between the tydeid subfamilies. However, it differs from that observed in Triophtydeidae, where the formula (3-1-3-3) is found in all genera.
In Caeculidae, the epimeral chaetotaxy is sometimes variable (Coineau, 1974: 149).
Variation of eugenital and aggenital setae
In the Tydeidae, the usual number of aggenital setae (ag) is four pairs. In a few cases, there are only three: Brachytydeus (Eotydeus) and Apolorryia. Asymmetry (3-4) is reported by Kaźmierski (1989: 296) in Brachytydeus (Q.) inconstans and B. (E.) mirabilis. Dastych (1990: 46) reported an additional aggenital seta in Pretriophtydeus tilbrooki (Triophtydeidae). Akyol (2011: 61) and Doğan et al. (2019: 118; 2022: 302) observed variations in the number of aggenital setae in Stigmaeidae. Fain and Nadchatram (1962: 77) described similar variations in Ereynetidae. Asymmetry (3-4) is reported by Khaustov (2022: 30) in Mediolata neocalifornica (Stigmaeidae). Coineau (1974: 149) observed similar variations in Caeculidae. Asymmetry in the number of aggenital setae in Syringophilidae is amply documented by Głowska and Skoracki (2009: figure 3, table I).
Eugenital setae (eu) are usually eupathidial and sexually dimorphic (present in males, absent in females) and many question marks are used in figure 1. They are sometimes difficult to see (André and Naudo, 1965: 676). The type-series of T. spathulatus contains 12 females (without eugenital setae) but no male. The female of Calotydeus croceus has no eugenital seta but the male has four pairs as indicated in figure 1. No variation is reported in Tydeidae: the absence of eugenital setae in females and the presence of four pairs in males seem to be the rule up to now.
Variations of genital setae
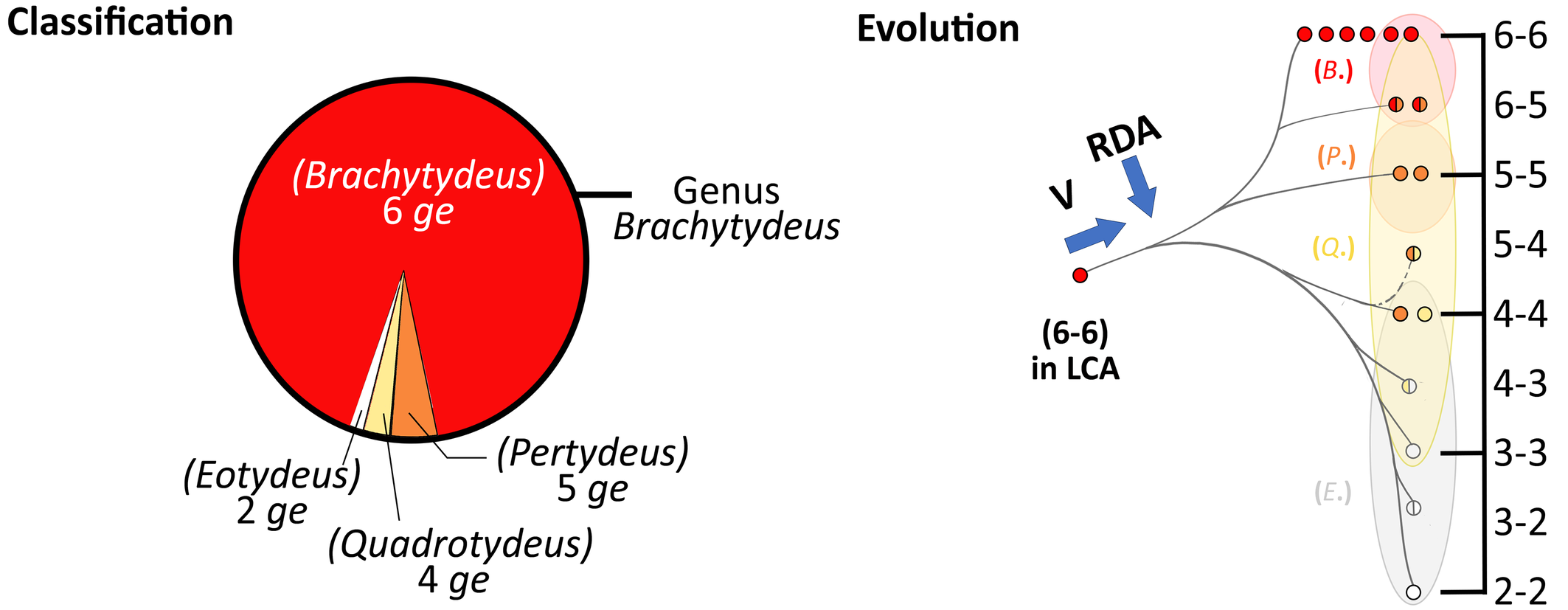
The usual and plesiomorph number of genital setae (ge) in Tydeidae is six pairs. A reduction in the number of genital setae is observed in several genera: Tydeus, Brachytydeus, Momenia and Neolorryia. However, the reduction is gradual in highly speciose genera, Tydeus and Brachytydeus, as illustrated in figure 1. This probably corresponds to the shrinkage of the cis-acetabulal area observed in Tydeidae by André and Fain (2000).
Firstly, observations have been disparate and have given rise to apparently inconsistent taxa. For example, the genus Homoiorthotydeus erected by Schiess (1981: 84) was synonymized with Tydeus by Kaźmierski (1989: 289; 1998a: 342). Kaźmierski (1989: 289) stated: ''It is inconsistent to assign to different genera any species with identical leg chaetotaxy only on the basis of genital chaetotaxy''. The interpretation changes if reduction gradients are taken into account: Homoiorthotydeus then becomes a subgenus of Tydeus, and Quadrotydeus, a subgenus of Brachytydeus, as illustrated in figures 1 and 2. In other words, the variation in the number of genitals is not a mere anomaly but a true vertition as indicated in figures 2 and 4. Asymmetry (3-2) is reported in B. (E.) mirabilis by Kaźmierski (1989: 301). A similar asymmetry in number of genital setae is observed 5 times in 20 females of Paratriophtydeus coineaui (Iolinidae): 14 times (3-3), 5 (4-3), 1 (4+4) (André, 1980, fig. 11D; André, 1981: 40).
Kaźmierski (1980: fig. 4) observed a 7th genital seta on one side of a male B. catenulatus. A similar anomaly was reported in Kazmierskius lundqvisti (5 out of 16 females) by Momen and Solhøy (1996: 25).
In Stigmaeidae and Raphignathidae, variations in the number of genital setae were reported by, respectively, Akyol (2011) and Akyol (2018).
Subgenera and fuzzy boundaries of genital setae
Based on Fig. 1, figure 6 of André (2025) is updated and its caption is revised (Figure 2 in this paper). The new version shows the overlap of subgenera delineated solely by genital chaetotaxy, a point already raised by Kaźmierski (1989: 300-301). B. (Q.) inconstans (n. comb.) is a good example of this, with 3 to 6 ge depending on individuals and which side of the body (= hemibody) is considered (Kaźmierski, 1989: 301). The trend observed in Brachytydeus, the gradual reduction of the genital chaetotaxy, is also found in Tydeus, with the subgenera Bakerpertydeus [5 (ge)] and Homoiorthotydeus [4 (ge)].
Evolutionary plasticity and fuzzy boundaries
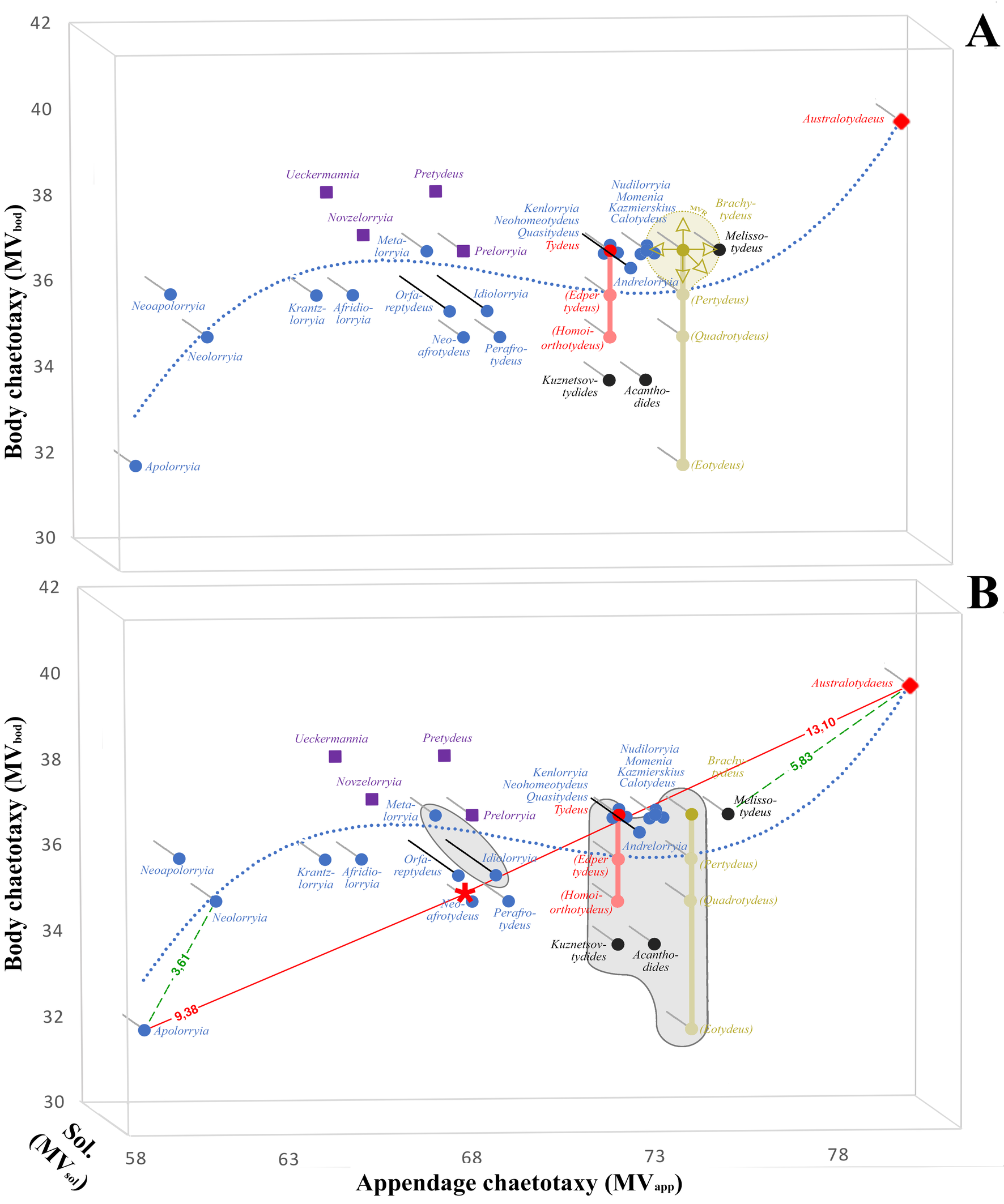
Plasticity of the genus Lorryia and the family Tydeidae has already been presented by André (2023, 2025). Figure 3a illustrates the tydeid genera and non-nominal subgenera in a 3-dimensional morphospace determined by meristic characters. The small symbols — different according to subfamilies — indicate the meristic values assigned to each genus and subgenus, these values being interpreted as hard borders. However, each genus and subgenus is also the centroid of a cluster, whatever the precise meaning of this term (see Estivill-Castro, 2002), composed of species and variant individuals. This meristic value range (MVR) is illustrated only for (Brachytydeus) and correspond to fuzzy boundaries. It is the narrowest volume which contains all the data of the taxon, excluding outliers (e.g., located 2 SD [standard deviation] from the rest of the data). The shape of the volume is variable and pertains to pattern recognition. In most cases, such an approach is impossible due to a lack of data.
Gaps, clusters and subfamilies
Figure 3b shows various gaps between genera. The most important one (5.39) separates Australotydaeus from Melissotydeus and isolates Australotydaeinae. A second gap detaches Apolorryia from Neolorryia (3.61). (Homoiorthotydeus) is separated from Perafrotydeus by only 3.00.
There is no discontinuity between Tydeinae (circles in fig. 3) and Pretydeinae (squares). However, the latter subfamily is easily recognized by a special chaetotaxy of legs III and IV as indicated in Fig. 1. At most, a central density-based cluster (Campello et al. 2020) can be perceived and would include the genera Tydeus and Brachytydeus.
An enumerative combinatorics of the morphospace delimited by extreme genera (i.e. Australotydaeus and Apolorryia) results in 528 combinations (22x8x3 along the 3 axes). If variants are excluded, only 34 vertices are occupied by males, which represents 6.4% of all the possibilities (12.9% with females lacking eugenitals). The gaps or discontinuities in the pattern or structure of possible combinations could result from our incomplete knowledge of the Tydeidae (only 3.72% of morphospecies have been described according to André (2025)) or from evolutionary processes that have yet to be explored.
Europeanism is also visible in figure 3b. The two central gray zones indicate that the holotype of the type species was collected in Europe.
Anomalies and vertitions in Tydeidae
The difference between a hard border and a fuzzy boundary is visually highlighted in figure 4. The hard border is visualized by a dark grey sphere and corresponds to the values in figure 1. In contrast, the fuzzy boundary takes variations into account and is represented by an ellipse around the sphere.

The difference between an anomaly and a vertition is also highlighted in figure 4. Evolutionary trends may be marked by vertitions as in Brachytydeus, or not as in Tydeus. Anomalies, devoid of any evolutionary significance, are observed for Neoafrotydeus.
Importance of variations: from genus to population level
Baker (1968a: 1002-1003, figs 42-44) described Lorryia macquillani on apple trees in Ireland. After studying the holotype, a new genus, Idiolorryia, was erected by André (1980: 117) to accommodate macquillani. A second species, named I. marci, was added by André (1984: 154-156, fig. 3). The second species was collected only in Ruette (Belgium), most often in thalli of Ramalina farinacea (lichen) growing on an ash tree (Fraxinus excelsior). The new species was similar to I. macquillani. In particular, the reticulate pattern was virtually identical. However, I. marci had only one seta on tibia II instead of two as in I. macquillani. In addition, the ornamented dorsal setae located on the opisthosoma and on some podomeres were finer and longer than in the Irish species. The underlying hypothesis (H0) — two species with dissimilar chaetotaxy assigned to the same genus — is not congruent with the definition of genera presented in this article, where each genus has its own phanerotaxy.
Other hypotheses exist: the Belgian specimens could be a variant of I. macquillani. Do they represent a new subgenus to be named I. (Newsubgenus) marci (H1), a subspecies, I. macquillani marci (H2), or a particular population collected from several lichens growing on a single tree (H3)? No observations allow us to decide between these hypotheses. I. macquillani has only been recaptured in Greece by Panou and Emmanouel (1995). Idiolorryia and its putative variants are illustrated in detail in figure 4.
Nomenclatural acts
Kazmierskius n. gen.
ZOOBANK: DC833E13-A8BF-4DC5-A08E-4525F3AFE127 ![]()
The first nomenclatural act concerns Kazmierskius (n. gen.). Kaźmierski (1998a: 337) first spotted the generic taxon, which he named ''Generic Unit cf. Lorryia'', but retained the original name, Tydeus lundqvisti, given by Momen and Solhøy (1996). The type species is Tydeus lundqvisti renamed K. lundqvisti (n. comb.). The new genus is dedicated to A. Kaźmierski in recognition of his numerous contributions, which significantly advanced the systematics of the Tydeidae.
Edpertydeus n. subgen.
ZOOBANK: F85799E1-EAFC-438A-A457-5F037A85A4DD ![]()
Next nomenclatural changes concern Edpertydeus (n. subgen.) with Tydeus tuttlei as type species renamed T. (E.) tuttlei (n. comb.), and T. (Homoiorthotydeus) (n. comb.) with H. quadrisetosus as type species renamed B. (H.) quadrisetosus (n. comb.). The gradual reduction of the genital chaetotaxy in Brachytydeus affects the subgenera Pertydeus [5 (ge)], Quadrotydeus [4 (ge)] and Eotydeus [2 (ge)]. Accordingly, the nomenclatural changes concern Brachytydeus (Pertydeus) (n. comb.) with T. (P.) schusteri as type species renamed B. (P.) schusteri (n. comb.), Brachytydeus (Quadrotydeus) (n. comb.) with Q. sleipneri as type species renamed B. (Q.) sleipneri (n. comb.) and B. (Q.) inconstans (n. comb.), as well as Brachytydeus (Eotydeus) (n. comb.) with E. mirabilis as type species renamed B. (E.) mirabilis (n. comb.).
Discussion and conclusions
Number of genera and subgenera
Figure 1 presents 29 genera and 5 non-nominal subgenera, valid taxa based on imagines. In addition, there are 2 genera inquirenda (Lasiotydaeus and Melanotydaeus) as well as Edlorryia based on the deutonymph only.
In the past (e.g. Thor, 1933; Baker, 1965, 1968a, 1970), tydeid genera were defined by a particular striation and the leg chaetotaxy was variable. Such a generic description based on morphometric characters in the broad sense, on a particular striation pattern in this case, is incompatible with a definition based on meristic characters as advanced by Kaźmierski (1989, 1998a, 1998b) or with a classification of genera also based on meristic characters as that proposed by André (2025). This paradigm shift results in the dismantling of the genus Paralorryia and the distribution of its species into six genera (André, 2026). Such an approach was already suggested by Fain (1963: 3) for Speleognathinae (Ereynetidae).
Normality and variations
Variants are not necessarily outliers (e.g., located 2 SD [standard deviation] from the rest of the data) and variations may be part of the normality even if the term ''normal'' is polysemous. Sometimes, ''it is consistent with our habits of mind to consider as abnormal anything relatively rare or exceptional'' (Bergson, 1932: 26; 1935: 26). ''Sometimes it designates a fact that can be described through statistical sampling; it refers to the mean of measurements made of a trait displayed by a species and to the plurality of individuals displaying this trait'' (Canguilhem, 1965: 200; 2008: 122).
In acarology too, the term is polysemous. Grandjean (1972: 457) suggested that vertitions were a normal trait but, according to Akyol (2011: 60, 62), ''′abnormal′ refers to a variation'' in chaetotaxy. The Spanish specimen of B. (B.) reticulata with no apotele IV is said to be abnormal (Kaźmierski and Sikora, 2008: 88). André (1981: 42) opposed ''normal setae'' to eupathidia (the term used by Grandjean to designate mechanoreceptor seta (e.g. 1964: 186) was ''ordinaire'' [ordinary]). The same meaning is found in the manual of acarology (Krantz 2009: 18, 50). However, this work does not address teratological systems such as that described by Stojnić et al. (2000).
In Tydeoidea, the term ''normal'' often referred to the usual situation: the usual ''position'' (André, 1981: 42), ''location'' (André and Fain, 2000: 420-421), number of setae (Khaustov et al. 2024: fig. 6), ''striation pattern'' (André, 1981: 32) or ''development'' (André,1981: 40). Legs were normal according to Baker (1945: 16).
Normality is not just an academic issue; it has a direct impact on taxonomy. The single specimen of Lorryia superba was considered abnormal (Kaźmierski, 1998: 301; Kaźmierski and Sikora, 2008: 88) or was treated as such (Mondin et al., 2016: 474; Kaźmierski et al., 2018: 813). In this context, Brachytydeus was considered a junior synonym of Lorryia (Akbari et al., 2015; Mondin et al., 2016; Kaźmierski et al., 2018; Da-Costa et al., 2019). Conversely, the single individual described by Oudemans was supposed to be normal and the genus Lorryia was maintained in its pristine state by André (2023). A similar case concerns the single individual of Brachytydeus globulipalpus: is it a valid species described by Kaźmierski (1998: 304) and based on a ''normal'' specimen treated by Kaźmierski and Sikora (2008: 82, 85), a variant of volgini, the initial determination made by Kaźmierski (1980: 184), or a variant of globulipalpus?
As pointed out by Coineau (1974: 141), individual variations are generally not appreciated by systematists who regret not finding in these characters the rigor they expected. Hard borders, as those in figure 1, are preferred to fuzzy boundaries. Yet, variations may be important: 67.6% of 68 adult specimens of Torotrogla merulae (Syringophilidae) present anomalies of aggenital setae (data from Głowska and Skoracki, 2009: 366). Normality and variations have also an influence on the biodiversity based on mites as demonstrated by Lienhard and Krisper (2021, especially figs 4, 5).
Discreteness vs continuity
The question of discreteness vs. continuity arises in the systematics of mites and concerns all taxa. This debate implies two philosophical approaches to interpreting natural systems. ''One of the most outstanding properties of natural diversity is its discreteness and order. Species can be identified and classified because of this property.'' (Alberch, 1989: 21). This perspective tends to define hard borders as illustrated in figures 1 and 2A.
In contrast, the idea of using fuzzy logic has not been widely adopted in taxonomy (Todisco and Nazari, 2023: 157) and has not been applied to mites. ''Taxonomy is a discrete ordering system imposed upon the continuous structure of the Tree of Life.'' (Zachos, 2016: 14) or ''the chaos that is life'' (Todisco and Nazari, 2023: 156). This approach tends to define fuzzy boundaries ''on a continuous process and a continuous organismic world with vague or fuzzy boundaries'' (Zachos, 2016: 5).
A final approach is to compare the variation in traits between taxa and simply state: ''from so simple a beginning endless forms most beautiful and wonderful have been, and are being, evolved'' (Darwin, 1859: 490).
Acknowledgements
I am grateful for the suggestions made by A. Khaustov and H. Proctor who analyzed drafts of this article and for remarks made by anonymous referees. Preliminary drafts were composed when I was hospitalized at HFR Riaz, special thanks to Dr. F. Cilla and the team of rehabilitation. A special thank also to my wife who supported me.
References
- Abé H. 2024. Distribution and morphological variation of the freshwater halacarid mite Lobohalacarus weberi (Romijn & Viets, 1924) (Acari, Halacaridae) in Japan. Ecologica Montenegrina, 74: 89-98. https://doi.org/10.37828/em.2024.74.12
- Akbari A., Irani-Nejad K.H., Khanjani M., Arzanlou M., Kaźmierski A. 2015. A new tydeid species (Acari: Tydeidae) with a key to Brachytydeus species from East Azerbaijan Province, Iran. Systematic and Applied Acarology, 20(4): 423-443. https://doi.org/10.11158/saa.20.4.7
- Akyol M. 2011. A new record of Storchia ardabiliensis and variations in the number of genital, aggenital and external clunal setae in two Storchia species (Acari: Stigmaeidae) from the aegean coast, Turkey. Systematic and Applied Acarology, 16(1): 59-66. https://doi.org/10.11158/saa.16.1.8
- Akyol M. 2018. Variations in the number of genital setae in Raphignathus hecmatanensis species (Acari: Raphignathidae). Kastamonu University Journal of Forestry Faculty, 18(2): 121-125. https://doi.org/10.17475/kastorman.305126
- Alberch P. 1989. The logic of monsters: Evidence for internal constraint in development and evolution. Geobios, 22 (Supplement 2): 21-57. https://doi.org/10.1016/S0016-6995(89)80006-3
- André, H. 1979 (1978). A generic revision of the family Tydeidae (Acari: Actinedida). I. Introduction, paradigms and general classification. Annales de la Société royale zoologique de Belgique, 108 : 189-208.
- André H.M. 1980. A generic revision of the family Tydeidae (Acari : Tydeidae). IV. Generic descriptions, keys and conclusions. Bulletin & annales de la Société entomologique de Belgique, 116: 103-130, 139-168.
- André H.M. 1981a. A generic revision of the family Tydeidae (Acari: Actinedida). II. Organotaxy of the idiosoma and gnathosoma. Acarologia, 22(1): 31-46.
- André, H. M. 1981b. A generic revision of the family Tydeidae (Acari: Actinedida). III. Organotaxy of the legs. Acarologia, 22(2): 165-178.
- André H.M. 1984. Tydeinae (Acari : Tydeidae) from Belgium. I. The genus Homeotydeus. Bulletin & annales de la Société entomologique de Belgique, 120: 117-122.
- André H.M. 1985. Redefinition of the genus Triophtydeus Thor, 1932 (Acari : Actinedida). Zoologische Mededelingen Leiden, 59: 189-195.
- André H.M. 2005. In search of the true Tydeus (Acari: Tydeidae). Journal of natural History, 39: 975-1001 https://doi.org/10.1080/00222930400002838
- André H.M. 2021. The Tydeoidea (Ereynetidae, Iolinidae, Triophtydeidae and Tydeidae) - An online database in the Wikispecies platform. Acarologia, 61(4): 1023-1035. https://doi.org/10.24349/6yc5-1lxw
- André H.M. 2023. Lorryia (Acariformes, Tydeidae): The evolutionary plasticity of an enigmatic genus. Acarologia, 63(3): 844-855. https://doi.org/10.24349/6btk-vacc
- André H.M. 2025 Sixty years after ''A review of the genera of the family Tydeidae» (Acariformes: Tydeoidea: Tydeidae). Acarologia, 65(1): 149-172. https://doi.org/10.24349/l5vb-767l
- André H.M. 2026. Dismantling of the genus Paralorryia (Acariformes: Tydeoidea: Tydeidae). Acarological Studies, 8(1). https://doi.org/10.47121/acarolstud.1775112
- André H.M., Ducarme X. 2003. Rediscovery of the genus Pseudotydeus (Acari: Tydeoidea), with description of the adult using digital imaging. Insect Systematics & Evolution, 34: 373-380. https://doi.org/10.1163/187631203X00027
- André H.M., Ducarme X., Lebrun, P. 2004 New ereynetid mites (Acari: Tydeoidea) from karstic areas: True association or sampling bias? Journal of Cave and Karst Studies, 66: 81-88.
- André H.M., Fain A. 2000. Phylogeny, ontogeny and adaptive radiation in the superfamily Tydeoidea (Acari: Actinedida), with a reappraisal of morphological characters. Zoological Journal of the Linnean Society, 130: 405-448. https://doi.org/10.1111/j.1096-3642.2000.tb01636.x
- André M., Naudo M.H. 1965. Pertydeus schusteri, n. sgen., n. sp., nouveau Tydeus à griffe pulvillaire (Tydeidae). Acarologia, 7(4): 673-682.
- André H.M., N′Dri J.K. 2013 (2012). Bréviaire de taxonomie des acariens. Abc Taxa 13, Bruxelles, Belgique. http://www.taxonomy.be/gti_abctaxa/volumes.
- Baker E.W. 1943. Nuevos Tydeidae Mexicanos (Acarina). Revista de la Sociedad Mexicana de Historia Natural, 4(3-4): 181-189.
- Baker E.W. 1945. Five mites of the family Ereynetidae from Mexico. Journal of the Washington Academy of Sciences, 35(1): 16-19.
- Baker E.W. 1965. A review of the genera of the family Tydeidae (Acarina). In: Naegele JA, editor. Advances in acarology 2. Ithaca (NY): Cornell University Press. p 96-133.
- Baker E.W. 1968a. The genus Lorryia. Annals of the Entomological Society of America, 61(4): 986-1008. https://doi.org/10.1093/aesa/61.4.986
- https:// doi.org/10.1093/aesa/61.4.986
- Baker E.W. 1968b. The genus Pronematus Canestrini. Annals of the Entomological Society of America, 61(5): 1091-1097. https://doi.org/10.1093/aesa/61.5.1091
- Baker E.W. 1970. The genus Tydeus: subgenera and species groups with descriptions of new species (Acarina: Tydeidae). Annals of the Entomological Society of America, 63(1): 163-177. https://doi.org/10.1093/aesa/63.1.163
- Baker E.W., Wharton G.W. 1952. An Introduction to Acarology. New York, The Macmillan Company.
- Bergson H. 1932. Les deux sources de la morale et de la religion, Félix Alcan, Paris.
- Bergson H. 1935. The two sources of morality and religion. Translated by R. Ashley Audra and C. Brereton with the assistance of W.H. Carter. Macmillan and Co., London.
- Berlese A. 1882. Acari, Myriopoda et Scorpiones hucusque in Italia reperta. Ordo Prostigmata (Trombidiidae). Padova, sumptibus auctoris. https://doi.org/10.5962/bhl.title.69269
- Booth R.G. 1983. A description of the juvenile life-stages of the Antarctic mite Ereynetes macquariensis. British Antarctic Survey Bulletin, 61: 7-12.
- Brunetti C., Siepel H., Fanciulli P.P., Nardi F., Convey P., Carapelli A. 2021. Two New Species of the Mite Genus Stereotydeus Berlese, 1901 (Prostigmata: Penthalodidae) from Victoria Land, and a Key for Identification of Antarctic and Sub-Antarctic Species. Taxonomy, 1: 116-141. https://doi.org/10.3390/taxonomy1020010
- Budd G.E. 2021. Morphospace. Current Biology, 31(19): 1181-1185. https://doi.org/10.1016/j.cub.2021.08.040
- Bukva V. 1990. Three species of the hair follicle mites (Acari: Demodicidae) parasitizing the sheep, Ovis aries L. Folia Parasitolica 37(1): 81-91.
- Campello R.J.G.B., Kröger P., Sander J., Zimek A. 2020. Density-based clustering. WileyInterdisciplinary Reviews: Data Mining and Knowledge Discovery, 10(2), Article e1343. https://doi.org/10.1002/widm.1343
- Canguilhem G. 1965. La Connaissance de la vie, Paris, Vrin.
- Canguilhem G. 2008. Knowledge of Life. Translated by S. Geroulanos & D. Ginsburg. Fordham University Press, New York.
- Carignan C., Casillas J.V., Chodroff E., Zellou G. 2022. Editorial: Fuzzy boundaries: Ambiguity in speech production and comprehension. Frontiers in Communication, 7: 1112753. https://doi.org/10.3389/fcomm.2022.1112753
- Coineau Y. 1974. Éléments pour une monographie morphologique, écologique et biologique des Caeculidae (Acariens). Mémoires du Muséum national d'histoire naturelle. Série A, Zoologie, 81(1): 1-299.
- Da-Costa T., Rodighero L.F., Silva G.L. da, Ferla N.J., Blochtein B. 2019. Two new species of Tydeidae (Acari: Prostigmata) associated with stingless bees. Zootaxa, 4652(1): 101-112. https://doi.org/10.11646/zootaxa.4652.1.4
- Darwin C. 1859. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life (1st ed.). John Murray, London. https://doi.org/10.5962/bhl.title.68064
- Darwin C. 1958. (Barlow N. ed.) The autobiography of Charles Darwin 1809-1882. With the original omissions restored. Edited and with appendix and notes by his grand-daughter Nora Barlow. Collins, London.
- Dastych H. 1990. Some notes on Antarctic mites (Acari). Entomologische Mitteilungen aus dem Zoologischen Museum Hamburg,, 10 (139/140): 43-56.
- De Vis, R.M.J., Lacerda, J.D.A., Moraes, G.J. de, Ueckermann, E.A. 2025. Pronematinae (Trombies: Iolinidae) from Brazil, with original and complementary descriptions of taxa as well as with a reappraisal of the subfamily and of Parapronematus. Acarologia, 65(2): 331-372. https://doi.org/10.24349/7w23-xsl9 https://doi.org/10.24349/7w23-xsl9
- Doğan S., Doğan S., Zeytun E. 2019. On newly discovered deutonymphal stage of Stigmaeus kumalariensis Akyol & Koç (Acari: Stigmaeidae) from Turkey, with numerical and structural variations in adults. Trakya University Journal of Natural Sciences, 20(2): 115-120. https://doi.org/10.23902/trkjnat.540747
- Doğan S., Fan, Q.-H., Uğurlu, Ş, Erman, O. 2022. First detection of numerical variations in aggenital setae of female Eustigmaeus segnis (Koch) (Acariformes: Stigmaeidae). In: Zhang, Z.-Q., Fan, Q.-H., Heath, A.C.G. & Minor, M.A. (Eds) (2022) Acarological Frontiers: Proceedings of the XVI International Congress of Acarology (1-5 Dec. 2022, Auckland, New Zealand). Magnolia Press, Auckland, 328 pp.
- Duden online. https://www.duden.de/woerterbuch. Accessed 2026.04.09.
- Estivill-Castro V. 2002. Why so many clustering algorithms - a position paper. ACM SIGKDD Explorations Newsletter, 4(1): 65-75. https://doi.org/10.1145/568574.568575
- Fain A. 1956. Nouvelles observations sur les Acariens de la famille Speleognathidae parasites des fosses nasales chez les batraciens, les oiseaux et les mammifères. Annales de parasitologie humaine et comparée, 31: 643-662. https://doi.org/10.1051/parasite/1956315643
- Fain A. 1963. Chaetotaxie et classification des Speleognathinae. Bulletin de l′Institut royal des Sciences naturelles de Belgique, 39(9): 1-80.
- Fain A., Nadchatram M. 1962. Acariens nasicoles de Malaisie I. Ereynetoides malayi n.g., n. sp., parasite d'un nectarin (Ereynetidae: Trombidiformes). Zeitschrift für Parasitenkunde, 22: 68-82. https://doi.org/10.1007/BF00260166
- Głowska E. & Skoracki M. 2009. Anomalous chaetotaxy in the quill mites Torotrogla merulae Skoracki et al., 2000 (Acari, Prostigmata, Syringophilidae). Acta Parasitologica, 54(4): 364-367. https://doi.org/10.2478/s11686-009-0046-y
- González-González J. 1998. Transformed and altered processes: foundations for a processual theory of biological knowledge. In: Ługowski, W. & Matsuno, K. (Eds) Uroboros or biology between mythology and philosophy. Warszawa Arboretum, Poland.
- Grandjean F. 1938. Retetydeus et les stigmates mandibulaires des Acariens prostigmatiques. Bulletin du Muséum national d'histoire naturelle, 10: 279-286.
- Grandjean F. 1949.Au sujet des variations individuelles et des polygones de fréquence. Comptes rendus hebdomadaires des séances de l′Académie des sciences, Paris, 229 : 801-804.
- Grandjean F. 1952. Sur les variations individuelles. Vertitions (écarts) et anomalies. Comptes rendus hebdomadaires des séances de l′Académie des sciences, Paris, 235 : 640-642.
- Grandjean F. 1964. New observations on Oribatid mites (3rd serie). 1. Anhomologous tautergy. 2. Sexual differentiation of setae in tarsus 1 male. Acarologia, 6(1): 170-198.
- Grandjean F. 1972. Caractères anormaux et vertitionnels rencontrés dans des clones de Platynothrus peltifer (Koch). Chapitre I à VI de la deuxième partie. Acarologia, 14(3): 454-478.
- Hammen, L. van der. 1976. Opilioacarida. Glossaire de la terminologie acarologique / (Glossary of acarological terminology), vol 2. Springer, Dordrecht. https:// doi.org/10.1007/978-94-010-1584-4_10
- Hammen L. van der. 1981. Numerical changes and evolution in actinotrichid mites (Chelicerata). Zoologische Verhandelingen, 182: 1-46.
- Kaźmierski A. 1980. Materiały do znajomości fauny Tydeidae (Acari: Prostigmata) w Polsce. I. Rodzaj Lorryia Oudemans. Prace Komisji Biologicznej, Towarzystwo Przyjaciół Nauk Poznańskie (PTPN), 54: 87-129.
- Kaźmierski A. 1989. Revision of the genera Tydeus Koch sensu André, Homeotydeus André and Orthotydeus André with description of a new genus and four new species of Tydeinae (Acari: Actinedida: Tydeidae). Mitteilungen Hamburgisches Zoologisches Museum und Institut, 86: 289-314.
- Kaźmierski A. 1996. A revision of the subfamilies Pretydeinae and Tydeinae (Acari, Actinedida: Tydeidae). Part II. The subfamilies Pretydeinae André, 1979 new taxa, species review, key and considerations. Mitteilungen aus dem Hamburger Zoologischen Museum und Institut 93: 171-198.
- Kaźmierski A. 1998a. Tydeinae of the world: generic relationships, new and redescribed taxa and keys to all species. A revision of the subfamilies Pretydeinae and Tydeinae (Acari:Actinedida: Tydeidae) - part IV. Acta Zoologica Cracoviensia 41(2): 283-455.
- Kaźmierski A. 1998b. Revision of the genera Tydeus Koch sensu André, Homeotydeus André and Orthotydeus André with description of a new genus and four new species of Tydeinae (Acari: Actinedida: Tydeidae). Mitteilungen Hamburgisches Zoologisches Museum und Institut, 86: 289-314.
- Kaźmierski A., Marciniak M., Sikora B. 2018. Tydeinae mites (Acariformes: Prostigmata: Tydeidae) from bird nests with description of three new species. Systematic and Applied Acarology, 23(5): 803-823. https://doi.org/10.11158/saa.23.5.3
- Kaźmierski A., Sikora B. 2008. On the morphological anomalies in Tydeoidea (Actinedida). In: Bertrand M., Kreiter S., McCoy K.D., Migeon A., Navajas M., Tixier M.-S., Vial L. (eds.) Integrative Acarology Proceedings of the 6th European Congress. Montpellier, European Association of Acarologists, pp. 82-88.
- Khaustov A.A. 2015. A new species of the genus Protopenthalodes (Acari: Penthalodidae) from Crimea. Acarina 23(2): 139-151.
- Khaustov A.A. 2022. Review of the Mediolata (Acari: Stigmaeidae) of Russia. Acarina, 30(1): 29-56. https://doi.org/10.21684/0132-8077-2022-30-1-29-56
- Khaustov A.A., Kolesniko V.B., Turbano I.S., Tolstiko A.V. 2024. A new troglobiont species of Rhagidiidae (Acari: Eupodoidea)from cave sites in the Ural Mountains (Russia). Acarologia, 64(3): 787-802. https://doi.org/10.24349/o7sw-gepq
- Kitajima E.W., Tassi A.D., Novellis V.M., Caceres S., Aguirre A., Costa N., Moraes G.J. De. 2011. Asymmetry in the number of solenidia on tarsi II of Brevipalpus (Acari: Tenuipalpidae) populations from Argentina. Zoosymposia, 6: 39-44. https://doi.org/10.11646/zoosymposia.6.1
- Klimov, P.B., Kolesnikov, V.B., Vorontsov, D.D., Ball A.D., Bolton S.J., Mellish C., Edgecombe G.D., Pepato A.R., Chetverikov L.E., He Q., Perotti M.A., Henk R. Braig H.R. 2025. The evolutionary history and timeline of mites in ancient soils. Scientific Reports, 15: 13555 https://doi.org/10.1038/s41598-025-96115-2
- Knop N., Hoy M. 1983. Biology of a tydeid mite, Homeopronematus anconai (n. comb.) (Acari: Tydeidae), important in San Joaquin Valley vineyards. Hilgardia, 51(5): 1-30. https://doi.org/10.3733/hilg.v51n05p030
- Krantz G.W. 2009. Form and function. In: Krantz G.W., Walter D.E. (eds) A manual of acarology, 3rd ed. Texas Tech University Press, Lubbock: 5-53.
- Langacker R.W. 2006. On the continuous debate about discreteness. Cognitive Linguistics, 17(1): 107-151. https://doi.org/10.1515/COG.2006.003
- Lawing A.M., Meik J.M., Schargel, W.E. 2008. Coding meristic characters for phylogenetic analysis: a comparison of step-matrix gap-weighting and generalized frequency coding. Systematic Biology, 57(1):167-173. https://doi.org/10.1080/10635150801898938
- Leponce M., Noti M.-I., Bauchau V., Wauthy G. 2001. ′Vertition′ of integumental organs in mites revisited: a case of fluctuating asymmetry. Mise au point sur les ′vertitions' des organes tégumentaires chez les acariens : un cas dˈasymétrie fluctuante. Comptes rendus de l′Académie des Sciences - Séries III - Sciences de la Vie, 324(5): 425-431. https://doi.org/10.1016/S0764-4469(01)01314-2
- Lienhard A., Krisper G. 2021 Hidden biodiversity in microarthropods (Acari, Oribatida, Eremaeoidea, Caleremaeus). Scientific Reports, 11. https://doi.org/10.1038/s41598-021-02602-7
- Lorençon J.R., Andrade S.C., Andrade D.J. 2016. Mites occurrence on Pachira aquatica Aubl. including aspects of external mouthpart morphology of Brachytydeus formosa (Acari: Tydeidae). Brazilian Journal of Biology, 76(1): 136-143. https:// doi.org/10.1590/1519-6984.15114 https://doi.org/10.1590/1519-6984.15114
- Maggenti M.A., Maggenti A.R., Gardner S. 2008. Dictionary of Invertebrate Zoology. Zea EBooks, 1-379.
- Marini F., Weyl P., Vidović B., Petanović R., Littlefield J., Simoni S., Lillo E. de, Cristofaro M., Smith L. 2021. Eriophyid mites in classical biological control of weeds: progress and challenges. Insects 12(6): 513. https://doi.org/10.3390/insects12060513
- Momen F.M., Lundqvist L. 1993. Inconsistencies in leg chaetotaxy in two species of Tydeid mites (Prostigmata: Tydeidae). International Journal of Acarology, 19(2): 137-144. https://doi.org/10.1080/01647959308683972
- Momen F.M., Solhøy T. 1996. A first record of the genus Tydeus in Himalaya, Tydeus lundqvisti nov. sp. (Acari: Actinedida: Tydeidae). Acarologia, 37(1): 23-25.
- Mondin A. de S., Nuvoloni F.M., Feres, R.J.F. 2016. Four new species of Lorryia (Acari: Tydeidae) associated with Hevea brasiliensis Muell. Arg. (Euphorbiaceae) in Brazil. Zootaxa, 4158(4): 473-490. https://doi.org/10.11646/zootaxa.4158.4.2
- Navia D., Ferreira C.B.S., Reis A.C., Gondi M.G.C. 2015. Traditional and geometric morphometrics supporting the differentiation of two new Retracrus (Phytoptidae) species associated with heliconias. Experimental & applied acarology 67(1): 87-12. https://doi.org/10.1007/s10493-015-9934-z
- Oudemans A.C. 1928 (1927). Acari uit Ambon. Zoologische Mededelingen, 10: 185-237.
- Palmer A.R., Strobeck C. 1986. Fluctuating asymmetry: measurement, analysis, patterns. Annual Review of Ecology and Systematics, 17: 391-421. https://doi.org/10.1146/annurev.es.17.110186.002135
- Panou H.N., Emmanouel N.G. 1995. New records of tydeid mites from Greece with description of Lorryia mantiniensis sp. nov. (Acari: Tydeidae). International Journal of Acarology, 21(1): 17-21. https://doi.org/10.1080/01647959508684038
- Paredes-León R., Corona-López A.M., Flores-Palacios A., Toledo-Hernández V.H. 2019. Camerobiid mites (Acariformes: Raphignathina: Camerobiidae) inhabiting epiphytic bromeliads and soil litter of tropical dry forest with analysis of setal homology in the genus Neophyllobius. European Journal of Taxonomy, 202: 1-25.
- Per S., Doğan S., Zeytun E., Ayyildiz N. 2017. Description of a new rake legged mite of the genus Allocaeculus (Acariformes: Caeculidae) from Turkey with description of variation in dorsal setation. Acarologia, 57(2): 369-382. https://doi.org/10.1051/acarologia/20164162
- Schiess T. 1981. Neue Tydeidenarten (Acari, Actinedida, Tydeidae) aus einem alpinen Rasen (Caricetum firmae, 2500 m) des Schweizer Nationalparkes. Entomologica Basiliensia, 6: 78-107.
- Sidorchuk, E.A., Khaustov, A.A. 2018. Two Eocene species of peacock mites (Acari: Tetranychoidea: Tuckerellidae). Acarologia, 58(1): 99-115. https:// doi.org/10.24349/acarologia/20184228. https://doi.org/10.24349/acarologia/20184228
- Skoracki M., Schmidt K.-H., Marciniak N., Marciniak M. 2018. A review of mites of the subfamily Speleognathinae (Acariformes: Ereynetidae) parasitizing respiratory tracts of birds in the Afrotropical region. Zootaxa, 4403(3): 401-440. https://doi.org/10.11646/zootaxa.4403.3.1
- Spain A.V. 1969. A new genus and species of Tydeidae (Acari: Prostigmata) from New Zealanad. Acarologia, 11: 23-28.
- Stojnić B., Sekulic D., Petanović R. 2000. A case of monstra duplicia in Tetranychus urticae Koch (Acari: Tetranychidae). Acarologia, 41(1-2): 227-232.
- Thor S. 1932. Opsereynetes, eine neue augentragende Gattung aus der Familie Ereynetidae Oudemans, 1931, in Norwegen gefunden. Zoologische Anzeiger, 99: 27-30.
- Thor S. 1933. Tydeidae, Ereynetidae. Das Tierreich, 60: 1-84.
- Todisco V., Nazari V. 2023. Is the concept of species a matter of sex or descent, or is it just ''fuzzy″? Biological Journal of the Linnean Society 140: 156-158. https://doi.org/10.1093/biolinnean/blad097
- Van Valen L. 1962. A study of fluctuating asymmetry. Evolution 16(2): 125-142. https://doi.org/10.1111/j.1558-5646.1962.tb03206.x
- Walter D.E. 2006. Invasive Mite Identification: Tools for Quarantine and Plant Protection. https://keys.lucidcentral.org/keys/v3/mites/Invasive_Mite_Identification/key/Whole_site/Introduction.htm
- Wauthy G., Noti M.-I., Leponce M., Bauchau V. 1998. Taxy and variations of leg setae in Tetranychus urticae (Acari: Prostigmata). Acarologia, 39(3): 233-255.
- Wood T.G. 1965. New and redescribed species of Tydeidae (Acari) from moorland soils in Britain. Acarologia, 7(4): 663-672.
- Zachos F.E. 2016. Species Concepts in Biology: Historical Development, Theoretical Foundations and Practical Relevance. Springer International Publishing Switzerland.



2026-03-15
Date accepted:
2026-04-20
Date published:
2026-05-05
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 André, Henri M.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)