



Complementary descriptions of two species of the subgenus Anthoseius De Leon (Parasitiformes: Phytoseiidae) collected from wild and cultivated apple trees in the western Himalayas, India
Döker, Ismail  1
; Singh, Robin
2
; Kaur, Paramjit
3
and Chinnamade Gowda, Channegowda
4
1
; Singh, Robin
2
; Kaur, Paramjit
3
and Chinnamade Gowda, Channegowda
4
1✉ Cukurova University, Agricultural Faculty, Department of Plant Protection, Acarology Lab, Adana, Türkiye.
2Punjab Agricultural University, Department of Entomology, 141004 Ludhiana, Punjab, India.
3Punjab Agricultural University, Department of Entomology, 141004 Ludhiana, Punjab, India.
4University of Agricultural Sciences, Department of Agricultural Entomology, Bangalore, India.
2026 - Volume: 66 Issue: 2 pages: 394-405
https://doi.org/10.24349/f2sw-8vzeZooBank LSID: D6519525-8730-4B70-8DAF-F0FBD10966E4
Original research
Keywords
Abstract
Introduction
Phytoseiid mites (Parasitiformes: Phytoseiidae) are a highly diverse group of predatory mites widely recognized for their importance in the biological control of phytophagous mites and other small arthropods (McMurtry et al. 2013; Kreiter et al. 2021; Ferragut and Navia 2024; Tsolakis and Ragusa 2016). Accurate species identification is a critical step for the effective implementation of integrated pest management programs (Gerson 2014). However, many earlier descriptions were based on limited material and insufficient diagnostic information, leading to taxonomic uncertainties that continue to complicate systematic and faunistic studies (Döker et al. 2025a). Consequently, detailed morphological examinations and redescriptions based on types or newly collected and well-preserved specimens remain essential for improving species delimitation and supporting reliable identification.
Within the subfamily Typhlodrominae, the subgenus Anthoseius De Leon is the most species-rich lineage, with approximately 400 nominal species, including about 60 from the Oriental region (Hernandes et al. 2012; Demite et al. 2026). Based on shared morphological characters, Chant and McMurtry (1994, 2007) established several species groups within this subgenus to facilitate classification. Among these, the rickeri species group was defined by a distinctive combination of characters, particularly the marked disparity in dorsal setal lengths, the presence of an elongate tubular calyx of spermatheca, and reduced cheliceral dentition. The group includes approximately 20 described species primarily distributed in India, Pakistan, China, and Southeast Asia (Wu et al. 2009; Demite et al. 2026). Furthermore, some authors have treated this group as a separate genus, Orientiseius Muma and Denmark, 1968 or as a subgenus (Kolodochka 2006). Nevertheless, the original descriptions of several included taxa lacked comprehensive morphological details, indicating the need for additional documentation based on newly collected material.
Although phytoseiid mites have been reported from various agricultural ecosystems, including citrus, grapevine, and apple orchards, their diversity in high-altitude environments remains insufficiently studied (Döker et al. 2025b; Robin et al. 2025). The western Himalayas represent a unique ecological region characterized by diverse agroecosystems, yet information on phytoseiid fauna from high-altitude apple habitats is still limited.
In the present study, we surveyed phytoseiid mites in high-altitude apple orchards in the north western Himalayas, India. Complementary descriptions of T. (A.) agilis (Chaudhri, 1975) and T. (A.) malicolus Wainstein and Arutunjan, 1967 are provided based on newly collected specimens. Typhlodromus (A.) agilis is recorded for the first time from India. In addition, comparative examination of original descriptions indicates that certain taxa previously described under different names represent junior synonyms of T. (A.) malicolus. The present study contributes to clarifying the taxonomy of species within the rickeri species group and enhances knowledge of phytoseiid species in high-altitude Himalayan ecosystems.
Materials and Methods
Field surveys were conducted in Lahaul and Spiti district of Himachal Pradesh, India. Adult phytoseiid mites were collected from apple trees by shaking branches over a white plastic tray using a wooden stick. Dislodged specimens were collected with a fine camel-hair brush (size 000), preserved in 70% ethanol and transferred to the laboratory. Specimens were cleared in 60% lactic acid and kept in cavity slides at 50 °C for 24 h. Mites were mounted on permanent microscope slides in Hoyer's medium under a stereozoom microscope (Olympus Magnus). Slides were dried in a hot-air oven at 55 °C for 48 h and sealed with nail polish. Measurements were taken using an Olympus CX-41 compound microscope equipped with a calibrated ocular micrometer. Illustrations were prepared with a camera lucida and finalized using Adobe Photoshop (CS6). Species identification followed the classification system of Chant and McMurtry (2007). Dorsal idiosomal setal nomenclature follows Lindquist and Evans (1965), as modified by Rowell et al. (1978). Ventral setal nomenclature follows Chant and Yoshida-Shaul (1991). Adenotaxy and poroidotaxy are interpreted according to Athias-Henriot (1971, 1975), and leg chaetotaxy follows Evans (1963).
All measurements are given in micrometers (µm) as means with ranges in parentheses. Dorsal shield length was measured along the midline from the anterior to the posterior margin. Genital shield length was measured along the midline, including the anterior hyaline part. Ventrianal shield length was measured along the midline, including the cribrum. Leg length was measured from the base of the coxa to the apex of the tarsus, excluding the ambulacrum.
Voucher specimens are deposited in the mite collections of the All India Network Project on Agricultural Acarology, University of Agricultural Sciences, Bangalore, India; the Acarology Laboratory, Department of Entomology, Punjab Agricultural University, Ludhiana, India; and the Acarology Laboratory, Department of Plant Protection, Çukurova University, Adana, Türkiye.
Results
Typhlodromus (Anthoseius) agilis (Chaudhri)
Orientiseius agilis Chaudhri, 1975: 189.
Material examined
Ten females collected on cultivated apple tree, Malus domestica (Borkh.) (Rosaceae), Kukumseri, Udaipur, Lahaul and Spiti, Himachal Pradesh, India, 32°42′03.4250″N, 76°41′19.8019″E, 29 September 2024, 2753 m above sea level, coll. P. Kaur and R. Singh.
Complementary description
Female (n = 10)
(Figures 1–2)

Dorsal idiosoma — (Figure 1a). Dorsal setal pattern 12A:8A (r3 and R1 off shield). Dorsal shield mostly smooth with some patches of lateral reticulations or striations; with five pairs of solenostomes (gd2, gd4, gd6, gd8, and gd9) and 16 pairs of poroids (id1, id2, id4, id5, id6, idm1, idm2, idm3, idm4, idm5, idm6, idx, is1, idl1, idl3, and idl4). Muscle-marks (sigillae) visible mostly on podosoma, length of dorsal shield 322 (315–325), width at level of s4 162 (160–163), width at level of S2 173 (170–175). Dorsal setae smooth except Z5 slightly serrated and J5 with one barb. Measurements of dorsal setae as follows: j1 26 (25–28), j3 49 (48–49), j4 23 (20–27), j5 22 (20–25), j6 33 (30–35), J2 42 (40–43), J5 5 (4–6), z2 15 (12–18), z3 56 (50–60), z4 20 (18–23), z5 13 (12–14), Z4 64 (63–65), Z5 81 (80–82), s4 58 (57–60), s6 67 (65–70), S2 66 (65–68), S4 37 (36–38), S5 19 (18–20), r3 35 (34–37), and R1 23 (20–25). Peritremes extending almost base of setae j1; solenostome gd3 and poroid id3 visible on peritrematal shield.
Ventral idiosoma — (Figure 1b). Ventral setal pattern 15:JV:ZV. Sternal shield poorly sclerotized, smooth; with three pairs of setae (st1–st3) and two pair of poroids (iv1 and iv2); distance between st1– st3 68 (65–70), distance between setae st2 68 (65–70); metasternal setae st4 and poroids iv3 on metasternal platelets. Genital shield smooth, with one pair of setae st5; 116 (112–120) long and 66 (65–67) width at level of st5; one pair of para-genital poroids iv5 on soft cuticle. Ventrianal shield elongated, longer than wide, smooth with a slight waist at level of gv3 solenostomes and setae JV3; with four pairs of pre-anal setae (JV1, JV2, JV3, and ZV2); one pair of para-anal setae PA, unpaired post-anal seta PST, and a pair of small crescentic pre-anal solenostomes gv3 posterior to JV2, distance between solenostomes 18 (17–22). Length of ventrianal shield 113 (110–119), width at level of ZV2 78 (75–82), width at level of muscle marks (widest point) 69 (68–70), width at level of waist (narrowest point) 61 (60–62). Four pairs of caudoventral setae (ZV1, ZV3, JV4, and JV5) and four pairs of poroids (three pairs of ivo, and ivp) on soft cuticle surrounding ventrianal shield. Setae JV5 smooth, 54 (53–55) in length.
Chelicera — (Figure 1c). Fixed digit 28 (27–30) long, with three apical teeth, and pilus dentilis; movable digit 31 (30–32) long, with three teeth.
Spermatheca — (Figure 1d). Calyx saccular or tubular, flaring distally, 16 (14–18) long, 5 (4–5) wide at middle point; atrium nodular, incorporated to base of calyx; major duct broad and minor duct visible.
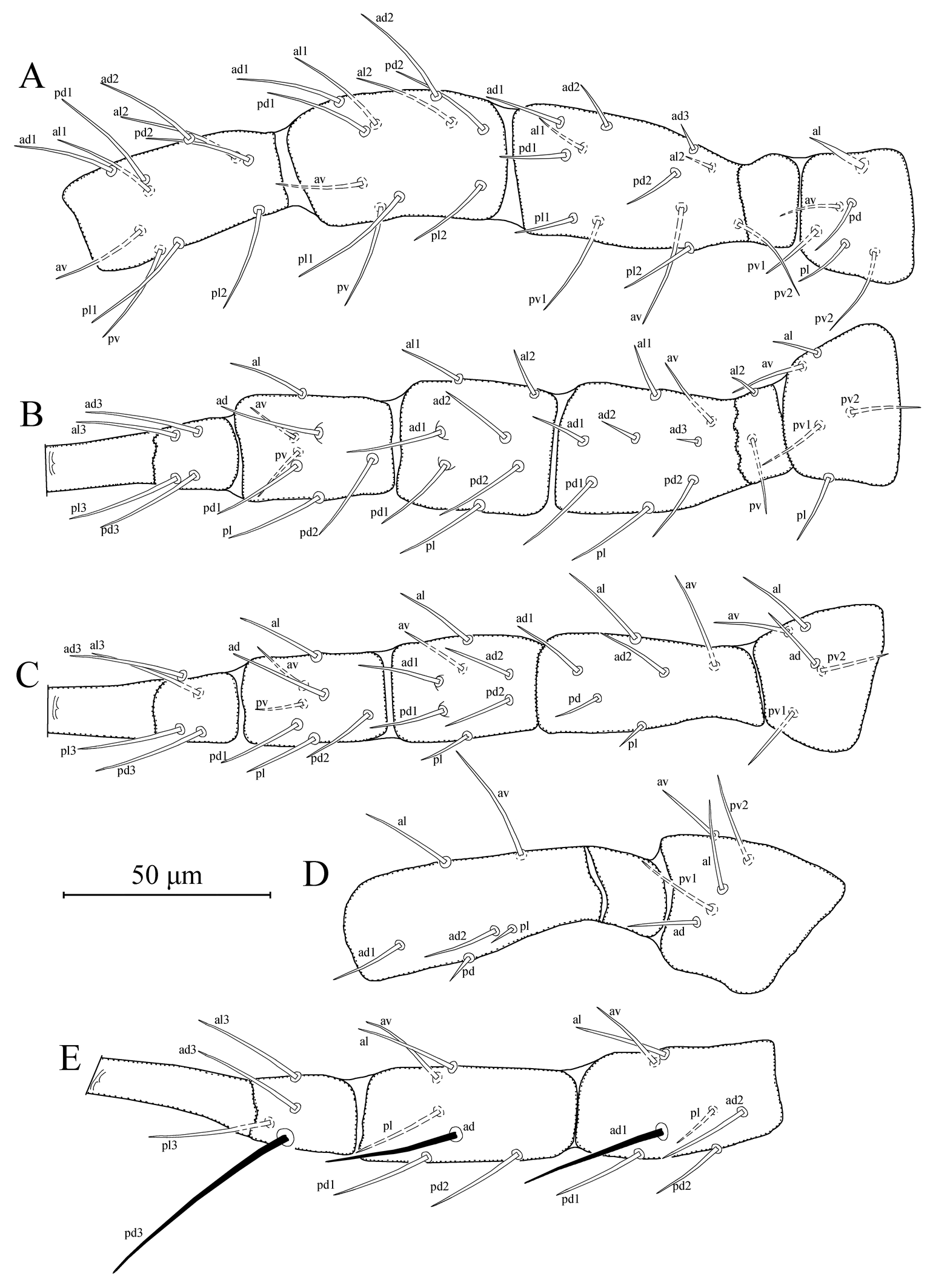
Legs — (Figures 2a–e). Length of legs I-IV 342 (335–345), 278 (275–280), 275 (270–280), and 373 (370–375), respectively. Chaetotaxy of legs as follows; leg I: coxa 0 0/1 0/1 0, trochanter 1 0/1 1/2 1, femur 2 3/1 2/2 2, genu 2 2/1 2/1 2, tibia 2 2/1 2/1 2. Leg II: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 2 3/1 2/1 1, genu 2 2/0 2/0 1, tibia 1 1/1 2/1 1. Leg III: coxa 0 0/1 0/1 0, trochanter 1 1/1 0/2 0, femur 1 2/1 1/0 1, genu 1 2/1 2/0 1, tibia 1 1/1 2/1 1. Leg IV: coxa 0 0/1 0/0 0, trochanter 1 1/1 0/2 0, femur 1 2/1 1/0 1, genu 1 2/1 2/0 1, tibia 1 1/1 2/0 1. Seta al on trochanter I subulate; three macrosetae present on genu, tibia and basitarsus of leg IV; SgeIV (ad1) 38 (36–40), StiIV (ad) 32 (31–33), and StIV (pd3) 55 (54–57) in length. Distance between pd3 and dorsal slit organ 48 (48–49).
Remarks
Typhlodromus (Anthoseius) agilis was first described by Chaudhri in 1975, based on a holotype female collected from Rubus sp. (Rosaceae) in the Northwest Frontier, Pakistan. This is the first report in India. The original description was brief, including only basic illustrations and setal measurements, and lacked several crucial diagnostic features that are now routinely used to distinguish phytoseiid species. Specifically, it did not include details such as the spermatheca morphology, number of dorsal solenostomes, leg chaetotaxy, and dentition of the cheliceral digits. The morphological characters and measurements of the current specimens are nearly identical to those provided for the holotype in the original description. However, Chaudhri (1975) reported the lengths of dorsal setae Z4 and Z5 as 87 and 57, respectively, whereas the measurements of these setae in the current specimens 64 (63–65) and 81 (80–82), respectively. In our opinion, these discrepancies likely result from a labeling error in the original description, wherein the notations of the setae may have been confused. This interpretation is supported by the original drawing, in which seta Z4 appears much shorter than Z5, consistent with the present specimens.
Moreover, the length of seta z5 was reported as 5 in the original description, compared to 13 (12–14) in the current specimens. Since this morphometric difference is the only notable discrepancy between the current specimens and the original description, we consider it a result of intraspecific variation or an error of the original description, and not sufficient to warrant the designation of a new species.
Wu and Lan (1994) and Wu et al. (2009) redescribed T. (A.) agilis from China; however, the setal measurements provided do not correspond with those in the original description or with the complementary description presented in this study. Therefore, the specimens reported as T. (A.) agilis by Wu and Lan (1994), and Wu et al. (2009) may represent another species and require reconsideration.
Typhlodromus (A.) agilis shows morphological affinity to T. (A.) hadii Chaudhri, 1965, T. (A.) lalazariensis (Chaudhri, 1975), T. (A.) malicolus Wainstein and Arutunjan, 1967, T. (A.) pruni Gupta, 1970, T. (A.) rickeri, and T. (A.) samliensis (Chaudhri, 1975), which are also placed in the rickeri species group and have nomenclatural priority over it. However, T. (A.) agilis can be distinguished from these closely related species by differences in dorsal setal measurements. In particular, the dorsocentral setae are shorter than those of T. (A.) rickeri, but longer than in the other related species mentioned above.
Typhlodromus (Anthoseius) malicolus Wainstein and Arutunjan
Typhlodromus malicolus Wainstein and Arutunjan, 1967: 1764.
Amblydromella dalii Rather, 1984: 105. New synonym.
Amblydromella viniferae Rather, 1987: 353. New synonym.
Anthoseius nobilis Kuznetsov, 1984: 389. Synonymy according to Kolodochka (2006).
Material examined
Ten females collected on wild apple tree, Malus sylvestris (L.) Mill. (Rosaceae), Kukumseri, Udaipur, Lahaul and Spiti, Himachal Pradesh, India, 32°42′03.4250″N, 76°41′19.8019″E, 29 September 2024, 2753 m above sea level, coll. P. Kaur and R. Singh.
Complementary description
Female (n = 10)
(Figures 3–4)
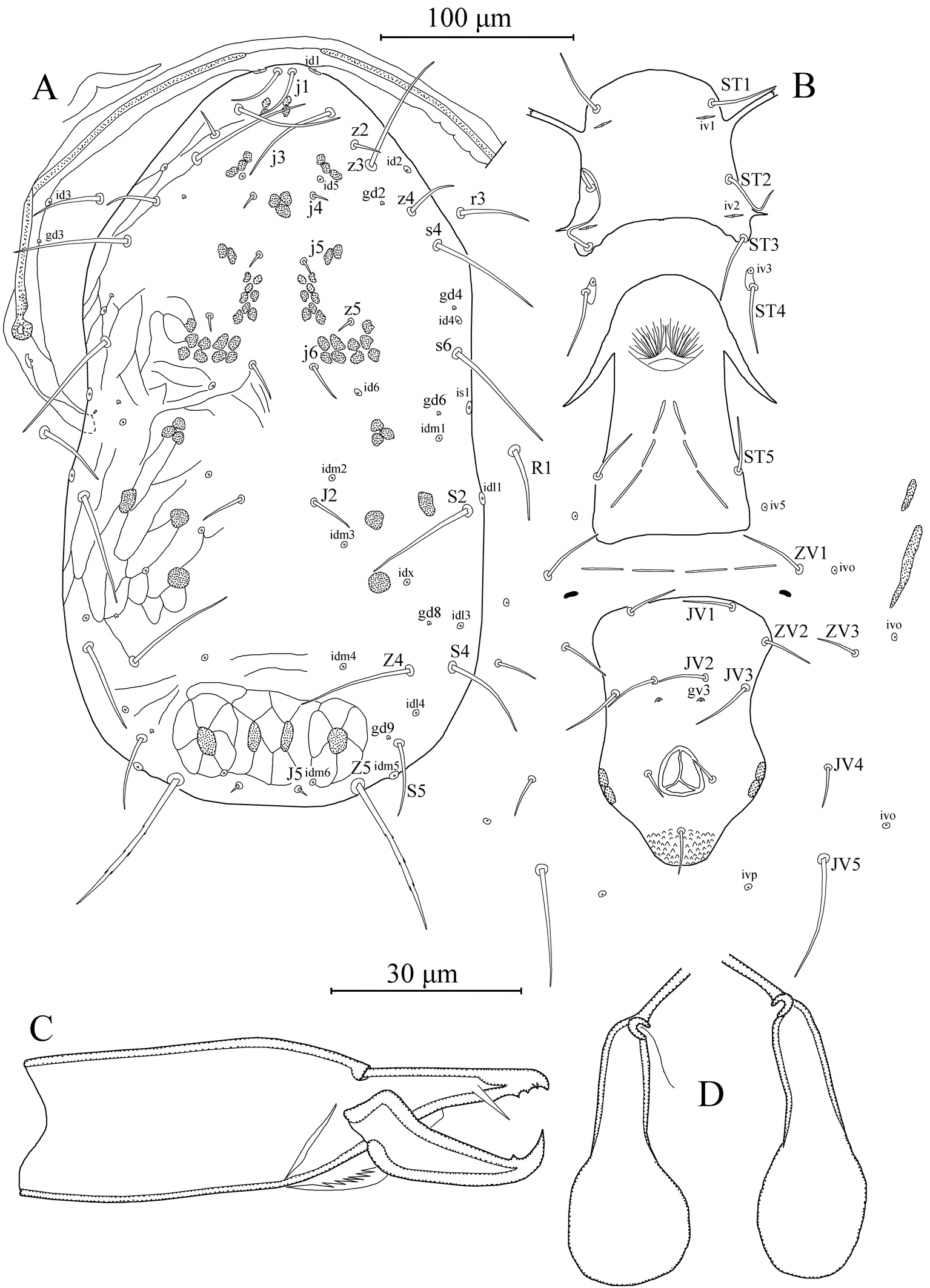
Dorsal idiosoma — (Figure 3a). Dorsal setal pattern 12A:8A (r3 and R1 off shield). Dorsal shield mostly smooth with some patches of faint reticulations or striations laterally and posteriorly; with five pairs of solenostomes (gd2, gd4, gd6, gd8, and gd9) and 16 pairs of poroids (id1, id2, id4, id5, id6, idm1, idm2, idm3, idm4, idm5, idm6, idx, is1, idl1, idl3, and idl4). Muscle-marks (sigillae) visible mostly on podosoma, length of dorsal shield 366 (360–375), width at level of s4 188 (183–190), width at level of S2 209 (203–213). Dorsal setae smooth except Z5 slightly serrated and J5 with one barb. Measurements of dorsal setae as follows: j1 27 (25–28), j3 51 (50–53), j4 8 (6–9), j5 8 (6–9), j6 20 (19–21), J2 23 (20–25), J5 5 (4–6), z2 15 (14–17), z3 59 (56–62), z4 24 (20–28), z5 8 (7–10), Z4 51 (48–55), Z5 78 (70–83), s4 57 (56–59), s6 60 (59–60), S2 58 (56–60), S4 41 (40–43), S5 33 (30–38), r3 32 (30–34), and R1 32 (30–36). Peritremes extending almost base of setae j1; solenostome gd3 and poroid id3 visible on peritrematal shield.
Ventral idiosoma — (Figure 3b). Ventral setal pattern 15:JV:ZV. Sternal shield poorly sclerotized, smooth; with three pairs of setae (st1–st3) and two pair of poroids (iv1 and iv2); distance between st1–st3 72 (71–73), distance between setae st2 72 (69–74); metasternal setae st4 and poroids iv3 on metasternal platelets. Genital shield smooth, with one pair of setae st5; 129 (124–135) long and 73 (67–77) width at level of st5; one pair of para-genital poroids iv5 on soft cuticle. Ventrianal shield elongated, longer than wide, smooth with a slight waist at level of gv3 solenostomes and setae JV3; with four pairs of pre-anal setae (JV1, JV2, JV3, and ZV2), seta ZV2 on soft cuticle in illustrated specimen; one pair of para-anal setae PA, unpaired post-anal seta PST, and a pair of small crescentic pre-anal solenostomes gv3 posterior to JV2, distance between solenostomes 21 (20–22). Length of ventrianal shield 127 (120–132), width at level of ZV2 84 (78–90), width at level of muscle marks (widest point) 75 (66–85), width at level of waist (narrowest point) 66 (62–75). Four pairs of caudoventral setae (ZV1, ZV3, JV4, and JV5) and four pairs of poroids (three pairs of ivo, and ivp) on soft cuticle surrounding ventrianal shield. Setae JV5 smooth, 57 (52–62) in length.
Chelicera — (Figure 3c). Fixed digit 31 (30–32) long, with three apical teeth, and pilus dentilis; movable digit 31 (30–32) long, with one tooth.
Spermatheca — (Figure 3d). Calyx saccular, flaring distally, 20 (18–22) long, 9 (7–10) wide at middle point; atrium nodular, incorporated to base of calyx; major duct broad and minor duct visible.

Legs — (Figures 4a–e). Length of legs I-IV 358 (350–370), 299 (290–310), 306 (295–315), and 398 (380–410), respectively. Chaetotaxy of legs same as those given for T. (A.) agilis. Seta al on trochanter I subulate; macrosetae present on genu and tibia of leg III and genu, tibia and basitarsus of leg IV; SgeIII (ad1) 30 (28–33), StiIII (ad) 28 (25–30) SgeIV (ad1) 41 (40–42), StiIV (ad) 39 (37–42), and StIV (pd3) 68 (65–69) in length. Distance between pd3 and dorsal slit organ 55 (53–55).
Remarks
Typhlodromus (Anthoseius) malicolus was first described by Wainstein and Arutunjan in 1967, based on female specimens from Malus sp. (Rosaceae) collected in Alma-Ata, Kazakhstan. The original description and the subsequent redescription of the holotype by Kolodochka (2006) are both in Russian and brief, providing only basic morphological characters, including dorsal setal measurements and the number of dorsal solenostomes. However, they lack details on leg chaetotaxy. The morphological characters and measurements of the current specimens are nearly identical to those provided for the holotype in both the original description and the redescription by Kolodochka (2006).
Rather (1984, 1987) described two species of the subgenus Anthoseius namely, T. (A.) dalii and T. (A.) viniferae. Interestingly, the female holotypes of both species, along with six paratypes of the former and two paratypes of the latter, were collected from the same host plant, Vitis vinifera L. (Vitaceae), at the same location, Dalgate, Jammu & Kashmir, India, on the same day and month, with only a two-year difference. The first species was collected in 1977, and the latter in 1979. Moreover, the original descriptions of both species are identical, including the illustrations (e.g. with all setae having the same orientation) and dorsal setal measurements, except for seta z3 (L3): 69 in T. (A.) dalii vs 59 in T. (A.) viniferae. Additionally, the morphological characters and measurements of these two species are consistent with the original description and redescription of the holotype of T. (A.) malicolus, as well as with the complementary description provided in this study. We therefore conclude that these two species are junior synonyms of T. (A.) malicolus.
This is the first report of T. (A.) malicolus under its currently accepted valid name in India. This species shows morphological affinities with T. (A.) rickeri and T. (A.) hadii. However, T. (A.) malicolus can be distinguished from T. (A.) rickeri by having shorter dorsal setae, especially dorsocentral setae. Moreover, the dorsal seta z4 is nearly twice as long in T. (A.) malicolus compared to T. (A.) hadii.
Discussion
This study improves the diagnosis of two members of the rickeri species group and contributes to taxonomic clarification within the subgenus Anthoseius. The examination of specimens identified as T. (A.) malicolus supports its current valid status and confirms the synonymy of T. (A.) dalii and T. (A.) viniferae, thereby reducing nomenclatural confusion within this species group.
Interestingly, our examination revealed that the anterolateral setae on the first trochanters of the two species included in this study are consistently modified into a subulate form, characterized by a robust base that gradually tapers to a fine point, as previously reported for several species belongs to the genera Euseius Wainstein and Neoseiulella Muma (Dayoub et al. 2025; Döker et al. 2024; Döker et al. 2025b, 2025c; Tsolakis and Ranja-William 2025; Robin et al. 2025). While the function of these modified setae remains unclear, it seems that this modification is common in various groups within the family Phytoseiidae. Further investigations should confirm whether this feature is present in other members of the rickeri species group as well as other Phytoseiidae species.
As conclusion, these findings contribute to a more stable taxonomy and provide additional morphological data that may support future systematic and comparative studies in Phytoseiidae.
Acknowledgments
We gratefully acknowledge the support of the Science and Engineering Research Board, Department of Science and Technology, Government of India, for providing funding for the field surveys conducted in the Trans Gangetic Plain and North Western Himalaya (DST project number CRG/2021/006434). We would also like to express our gratitude to the All India Network Project on Agricultural Acarology (project number ICAR-34), Department of Entomology, Punjab Agricultural University, Ludhiana, and the Department of Entomology, University of Agricultural Sciences, GKVK, Bangalore, for providing research facilities.
References
- Athias-Henriot C. 1971. Nouvelles notes sur les Amblyseiini (Gamasides podospermiques, Phytoseiidae) I. La dépilation des genoux et tibias des pattes. Acarologia, 13: 4-15.
- Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini. II. Le relevé organotaxique de la face dorsale adulte (Gamasides protoadéniques, Phytoseiidae). Acarologia, 17: 20-29.
- Chant D.A., McMurtry J.A. 1994. A review of the subfamilies Phytoseiinae and Typhlodrominae (Acari: Phytoseiidae). International Journal of Acarology, 20: 223-310. https://doi.org/10.1080/01647959408684022
- Chant D.A., McMurtry J.A. 2007. Illustrated keys and diagnoses for the genera and subgenera of the Phytoseiidae of the world (Acari: Mesostigmata). West Bloomfield, Indira Publishing House, 219 pp.
- Chant D.A., Yoshida-Shaul E. 1991. Adult ventral setal patterns in the family Phytoseiidae (Acari: Gamasina). International Journal of Acarology, 17: 187-199.c https://doi.org/10.1080/01647959108683906
- Chaudhri W.M. 1965. Two new species of mites from Pakistan (Acarina: Phytoseiidae). Acarologia, 7: 632-635.
- Chaudhri W.M. 1975. Descriptions of the mites of the genus Amblydromella (Acarina: Phytoseiidae) from Pakistan. Pakistan Journal of Agricultural Science, 12: 201-220.
- Dayoub A.M., Döker I., Ueckermann E.A. 2025. Three new records of Phytoseiidae (Acari: Mesostigmata) in Syria, with a key to Syrian species. Systematic and Applied Acarology, 30: 592-610. https://doi.org/10.11158/saa.30.3.7
- Demite P.R., de Moraes G.J., McMurtry J.A., Denmark H.A., Castilho R.C. 2026. Phytoseiidae database. [accessed 2026 Febuary 27]. https://www.lea.esalq.usp.br/phytoseiidae/
- Döker I., Atchia I., Bolton, S.J. 2025a. Complementary descriptions of three nominal phytoseiid mite species (Parasitiformes: Phytoseiidae) invalid under the International Code of Zoological Nomenclature. Systematic and Applied Acarology, 30: 1007-1016.c https://doi.org/10.11158/saa.30.6.4
- Döker I., Çelik S.O., Karaca, M.M. 2025c. Review of Euseius Wainstein (Parasitiformes: Phytoseiidae) in Türkiye: An integrative taxonomic approach using morphological and molecular data with a new combination Euseius degenerans (Berlese). Systematic and Applied Acarology, 30: 560-591. https://doi.org/10.11158/saa.30.3.6
- Döker I., Khaustov V.A., Joharchi O., Khaustov A., Kazakov D.V., Meshkov Y.I. 2024. Integrative taxonomy demonstrates synonymy between Euseius amissibilis Meshkov and Euseius gallicus Kreiter & Tixier (Acari: Phytoseiidae). Systematic and Applied Acarology, 29: 60-77. https://doi.org/10.11158/saa.29.1.5
- Döker I., Robin, Jose A., Kaur P., Gowda C.C., Brar-Bhullar M. 2025b. Some species of Euseius Wainstein (Parasitiformes: Phytoseiidae) from India: description of a new species, complementary description of known species with new synonyms. Systematic and Applied Acarology, 30: 1297-1324. https://doi.org/10.11158/saa.30.7.9
- Evans G.O. 1963. Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bulletin of the British Museum (Natural History) Zoology, 10: 275-303. https://doi.org/10.5962/bhl.part.20528
- Ferragut F., Navia D. 2024. The subfamily Typhlodrominae Wainstein (Mesostigmata: Phytoseiidae) in the Serra do Espinhaço, Brazil, with the description of a new genus and two new species. International Journal of Acarology, 50: 87-108.c https://doi.org/10.1080/01647954.2023.2290665
- Gerson U. 2014. Pest control by mites (Acari): present and future. Acarologia, 54: 371-394. https://doi.org/10.1051/acarologia/20142144
- Gupta S.K. 1970. Some predatory mites of the family Phytoseiidae from West Bengal, India (Acarina: Mesostigmata). Oriental Insects, 4: 185-191. https://doi.org/10.1080/00305316.1970.10433954
- Hernandes F.A., Kreiter S., Tixier M.-S. 2012. The first electronic polytomous key to the world species of the subgenus Typhlodromus (Anthoseius) De Leon (Acari: Phytoseiidae). Zootaxa, 3451: 46-59. https://doi.org/10.11646/zootaxa.3451.1.4
- Kolodochka L.A. 2006. Phytoseiid mites of the Palaearctic Region (Parasitiformes, Phytoseiidae): faunistic, taxonomy, ecomorphology, evolution. Vestnik Zoologii, suppl. 21, 250pp. [in Russian].
- Kuznetsov N.N. 1984. New species of predacious mites of the family Phytoseiidae (Parasitiformes) in the USSR. Entomologicheskoe Obozrenie, 63: 384-397 [in Russian].
- Kreiter S., Payet R.-M., Azali H.A. 2021. Phytoseiid mites (Acari: Mesostigmata) of Anjouan Island (Comoros Archipelago). Acarologia, 61: 62-83.c https://doi.org/10.24349/acarologia/20214418
- Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina Acarina: Mesostigmata. The Memoirs of the Entomological Society of Canada, 47: 1-64. https://doi.org/10.4039/entm9747fv
- McMurtry J.A., Moraes G.J. de, Sourassou N.F. 2013. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Systematic and Applied Acarology, 18: 297-320. https://doi.org/10.11158/saa.18.4.1
- Muma M.H., Denmark H.A. 1968. Some generic descriptions and name changes in the family Phytoseiidae (Acarina: Mesostigmata). The Florida Entomologist, 51, 229-240. https://doi.org/10.2307/3493424
- Rather A.Q. 1984. New species and a new record of the genus Amblydromella Muma (Acarina, Phytoseiidae) from India. Entomologist's Monthly Magazine, 120: 103-108.
- Rather A.Q. 1987. Some new records and a new species of phytoseiid mites (Acarina: Mesostigmata) from Jammu and Kashmir (India). Zeitschrift fur Angewandte Zoologie, 74, 353-359.
- Robin, Döker I., Kaur P., Brar-Bhullar M., Gowda C.C. 2025a. A new species of Neoseiulella Muma (Parasitiformes: Phytoseiidae) from Northwestern India. Acarologia, 65, 762-770. https://doi.org/10.24349/wpop-gesj
- Rowell H.J., Chant D.A., Hansell R.I.C. 1978. The determination of setal homologies and setal patterns on the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata). The Canadian Entomologist, 110: 859-876.c https://doi.org/10.4039/Ent110859-8
- Tsolakis H., Ranja-William F.A. 2025. New records of phytoseiid mites from Madagascar, with descriptions of two new species. Acarologia 65: 448-460.c https://doi.org/10.24349/t5zy-f3cc
- Tsolakis H., Ragusa S. 2016. On the identity of Neoseiulus fallacis (Garman 1948) (Parasitiformes, Phytoseiidae) redescription of the species and description of the new species Neoseiulus garmani. International Journal of Acarology, 42: 394-404.c https://doi.org/10.1080/01647954.2016.1205134
- Wainstein B.A., Arutunjan E.S. 1967. New species of predaceous mites of the genera Typhlodromus Scheuten and Paraseiulus Muma (Parasitiformes, Phytoseiidae). Zoologicheskii Zhurnal, 46: 1764-1770 [in Russian].
- Wu W.N., Lan W.M. 1994. Notes on the agilis group of the genus Typhlodromus from China (Acari: Phytoseiidae). Acta Zootaxonomica Sinica, 19: 426-432 [in Chinese].
- Wu W.N., Ou J.F., Huang J.L. 2009. Fauna Sinica, Invertebrata vol. 47. Arachnida Acari: Phytoseiidae. Beijing, Science Press, 511 pp. [In Chinese with English abstract].



2026-03-05
Date accepted:
2026-04-18
Date published:
2026-05-05
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Döker, Ismail; Singh, Robin; Kaur, Paramjit and Chinnamade Gowda, Channegowda
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)