



A redefinition of Vikramus Denmark & Evans (Parasitiformes: Mesostigmata) with a complementary description of Vikramus argentinus (Guanilo & Moraes) comb. nov.
Döker, Ismail  1
; Demite, Peterson R.
2
; Atchia, Isabelle
3
; Gasparro-Brewer, Jude
4
and Bolton, Samuel J.
5
1
; Demite, Peterson R.
2
; Atchia, Isabelle
3
; Gasparro-Brewer, Jude
4
and Bolton, Samuel J.
5
1✉ Cukurova University, Agricultural Faculty, Department of Plant Protection, Acarology Laboratory, 01330, Adana, Türkiye.
2Campus Bonfim, Instituto Federal de Roraima (IFRR), Bonfim, Roraima, Brazil.
3Florida Department of Agriculture and Consumer Services, Florida State Collection of Arthropods (FDACS) 32608, Gainesville, FL, USA.
4Florida Department of Agriculture and Consumer Services, Florida State Collection of Arthropods (FDACS) 32608, Gainesville, FL, USA.
5Florida Department of Agriculture and Consumer Services, Florida State Collection of Arthropods (FDACS) 32608, Gainesville, FL, USA.
2026 - Volume: 66 Issue: 2 pages: 307-316
https://doi.org/10.24349/p1bw-itsdZooBank LSID: 4026BA84-18D0-4D0B-9007-D540593D24FD
Original research
Keywords
Abstract
Introduction
The family Phytoseiidae (Mesostigmata) includes numerous species of predatory mites that are widely studied because of their ecological importance and their use in biological control (McMurtry et al. 2013; Gerson 2014; Knapp et al. 2018; Lofego et al. 2024; Demite et al. 2026). As in many mite families, the taxonomy of Phytoseiidae relies heavily on morphological characters; however, generic boundaries within the family remain taxonomically challenging, particularly in groups traditionally delimited based on a restricted set of morphological characters.
The genus Vikramus Denmark & Evans, 2019 was established relatively recently as a monotypic genus, based solely on its type species, Vikramus cordobus. Although the genus was separated from Neoseiulus Hughes, 1948 primarily on the basis of the morphology of the dorsal setae, its original description was brief and did not provide a sufficiently detailed morphological characterization to clearly justify its generic status. As a result, the placement and taxonomic status of Vikramus have remained uncertain, especially in relation to the morphologically similar species Neoseiulus argentinus Guanilo & Moraes, 2008, currently assigned to the cucumeris species group and tunus species subgroup within Neoseiulus.
At the same time, Neoseiulus sensu Chant & McMurtry, 2003 is widely recognized as polyphyletic, and several of its species groups and species subgroups include taxa that differ markedly in overall morphology (Zannou et al. 2006; Tsolakis et al. 2012; Tsolakis and Ragusa 2016; Döker et al. 2023a; Ferragut and Navia 2022; Rahmani-Piyani et al. 2025). This taxonomic instability suggests the need for a re-evaluation of recently proposed genera that were separated from Neoseiulus without comprehensive comparative analyses. In such cases, re-examination of type material is essential to assess whether proposed genera represent distinct morphological lineages or are artificial segregates based on incomplete character sets.
In the present study, we re-examine the holotype female of V. cordobus and detailed photos of the type series of N. argentinus. Given that these two species show a high degree of morphological similarity, this examination was undertaken to clarify their taxonomic relationship and to determine whether they represent distinct species or the same taxon. In addition, we provide a redefinition of the genus Vikramus based on a consistent combination of diagnostic characters. Finally, we discuss the taxonomic status of Vikramus and justify its recognition as a separate valid genus. This study contributes to a clearer understanding of generic boundaries within Phytoseiidae and provides a reference for future taxonomic studies of related taxa.
Materials and methods
Mite specimen was examined using the following: 1) a Zeiss Axio Imager M2 compound microscope fitted with differential interference contrast optics and an AxioCam 820 mono camera; 2) a Zeiss Axioscope 5 compound microscope equipped with phase-contrast optics and a Canon EOS 90D camera. Specimen was imaged with oil lenses (63x or 100x; 1.4 NA) and separate images were stitched together with PTGui (Version 13.0) to produce large panoramas. Z-stacks were merged using Helicon Focus version 7.6.4 Pro (Helicon Soft Ltd., 2000). Illustrations were produced digitally with Adobe Photoshop CS2. Measurements of the dorsal shield were taken along the midline from the anterior margin to the posterior margin, whereas the length of the ventrianal shield, including the cribrum, was measured along the midline. Leg lengths were measured from the base of the coxa to the distal end of the tarsus, excluding the ambulacrum. All measurements were obtained using ZEN 2012 software (version 8.0) and are presented in micrometers. Terminology and nomenclature of dorsal setae follow Lindquist and Evans (1965), as adapted for Phytoseiidae by Rowell et al. (1978). Ventral setal nomenclature follows Chant and Yoshida-Shaul (1991), while the nomenclature of dorsal solenostomes (gland pores) follows Athias-Henriot (1975). Leg chaetotaxy is based on Evans (1963).
Results
Vikramus Denmark & Evans
Vikramus Denmark & Evans, 2019: 98.
Type species — Vikramus cordobus Denmark & Evans, 2019: 99, junior subjective synonym. (= Neoseiulus argentinus Guanilo & Moraes, 2008 in Guanilo et al. 2008: 17.).
Valid name of type species — Vikramus argentinus (Guanilo & Moraes, 2008) comb. nov.
Diagnosis
This genus characterized by combination of following characters: shape of ventrianal shield, morphology and position of preanal solenostomes, morphology of spermatheca, and morphology of dorsal setae. Ventrianal shield narrow, elongated, much longer than wide, width at level of seta ZV2 subequal to width at level of anal opening; with a slight waist at level of JV2; lateral margins progressively narrowing from level of para-anal setae or muscle marks towards cribrum. Solenostomes gv3 small, rounded, located well posteromedian to seta JV2, closer to JV2 than anal opening; relatively well-separated and closer to base of setae JV2. Spermatheca with calyx simple, cup-shaped. Atrium small, nodular, attached to calyx, with a very small neck, never forked at juncture with major duct. Dorsal setae and ventral seta JV5 clubbed, strongly serrated, except j4, j5, j6, J2, and z5 smooth and sharply pointed. These characters represent unique set of morphological features defining this genus. No exceptions to this character combination known.
Other morphological character states
Dorsal idiosoma: Dorsal shield sclerotized, strongly reticulated, oval, with slight waist at level of Z1; setal pattern 10A:9B (r3 and R1 off shield). Dorsal setae clubbed and strongly serrated, except j4, j5, j6, J2, and z5 smooth and sharply pointed. Dorsal solenostomes simple, rounded; complete set of solenostomes present (seven pairs).
Ventral idiosoma: Sternal shield smooth, except some lateral striations, with three pairs of setae (ST1–ST3); length/width ratio subequal; posterior margin straight. Setae ST4 on metasternal platelets. Genital shield smooth, as wide as ventrianal shield. Ventrianal shield reticulated and elongated, much longer than wide; its shape is one of the primary morphological features distinguishing this genus, as described above; with three pairs of preanal setae (JV1, JV2, and ZV2) arranged in broad triangular pattern. Four pairs of caudoventral setae (JV4, JV5, ZV1, and ZV3) on soft cuticle surrounding ventrianal shield; two pairs of metapodal plates present. Ventral setae simple, smooth except JV5 slightly clubbed and serrated in apical half.
Peritrematal shield: Peritrematal shield fused with dorsal shield. Peritremes stippled type; long, extending beyond insertion of seta j1. Solenostome gd3 and poroid id3 present on peritrematal shield.
Spermatheca: Calyx of spermatheca cup-shaped with well sclerotized wall; atrium nodular attached to calyx with a very small neck, not always visible, never forked at juncture with a thin major duct. The spermatheca is one of the primary morphological features recognizing this genus.
Chelicera: Fixed digit with five teeth, four of them located subapically and one slightly more prominent than others, near base of pilus dentilis; movable digit with only one tooth. Second segment of chelicera of normal proportions; length ratio of second cheliceral segment to movable digit = 2.50–2.70: 1.0.
Legs: Legs without macrosetae. Leg chaetotaxy as follows: leg I: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 2 3/1 2/2 2, genu 2 2/1 2/1 2, tibia 2 2/1 2/1 2. Leg II: coxa 0 0/1 0/1 0, trochanter 1 0/1 0/2 1, femur 2 3/1 2/1 1, genu 2 2/1 2/0 1, tibia 1 1/1 2/1 1. Leg III: coxa 0 0/1 0/1 0, trochanter 1 1/1 0/1 1, femur 1 2/1 1/0 1, genu 1 2/1 2/0 1, tibia 1 1/1 2/1 1. Leg IV: coxa 0 0/1 0/0 0, trochanter 1 1/1 0/1 1, femur 1 2/1 1/0 1, genu 1 2/1 2/0 1, tibia 1 1/1 2/0 1. Leg chaetotaxy stable at generic level; interspecific variation, if present in future-described species, expected to be restricted to genu II.
Vikramus argentinus (Guanilo & Moraes) comb. nov. (Figures 1–2)
Neoseiulus argentinus Guanilo & Moraes, in Guanilo et al. 2008: 17.
Vikramus cordobus Denmark & Evans 2019: 99. syn. nov.
Material examined
Holotype female, Vikramus cordobus, on Solanum elaeagnifolium Cavanilles, (Solanaceae), January 1980, Cordoba Province, Argentina, coll. C.C. Orr. Photos of the holotype and three female paratypes of N. argentinus, on S. americanum Miller (Solanaceae), 5 May 2007, Chajarí, Taue, Entre Ríos Province, Argentina, coll. A. D. Guanilo.
Diagnosis
As in genus diagnosis.
Redescription
Female (n = 1, holotype female of V. cordobus). (Figures 1–2)
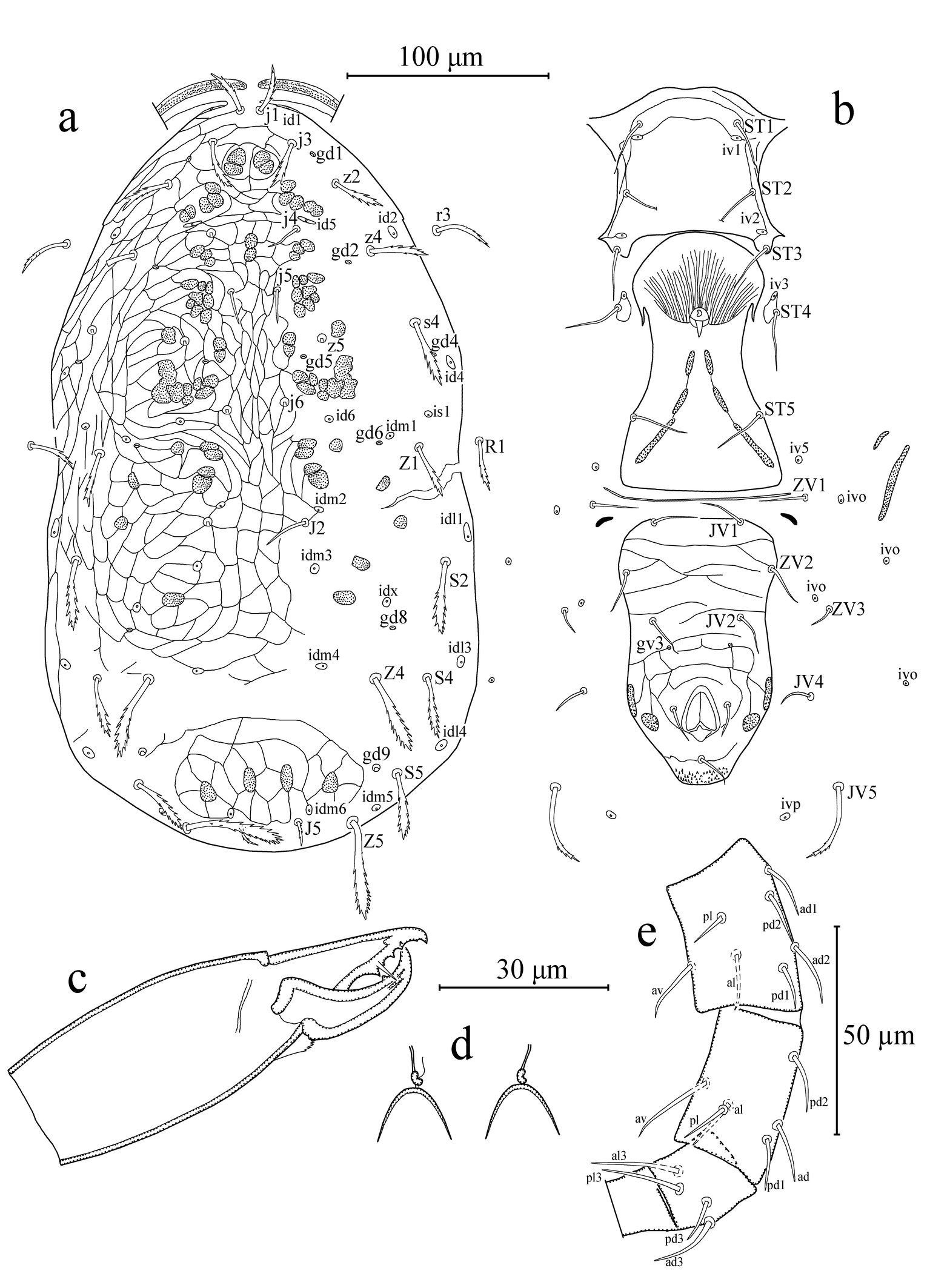
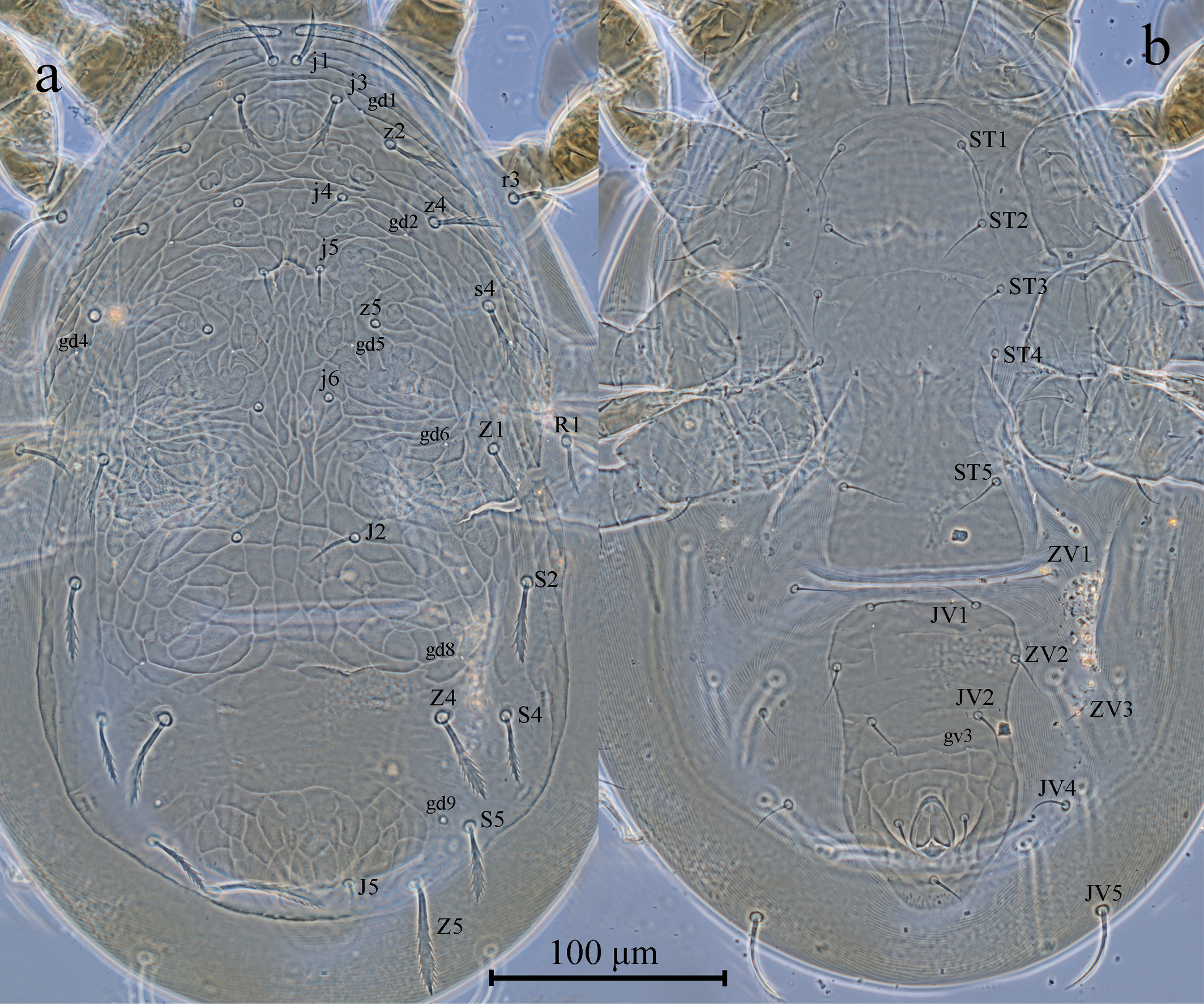
Dorsal idiosoma (Figures 1a, 2a). Dorsal setal pattern 10A: 9B (r3 and R1 off shield). Dorsal shield sclerotized, strongly reticulated, oval with slight waist at level of Z1; with seven pairs of solenostomes (gd1, gd2, gd4, gd5, gd6, gd8, and gd9); 16 pairs of visible poroids (sensillae) (id1, id2, id4, id5, id6, idm1, idm2, idm3, idm4, idm5, idm6, idx, is1, idl1, idl3, and idl4). Muscle marks (sigilla) mostly visible on podosoma. Length of dorsal shield 360, width at level of s4 196, width at level of S2 210. Dorsal setae clubbed, strongly serrated, except j4, j5, and probably j6 and z5 simple, smooth. Measurements of dorsal setae as follows: j1 24, j3 25, j4 17, j5 17, j6 (broken), J2 (broken), J5 12, z2 27, z4 30, z5 (broken), Z1 27, Z4 41, Z5 49, s4 33, S2 35, S4 30, S5 32, r3 28, and R1 25.
Peritrematal shield. Fused with dorsal shield at level of seta j1. Peritremes stippled type with three to four lines of microvilli, extending between setae j1. Solenostomes gd3, and poroid id3 in association with gd3 visible. Solenostome gdp inserted posterior to stigmatic opening.
Ventral idiosoma (Figures 1b, 2b). Ventral setal pattern 14:JV–3:ZV. Sternal shield sclerotized, mostly smooth, except some lateral striations, with three pairs of setae (ST1–ST3) and with two pairs of poroids iv1 and iv2; distance between ST1–ST3 63, distance between setae ST2 63; metasternal setae ST4 and poroid iv3 on metasternal platelets. Genital shield smooth, with one pair of setae ST5; 124 long, and 66 width at level of ST5; one pair of para-genital poroids iv5 on soft cuticle. Ventrianal shield elongated, much longer than wide with a slight waist, reticulated; with three pairs of preanal setae (JV1, JV2 and ZV2), one pair of para-anal setae (PA), and unpaired post-anal seta (PST); one pair of small, rounded preanal solenostomes (gv3) located posteromedian to JV2, distance between solenostomes 31. Length of ventrianal shield 128, width at level of ZV2 78, width at level of para-anal setae 70. Four pairs of caudoventral setae (JV4, JV5, ZV1, and ZV3); five pairs of poroids (four pairs of ivo, and ivp); two pairs of metapodal plates on soft cuticle surrounding ventrianal shield. Primary metapodal plate 39 long, 3 wide, secondary metapodal plate 12 long, 2 wide. Setae JV5 slightly clubbed, serrated in apical half, 39 in length.
Gnathosoma (Figure 1c). Subcapitulum slightly sclerotized, with three pairs of hypostomal setae h1 22, h2 22, and h3 21 in length, and with one pair of palp coxal seta pc 24 in length; internal malae paired, not extending tip of corniculi; deutosternal groove narrow 3–4 in width, with seven transverse rows of denticles, each row with two lateral denticles. Second segment of chelicera 75 in length. Antiaxial poroid and external arthrodial brush visible. Fixed digit 28 long, with five teeth, and pilus dentilis; movable digit 27 long, with one tooth.
Spermatheca (Figure 1d). Calyx bell-shaped, 10 long, 12 wide at widest point (apical point); atrium knobbed, attached to calyx with a very small neck, absent in one side, probably due to its position or mounting; major duct narrow, minor duct visible.
Legs (Figure 1e). Length of legs I-IV 287, 220, 206, and 300, respectively. Chaetotaxy of legs as in genus diagnosis. Legs without macrosetae.
Remarks
Vikramus argentinus comb. nov. was described by Guanilo and Moraes (in Guanilo et al. 2008) from specimens collected from Solanum americanum Mill. (Solanaceae) in Chajarí, Entre Ríos Province, Argentina. The original description is detailed and includes most of the diagnostic characters. Vikramus cordobus was later described by Denmark and Evans (2019) based on a single female holotype collected from S. elaeagnifolium Cavanilles (Solanaceae) in Córdoba Province, Argentina. We examined the holotype of V. cordobus, deposited in the mite collection of the Florida State Collection of Arthropods (FSCA), Gainesville, FL, USA. In addition, we also examined photos of the holotype and three female paratypes of V. argentinus comb. nov.
Several inconsistencies were found between our examination and the original descriptions of V. argentinus comb. nov. and V. cordobus. In their original descriptions, nine setae were reported on genu II for V. argentinus comb. nov. and seven for V. cordobus, whereas we consistently observed eight setae on both genu II of the holotype of V. cordobus. Additionally, the original description of V. argentinus comb. nov. indicated two teeth on the movable digit of the chelicera, and V. cordobus was described as having no tooth, while our examination of the holotype of V. cordobus revealed a single tooth. Furthermore, the number of dorsal solenostomes differed considerably: six pairs (gd5 absent) were reported for V. argentinus comb. nov. and five pairs (gd5 and gd6 absent) for V. cordobus, whereas we observed seven pairs of solenostomes on the holotype of V. cordobus. Because the number of setae on genu II, dentition of the movable digit, and the number of dorsal solenostomes are important and mostly stable diagnostic characters for phytoseiids (Athias-Henriot 1977, 1978; Döker et al. 2018; Kolodochka 2006; Khaustov et al. 2022; Papadoulis et al. 2009), we contacted Dr. Gilberto José de Moraes to re-examine the type specimens of V. argentinus comb. nov. deposited at ESALQ-USP, Piracicaba, São Paulo, Brazil. His re-examination revealed the following: in six of the seven available specimens, only one tooth is present on the movable digit, although one specimen appears to show a second basal tooth, which may represent an artifact. Regarding genu II, six specimens possess eight setae, whereas a single specimen shows nine setae due to the apparent presence of an enlarged pv seta on one side. Although possibility of nine setae on genu II mentioned in one specimen, our careful examination of the video recordings showed that there are eight setae, which is also consistent with our observations on the V. cordobus holotype. We also confirmed the presence of seven pairs of solenostomes, including gd5, on the dorsal shield, consistent with our observations of the V. cordobus holotype.
Based on these observations and complete similarity between the two species, we conclude that V. cordobus, is a junior subjective synonym of V. argentinus comb. nov.
Taxonomic status of the genus Vikramus and justification for its recognition as a separate valid genus
The genus Vikramus was established by Denmark and Evans (2019) based primarily on the dorsal setal pattern and its morphology, the number of setae on the ventrianal shield, and chelicera dentition. It is currently represented only by its type species, V. cordobus a junior subjective synonymy of V. argentinus comb. nov. The latter valid species was originally placed in the cucumeris species group and tunus species subgroup of Neoseiulus, most probably on the basis of the dorsal setal pattern (10A:9B) and the presence of strongly serrated dorsal setae. Despite the synonymy of its type species, Vikramus is retained here as a valid genus. This decision is supported by the distinctly modified dorsal setal morphology of V. argentinus comb. nov. and by increasing evidence that Neoseiulus Hughes sensu Chant and McMurtry (2003) represents a polyphyletic genus (Zannou et al. 2006; Tsolakis et al. 2012; Tsolakis and Ragusa 2016; Döker et al. 2023a; Ferragut and Navia 2022; Rahmani-Piyani et al. 2025). Species assigned to the tunus species subgroup share some morphological characters with V. argentinus comb. nov., such as the dorsal setal pattern and modified dorsal setae, but differ from most other species traditionally assigned to Neoseiulus (Cavalcante et al. 2017).
Molecular evidence further supports the distinct position of the tunus species subgroup. Based on ITS and 12S rRNA gene markers, Tsolakis et al. (2012) showed that N. tunus (De Leon, 1967) clusters with Kampimodromus aberrans (Oudemans, 1930) and K. corylosus Kolodochka, 2003 in both parsimony and Bayesian analyses. These latter species are currently placed in the subtribe Kampimodromina Chant and McMurtry, 2003 in the tribe Kampimodromini Kolodochka, 1998, which are characterized by the absence of dorsal seta S4. Although some morphological similarities, such as serrated dorsal setae and an elongated ventrianal shield, exist between Kampimodromus Nesbitt, 1951 and members of the tunus species subgroup, they are currently placed in two different tribes. This suggests that phylogenetic relationships inferred from molecular data cannot be reliably established solely on the presence or absence of a single dorsal seta. These observations indicate limitations of the classification system proposed by Chant and McMurtry (2007), which relies heavily on dorsal setal patterns and, even in some cases, ratios between certain dorsal setae for separating tribes. Several examples illustrate this problem. For instance, Typhlodromus (Anthoseius) cephalochaitosus Moraes et al. 2001, described from Kenya, lacks dorsal seta z3, yet was placed in Typhlodromus Scheuten a genus otherwise characterized by the consistent presence of this seta across more than 400 nominal species. Similarly, some species currently assigned to Neoseiulella Muma based on their dorsal setal pattern show strong overall morphological affinity with species of Typhlodromus (Anthoseius), particularly in dorsal and ventral shields morphology, the presence of knobbed dorsal setae and ventral seta JV5, knobbed macrosetae on legs, ventrianal shield shape, morphology and position of preanal pores, and cheliceral dentition (Chant and Yoshida-Shaul 1989; Kanouh et al. 2012; Tsolakis and Ragusa 2015; Döker et al. 2023b; Robin et al. 2025). In addition, recent discussions on the validity of Moraeseius Chant and McMurtry further emphasize the risks of defining genera based solely on the absence or presence of a single dorsal seta (Döker et al. 2025). Together, these cases demonstrate that phylogenetically coherent, monophyletic taxa should not be defined using characters that require multiple exceptions. Consequently, when defining genera, and especially higher taxa such as tribes or subfamilies, reliance on single characters should be avoided. Many of these characters are highly prone to homoplasy, often resulting from convergent or parallel evolution, and the sole use of a single character for defining higher taxa may lead to artificial and non-monophyletic groupings. As currently applied, the system of Chant and McMurtry (2007) often creates practical difficulties in recognizing genera within Amblyseiinae and even across the family Phytoseiidae. Genera such as Transeius sensu Chant and McMurtry (2004), Typhlodromips sensu Chant and McMurtry (2005), and Neoseiulus sensu Chant and McMurtry (2003) are increasingly recognized as polyphyletic and will likely require further subdivision to achieve a more realistic and evolutionarily consistent classification (Çobanoğlu et al. 2018; Khaustov et al. 2020, Döker et al. 2022).
In this context, Vikramus is provisionally retained within the tribe Neoseiulini, although it shows closer morphological affinities to some members of Kampimodromini, particularly in the elongated ventrianal shield and modified dorsal setae. We refrain from transferring Vikramus to Kampimodromini at this stage, as the tribe itself also requires substantial revision.
Importantly, V. argentinus comb. nov. differs clearly from species of the tunus species subgroup by its anteriorly narrower ventrianal shield, clubbed dorsal setae, and simple spermatheca. In contrast, N. tunus typically exhibits a broader ventrianal shield, large crescentic preanal solenostomes located close together in line with setae JV2, and a forked atrium at the junction with the major duct (based on our examination of numerous specimens collected from Brazil). Interestingly, V. argentinus comb. nov. shares several characters, including an elongated ventrianal shield and clubbed dorsal setae, with species currently assigned to Asperoseius Chant, 1959, suggesting a possible relationship among these taxonomic groups. Future molecular and integrative studies will be essential to clarify the phylogenetic relationships among V. argentinus comb. nov., species of the tunus species subgroup, and members of Kampimodromini, and to establish a more evolutionarily consistent classification within Phytoseiidae.
Acknowledgements
We are grateful to Prof. Dr. Gilberto José de Moraes and Mr. José Dantas Araújo Lacerda (Universidade de São Paulo, Piracicaba, Brazil) for sharing related photos and video records of the type specimens of Neoseiulus argentinus.
References
- Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini. II. Le relevé organotaxique de la face dorsale adulte (Gamasides, protoadéniques, Phytoseiidae). Acarologia, 17: 20-29.
- Athias-Henriot C. 1977. Nouvelles notes sur les Amblyseiini. III. Sur le genre Cydnodromus: Redéfinition, composition (Parasitiformes, Phytoseiidae). Entomophaga, 22: 61-73. https://doi.org/10.1007/BF02372991
- Athias-Henriot, C. 1978. Définition de Dictyonotus nov. gen., avec description de deux espèces nouvelles de France Méridionale (Gamasides, Phytoseiidae). Entomophaga, 23: 189-194. https://doi.org/10.1007/BF02371727
- Cavalcante A.C.C., Demite P.R., Amaral F.S.R., Lofego A.C., Moraes G.J. de 2017. Complementary description of Neoseiulus tunus (De Leon) (Acari: Mesostigmata: Phytoseiidae) and observation on its reproductive strategy. Acarologia, 57: 591-599. https://doi.org/10.24349/acarologia/20174178
- Chant D.A. 1959. Phytoseiid mites (Acarina: Phytoseiidae). Part I. Bionomics of seven species in southeastern England. Part II. A taxonomic review of the family Phytoseiidae, with descriptions of thirty-eight new species. Can. Entomol., 61: 1-166. https://doi.org/10.4039/entm9112fv
- Chant D.A., McMurtry J.A. 2003. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae): Part I. Neoseiulini new tribe. Int. J. Acarol., 29: 3-46. https://doi.org/10.1080/01647950308684319
- Chant D.A., McMurtry J.A. 2004. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae): Part III. The tribe Amblyseiini Wainstein, subtribe Amblyseiina n. subtribe. Int. J. Acarol., 30: 171-228. https://doi.org/10.1080/01647950408684388
- Chant D.A., McMurtry, J.A. 2005. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae) Part VII. Typhlodromipsini n. tribe. Int. J. Acarol., 31: 315-340. https://doi.org/10.1080/01647950508683673
- Chant D.A., McMurtry J.A. 2007. Illustrated keys and diagnoses for the genera and subgenera of the Phytoseiidae of the world (Acari: Mesostigmata). West Bloomfield, Indira Publishing House, 219 pp.
- Chant D.A., Yoshida-Shaul E. 1989. A world review of the tiliarum species group in the genus Typhlodromus Scheuten (Acari: Phytoseiidae). Can. J. Zool., 67: 1006-1046. https://doi.org/10.1139/z89-144
- Chant D.A., Yoshida-Shaul E. 1991. Adult ventral setal patterns in the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol., 17: 187-199. https://doi.org/10.1080/01647959108683906
- Çobanoğlu S., Faraji F., Cılbırcıoğlu C. 2018. Re-descriptions of Amblyseius decolor (Westerboer) and Proprioseiopsis sororculus (Wainstein) (Acari: Phytoseiidae) based on the specimens collected in Turkey. Acarologia, 58: 825-836. https://doi.org/10.24349/acarologia/20184272
- De Leon D. 1967. Some mites of the Caribbean Area. Part I. Acarina on plants in Trinidad, West Indies. Allen Press Inc., Lawrence, Kansas, USA, pp. 1-66.
- Demite P.R., Moraes G.J. de, McMurtry J.A., Denmark H.A., Castilho R.C. 2026. Phytoseiidae database. [accessed 2026 January 28]. https://www.lea.esalq.usp.br/phytoseiidae/
- Denmark H.A., Evans G.A. 2019. Additions to the world fauna of the family Phytoseiidae (Acari: Mesostigmata) with an illustrated key to the subfamilies, tribes, subtribes and genera of Phytoseiidae of the world. West Bloomfield, Indira Publishing House, 315 pp.
- Döker I., Celik S.O., Karaca M.M. 2025. Review of Euseius Wainstein (Parasitiformes: Phytoseiidae) in Türkiye: An integrative taxonomic approach using morphological and molecular data with a new combination Euseius degenerans (Berlese). Syst. Appl. Acarol., 30: 560-591. https://doi.org/10.11158/saa.30.3.6
- Döker I., Demard E.P., Bolton, S.J. Qureshi J.A. 2023a. Description of Neoseiulus hexaporus sp. nov. and a redescription of Neoseiulus mumai (Acari: Mesostigmata: Phytoseiidae) from Florida citrus groves. Ann. Entomol. Soc. Am., 116: 225-234. https://doi.org/10.1093/aesa/saad015
- Döker I., Joharchi O., Karut K., Kazak C. 2023b. Description of Typhloseiulus anatolicus sp. nov. and redescription of two new records of Phytoseiidae (Acari: Mesostigmata) from Turkey. Acarologia, 63: 553-568. https://doi.org/10.24349/r4a9-vy9o
- Döker I., Karut K., Karaca M.M., Cargnus E., Kazak C. 2018. Internal Transcribed Spacer (ITS) sequences of some Kampimodromus (Acari: Phytoseiidae) species: Is Kampimodromus ragusai a valid species or a synonym of Kampimodromus aberrans? Syst. Appl. Acarol., 23: 2237-2243. https://doi.org/10.11158/saa.23.11.15
- Döker I., Khaustov V.A., Joharchi O., Jung C., Marchenko I.R. 2022. Variability of dorsal shield in different populations of Amblyseius ishizuchiensis Ehara (Acari: Phytoseiidae). Acarologia, 62: 916-926. https://doi.org/10.24349/9cvj-j2ia
- Evans G.O. 1963. Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bull. Br. Mus. Nat. Hist., Zool., 10: 275-303. https://doi.org/10.5962/bhl.part.20528
- Ferragut F., Navia D. 2022. The genus Neoseiulus Hughes (Mesostigmata: Phytoseiidae) in the Espinhaço Range, a great reservoir of biodiversity. Zootaxa, 5120: 523-542. https://doi.org/10.11646/zootaxa.5120.4.4
- Garman P. 1948. Mite species from apple trees in Connecticut. Bull. Conn. Agric. Exp. Stn., 520: 1-27.
- Gerson U. 2014. Pest control by mites (Acari): present and future. Acarologia, 54: 371-394. https://doi.org/10.1051/acarologia/20142144
- Guanilo A.D., Moraes G.J. de, Toledo S., Knapp M. 2008. Phytoseiid mites (Acari: Phytoseiidae) from Argentina, with description of a new species. Zootaxa, 1884: 1-35. https://doi.org/10.11646/zootaxa.1884.1.1
- Hughes A.M. 1948. The mites associated with stored food products. London, Ministry of Agriculture and Fisheries, H. M. Stationary Office, 168 pp.
- Kanouh M., Kreiter S., Douin M., Tixier M.-S. 2012. Revision of the genus Neoseiulella Muma (Acari: Phytoseiidae): re-description of species, synonymy assessment, biogeography, plant supports and key to adult females. Acarologia, 52: 259-348. https://doi.org/10.1051/acarologia/20122048
- Khaustov V.A., Döker I., Joharchi O., Kazakov D.V., Khaustov A.A., Moradi M., Fang X.-D., Klimov P. 2022. A new, broadly distributed species of predacious mites, Neoseiulus neoagrestis sp. nov., (Acari: Phytoseiidae) discovered through GenBank data mining and extensive morphological analyses. Syst. Appl. Acarol., 27: 2038-2061. https://doi.org/10.11158/saa.27.10.14
- Khaustov V.A., Döker I., Joharchi O., Khaustov A.A. 2020. A new record and a redescription of Typhlodromips montanus (Wainstein, 1962) comb. n. (Acari: Phytoseiidae) from the Altai Republic, Russia. Acarina, 28: 203-211. https://doi.org/10.21684/0132-8077-2020-28-2-203-211
- Knapp M., van Houten Y., van Baal E., Groot T. 2018. Use of predatory mites in commercial biocontrol: current status and future prospects. Acarologia, 58 (Suppl.): 72-82. https://doi.org/10.24349/acarologia/20184275
- Kolodochka L.A. 1998. Two new tribes and the main results of a revision of Palearctic phytoseiid mites (Parasitiformes, Phytoseiidae) with the family system concept. Vestn. Zool., 32: 51-63 [in Russian].
- Kolodochka L.A. 2003. A new species of the genus Kampimodromus (Parasitiformes, Phytoseiidae) from Ukraine and Moldova. Acarina, 11: 51-55.
- Kolodochka L.A. 2006. Phytoseiid mites of the Palaearctic Region (Parasitiformes, Phytoseiidae): faunistic, taxonomy, ecomorphology, evolution. Vestn. Zool., suppl. 21, 250pp. [in Russian].
- Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Mem. Ent. Soc. Can., 47: 1-64. https://doi.org/10.4039/entm9747fv
- Lofego A.C., Barbosa M. F. de C., Demite P. R., Moraes G. J. de 2024. Phytoseiidae (Acari: Mesostigmata) of the subfamily Amblyseiinae from Brazil. Zootaxa, 5439: 1-306. https://doi.org/10.11646/zootaxa.5439.1.1
- McMurtry J.A., Moraes G.J. de., Sourassou N.F. 2013. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol., 18: 297-320. https://doi.org/10.11158/saa.18.4.1
- Moraes G.J. de, Oliveira A.R., Zannou I.D. 2001. New phytoseiid mites (Acari: Phytoseiidae) from tropical Africa. Zootaxa, 8: 1-10. https://doi.org/10.11646/zootaxa.8.1.1
- Nesbitt H.H.J. 1951. A taxonomic study of the Phytoseiidae (Family Laelaptidae) predaceous upon Tetranychidae of economic importance. Zool. Verh., 12: 64 pp.+ 32 plates.
- Oudemans A.C. 1930. Acarologische Aanteekeningen. CI. Entomol. Ber., 8: 48-53.
- Papadoulis G.Th., Emmanouel N.G., Kapaxidi E.V. 2009. Phytoseiidae of Greece and Cyprus (Acari: Mesostigmata). West Bloomfield, Indira Publishing House, 200 pp.
- Rahmani-Piyani A., Shishebor P., Faraji F. 2025. A complementary description of Neoseiulus longilaterus (Athias-Henriot) (Acari: Phytoseiidae) based on additional specimens from Iran. Syst. Appl. Acarol. 30: 641-652. https://doi.org/10.11158/saa.30.4.1
- Robin, Döker I., Kaur P., Bhullar M.B., Gowda C.C. 2025. A new species of Neoseiulella Muma (Parasitiformes: Phytoseiidae) from Northwestern India. Acarologia, 65: 762-770. https://doi.org/10.24349/wpop-gesj
- Rowell H.L., Chant D.A., Hansell R.I.C. 1978. The determination of setal homologies and setal patterns on the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata). Can. Entomol., 110: 859-876. https://doi.org/10.4039/Ent110859-8
- Tsolakis H., Ragusa S. 2015. Considerations on systematics of the Phytoseiidae (Acari: Mesostigmata), with definition of a new species group and description of a new species. Zootaxa, 3926: 229-243. https://doi.org/10.11646/zootaxa.3926.2.4
- Tsolakis H., Ragusa S. 2016. On the identity of Neoseiulus fallacis (Garman 1948) (Parasitiformes, Phytoseiidae) redescription of the species and description of the new species Neoseiulus garmani. Int. J. Acarol., 42: 394-404. https://doi.org/10.1080/01647954.2016.1205134
- Tsolakis H., Tixier M.-S., Kreiter S., Ragusa S. 2012. The concept of genus within the family Phytoseiidae (Acari: Parasitiformes): historical review and phylogenetic analyses of the genus Neoseiulus Hughes. Zool. J. Linn. Soc., 165: 253-273. https://doi.org/10.1111/j.1096-3642.2011.00809.x
- Zannou I.D., Moraes G.J. de, Ueckermann E.A., Oliveira A.R., Yaninek J.S., Hanna R. 2006. Phytoseiid mites of the genus Neoseiulus Hughes (Acari: Phytoseiidae) from sub-Saharan Africa. Int. J. Acarol., 32: 241-276. https://doi.org/10.1080/01647950608684467



2026-02-05
Date accepted:
2026-04-03
Date published:
2026-04-10
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Döker, Ismail; Demite, Peterson R.; Atchia, Isabelle; Gasparro-Brewer, Jude and Bolton, Samuel J.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)