



First records of the marine water mite family Pontarachnidae Koenike, 1910 from New Zealand (Acari: Hydrachnidia), with the description of one new species
Pešić, Vladimir  1
; Batista-Ribeiro, Dante
2
; Pepato, Almir R.
3
and Smit, Harry
4
1
; Batista-Ribeiro, Dante
2
; Pepato, Almir R.
3
and Smit, Harry
4
1✉ Department of Biology, University of Montenegro, Cetinjski put b.b., 81000 Podgorica, Montenegro.
2Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Zoologia, Laboratório de Sistemática e Evolução de Ácaros Acariformes, Av. Antônio Carlos, 6627, Pampulha, Belo Horizonte - MG, Brazil.
3Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Zoologia, Laboratório de Sistemática e Evolução de Ácaros Acariformes, Av. Antônio Carlos, 6627, Pampulha, Belo Horizonte - MG, Brazil.
4Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, the Netherlands.
2026 - Volume: 66 Issue: 1 pages: 134-148
https://doi.org/10.24349/rje7-i3neZooBank LSID: 26C4BB41-22E5-481D-91C2-2FD5F1C30704
Original research
Keywords
Abstract
Introduction
Pontarachnidae Koenike, 1910 is the only family of Hydrachnidia that occurs in the marine environments, primarily in subtropical and tropical regions. A few species have been described from brackish or maybe even fresh water, but almost always in estuaries or close to the sea (Cook 1986, 1996; Smit 2007; Pešić 2013b). Two species from South Korea are adapted to live in marine interstitial habitats (Pešić 2013a). At the global level, the highest number of pontarachnid species is found in the tropical central Indo-Pacific region (Pešić et al. 2012a), suggesting that temperature gradients may play an important role in shaping the diversity of this group. Nothing is known about the life cycle of the species (Cook 1996). In most water mites, the larval stage is parasitic on insects; however, because few insects inhabit marine environments, this stage may be absent or bypassed in pontarachnids (Smit and Alberti 2010). The diet of pontarachnids also remains unknown and requires laboratory studies to determine their food preferences (Pešić et al. 2019).
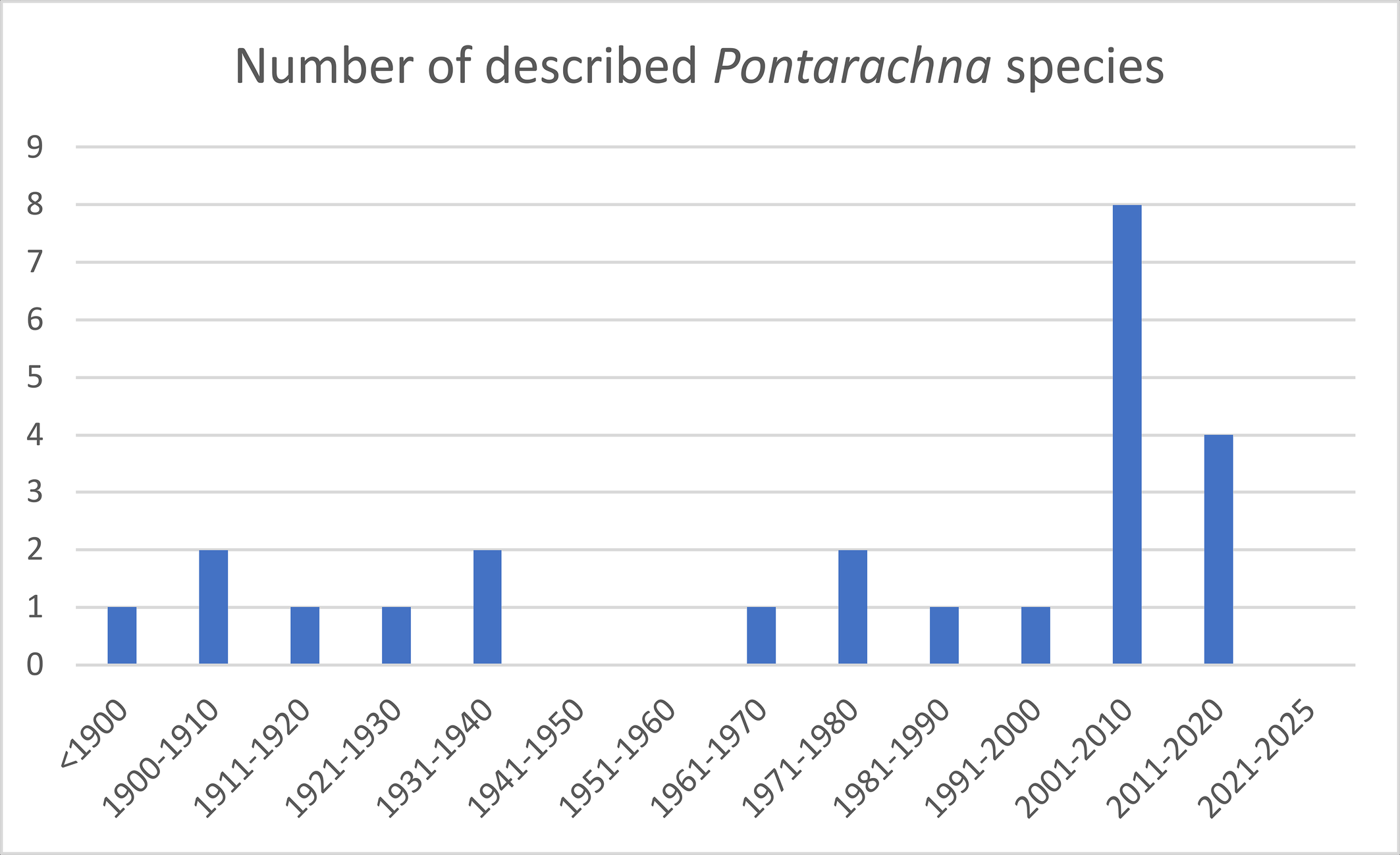
Within the Pontarachnidae two genera are known, i.e. Pontarachna Philippi, 1840 and Litarachna Walter, 1925. Worldwide 34 Litarachna species and 24 Pontarachna species are known (Asadi et al. 2019, Chatterjee et al. 2019; Pešić et al. 2012, 2019, 2020; Montes-Ortiz et al. 2021). Most Litarachna species were described in the 21th century (Figure 1), the same applies to Pontarachna (Figure 2). A list of Litarachna and Pontarachna species can be found in the World Register of Marine Species (WORMS https://www.marinespecies.org/ ![]() ).
).
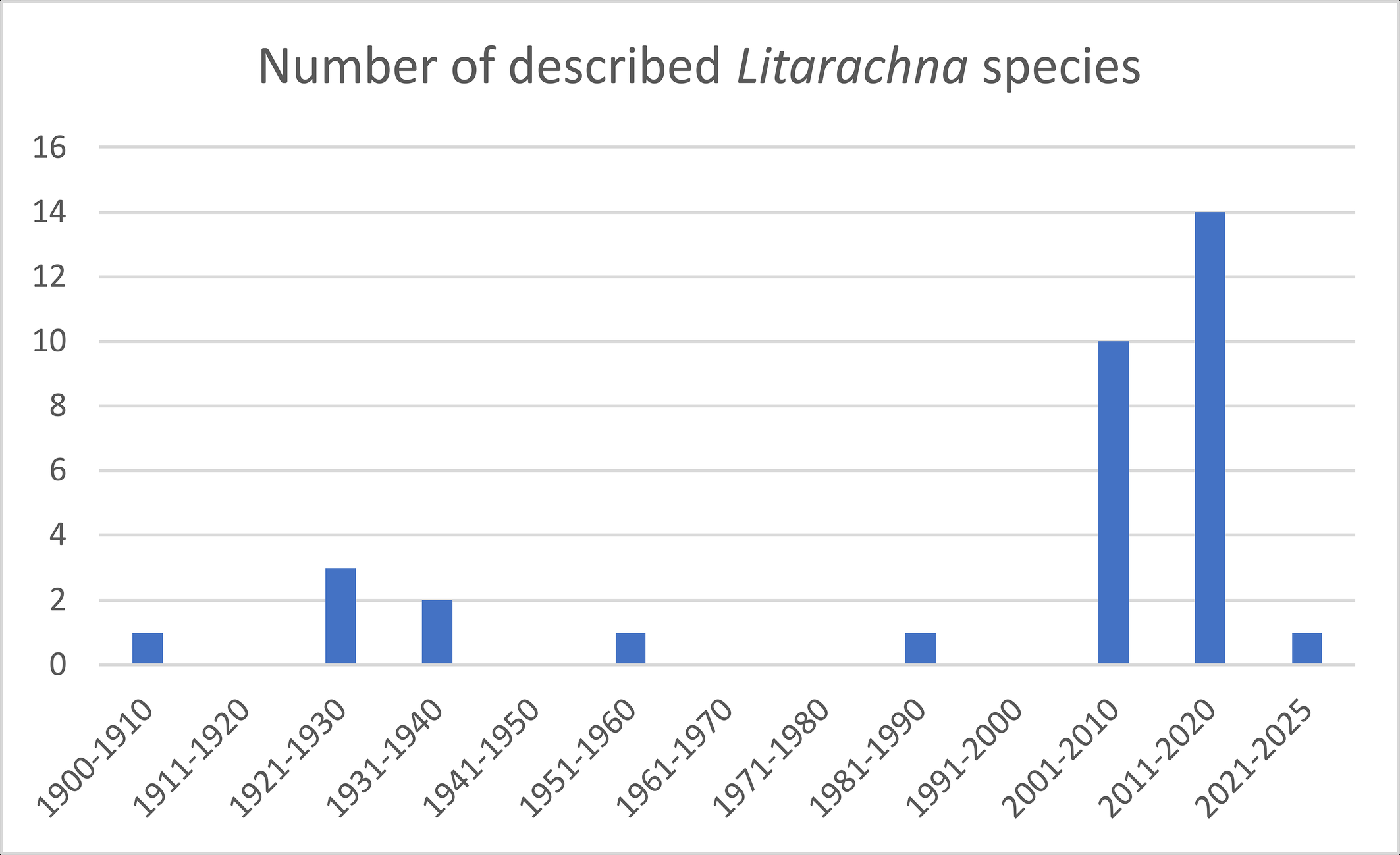
This paper provides the first records of the genus Litarachna for New Zealand. One species is described as new for science, and one species is reported for the first time for New Zealand. Finally, we provide a worldwide key for the genus.
Material and methods
Material from the Clive River was collected by AP and HS, material from other locations was collected by AP. Material of the Clive River was fixed in ethanol 96% and Koenike-fluid, material from other locations in ethanol 96%.
The holotype is lodged the National Museum of New Zealand, Wellington (NMNZ), paratypes and non-type material in the New Zealand Arthropod Collection of Landcare Research (NZAC), Auckland and Naturalis Biodiversity Center, Leiden (RMNH).
Morphological nomenclature follows Wiles et al. (2002) and Pešić et al. (2019). The following abbreviations are used: Cx-I to -IV – first to forth coxae; Cxgl-4 – coxoglandulare 4 (''E4» according to Wiles et al. 2002); I-Leg-1 to -6 – first to sixth segments of first leg; n – number of specimens examined; P1 to P5 – palp segments 1 to 5; V1 to V3 – first to third ventral setae sensu Wiles et al. (2002); V3 gland – ventral glandularium sensu Wiles et al. (2002); W1 to W3 – first to third wheel-like acetabula. All measurements are given in µm.
Systematics
Family Pontarachnidae Koenike, 1910
Genus Litarachna Walter, 1925
Litarachna amnicola Cook, 1986
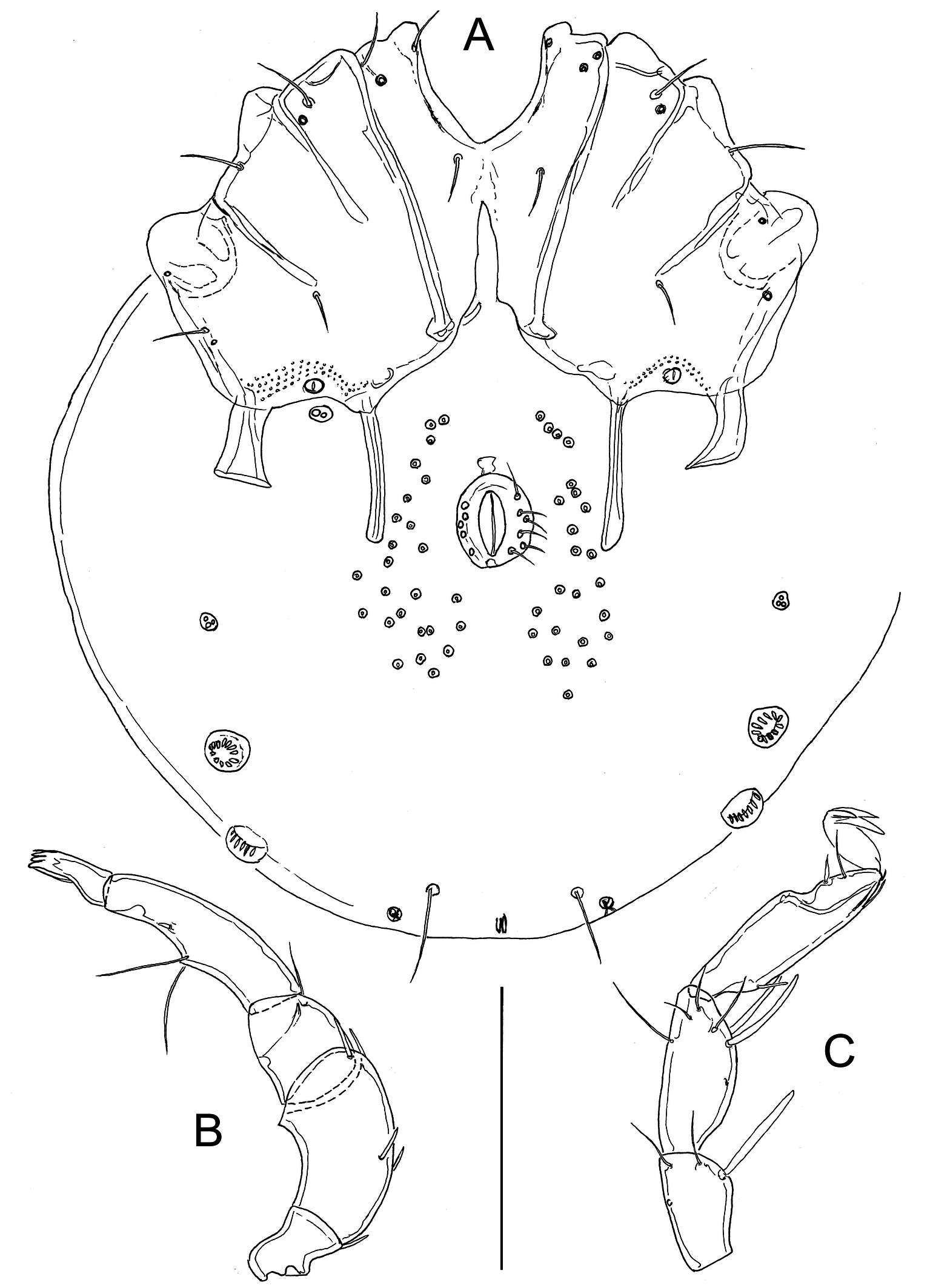
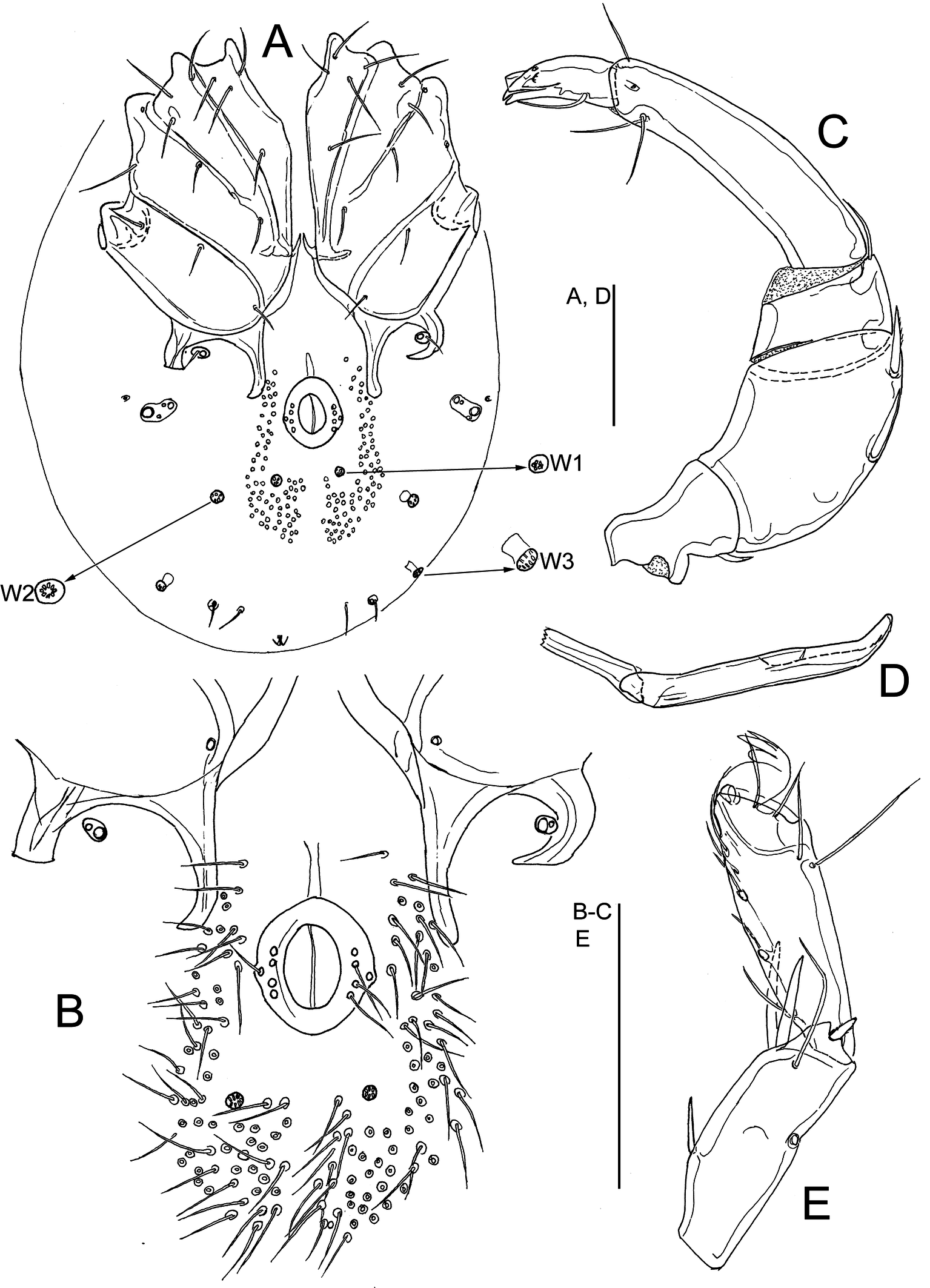
Figures 3–4
Material examined — North Island. 2/3/0, Clive River at Clive Bridge, Clive, about 1700 m upstream from the sea, floating freshwater macrophytes, 39°34.879″S 176°55.011″E, 20 Feb. 2025, leg. H. Smit (NZAC); 3/13/0, Estuary Clive River, Clive, close to the sea, in a lagoon dammed from the sea by a sandspit, 39°34.422″S 176°55.504″E, 19 Feb. 2025, leg. H. Smit, one male and one female dissected and slide mounted (RMNH).
Remarks — Initially, the species was described from the interstital in a river several kilometers from the sea in Tasmania, Australia (Cook 1986). Later on, Pesic & Smit (2009) reported the species from an estuary, also on Tasmania. The latter authors were not 100% sure about the identification. According to Cook (1986), the species has no swimming setae, but the specimens of Pesic and Smit did have swimming setae. Moreover, P2 has a small distoventral extension, not present in Cook's specimens. The specimens from New Zealand also have swimming setae. As the specimens from Pesic and Smit (2009) and from New Zealand match the description of L. amnicola, including V3 gland fused with Cx-IV, a rare character in Litarachna, we think that all these specimens are conspecific with L. amnicola. The swimming setae are rather fragile, and probably lost in Cook's two males.
We found one male from the Clive river estuary that differs by the increased number of setae in the sclerotized ring of the male genital field (5-6 compared to 4 setae in typical specimens of L. amnicola). Below we give a description of this specimen. A final decision on the taxonomic position of this specimen requires molecular analysis to assess the possibility that it is a new species within the L. amnicola complex.
Morphology — Male — Idiosoma 338 long and 270 wide. First coxal plates fused medially; suture line Cx-I/II complete, suture line Cx-II/III and Cx-III/IV incomplete; gnathosomal bay 43 long; coxal field 137 long and 230 wide. Lateral apodemes of Cx-IV shorter than medial apodemes, the latter extending beyond anterior margin of genital field (Figure 3A). Genital field consisting of a sclerotized ring, 33 long and 27 wide, with five to six pairs of setae, about 25 pairs of perigenital setae free in integument around the sclerotized ring. A pair of small platelets with Cxgl-4 (E4 according to Wiles et al. 2002) and associated setae placed between the posterior lateral and medial apodemes of Cx-IV; the large glandularia-like structure (V3 gland according to Wiles et al. 2002) fused with Cx-IV. Posterior to the genital field a pair of platelets with three pores and two pairs of large wheel-like acetabula, with many radiating spokes. Excretory pore unsclerotized, near the posterior idiosoma margin. Dorsal length/height of palp segments: P1, 16/16; P2, 69/34; P3, 29/30; P4, 77/23; P5, 28/13; dorsal length P5/P4 ratio 0.9; P2 with small ventrodistal peg-like projection; ventral margin of P4 with a strongly developed setal tubercle (Figure 3B). Dorsal lengths of I-Leg-3–6: 38, 44, 59, 77; dorsal lengths of IV-Leg: 44, 39, 56, 84, 98, 119. Number of swimming setae: II-Leg-5, 1; III-Leg-4, 1; III-Leg-5, 2; IV-Leg-4, 1; IV-Leg-5, 2.
In addition, we provide a description of a female collected together with above described male.
Female — Similar to male except in the shape of genital field (Figure 4A). Idiosoma 373 long and 297 wide. Gnathosomal bay 47 long; coxal field 142 long and 244 wide. Medial apodemes of Cx-IV extending to the posterior margin of genital field; genital field 66 long, pre- and post-genital sclerites bowed, pre-genital sclerite 45 wide, post-genital sclerite 46 wide. Dorsal length/height of palp segments: P1, 17/16; P2, 70/33; P3, 26/28; P4, 78/23; P5, 30/11. Chelicera 167 long. Dorsal lengths of I-Leg: 39, 36, 42, 42, 58, 75; dorsal lengths of IV-Leg: 59, 44, 56, 88, 100, 116. Swimming setae as in male.

Litarachna quinqueseta Pešić & Smit n. sp.
ZOOBANK: 6758FDAB-ECF9-4234-9763-5B9598285A25 ![]()
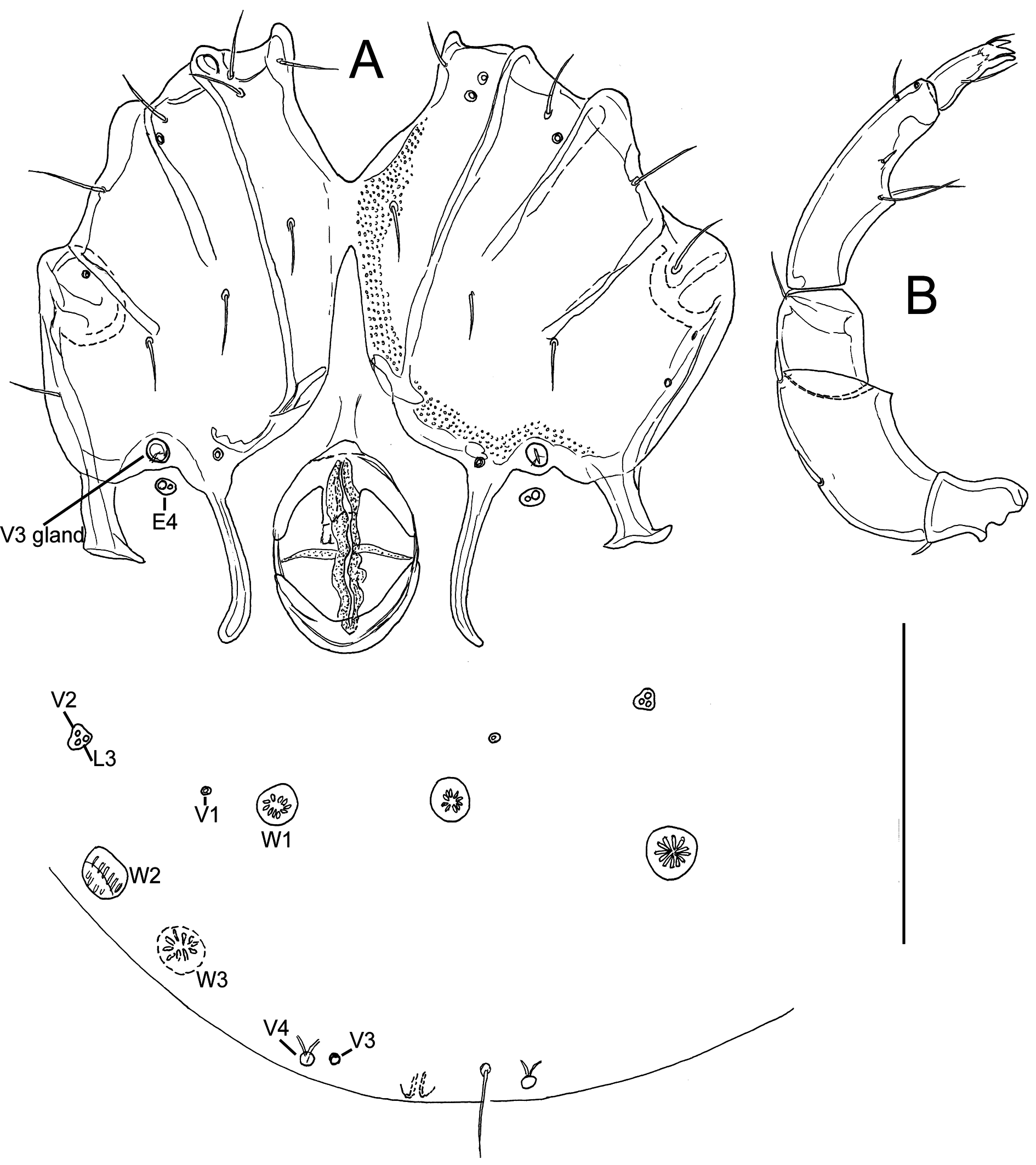
Figures 6–7
Material examined — Holotype, male, North Island, Tawharanui Peninsula, Tawharanui Regional Park, collected from intertidal rocky shore algae, 36°22′44.8″S 174°48′31″E, 6 Dec. 2022, leg. A. Pepato, dissected and slide mounted (NMNZ). Paratypes: 1/2/1, same data as the holotype, all slide mounted; 1/1/1, Tanogio, Tangoio beach, collected from intertidal rocky shore algae, 39°20′13″S 176°55′28″E, 9 Dec. 2022, leg. A. Pepato & P.B. Klimov, all slide mounted (RMNH).
Diagnosis — Sclerotized ring of male genital field with five setae; P5 distinctly shortened, dorsal length P5/P4 ratio 0.31-0.38; chelicera with a straight, pointed claw.
Description — General features — Cx-I separated medially; suture lines Cx-I/II and Cx-III/IV complete, suture line Cx-II/III incomplete; posterior margin of Cx-IV with two pairs of apodemes of moderate length, the medial longer than the lateral ones; a pair of small platelets with Cxgl-4 and associated setae free in the integument between the posterior apodemes of Cx-IV; posterior to the genital field a pair of platelets with three pores and a glandularium-like structure, and three pairs of small wheel-like acetabula, with relatively few radiating spokes; excretory pore unsclerotized, near posterior end of idiosoma (Figures 6A, 7A). P4 slightly bowed, shorter than P2+3, ventral margin without a tubercle. Chelicera with a straight, pointed claw. Number of swimming setae: I-Leg-5, 1 (short; as illustrated in Figure 6E) II-Leg-5, 1; III-Leg-4, 1; III-Leg-5, 1; IV-Leg-4, 1; IV-Leg-5, 2. Male: Medial apodemes of Cx-IV extending beyond the anterior margin of genital field, genital field consisting of a sclerotized ring with five pairs of setae, one seta more laterally than the four other setae; many perigenital setae (60-70 pairs) free in the integument around the genital field (Figure 6B). Female: Medial apodemes of Cx-IV extending to the posterior margin of genital field, pre- and post-genital sclerites bowed (Figure 7A).
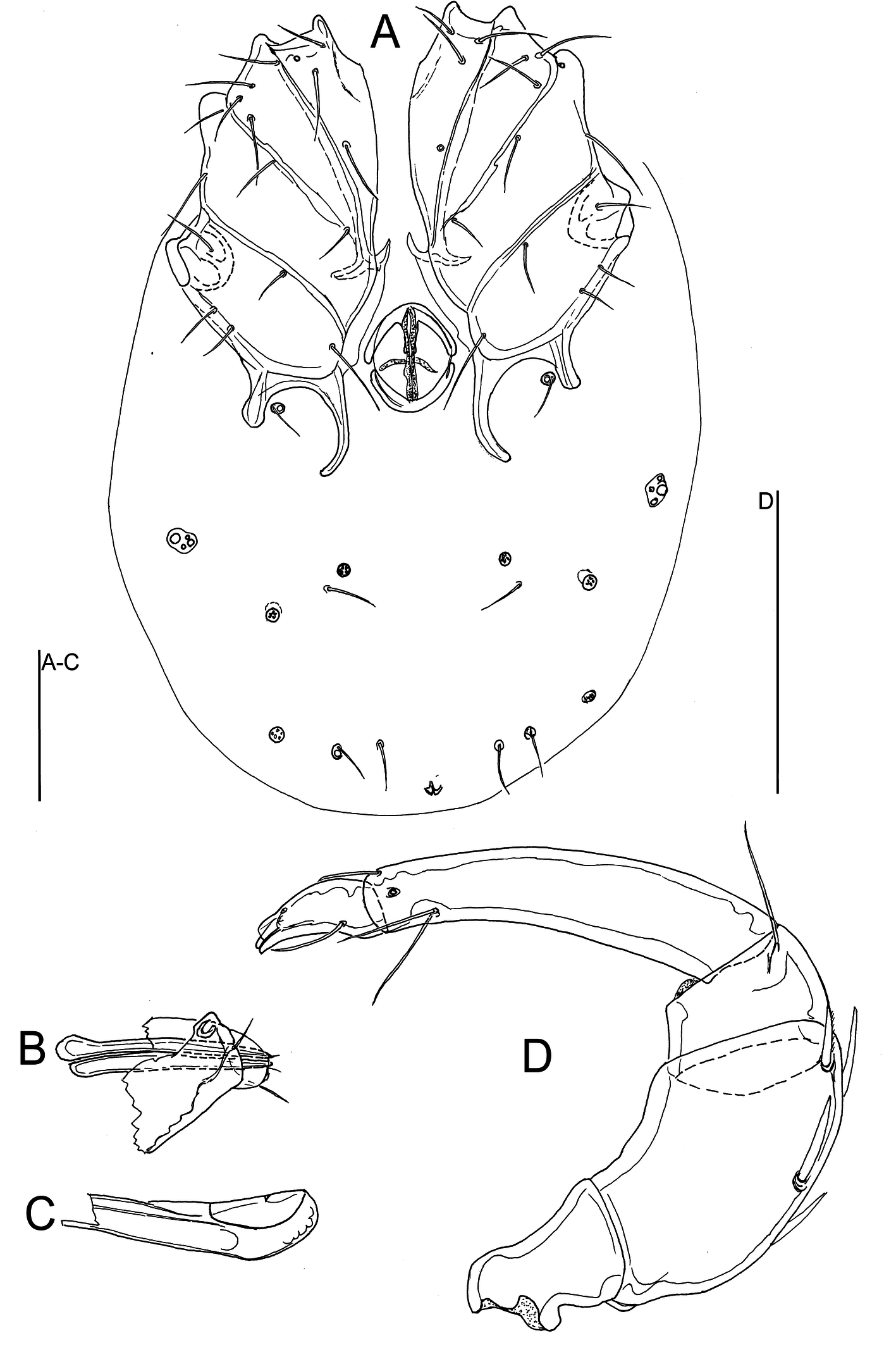
Measurements — Male (holotype) — Idiosoma 434 long and 331 wide; coxal field 211 long and 263 wide; sclerotized ring of genital field 48 long and 42 wide. Dorsal length/height of palp segments: P1, 25/22; P2, 94/54; P3, 33/47; P4, 109/27; P5, 41/17; dorsal length P5/P4 ratio 0.38. Chelicera 295 long, basal segment 194 long; gnathosoma 134 long.
Dorsal lengths of I-Leg: 47, 39, 55, 63, 89, 108; dorsal lengths of IV-Leg-2-6: 58, 64, 100, 122, 128.
Female (n = 1) — Idiosoma 533 long and 403 wide; coxal field 242 long and 323 wide; genital field 79 long, pre-genital sclerite 61 wide, post-genital sclerite 56 wide. Dorsal length/height of palp segments: P1, 25/25; P2, 117/66; P3, 42/56; P4, 142/31; P5, 44/19; dorsal length P5/P4 ratio 0.31. Chelicera 336 long, claw 147 long; gnathosoma 163 long.
Dorsal lengths of I-Leg-2-6: 50, 70, 72, 98, 113; dorsal lengths of IV-Leg-3-6: 77, 116, 141, 147.
Deutonymph — As in adults but lacking a genital field. Idiosoma 291 long and 225 wide. Coxal field 127 long and 184 wide. Gnathosoma 94 long; chelicera 181 long; palp as in adults.
Etymology — The specific epithet is derived from the Latin quinque (five) and setae, referring tothe presence of five setae on the sclerotized ring of male genital field.
Remarks — In regard to the shape of the genital field (in male a patch with perigenital setae distinctly longer than wide, not widened posteriorly) and palp (P4 being much shorter than P2+3 but distinctly longer than P2, P5 shorter than half of P4, not stylet-like), the new species from New Zealand resembles L. thetis Pešić & Smit, 2016. The latter species was originally, established by Pešić & Smit (2016) as a new name for populations from the Sea of Japan (Uchida, 1935) and from South Korea, which were originally assigned by Pešić et al. (2008) to L. denhami. Recently, L. thetis was reported from Hainan Island, China (Li et al., 2025).
The new species can be separated from L. thetis in having five pairs (four on a medial row and one lateral of this row) of setae in sclerotized ring of male genital field (four pairs of setae in L. thetis) and P5 distinctly shortened, dorsal length P5/P4 ratio 0.31-0.38 (in L. thetis and P5 nearly half as long as P4). Moreover, Litarachna quinqueseta n. sp. is unique and can be separated from all other species of the genus by the shape of chelicera with a straight pointed claw.
Discussion
So far, water mites of the marine family Pontarachnidae have not been recorded from New Zealand. In this study we report two pontarachnid species, one of them described as new to science. All specimens were collected on the North Island. The North Island possesses a warm temperate climate characterized by mild winters and subtropical summer conditions. In contrast, the South Island experiences a cold temperate climate with precipitation distributed relatively evenly throughout the year, though rainfall typically reaches its annual minimum during the winter (Willis et al. 2007).
Most of the 54 known species of pontarachnid mites are found in subtropical and tropical regions, much fewer species are known from cold temperate zones (e.g., Northwest Pacific) (see Pešić et al. 2012 for discussion). Large parts of the world, e.g. major parts of the temperate and tropical Atlantic Ocean, still have no reported data on pontarachnid mites, indicating the need for a more extensive sampling campaign to gain a clearer insight into the diversity and distribution of this globally neglected group of marine meiofauna.
Key to the Litarachna species of the world
Gonopore surrounded by a sclerotized ring bearing a few pairs of setae, few to many perigenital setae free in the surrounding integument
...... Males
Genital field with well-developed pre-and postgenital sclerites
...... Females
Key to the males
Males are unknown for Litarachna enigmatica Pešić, Durucan & Chatterjee, 2018, L. gracilis Pešić, 2013, L. haleioides Smit, 2016, L. minuta Pešić, Chatterjee & Marshall, 2013, L. sabangensis K.O. Viets, 1984 and L. triangularis Smit, 2009.
1. P2 without a ventral extension
...... 2
— P2 with a ventral extension
...... 19
2(1) P4 with ventral setal tubercles
...... 3
— P4 without ventral setal tubercles
...... 15
P2 without a ventral extension, P4 with setal tubercles
3(2) Sclerotized ring of genital field with two pairs of setae
...... Litarachna guadeloupensis Pešić, Chatterjee & Schizas, 2020 (Caribbean Sea, Guadeloupe)
— Sclerotized ring of genital field with more than two pairs of setae
...... 4
4(3) Ocular pigment absent; sclerotized ring with three pairs of setae
...... Litarachna lukai Pešić, 2013 (Yellow Sea, South Korea)
— Ocular pigment present; sclerotized ring with four pairs of setae
...... 5
5(4) Cx-I fused medially
...... 6
— Cx-I separated medially
...... 11
6(5) Genital field with more than 15 pairs of perigenital setae
...... 7
— Genital field with 2-5 pairs of perigenital setae
...... 8
7(6) Genital field with about 20 pairs of perigenital setae, V3 gland fused with Cx-IV
...... Litarachna amnicola Cook, 1986 (Tasmania, New Zealand)
— Genital field with about 50 pairs of perigenital setae, V3 gland not fused with Cx-IV
...... Litarachna caribica Pešić, Chatterjee & Schizas, 2008 (Caribbean Sea, Netherlands Antilles; Pacific Ocean, Panama)
8(6) Genital field with five pairs of perigenital setae, P4 near setal tubercles a peg-like seta
...... Litarachna lopezae Pešić, Chatterjee, Alfaro & Schizas, 2014 (Caribbean Sea, Puerto Rico)
— Genital field with 2-3 pairs of perigenital setae, P4 without peg-like seta
...... 9
9(8) Genital field with three pairs of perigenital setae, V3 gland on one platelet with Cxgl-4
...... Litarachna degiustii Cook, 1958 (Caribbean Sea, Bahamas, Curaçao)
— Genital field with 2-3 pairs of perigenital setae, V3 gland and Cxgl-4 separated
...... 10
10(9) Wheel-like acetabula very large (maximum diameter > 15 µm), genital field wider than long
...... Litarachna indica Pešić, Chatterjee & Ingole, 2012 (Arabian Sea, India)
— Wheel-like acetabula smaller, genital field longer than wide
...... Litarachna brasiliensis Smit, 2007 (Brazil)
11(5) Medial posterior apodemes of Cx-IV a hammer-shaped, genital field with four pairs of perigenital setae
...... Litarachna marshalli Wiles, Chatterjee & De Troch, 2002 (South Africa)
— Medial posterior apodemes of Cx-IV regular, not a hammer-shaped, genital field with more than four pairs of perigenital setae
...... 12
12(11) Cxgl-4 on the same platelet with V3 gland
...... Litarachna duboscqi Walter, 1925 (Mediterranean Sea, Black Sea)
— Cxgl-4 separated from V3 gland
...... 13
13(12) V3 gland and Cxgl-4 between posterior apodemes of Cx-IV, genital field with about ten pairs of perigenital setae
...... 14
— V3 gland and Cxgl-4 not between posterior apodemes of Cx-IV, posteriorly to posterolateral apodeme of Cx-IV, genital field with about 25-35 pairs of perigenital setae
...... Litarachna sagamiensis Moto & Abé, 2013 (Japan)
14(13) Medial apodemes of Cx-IV short, not reaching anterior margin of genital field
...... Litarachna antalyaensis Pešić, Durucan & Chatterjee, 2018 (Mediterranean Sea, Türkiye)
— Medial apodemes of Cx-IV longer, extending beyond the anterior margin of genital field
...... Litarachna hongkongensis Smit, 2002 (South China Sea, China)
P2 without a ventral extension, P4 without setal tubercles
15(2) Chelicera with a straight, pointed claw
...... Litarachna quinqueseta n. sp. (New Zealand)
— Chelicera with a curved claw
...... 16
16(15) Sclerotized ring with two pairs of setae, medial apodemes of Cx-IV very short
...... Litarachna muratsezgini Pešić, Durucan & Zawal, 2019 (Mediterranean Sea, Türkiye)
— Sclerotized ring with four pairs of setae, medial apodemes of Cx-IV much longer
...... 17
17(16) P3 dorsally longer than high, P4 much shorter than P2+3, but noticeably longer than P2, P5 very slender, stylet-like, about four times long as the distal height of P4
...... Litarachna denhami (Lohmann, 1909) (Western Australia, South Africa, India, Egypt, New Guinea)
— P3 proximally as high as long or higher, P4 shorter than P2+3 but a little longer than P2, P5 less slender, about twice as long as the distal height of P4
...... 18
18(17) P5 shorter than ½ of P4, P4 ventral setae not at the same level, patch of perigenital setae longer than wide, posteriorly not widened
...... Litarachna thetis Pešić & Smit, 2016 (South Korea, Japan, China)
— P5 longer than ½ of P4, P4 ventral setae at the same level, patch of perigenital setae wide as long, posteriorly widened
...... Litarachna communis Walter, 1925 (Mediterranean Sea, Black Sea)
P2 with a ventral extension
19(1) P4 without ventral setal tubercles
...... 20
— P4 with ventral setal tubercles
...... 24
P2 with a ventral extension, P4 without setal tubercles
20(19) Genital field with two pairs of perigenital setae, P4 with a minute denticle-like extension near ventral setae
...... Litarachna kamui Uchida, 1935 (Japan)
— Genital field with many perigenital setae, P4 without a minute denticle-like extension near ventral setae
...... 21
21(20) Genital field lying posterior to apodemes of Cx-IV, patch of perigenital setae extending almost to posterior idiosoma margin
...... Litarachna muelleri Smit, 2008 (Malaysia)
— Genital field lying between apodemes of Cx-IV, patch of perigenital setae not extending far beyond genital field
...... 22
22(21) Medial apodemes of Cx-IV long, extending beyond posterior margin of genital field, palp long (P2 170, P4 162)
...... Litarachna halei (Womersley, 1937) (Western and southern Australia)
— Medial apodemes of Cx-IV shorter, extending not or just beyond posterior margin of genital field, palp shorter (P2 < 70, P4 < 90)
...... 23
23(22) Sclerotized ring with 2–3 setae on each side
...... Litarachna bruneiensis Pešić, Chatterjee, Marshall & Pavićević, 2011 (Brunei Darussalam)
— Sclerotized ring with four pairs of setae on each side
...... Litarachna belicensis Montes-Ortiz, Goldschmidt, Vásquez-Yeomans & Elías-Gutiérrez, 2021 (Carribean Sea, Belize)
P2 with a ventral extension, P4 with setal tubercles
24(19) P4 without a peg-like setae near setal tubercles
...... 25
— P4 with a peg-like seta near setal tubercles
...... 26
25(24) Medial apodemes of Cx-IV very long, much longer than lateral apodemes; V3 gland and Cxgl -4 between posterior apodemes of Cx-IV
...... Litarachna cawthorni Wiles, Chatterjee & De Troch, 2002 (Kenya)
— Medial apodemes of Cx-IV little longer than lateral apodemes; V3 gland and Cxgl-4 not between posterior apodemes of Cx-IV
...... Litarachna curtipalpis Smit, 2003 (Western Australia, Singapore, China)
26(24) V3 gland fused with Cx-IV
...... 27
— V3 gland not fused with Cx-IV
...... Litarachna smiti Pešić, Chatterjee & Ahmed Abada, 2008 (Red Sea, Saudi Arabia)
27(26) Medial apodemes of Cx-IV longer, extending to the posterior margin of genital field
...... Litarachna bartschae Smit, 2003 (Western Australia, Brunei Darussalam, Singapore)
— Medial apodemes of Cx-IV short, reaching anterior margin of genital field
...... Litarachna madagascariensis Pešić, 2013 (Madagascar)
Key to the females
Females are unknown of the following species: Litarachna guadeloupensis.
1 Postgenital sclerite with a pair of wheel-like acetabula
...... Litarachna enigmatica Pešić, Durucan & Chatterjee, 2018 (Mediterranean Sea, Türkiye)
— Postgenital sclerite without a pair of wheel-like acetabula
...... 2
2(1) P2 without a ventral extension
...... 3
— P2 with a ventral extension
...... 20
3(2) P4 with ventral setal tubercles
...... 4
— P4 without ventral setal tubercles
...... 16
P2 without ventral extension, P4 with setal tubercles
4(3) Cx-I medially fused
...... 5
—Cx-I medially separated
...... 11
5(4) V3 gland fused with Cxgl-4, medial apodemes of Cx-IV extending to posterior margin of genital field
...... Litarachna degiustii
—V3 gland separated from Cxgl-4, medial apodemes of Cx-IV shorter, extending not or only slightly beyond posterior margin of genital field
...... 6
6(7) V3 gland fused with Cx-IV
...... 7
— V3 gland not fused with Cx-IV
...... 9
7(6) Lateral apodemes of Cx-IV indistinct, Cxgl-4 fused with Cx-IV
...... Litarachna minuta Pešić, Chatterjee & Marshall, 2013 (Brunei Darussalam)
— Lateral apodemes of Cx-IV distinct, Cxgl-4 not fused with Cx-IV
...... 8
8(7) P4 near setal tubercles a peg-like seta
...... Litarachna lopezae
— P4 without peg-like seta
...... Litarachna amnicola
9(6) Anterior pregenital sclerite arrow-shaped
...... Litarachna brasiliensis
— Anterior pregenital sclerite more rounded
...... 10
10(9) Wheel-like acetabula very large
...... Litarachna indica
— Wheel-like acetabula smaller
...... Litarachna caribica
11(4) V3 gland and Cxgl-4 fused
...... Litarachna duboscqi
— V3 gland and Cxgl-4 separated
...... 12
12(11) Medial apodemes long, extending well beyond posterior margin of genital field
...... 13
— Medial apodemes short, extending not or only slightly beyond posterior margin of genital field
...... 14
13(12) V3 gland and Cxgl-4 between posterior apodemes of Cx-IV
...... Litarachna hongkongensis
— V3 gland and Cxgl-4 not between posterior apodemes of Cx-IV, located posteriorly to posterolateral apodeme of Cx-IV
...... Litarachna sagamiensis
14(12) Anterior and posterior genital sclerite touching
...... Litarachna marshalli
— Anterior and posterior genital sclerites not touching
...... 15
15(14) Ocular pigment absent; medial and lateral apodemes of Cx-IV or equal length
...... Litarachna lukai
— Ocular pigment present; medial apodemes of Cx-IV longer than lateral apodemes
...... Litarachna antalyaensis
P2 without ventral extension, P4 without setal tubercles
16(3) Chelicera with a straight, pointed claw
...... Litarachna quinqueseta n. sp.
— Chelicera with a curved claw
...... 17
17(16) Medial apodemes of Cx-IV broad, shorter than the lateral ones
...... Litarachna muratsezgini
— Medial apodemes of Cx-IV longer than the lateral ones
...... 17
18(17) P3 dorsally longer than high, P4 much shorter than P2+3, but noticeably longer than P2, P5 very slender, stylet-like, about four times long as the distal height of P-4
...... Litarachna denhami
— P3 proximally as high as long or higher, P4 shorter than P2+3 but a little longer than P2, P5 less slender, about twice as long as the distal height of P4
...... 19
19(18) P5 shorter than ½ of P4, P4 ventral setae not at the same level
...... Litarachna thetis
— P5 longer than ½ of P4, P4 ventral setae at the same level
...... Litarachna communis
P2 with a ventral extension
20(2) Ocular pigment absent; genital field very long, P4 with a small ventral peg-like seta
...... Litarachna gracilis Pešić, 2013 (Yellow Sea, South Korea)
— Ocular pigment present; genital field not very long, but if long, P4 without a peg-like seta
...... 21
21(20) P4 without seta tubercles
...... 22
— P4 with setal tubercles
...... 28
P2 with a ventral extension, P4 without seta tubercles
22(21) Genital sclerites forming a complete ring
...... 23
— Genital sclerites not forming a complete ring
...... 27
23(22) Medial apodemes of Cx-IV long, more than three times as long as lateral apodeme
...... Litarachna halei
— Medial apodeme less long, less than two times as long as lateral apodeme
...... 24
24(23) Medial and lateral apodeme of Cx-IV of same length
...... Litarachna muelleri
— Medial apodeme of Cx-IV distinctly longer than lateral apodeme
...... 25
25(24) Most anterior pair of wheel-like acetabula (W1) lying close to medial apodeme of Cx-IV and distanced from each other
...... Litarachna bruneiensis
— Most anterior pair of wheel-like acetabula lying distanced from medial apodeme of Cx-IV and lying much closer to each other
...... 26
26(25) Ventral extension of P2 relatively small
...... Litarachna belicensis
— Ventral extension of P2 relatively large
...... Litarachna triangularis Smit, 2009 (Singapore)
27(22) Genital field very long (2.5 times long as wide), P4 with a minute denticle-like extension near ventral setae
...... Litarachna kamui
— Genital field short, P4 without a minute denticle-like extension near ventral setae
...... Litarachna haleioides Smit, 2016 (Eastern Australia)
P2 with a ventral extension, P4 with setal tubercles
28(21) P4 with a peg-like seta near setal tubercles
...... 29
— P4 without a peg-like seta near setal tubercles
...... 31
29(28) V3 gland fused with Cx-IV
...... 30
— V3 gland not fused with Cx-IV
...... Litarachna smiti
30(29) Anterior and posterior genital sclerite distanced, medial apodemes of Cx-IV long, extending well beyond posterior margin of genital field
...... Litarachna bartschae
— Anterior and posterior genital sclerite less distanced, medial apodemes of Cx-IV less long, just extending beyond posterior margin of genital field
...... Litarachna madagascariensis Pešić, 2013 (Madagascar)
31(28) Suture lines of Cx-III/IV complete
...... Litarachna cawthorni
— Suture lines of Cx-III/IV incomplete
...... 32
32(31) Pregenital sclerite broad, ventral extension of P2 short, triangular
...... Litarachna curtipalpis
— Pregenital sclerite narrow, sickle-shaped, ventral extension of P2 longer, more slender
...... Litarachna sabangensis K.O. Viets, 1984 (Philippines)
Acknowledgements
HS is indebted to Truus van der Pal (Alkmaar) for her assistance during the fieldwork, AP is indebted to Zhi-Qiang Zhang (Landcare Research) for Local Organizing Committee of the XVI International Congress of Acarology for funding his travel to New Zealand, DBR is indebted to the Fundação de Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for their scholarship, and Conselho Nacional de Desenvolvimento Científico e Tecnológico, MCTI/CNPQ Nº 16/2024 - Project funded under Track 1: Cooperative Project (402926/2024-2). We thank three anonymous reviewers whose constructive comment improved the manuscript.
References
- Asadi M., Smit H., Pešić V. 2019. A new marine mite (Acari: Hydrachnidia: Pontarachnidae). Cah. Biol. Mar., 60: 567-570.
- Chatterjee T., Schizas NV., Pešić V. 2019. A checklist of Pontarachnidae (Acari: Hydrachnidia) and notes on distributional patterns of the species. Zootaxa, 4619(3): 527-544. https://doi.org/10.11646/zootaxa.4619.3.6
- Cook DR. 1986. Water mites from Australia. Mem. Amer. Ent. Inst., 40: 1-568.
- Cook DR. 1996. A freshwater species of Pontarachna (Acari, Pontarachnidae) from South Africa, with a discussion of genital acetabula in the family. An. Inst. Biol. UNAM Zool., 67: 259-264.
- Li H.-T., Smit H., Zhan B-X., Jin D-C., Guo J-J. 2025. A comprehensive morphological descriptions of three marine water mites (Acari: Hydrachnidia: Pontarachnidae) from Hainan Island, China. Acarologia 65(3): 786-814. https://doi.org/10.24349/jiun-ayx5
- Montes-Ortiz L., Goldschmidt T., Vásquez-Yeomans L., Elías-Gutiérre M. 2021. A new species of Litarachna Walter, 1925 (Acari: Hydrachnidia: Pontarachnidae) from Corozal Bay (Belize), described based upon morphology and DNA barcodes. Acarologia, 61(3): 602-613. https://doi.org/10.24349/r7no-Ludg
- Pešić, V. 2013a. Pontarachnid mites from marine interstitial, with a description of three new species from South Korea (Acari: Hydrachnidia: Pontarachnidae). Zootaxa, 3701(1): 83-92. https://doi.org/10.11646/zootaxa.3701.1.7
- Pešić, V. 2013b. A new marine water mite species (Acari, Hydrachnidia, Pontarachnidae) from a coastal lake in Southeast Madagascar. Mar. Biol. Res., 9: 312-315. https://doi.org/10.1080/17451000.2012.739697
- Pesic V. & Smit H. 2009. New records of water mites (Acari: Hydrachnidia) from Tasmania, with description of three new species. Zootaxa, 2070: 53-62. https://doi.org/10.11646/zootaxa.2070.1.2
- Pešić V., Chatterjee T., Chan BKK., Ingole B. 2008. Marine water mites (Acari: Hydrachnidia: Pontarachnidae) from Taiwan, Korea and India, with the first description of the male of Pontarachna australis Smit, 2003. Syst. Appl. Acarol., 13: 70-74. https://doi.org/10.11158/saa.13.1.8
- Pešić V., Chatterjee T., De Troch M., Ingole B. 2009. New records of marine water mites (Acari: Hydrachnidia: Pontarachnidae) from the Pujada Bay (west Pacific Ocean) and the Andaman Sea (Indian Ocean). Acta Entomol. Serbica, 14(11): 129-132.
- Pešić V., Chatterjee T., Ingole B., Velip D., Pavićević A. 2012. A new species of Litarachna Walter, 1925 (Acari: Hydrachnidia) from the West Indian Coast, with a discussion on the diversity of the family Pontarachnidae Koenike, 1910. Cah. Biol. Mar., 53: 547-553.
- Pešić V., Smit H. 2016. On the identity of Litarachna divergens Walter, 1925 (Acari, Hydrachnidia: Pontarachnidae), with description of one new species. Mar. Biodivers., 46(1): 51-57. https://doi.org/10.1007/s12526-015-0316-x
- Pešić V., Durucan F., Zawal A. 2019. Marine mites (Acari: Hydrachnidia) of the Mediterranean Sea: Descriptions of two new species, key for identification and future prospects. Zootaxa, 4585(3): 501-516. https://doi.org/10.11646/zootaxa.4585.3.6
- Pešić V., Chatterjee T., Schizas NV. 2020. A new marine mite of the genus Litarachna Walter, 1925 from Guadeloupe, Caribbean Sea (Acari, Hydrachnidia, Pontarachnidae). Ecolog. Montenegrina, 38: 205-209. h https://doi.org/10.37828/em.2020.38.28
- Smit H. 2003. Five new species of the water mite family Pontarachnidae from Western Australia (Acari: Hydrachnidia). In: Wells FE,, Walker DI,, Jones DS, (Eds.), The Marine Flora and Fauna of Dampier, Western Australia 2. Western Australian Museum, Perth, pp. 547-562.
- Smit H. 2007. Litarachna brasiliensis n. sp., the first member of the water mite family Pontarachnidae (Acari: Hydrachnidia) from South America. Syst. Appl. Acarol., 12: 141-146. https://doi.org/10.11158/saa.12.2.8
- Smit H., Alberti G. 2010. The water mite family Pontarachnidae, with new data on its peculiar morphological structures (Acari: Hydachnidia). In: Sabelis M., Bruin, J. (Eds) Trends in Acarology. Springer Verlag, Amsterdam, pp. 71-79. https://doi.org/10.1007/978-90-481-9837-5_11
- Uchida T. 1935. Marine water mites from Japan. J Fac Sci Hokkaido Univ, Sapporo (s. 6), Zool., 4(4): 183-195.
- Wiles PR., Chatterjee T., De Troch M. 2002. Two new and one known marine water mite (Acari: Hydrachnidia: Pontarachnidae) from South-East Africa. J. Nat. Hist., 36: 1987-1994. https://doi.org/10.1080/00222930110068583
- Willis TJ., Handley SJ., Chang FH., Law CS., Morrisey DJ., Mullan AB., Pinkerton M., Rodgers KL., Sutton P.H., Tai A. 2007. Climate change and the New Zealand marine environment. NIWA Client Report for the Department of Conservation: NEL2007-025 October 2007, NIWA Project: DOC08305.



2025-10-24
Date accepted:
2026-02-13
Date published:
2026-02-19
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2026 Pešić, Vladimir; Batista-Ribeiro, Dante; Pepato, Almir R. and Smit, Harry
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)