



Description of immatures and ontogenetic development of Africoseiulella flechtmanni Kreiter, type species of the enigmatic family Africoseiulellidae (Acari: Mesostigmata: Phytoseioidea)
Ferragut, Francisco  1
1
1✉ Instituto Agroforestal Mediterráneo. Universidad Politécnica de Valencia. Camino de Vera, s/n. 46022 Valencia, Spain.
2025 - Volume: 65 Issue: 4 pages: 1285-1301
https://doi.org/10.24349/4q3d-f3ezZooBank LSID: C00E4863-F9FA-43F2-B70C-B36A7CA76FEF
Original research
Keywords
Abstract
Introduction
Africoseiulella flechtmanni Kreiter, a peculiar mesostigmatic mite, was first described from an unidentified low shrub in a palm growing inside the Tunisian desert (Kreiter & Tixier, 2006). Aware of the taxon´s originality, the authors erected the new genus Africoseiulella Kreiter to accommodate it, and paid particular attention to its taxonomic position within the family Phytoseiidae. They concluded that it is probably a member of the tribe Metaseiulini Chant & McMurtry in the subfamily Typhlodrominae, although they expressed doubts about this placement. Later, El-Banhawy et al. (2025) described a second species in the genus, A. sedishazlensis El-Banhawy, Nasr & Ramadan, from Egypt, and Mirza et al. (2025) transferred the previously described Nabiseius arabicus Negm & Alatawi to Africoseiulella. Recently, Kazemi et al. (2025) described a fourth species, A. hajiqanbari Kazemi, Klompen & Faraji, whose females were collected from the house cricket Acheta domesticus (L) in Iran, apparently displaying phoretic behaviour. These latter authors conducted a thorough comparative study of the most significant characters of the genus (the reduction in the number of leg tarsi II–IV and palp setae, the undivided palp tarsus apotele, and the presence of a membranous lobe associated with the movable digit of chelicera) among the families of Phytoseioidea. They concluded that the absence of basitarsal setae al3 and pl3 on tarsi II–IV, of ad2 on tarsus IV, the undivided palp tarsal apotele and the lobe attached to the cheliceral movable digit are unique and distinctive features in the superfamily Phytoseioidea and very rare within the whole order Mesostigmata. After that, they redefined Africoseiulella, removed it from Phytoseiidae, and established a new family, Africoseiulellidae Kazemi & Klompen, to accommodate the genus.
The type species of the genus, A. flechtmanni, was originally described from females, males, and deutonymphs, and later redescribed by Kazemi et al. (2025) using the same type material. In both cases the deutonymphal stage was not completely characterized, probably due to the bad condition of specimens, and the larva and protonymph were, still, undescribed. In a survey in the Canarian Island of Fuerteventura I had the opportunity to collect an abundant sample of this species in a semidesert environment, within the bed of a dry ravine, which geomorphologically resembles the North African wadis. The sample included all motile developmental stages, which allowed for a complete description of immatures and the documentation of the changes occurring during development in the appearance of idiosomal, leg and palp setae, as well as the cuticular structures, pores and poroids. Hence, this is the first study on the ontogeny of this species and family. The complete ontogenetic description will enable further research to clarify the taxonomic affinities within the subfamily Phytoseioidea and elucidate the independence of characters in the absence of ontogenetic correlations among them.
Material and methods
The mites were collected by the author in Fuerteventura Island and extracted from plants beating twigs and leaves on a fine sieve. Specimens were mounted and clarified in Heinze-PVA medium and stored in an oven at 50 °C until total clarification. After that, they were examined using differential interference contrast (DIC) in a compound microscope Nikon Eclipse Ni-U (Nikon Corporation, Tokyo, Japan). Images were captured with a digital camera Nikon DS-Fi3 (Nikon Corporation, Tokyo, Japan) and edited in Lightroom and Photoshop CC (© Adobe Systems Inc.). Measurements of morphological traits were obtained with the software NIS-Elements D 3.1 and are given in the text in micrometres (μm). They are represented as the range of all the specimens measured. The length of the first cheliceral segment was taken from its base to the base of the second segment; the second segment was measured from the base to the apex of the fixed digit. The cheliceral fixed digit length was measured from dorsal lyrifissure to the tip; the movable digit from basal articulation to tip. The length of adult legs was obtained from the base of the coxa to the end of tarsus, excluding the ambulacrum.
The generic concept is that proposed by Kreiter (in Kreiter and Tixier, 2006), recently modified by Kazemi et al. (2025). Idiosomal setal nomenclature follows that of Lindquist and Evans (1965) as adapted by Rowell et al. (1978). Leg and palp chaetotaxy is that proposed by Evans (1963; 1964) and Rowell and Chant (1979). Nomenclature for idiosomal solenostomes and poroids follows Athias-Henriot system (Athias-Henriot, 1975) later modified by Moraza (2025).
Results
Family Africoseiulellidae Kazemi & Klompen, 2025
Genus Africoseiulella Kreiter, 2006
Africoseiulella flechtmanni Kreiter, 2006
Specimens examined and depository
Twenty-two females, nine males, eight deutonymphs, three protonymphs, two larvae on Heliotropium ramosissimum (Boraginaceae) and four females on Ononis hesperia (Fabaceae); Tindaya, Barranco de Esquinzo, 28º37′47″N, 14º00′18″W, 59 m a.s.l. (above sea level); 26 May 2024. Three females and one deutonymph on Suaeda sp. (Amaranthaceae); El Cotillo, Aljibe de la Cueva beach, 28º39′53″N, 14º00′41″W, 18 m a.s.l.; 27 May 2024. Both localities in the island of Fuerteventura, Canary Islands.
Eleven females, one male, and four deutonymphs were deposited in the Museo de Ciencias Naturales de Tenerife, Santa Cruz de Tenerife, Canary Islands. The remaining specimens are in the Acari collection, Instituto Agroforestal Mediterráneo, Universidad Politécnica de Valencia.
Geographical distribution
Described from a palm oasis in the Tunisian desert. This is the second record of the species.
Description of motile immature stages
Larva (two larvae examined and measured. Figures 1 a–b, 2 a–c)
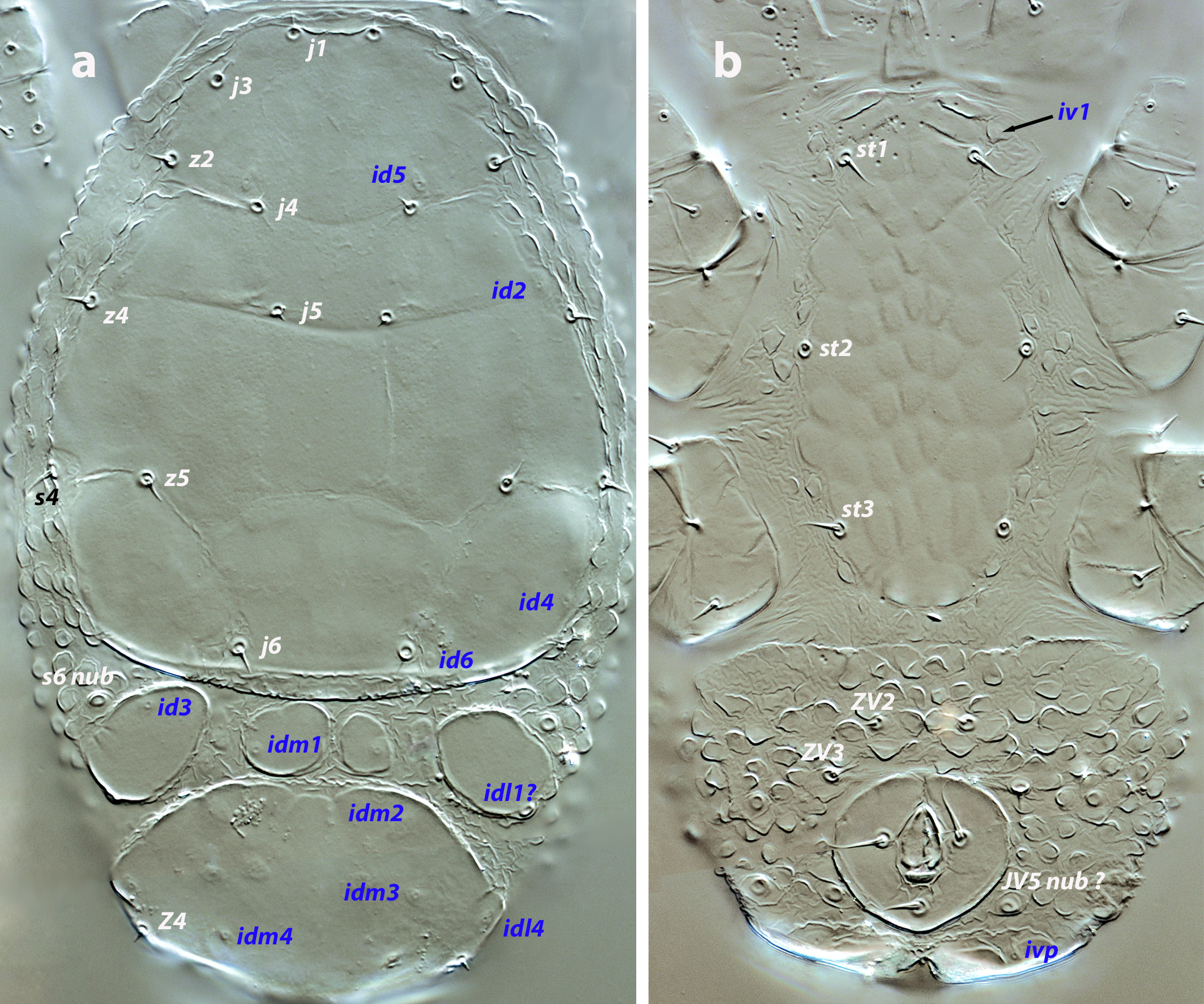
Dorsal and lateral idiosoma —(Figure 1a). Podonotal shield 121–123 long, 96–99 wide at level of s4. Seven pairs of short and pointed setae: j1, j3, j4, j5, j6, z2, z5 on the shield, z4, s4 off the shield. Shield evenly smooth, bearing poroids id2, id4, id5, and id6. Soft integument around the shield forming pebble protuberances. Mesonotal region with three-four shields (mesonotal scutella) arranged as follows: two lateral shields with poroids id3 and idl1?, and a median shield which can be single or divided into two, bearing poroids idm1. Pygidial shield (44 long, 42–45 wide) bearing the setae Z4 and the poroids idm2, idm3, idm4, and idl4. Lengths of j1 6, j3 5–6, j4, j5 4, j6 6, z2, z4, z5 5–6, Z4 4–5, s4 5.
Ventral idiosoma — (Figure 1b). Total length of tritosternum 57–60, distally divided into two laciniae, 25–29 long; each lacinia with just 2–3 barbs. Sternal shield 85–90 long, 39–43 wide at level of setae st2. Setae st1–st3 present; poroid iv1 off the shield. Surface of the shield ornamented, with piriform or elliptical cells oriented axially. Soft integument between the sternal and anal plates with protuberances and folds, scarcer and more isolated than on the dorsum. A pair of subcuticular ''funicules» (sensu Athias-Henriot, 1971) anterior to the anal plate. Anal plate 24–27 long, 36 wide. Setae ZV2 and ZV3, some setal nubs (JV5?), and poroid ivp surrounding the anal valves. Setae ZV3 twice as long as ZV2, 6–7 and 3 long, respectively. Cribrum not developed.
Gnathosoma — Base of gnathosoma 35–37 long, 37–41 wide. Anterior margin of tectum straight, not extended forward. First cheliceral segment 7-10; second segment 47–50 long. Hypostomal setae h1 8–9, h2 5. Fixed digit of chelicera 9 long, movable digit 8–9 long; dentition of both digits undiscernible. Chaetotactic formula of palps (from trochanter to tarsus): 0-4-5-12-11. Anterolateral setae on palp femur and genu thickened, but not expanded distally. Apotele conical, undivided, 7 long.
Download as
Stage
Podonotum
Opisthonotum
Intercoxal region
Opisthogastric region
L
j1, j3, j4, j5, j6, z2, z4, z5, s4
Z4
st1, st2, st3
ZV2, ZV3, pa1, pa2, ps
PN
j1, j3, j4, j5, j6, z2, z4, z5, s4, s6, r3
J2, J5, Z4, Z5, S2, S5
st1, st2, st3
JV2, JV5, ZV2, ZV3, pa1, pa2, ps
DN
j1, j3, j4, j5, j6, z2, z3, z4, z5, s4, s6, r3
J2, J5, Z4, Z5, S2, S5
st1, st2, st3, st4,st5
JV2, JV5, ZV2, ZV3, pa1, pa2, ps
A
j1, j3, j4, j5, j6, z2, z3, z4, z5, s4, s6, r3
J2, J5, Z4, Z5, S2, S5
st1, st2, st3, st4, st5
JV2, JV5, ZV2, ZV3, pa1, pa2, ps
Stage
Gland openings
Dorsal and lateral poroids
Ventral poroids
L
–
id2, id3, id4, id5, id6, idl1, idl4, idm1, idm2, idm3, idm4
iv1, ivp
PN
gp, gvi, gv3
id2, id3, id4, id5, id6, idl1, idl2, idl3, idl4, idl5, idm1, idm2, idm3, idm4, idm5,ip
iv1, iv2, ivo1, ivo2, ivp
DN
gp, gvi, gv3
id1, id2, id3, id4, id5, id6, idl1, idl2, idl3, idl4, idl5, idm1, idm2, idm3, idm4, idm5, ip
iv1, iv2, iv3, iv5, ivo1, ivo2, ivp, ip
A
gp, gvi, gv3
id1, id2, id3, id4, id5, id6, idl1, idl2, idl3, idl4, idl5, idm1, idm2, idm3, idm4, idm5, ip
iv1, iv2, iv3, iv5, ivo1, ivo2, ivp, ip
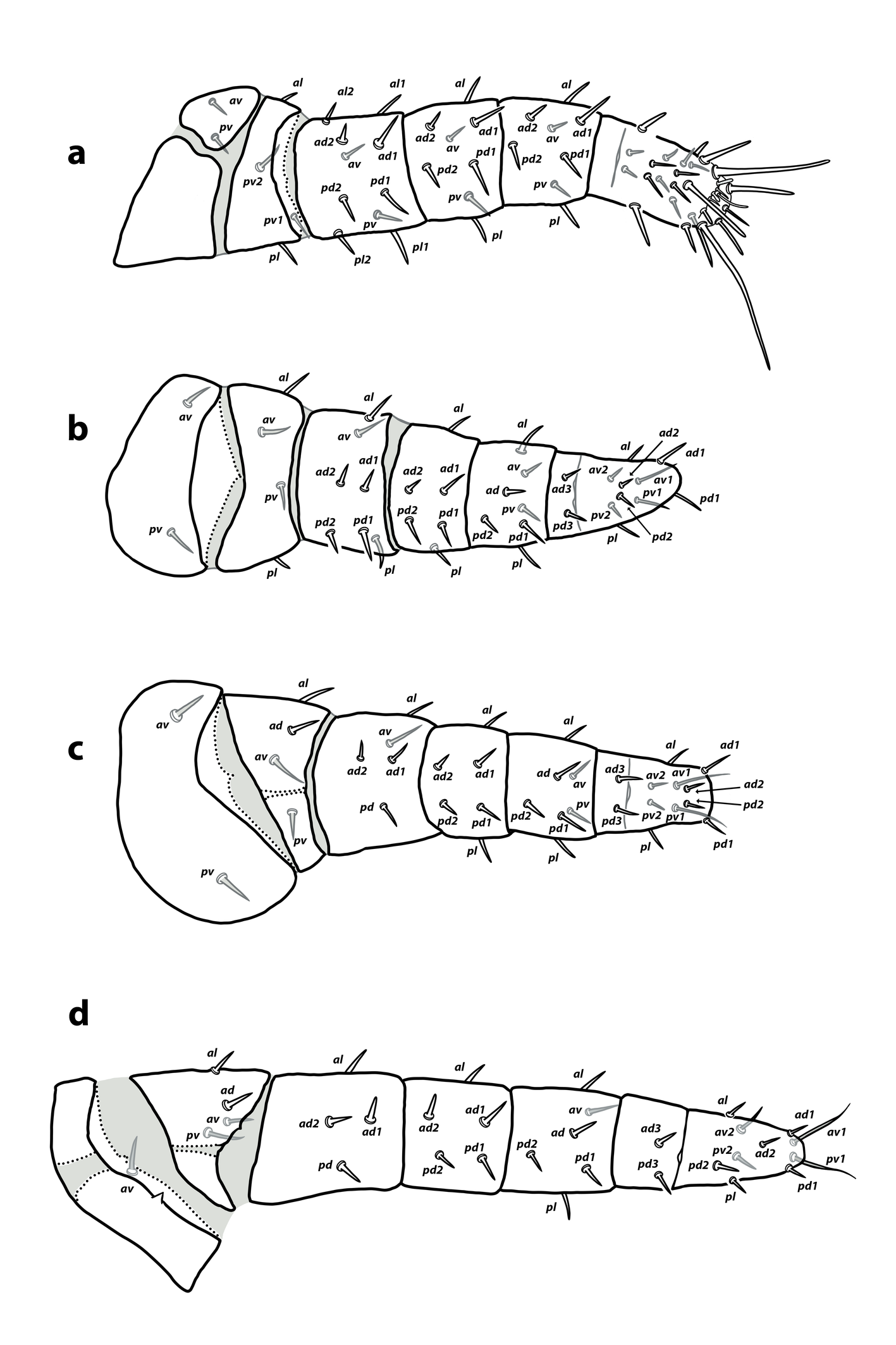
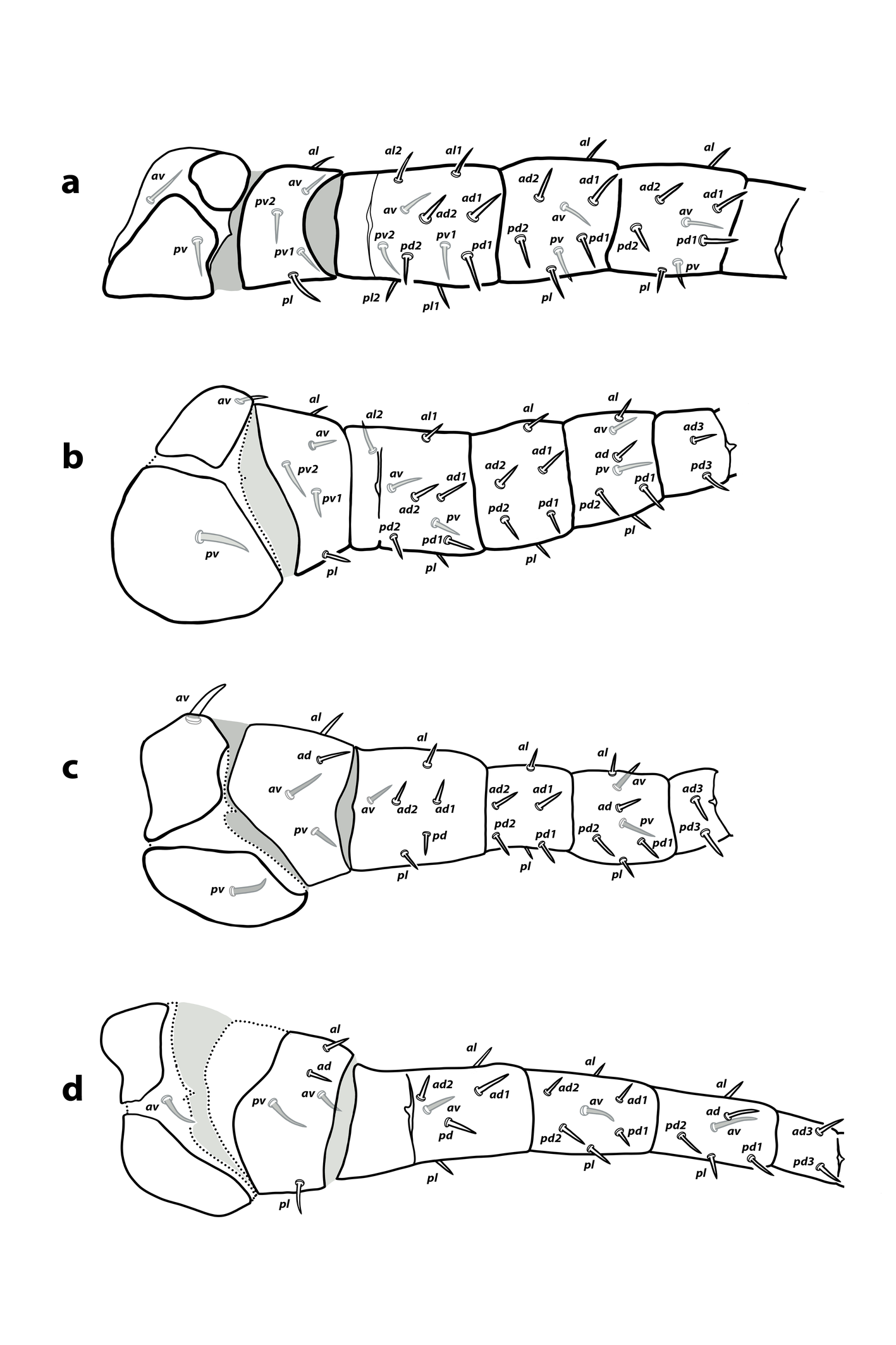
Legs — (Figure 2 a–c). Coxa of leg I with discontinuous sclerotization at the dorsal side. Interruptions of sclerotization not observed on other coxae or leg segments. Setae on legs short, smooth and acuminate, except the longer ventral setae on tarsi I–III. Chaetotaxy of leg segments I–III: coxae 2-2-2; trochanters 4-4-4; femora 10-7-5; genua 8-6-6; tibiae 8-7-7; basitarsi 0-2-2. Detailed leg chaetotaxy in Table 2. Setal formulae from trochanter to basitarsus as follows. Leg I: trochanter 1 0/0, 0/2 1; femur 2 2/1, 2/1 2; genu 1 2/1, 2/1 1; tibia 1 2/1, 2/1 1. Leg II: trochanter 1 0/1, 0/1 1; femur 1 2/1, 2/0 1; genu 1 2/0, 2/0 1; tibia 1 1/1, 2/1 1; basitarsus 0 1/0, 1/0 0. Leg III: trochanter 1 1/1, 0/1 0; femur 1 2/1, 1/0 0; genu 1 2/0, 2/0 1; tibia 1 1/1, 2/1 1; basitarsus 0 1/0, 1/0 0. Without macrosetae.
Download as
Leg
Stage
Coxa
Trochanter
Femur
Genu
Tibia
Basitarsus
I
L
av, pv
al, pl, pv1, pv2
al1, al2, ad1, ad2, av, pl1, pl2, pd1, pd2, pv
al, ad1, ad2, av, pl, pd1, pd2, pv
al, ad1, ad2, av, pl, pd1, pd2, pv
–
PN
av, pv
al, pl, pv1, pv2
al1, al2, ad1, ad2, av, pl1, pl2, pd1, pd2, pv
al, ad1, ad2, av, pl, pd1, pd2, pv
al, ad1, ad2, av, pl, pd1, pd2, pv
–
DN
av, pv
al, av, pl, pv1, pv2
al1, al2, ad1, ad2, av, pl1, pl2, pd1, pd2, pv1, pv2
al, ad1, ad2, av, pl, pd1, pd2, pv
al, ad1, ad2, av, pl, pd1, pd2, pv
–
A
av, pv
al, av, pl, pv1, pv2
al1, al2, ad1, ad2, av, pl1, pl2, pd1, pd2, pv1, pv2
al, ad1, ad2, av, pl, pd1, pd2, pv
al, ad1, ad2, av, pl, pd1, pd2, pv
–
II
L
av, pv
al, av, pl, pv
al, ad1, ad2, av, pl, pd1, pd2
al, ad1, ad2, pl, pd1, pd2
al, ad, av, pl, pd1, pd2, pv
ad3, pd3
PN
av, pv
al, av, pl, pv
al, ad1, ad2, av, pl, pd1, pd2
al, ad1, ad2, pl, pd1, pd2
al, ad, av, pl, pd1, pd2, pv
ad3, pd3
DN
av, pv
al, av, pl, pv1, pv2
al1, al2, ad1, ad2, av, pl, pd1, pd2, pv
al, ad1, ad2, pl, pd1, pd2
al, ad, av, pl, pd1, pd2, pv
ad3, pd3
A
av, pv
al, av, pl, pv1, pv2
al1, al2, ad1, ad2, av, pl, pd1, pd2, pv
al, ad1, ad2, pl, pd1, pd2
al, ad, av, pl, pd1, pd2, pv
ad3, pd3
III
L
av, pv
al, ad, av, pv
al, ad1, ad2, av, pd
al, ad1, ad2, pl, pd1, pd2
al, ad, av, pl, pd1, pd2, pv
ad3, pd3
PN
av, pv
al, ad, av, pv
al, ad1, ad2, av, pd
al, ad1, ad2, pl, pd1, pd2
al, ad, av, pl, pd1, pd2, pv
ad3, pd3
DN
av, pv
al, ad, av, pv
al, ad1, ad2, av, pl, pd
al, ad1, ad2, pl, pd1, pd2
al, ad, av, pl, pd1, pd2, pv
ad3, pd3
A
av, pv
al, ad, av, pv
al, ad1, ad2, av, pl, pd
al, ad1, ad2, pl, pd1, pd2
al, ad, av, pl, pd1, pd2, pv
ad3, pd3
IV
PN
av
al, ad, av, pv
al, ad1, ad2, pd
al, ad1, ad2, pd1, pd2
al, ad, av, pl, pd1, pd2
ad3, pd3
DN
av
al, ad, av, pl, pv
al, ad1, ad2, av, pl, pd
al, ad1, ad2, av, pl, pd1, pd2
al, ad, av, pl, pd1, pd2
ad3, pd3
A
av
al, ad, av, pl, pv
al, ad1, ad2, av, pl, pd
al, ad1, ad2, av, pl, pd1, pd2
al, ad, av, pl, pd1, pd2
ad3, pd3
Protonymph (three protonymphs examined and measured. Figures 2 d; 3 a–b; 8 h)
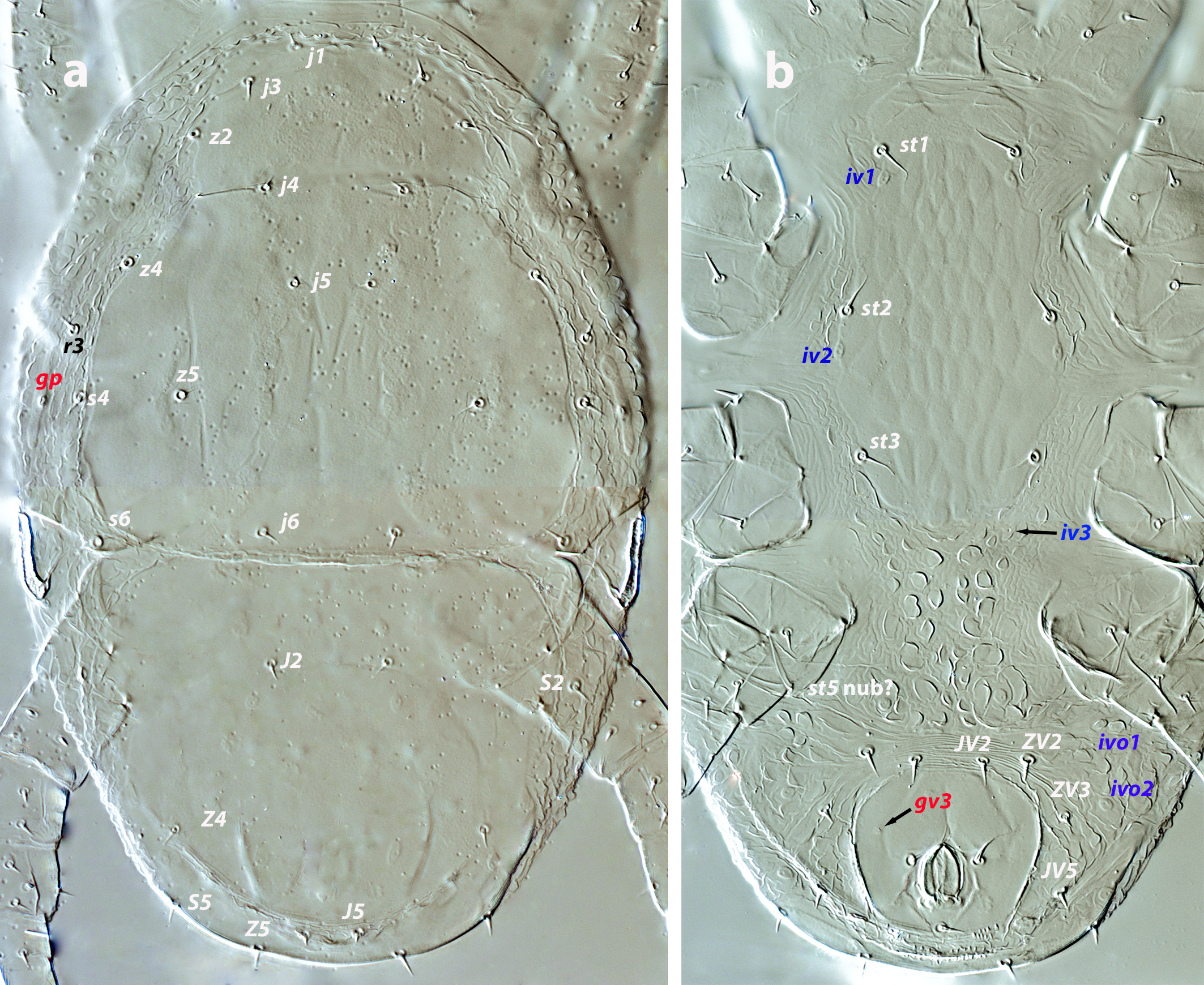
Dorsal and lateral idiosoma — (Figure 3a). Podonotal shield 121–133 long, 117–122 wide at level of s4. Seven pairs of short and pointed setae on the shield: j1, j3, j4, j5, j6, z2, z5; z4, s4, s6 and r3 off the shield. Surface of the shield smooth but showing the characteristic punctate pattern, with poroids id2, id4, id5, and id6. Pygidial shield 89–102 long, smooth, with five pairs of setae (J2, J5, Z4, Z5, S5) and nine pairs of poroids (id3, idm1, idm2, idm3, idm4, idm5, idl2, idl3, idl4); setae S2 and poroids idl1 and idl5 on the lateral integument. Podonotal and pygidial shields contiguous, without soft cuticle between them or mesonotal scutella. Lengths of j1, j3 5–6, j4, j5 3–4, j6, J2 4–5, J5 3, z2, 5–6, z4 6, z5, Z4 4–5, Z5 6, s4 6–7, s6 6, S2, S5 5–6. Setae r3 4–5. Peritremes 19–21 long, covered internally by microvilli, with a posterior extension bearing the gland pore gvi (gdp in Athias-Henriot´s system). Primordial peritrematal shield anterior and separated from the peritreme, 19–21 long, not reaching the level of setae r3 and bearing the solenostome gp (gd3 in Athias-Henriot´s system) and the poroid ip.
Ventral idiosoma — (Figure 3b). Tritosternum 62–66 long, its base 14 wide, with two laciniae barely barbed, 36–43. Sternal shield 90–99 long, 46–47 wide at level of setae st2. Setae st1–st3 and poroids iv1 and iv2 present. Shield ornamented with a pattern similar to that of larva. Anterior and lateral soft integument folded, posterior integument with protuberances. Preanal pores gv3 present, but hardly discernible. Region surrounding the anal plate transversally striated or folded, with setae JV2, ZV2, ZV3, JV5, and poroids ivo1, ivo2, and ivp. Setae JV2, ZV2, and ZV3 similar in length (6–7 long).
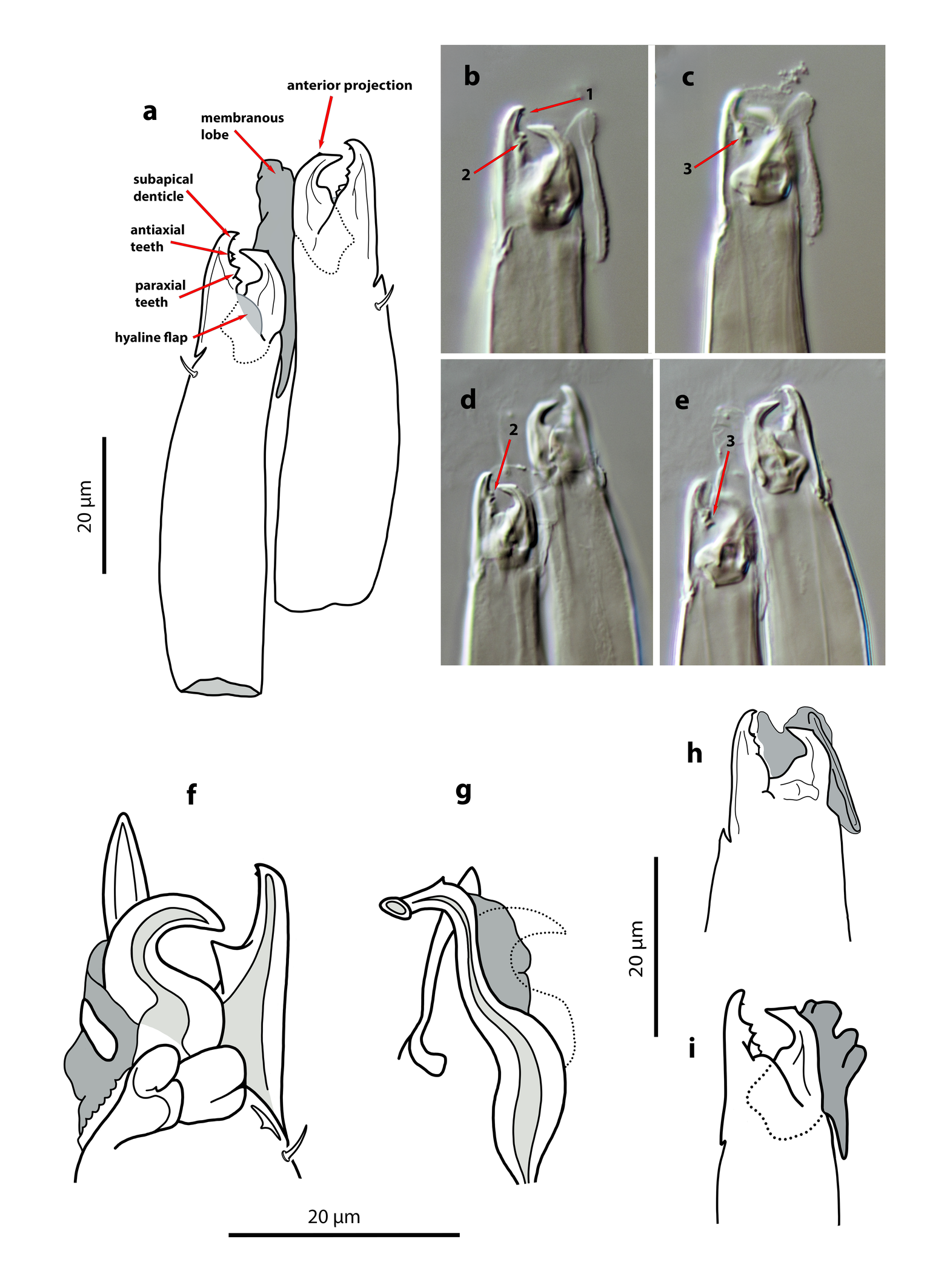
Gnathosoma — (Figure 8h). Base of gnathosoma 55–63 long, 51–65 wide. Anterior margin of gnathotectum similar in shape but smaller than in adult. Deutosternal channel with ten rows of single and median spicules. First cheliceral segment 7–10; second segment 47–50 long. Setae h1 7–9, h2 4–5, h3 5–6, pc 6. Fixed digit of chelicera 10–12 long, with two small teeth in the middle and two at the base of the digit, subapical denticle undiscernible; movable digit 8–9 long, edentate. Without pilus dentilis. Large membranous lobe attached to movable digit. Chaetotactic formula of palps (trochanter to tarsus): 1-4-5-12-12. Setae al on palpfemur and palpgenu acicular. Apotele conical, 7–8 long.
Legs — (Figure 2d). Sclerotization of coxae I–II and trochanter III interrupted; not as clear in other segments due to the poor degree of sclerotization. Setal morphology similar to that of larva. Chaetotaxy of leg segments I–IV (Figure 2 d; Table 2): coxae 2-2-2-1; trochanters 4-4-4-4; femora 10-7-5-4; genua 8-6-6-5; tibiae 8-7-7-6; basitarsi 0-2-2-2. Setal formulae of the three first pair of legs similar to those in the larva (Table 2). Setal formula of leg IV (from trochanter to basitarsus): trochanter 1 1/1, 0/1 0; femur 1 2/0, 1/0 0; genu 1 2/0, 2/0 0; tibia 1 1/1, 2/0 1; basitarsus 0 1/0, 1/0 0. Without macrosetae.
Deutonymph (nine deutonymphs examined; five measured. Figures 4 a–b; 5; 8i)
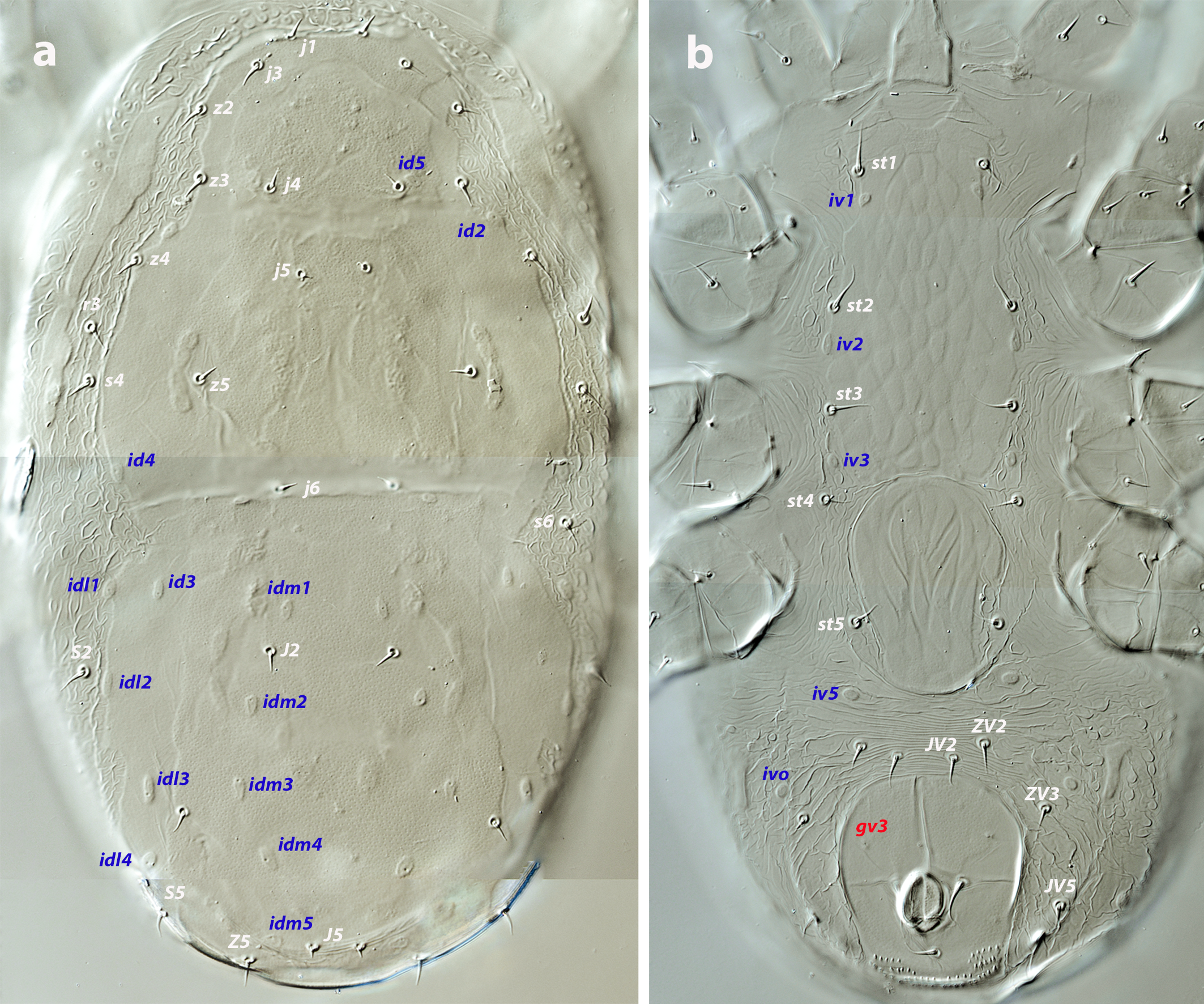
Dorsal and lateral idiosoma — (Figure 4a). Podonotal and pygidial shields fused into a holodorsal shield, 234–246 long, 105–117 wide. Anterolateral margin of dorsal shield between setae z2–z4 concave, with setal insertions and poroid id2 in marginal position. Surface of the shield smooth, finely punctate, apparently more sclerotized on the inner region than on the margins. Absence of solenostomes; poroids sticking out from the surface, elongated; the complete set of sixteen poroids present. All the dorsal setae very short and blunt, subequal in length except the minute J5. Setae s4, s6, S2, and r3 on the soft cuticle. Lengths of j1, j3 5–6, j4, j5, J2 4–5, J5 3, z2, z3 5–6, z4 6–7, z5 5–6, Z4 5–7, Z5 6–8, s4 6–7, s6 6–8, S2, S5 6–7. Setae r3 5–6. Integument around the shield folded, with pebble protuberances. Peritremes short, 23–26, isolated from the anterior part of the peritrematal shield, which ends at the level of setae z2 and is not fused with the dorsal shield.
Ventral idiosoma — (Figure 4b). Tritosternum 66–70 long, with two laciniae, 39–45 long. Sternal shield 90–95 long, 47–50 wide, bearing setae st1–st3 and poroids iv1–iv3; both setae and poroids on the margins. Shield ornamented with a reticulated pattern consisting of elongate and contiguous cells oriented axially. Setae st4 on the soft integument. Inverted tongue-shaped epigynal shield with longitudinal grooves; 56–62 long, 37–44 wide; setae st5 and poroids iv5 off the shield. Anal shield 48–54 long, 42–47 wide, smooth, with preanal pores. Cuticle between epigynal and anal shields transversally striated, mostly plicate on both sides of the anal plate. Setae ZV2, ZV3, JV2, JV5, and poroids ivo1, ivo2, and ivp around the shield. Setae JV5 5–7 long.
Gnathosoma — (Figure 8i). Base of gnathosoma 51–60 long, 43–55 wide. Hypostomal and capitular setae h1 9–11, h2 5–7, h3 6–9, pc 7–9. First cheliceral segment 11–14; second segment 49–58 long. Fixed digit of chelicera 15–18 long, with a small subapical denticle, two teeth at mid-length and two contiguous basal teeth, just anterior to the hyaline flap; movable digit 13–15 long, without dentition; both digits with the same morphology as in female. Pilus dentilis absent. Membranous lobe attached to the movable digit. Palp chaetotaxy from trochanter to tarsus: 2-5-5-12-12.
Legs — (Figure 5)″ Coxae I–IV and, occasionally, trochanters III–IV with interrupted dorsal sclerotization. Setal morphology as in the larva and protonymph. Chaetotaxy of leg segments I–IV: coxae 2-2-2-1; trochanters 5-5-4-5; femora 11-9-6-6; genua 8-6-6-7; tibiae 8-7-7-6; basitarsi 0-2-2-2 (Figure 5; Table 2). Setal formulae from trochanters to basitarsi as follows. Leg I: trochanter 1 0/1, 0/2 1; femur 2 2/1, 2/2 2; genu 1 2/1, 2/1 1; tibia 1 2/1, 2/1 1. Leg II: trochanter 1 0/1, 0/2 1; femur 2 2/1, 2/1 1; genu 1 2/0, 2/0 1; tibia 1 1/1, 2/1 1; basitarsus 0 1/0, 1/0 0. Leg III: trochanter 1 1/1, 0/1 0; femur 1 2/1, 1/0 1; genu 1 2/0, 2/0 1; tibia 1 1/1, 2/1 1; basitarsus 0 1/0, 1/0 0. Leg IV: trochanter 1 1/1, 0/1 1; femur 1 2/1, 1/0 1; genu 1 2/1, 2/0 1; tibia 1 1/1, 2/0 1; basitarsus 0 1/0, 1/0 0. Without leg macrosetae.
Complementary description of adults
Female (twenty-nine females examined; seven females measured. Figures 6 a–c; 8 a–e)
Dorsal shield pyriform, with a characteristic punctate pattern more evident on the podonotum (Figure 6a); 289–322 long, much wider in the opisthosoma (159–179) at level of S2, than on the podosoma (131–143 at level of s4). Lateral margins irregular, always concave at the level of setae z3, s4, and between s4 and s6, and strongly constricted between podosoma and opisthosoma. All the mentioned setae inserted on the shield margin. Sixteen pairs of short dorsal setae. Lengths of j1 6–7, j3 8–9, j4 6–7, j5 4–5, j6, J2 6–8, J5 4–5, z2, z3, z4 8–9, z5 7–8, Z4 6–8, Z5, s4, s6, S2, S5 8–9. Setae r3 7–8; absence of R1. Dorsal solenostomes absent, but with the complete set of sixteen pairs of poroids, large, elliptical, about 7–8 long, most of them longitudinally oriented. Peritrematal shield broadly merged with the dorsal shield between z2 and z3, bearing the gland pore gp and the poroid ip at the anterior level of the peritremes, and the pore gvi on the poststigmatic extension of the shield. Peritremes very short, 40–45 long, reaching anteriorly the level of setae s4 and gland pore gp; internal groove evenly covered by papillae. Base of tritosternum broad, subrectangular, distinctly wider than the combined width of the two laciniae, 15 wide, 12 long; total length of tritosternum 76–81, laciniae, 48–52. Sternal shield elongate and trapezoidal (Figure 6b), without anterior arms, bearing setae st1–st2 and poroids iv1–iv2; 80–86 long, 43–49 wide at the level of st2; anterior margin concave, with two lateral and sclerotized areas transversally striated, posterior margin subtriangular and weakly sclerotized; surface of the shield with a delicate pattern of elongate cells oriented axially. Setae st3–st4 and poroid iv3 free on the soft cuticle. Epigynal shield 106–118 long, covered by longitudinal furrows; posterior margin concave; distance st5–st5 52–56. Ventrianal shield vase-shaped, anterior margin concave (Figure 6c); 82–94 long, 64–69 wide at level of setae ZV2, 56–62 wide at the level of anus. Surface evenly smooth, with two pairs of preanal setae (ZV2, JV2) and a pair of small and punctiform preanal solenostomes gv3, situated posterior and distant to the insertions of JV2, and separated 32–41 μm from each other. Most of spicules of cribrum joined in both laterals, arranged into three-four rows, with a bridge of two rows of spicules posterior to post-anal seta and connecting the two lateral areas. Opisthogastric soft integument with setae ZV3 and JV5, two pairs of poroids ivo and poroid ivp, and a bacillar platelet placed near the anterior corners of the ventrianal shield, in the triangle formed by the setae ZV2, ZV3 and anterior poroid ivo. Setae JV5 8–9, placed at the level of anal opening. A single metapodal shield, very long (40–46) and narrow, with a longitudinal ridge. Major duct of spermatheca 20–28 long, well-sclerotized, cylindrical, with a ring near the calyx; atrium very small, at the same bases of calyx; calyx shallow, 3–4 deep, 9–10 wide near the vesicle. Base of gnathosoma 40–47 long, 52–58 wide. Tectum developed as a narrow projection rounded anteriorly. Subcapitular setae short, h1 9–11, h2 6–7, h3 8–9, pc 9–11. Fist cheliceral segment 14–19, second segment 60–65. Fixed digit of chelicera straight, 16–18 long, with a small subapical denticle, two median teeth, and two basal teeth, pilus dentilis not discerned in the examined specimens; movable digit curved, with a broad base abruptly reduced in width at the middle and ending into a sharp and angled hook, 13–15 long, without teeth (Figures 8 a–e); a large and hyaline lobe attached to the movable digits and a hyaline flap at the base of fixed digit (Figure 8a). Palp 53–60 long, setation as in deutonymph. Palpfemur with setae al blunt, not expanded distally; palpgenu without al2. Palptarsus apotele undivided.

Legs — Short; lengths of leg I 152–169, leg II 126–148, leg III 136–152, leg IV 163–177. Coxa II with seta pv more than twice as long as seta av; seta av on coxa III spur-like. Leg chaetotaxy of segments I–IV and setal formulae similar to deutonymph. Without differentiate macrosetae.
Male (nine males examined; five males measured. Figures 7 a–c, 8 f–g)
Dorsal shield suboval, 242–255 long, 123–138 wide, attached to the peritrematal shield at level of setae z4. Shield surface finely punctate, but porous ornamentation typical of the female not visible (Figure 7a). Rectangular unsclerotized area inside the shield, between setae z2 and z4, bearing in its internal margin the seta z3 and the poroid id2. This area corresponds to the marginal concavity between z2–z4 in the female shield and results from the merging of the male peritrematal and dorsal shields. Sixteen pairs of short dorsal setae. Lengths of j1 6–7, j3 7–8, j4 5–6, j5 4–5, j6 6–7, J2 5–6, J5 3–4, z2 7–8, z3 8, z4 7, z5 6–7, Z4 6–7, Z5 and s4 8, s6 7–8, S2 and S5 8, r3 6–7. Setae R1 absent, r3 always off the shield. Peritremes short, reaching the level of pore gp on the peritrematal shield and almost the insertions of s4 on the dorsum. Tritosternum relatively short, with a broad base 14 wide, 10 long, divided into two laciniae; total length of tritosternum 58, of the laciniae 40–42. Sternogenital shield elongate (Figure 7b), anterior margin concave. Poroid iv1 posterior and close to the insertions of st1, poroid iv2 free on the soft integument between st3 and st4. Lateral margins invaginated at the level of st4; posterior margin rounded. Setae st1–st3 on the margin of the shield, st4–st5 off the shield. Surface mostly smooth, but in the posterior part it is punctate and forms five pairs of areolae of different size arranged in a characteristic pattern. Distance st1–st5 113–126, st3–st3 36–41. Poroids iv5 between the sternogenital and ventrianal shields. Ventrianal shield divided into three plates (Figure 7c), the central plate being larger and columnar in shape, bearing the setae ZV2 and JV2, the small and punctiform solenostomes gv3 posteroantiaxial to JV2 (34–38 between them), the anal opening, and the para-anal and postanal setae. The lateral plates bear one poroid ivo and, sometimes, the setae ZV3. Setae JV5 in posterolateral position in relation to the shield, somewhat anterior to the large poroid ivp. The anterior part of the central plate is transversally striated and both lateral plates have longitudinal striae. Although the lateral shields were previously reported attached to the central plate, this character is variable, as some of the males examined have the three plates separated (as can be seen in Figure 7c). Cribrum as in females. All the opisthoventral setae short o very short; JV5 6–7 long. Fixed digit of chelicera straight, distally attenuated,17–18 long, with a wide base and a strong median tooth, apparently with a minute subapical tooth; movable digit 15–17, edentate, with a broad and rounded bases and a strongly angled hook (Figure 8f). Spermatodactyl sinuous, with a broad shaft 13–15 long, a short foot 5–7, and a small and pointed lateral process (Figure 8g). Vellum present. Most of leg setae very short and pointed, some of them clearly subconical, spur-like; e.g., seta av on coxa I, pv on coxa II, and av, pv, on coxa III. Seta v on coxa IV setaceous. Leg chaetotaxy of segments I–IV and setal formulae similar to deutonymph. Without macrosetae.

Remarks
At first glance, A. flechtmanni is a small, slow-moving mite with a fusiform idiosoma and short legs. It was found at high densities inside the inflorescences of Heliotropium ramosissimum, a plant native to Tunisia, Morocco, and the Macaronesian islands (Sauerbier et al., 2023). This particular morphology may represent an adaptation to life in highly confined spaces. Because no other microarthropods were observed on the plants, the mite presumably feeds on plant resources, most likely pollen.
In their comparative study of the genus, Kazemi et al. (2025) highlighted both the shared and distinctive morphological traits between Africoseiulella and other phytoseioid families, supporting their decision to establish a new family. Examination of the immature stages of A. flechtmanni provides further insights and additional arguments reinforcing the distinction between Africoseiulellidae and Phytoseiidae. Although descriptions of phytoseiid immatures are still the exception rather than the rule, available data indicate that larvae are generally weakly sclerotized and that the outlines of dorsal shields are often poorly defined. By contrast, the larva of A. flechtmanni possesses well-sclerotized dorsal shields and, remarkably, two pairs of large mesonotal scutella, structures that have never been described in phytoseiids (for a list of papers describing immature phytoseiids up to 2017, see Ma et al., 2018). The occurrence of larval mesonotal scutella has been reported in species from other phytoseioid families, such as the Blattisociidae Lasioseius allii Chant (illustrated in Lindquist & Evans, 1965), L. corticeus Lindquist, L. serripes Moraza & Lindquist (Lindquist, 1971; Moraza & Lindquist, 2018), and in the Treatiidae Hemipteroseius womersleyi Evans (Prasad, 2018). In contrast, the larvae of other members of Treatiidae lack developed scutella between the podonotal and pygidial shields, e.g., Nabiseius duplicisetus Chant & Lindquist and N. palifer Joharchi & Khaustov (Prasad, 2018; Joharchi et al. 2022).
Ontogenetic changes in the development of gnathosomal structures — In the subcapitulum, the hypostomatic setae h1, h2 are present in the larva, with h3 and the capitular cs added in the protonymph, following the common pattern observed in Gamasina (Lindquist and Evans, 1965). Likewise, palp setation from trochanter to genu is consistent with that described by Evans (1964) for the free-living Gamasina with the exception of palpgenu seta al2 which is not expressed in Africoseiulella. One seta v is added at the protonymphal stage on the trochanter, while one seta v2 and a third dorsal seta d3 are incorporated into the trochanter and femur, respectively, in the deutonymph. The chaetome of tibia and tarsus in the larva is 12 and 11 setae, respectively. An additional seta is added to tarsus in the protonymph, and the complement of 12 setae on each of the two distal segments is apparently determined at the protonymphal stage. The apotele is undivided and conical in shape from the larval stage onward.
The examination of the chelicerae has revealed some differences from previous information. This structure was originally described as having an edentate fixed digit (Kreiter & Tixier, 2006), and according to Kazemi et al. (2025) the fixed digit lacks a subapical denticle and bears only a median tooth. However, the presence of a minute denticle near the apex, as well as teeth in both paraxial and antiaxial positions, is evident in the specimens from Fuerteventura (Figure 8 a–e). This figure shows different views of the chelicera in which two contiguous teeth can be observed at mid-length, along with two additional contiguous teeth in a basal position, located just anterior to the hyaline basal lobe. All these structures, particularly the anterior projection and the small subapical tooth, are extremely small and can only be clearly observed at maximum magnification and when the cheliceral digits are fully opened.
The general appearance of the chelicerae and the dentition in the immature stages is similar to that of the female, although the smaller structures can only be distinguished in the deutonymph and it remains unclear whether they are also present in the protonymph. Unfortunately, the positioning of the chelicerae in the larva did not allow observation of the internal dentition. The hyaline lobe associated with the movable digit is present in all mobile stages but I could not identify any structure similar to a pilus dentilis.
Ontogeny of idiosomal and leg setation — All podonotal setae are expressed in the larval stage except for s6 and the lateral r3, which appear in the protonymph, and z3 in the deutonymph (Table 1). All opisthonotal setae occur in the protonymph, with only Z4 present on the larval pygidial shield. This pattern of setal additions coincides with that of phytoseiids, except in the opisthonotum, where phytoseiid larvae exhibit greater variation in the number of setae, ranging from 1 to 5, although Z4 remains the unique constant seta. On the ventral surface, JV2 and JV5 first appear in the protonymph, and st4, st5 in the deutonymph, while the remaining setae are already present at the larval stage.
As pointed out by Kazemi et al. (2025), the adult leg chaetome is reduced in comparison to the common pattern in other mesostigmatic families. By contrast, the leg chaetome in larvae and protonymphs generally matches the phytoseiid pattern, except that the two lateral setae on basitarsi II–IV are absent. There are two discrepancies in notation of setae on trochanter and tibia IV between the redescription by Kazemi et al. (2025) and the specimens I examined. According to the mentioned authors, the adult setal formula of trochanter IV is 1 1/1, 1/1 0, indicating the presence of a posterodorsal seta and the absence of posterolateral seta; however, in my opinion, the correct notation is 1 1/1, 0/1 1, as can also be seen in Figure 26 of the cited publication. Likewise, the reported formula for tibia IV (1 2/1, 1/0 1) should be stated as 1 1/1, 2/0 1, because a second pd seta is clearly in line with the other seta posterodorsal. The reduction of leg setae becomes evident in the deutonymph and remains unchanged in the adults. Compared to the most common pattern in Phytoseiidae (that can be drawn from the literature; e.g., Evans, 1963; Rowell and Chant, 1979; Aponte and McMurtry, 1987; Ma et al. 2018, 2020, 2024; Khaustov, 2020; Döker et al. 2023, 2024), the chaetotaxy of A. flechtmanni includes 15 absences, six of which occur on the first pair of legs. On leg I, the seta ad on trochanter, ad3 on femur, and the setae al2 and pl2 of the genu and tibia are missing; on femur II, seta ad3 is absent; and on trochanter III, seta pv2 is lacking. To these absences must be added those previously mentioned lateral setae on basitarsi II–IV, and seta ad2 on telotarsus IV.
Larva and protonymph shares the same chaetotaxy on the first three leg pairs. There are 11 setal ontogenetic additions on leg segments (excluding telotarsi) (Table 2). The following setae are added to the deutonymphal complement: on trochanter I seta av, on femur I seta pv2; on trochanter II seta pv2, on femur II setae al2 and pv; on femur III seta pl. In the leg IV the setae pl on the trochanter, and av, pl on both femur and genu are added in the moult to the deutonymphal instar.
Ontogenetic development of gland pores and poroids — This species lacks dorsal solenostomes and only has the peritrematal gland pore gp and the ventral solenostomes gvi (associated with the posterior end of the peritrematal shield) and gv3, all of which develop in the protonymphal stage.
The larva possesses a complement of 13 pairs of poroids (Table 1), 11dorsal poroids: id2, id3, id4, id5, id6, idl1, idl4, idm1, idm2, idm3, idm4, and the ventral poroids iv1 and ivp. The anterodorsal poroid id1 first appears in the deutonymph, while idl2, idl3, idl5, idm5, and ip are added in the protonymph. The protonymph also expresses the ventral poroids iv2, ivo1, and ivo2. The deutonymphal stage incorporates the dorsal poroid id1 and the ventral poroids iv3, iv5, and ip. It is not clear whether id1 is first expressed in the deutonymph, where it is distinctly noticeable, or in the protonymph. In the latter stage, the cuticle bearing this poroid appears corrugated in mounted specimens, making observation difficult and uncertain.
The full set of peritrematal pores and poroids is already present in the protonymphal stage, whereas the anterior and posterior portions of the future peritrematal shield remain separated throughout the juvenile active instars. These portions do not fuse with each other, nor with the dorsal shield, until the adult stage is reached.
Acknowledgements
The author was provided with a permission to collect plants by the Cabildo de Fuerteventura (Resolución 1087/2024). I am grateful to Dr. Shahrooz Kazemi for sharing valuable information and unpublished drawings of the fourth species of Africoseiulella collected in Iran and for the collaborative and fruitful exchange of ideas on the genus Africoseiulella. I appreciate the interest with which three anonymous reviewers read and criticized the initial version of the manuscript. Their comments have substantially improved the final result.
References
- Aponte O.R., McMurtry J.A. 1987. Description of the immature and adult stages of Amblyseius colimensis n. sp. (Acari: Phytoseiidae) from Mexico. Acarologia, 28(3): 201-220.
- Athias-Henriot C. 1971. Un progres dans la connaissance de la composition métamérique des gamasides : leur sigillotaxie idiosomale (Arachnida). Bull. Soc. Zool. France, 96: 73-85.
- Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini II. Le rélève organotaxique de la face dorsal adulte (Gamasides protoadeniques, Phytoseiidae). Acarologia, 17(1): 20-29.
- Döker I., Joharchi O., Karut K., Kazak, C. (2023) Description of Typhloseiulus anatolicus sp. nov. and redescription of two new records of Phytoseiidae (Acari: Mesostigmata) from Turkey. Acarologia 63(2): 553-568. https://doi.org/10.24349/r4a9-vy9o
- Döker I., Atchia I., Jose, A., Bolton, S.M. (2024) Complementary description of two Anthoseius De Leon (Acari: Phytoseiidae) species based on their holotypes with new synonyms. Acarologia 64(1): 192-201. https://doi.org/10.24349/gt2k-nkkv
- El-Banhawy E.S.M., Nasr A.K., Ramadan M.M. 2025. Description of a new species of Africoseiulella Kreiter (Acari: Phytoseiidae) from Egypt. Zootaxa, 5633(2): 244-250. https://doi.org/10.11646/zootaxa.5633.2.2
- Evans G.O. 1963. Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bull. Br. Mus. (Nat. Hist.), Zool., 10: 275-303. https://doi.org/10.5962/bhl.part.20528
- Evans G.O. 1964. Some observations on the chaetotaxy of the pedipalps in the Mesostigmata (Acari). Ann. Mag. Nat. Hist., 13(6): 513-527. https://doi.org/10.1080/00222936308651393
- Joharchi O., Khaustov V., Stanyukovich, M. 2022. First record of family Otopheidomenidae Treat (Acari: Mesostigmata) in Central Asia, with description of a new species of Nabiseius Chant & Lindquist. Acarologia 62(2): 431-445. https://doi.org/10.24349/xo13-9sb4
- Kazemi S., Klompen H., Faraji F. 2025. Africoseiulella Kreiter (Acari, Mesostigmata): description of a new species, taxonomic affinities within Phytoseioidea, establishment of a new family, and revised classification of Treatiinae Wainstein. Syst. Appl. Acarol., 30(8): 1474-1496. https://doi.org/10.11158/saa.30.8.9
- Khaustov V. A. (2020) Review of Amblyseius Berlese (Acari: Phytoseiidae) in Western Siberia, Russia. Acarologia 60(4): 769-80. https://doi.org/10.24349/acarologia/20204401
- Kreiter S., Tixier M.-S. 2006. A new genus and species of phytoseiid mites (Acari: Mesostigmata) from southern Tunisia, with discussion of its phylogenetic position. Zootaxa, 1237(1): 1-18. https://doi.org/10.11646/zootaxa.1237.1.1
- Lindquist E.E. 1971. New species of Ascidae (Acarina: Mesostigmata) associated with forest insect pests. Can. Entomol., 103(7): 919-942. https://doi.org/10.4039/Ent103919-7
- Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina. Mem. Entomol. Soc. Canada, 47: 1-64. https://doi.org/10.4039/entm9747fv
- Ma M., Fan Q-H., Zhang Z-Q. 2018. Morphological ontogeny of Amblydromalus limonicus (Acari: Phytoseiidae). Syst. Appl. Acarol., 23(9): 1741-1765. https://doi.org/10.11158/saa.23.9.3
- Ma M., Fan Q-H., Zhang Z-Q. 2020. Neoseiulus kikuyu sp. nov. (Mesostigmata: Phytoseiidae): descriptions of all life stages from New Zealand. Syst. Appl. Acarol., 25(11): 2098-2114. https://doi.org/10.11158/saa.25.11.13
- Ma M., Zhang, K., Fan Q-H., Zhang Z-Q. 2020. Description of ontogenetic changes in the morphology of Amblyseius herbicolus (Chant, 1959) and Amblyseius lentiginosus Denmark & Schicha, 1974 (Acari: Phytoseiidae). Zootaxa 5485(1): 007-037. https://doi.org/10.11646/zootaxa.5485.1.4
- Mirza J.H., Kamran M., Khan E.M., Alatawi, F.J. 2025. First record of Africoseiulella Kreiter (Parasitiformes: Phytoseiidae) with the complementary description of Africoseiulella arabicus (Negm & Alatawi) comb. nov. from Saudi Arabia. Acarologia, 65(3): 815-826. https://doi.org/10.24349/n2ln-2wlc
- Moraza M.L. 2025. Glandular and non-glandular cuticular organs on the idiosoma of Gamasina mites (Acari: Mesostigmata). Persian J. Acarol., 14(1): 151-171. https://doi.org/10.22073/pja.v14i1.86027
- Moraza M.L., Lindquist E.E. 2018. A new species-group with new species of the genus Lasioseius (Acari: Mesostigmata: Blattisociidae) associated with Neotropical hispine beetles in furled leaves of Heliconia. Acarologia, 58(1): 62-98. https://doi.org/10.24349/acarologia/20184227
- Prasad W. 2018. Description of motile immature stages of Hemipteroseius indicus (Krantz & Khot) (Acari: Otopheidomenidae). Persian J. Acarol., 7(1): 1-40. http://doi.org/110.22073/pja.v7i1.32053
- Rowell H.J., Chant D.A. 1979. Observations on the ontogeny of setae in the family Phytoseiidae (Acarina: Gamasina). Can. J. Zool., 57(3): 670-682. https://doi.org/10.1139/z79-080
- Rowell H.J., Chant D.A., Hansell R.I.C. 1978. The determination of setal homologies and setal patterns on the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata). Can. J. Zool., 110: 859-876. https://doi.org/10.4039/Ent110859-8
- Sauerbier H., Cabrera F., Muer, T. 2023. Flora Vascular de Canarias. Publ. Turquesa, La Laguna, Tenerife. 1514 pp.



2025-08-23
Date accepted:
2025-12-05
Date published:
2025-12-15
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2025 Ferragut, Francisco
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)