



The effect of Tetranychus urticae Koch (Acari: Tetranychidae) infestation on some plant nutrients and growth parameters of common bean under laboratory conditions
Türkoğlu, Huriye  1
; Çetin, Hüseyin
2
and Elma, Fatma Nur
3
1
; Çetin, Hüseyin
2
and Elma, Fatma Nur
3
1Department of Plant Protection, Faculty of Agriculture, Selcuk University, Konya, Türkiye.
2Department of Plant Protection, Faculty of Agriculture, Selcuk University, Konya, Türkiye.
3✉ Department of Plant Protection, Faculty of Agriculture, Selcuk University, Konya, Türkiye.
2024 - Volume: 64 Issue: 4 pages: 1175-1181
https://doi.org/10.24349/aa26-lh0hOriginal research
Keywords
Abstract
Introduction
Common bean (Phaseolus vulgaris L.) is one of the most important food legumes and widely grown crops in the world. It is a valuable source of vegetable protein, characterized by low fat content, adequate levels of carbohydrates and proteins, and richness in essential minerals such as potassium, calcium, magnesium, and phosphorus (Akçin 1988). Common bean can be grown almost anywhere in Turkey (Ülker and Ceyhan 2008). Various pests affect bean production, among them the two-spotted spider mite, Tetranychus urticae Koch, is a polyphytophagous species and is one of the most important cosmopolitan pests of many economically important crops cultivated both in greenhouses and open fields (Elma and Alaoğlu 2008; Polat and Kasap 2011; Çobanoğlu and Kumral 2014). This mite can cause injuries to plants leading to significant yield losses and deteriorataion in the quality of crop production (Abdel-Wali et al. 2012; Tehri et al. 2014). Due to its short life cycle, arrhenotokous parthenogensis reproduction and high fecundity, T. urticae can develop quickly resistance to several classes of pesticides (Whalon et al. 2008; Atalay and Kumral 2013). Given the increasing negative impact of chemical control methods on both the environment and human health, research into alternative control strategies for T. urticae has gained prominence in recent years (Erdoğan 2016; Mammadova et al. 2023).
Understanding the effect of mite infestation on host plant nutrient content can provide valuable data for developing sustainable and effective integrated pest management strategies. Numerous studies have examined the effects of plant nutrients on mite feeding and development (Kytö et al. 1996; Harris et al. 1998; Ribeiro et al. 2012; Nansen et al. 2013). For example, Ribeiro et al. (2012) demonstrated that potassium application on strawberry plants infested with T. urticae led to a decline in mite populations.
However, few studies have investigated the reciprocal impact of mite infestation on the chemical composition of host plants. This study addresses a gap in research by investigating the impacts of T. urticae infestation on nutrient levels in common bean plants, a topic largely unexplored previously. Koçak and Erdal (1999) observed that mite feeding on bean plants result in a decrease in nitrogen and micronutrients like iron and copper in the leaves, while phosphorus and manganese increased, and potassium and zinc levels remained relatively stable. Similarly, Kamel et al. (2019) reported a decrease in potassium and phosphorus contents in mite-damaged pea leaves, accompanied by an increase in nitrogen concentrations. These findings highlight the dynamic changes in plant nutrients induced by T. urticae feeding. By evaluating the specific changes in macronutrient and micronutrient content, as well as growth parameters, in common bean plants infested by T. urticae at different plant growth stages, this study aims to contribute to a better understanding of the effect of mite feeding on the nutrient content of plant.
Material and methods
Mites Rearing
Mites used in this study were sourced from the laboratory of the Plant Protection Department (Faculty of Agriculture of Selcuk University). They were reared and maintained on common bean (Phaseolus vulgaris) plants under laboratory conditions at 28±2 °C, 65±10% RH, and 16 L /8 D / photoperiod.
Plant cultivation
We used in this study the bean cultivar'Yunus 90' because it is widely cultivated in Turkey and is resistant to bacterial and viral diseases (Anonymous 2023). Seeds were obtained from Eskişehir (Turkey) Transitional Zone Agricultural Research Institute. Before sowing, seeds were sterilized by soaking in sodium hypochlorite and then in 75% ethanol, then washed with distilled water. The sterilized seeds were kept in distilled water for 2-3 hours. Afterwards, the germinated seeds were transferred to pots containing soil and peat mixture (1:1). According to the results of soil analysis, a fertilization program was suitably created and routine cultural practices for plants were done.
Bioassay
Artificial infestation with T. urticae was performed at rates of 0, 20, 40, 80 adult mite per plant in 3 groups of plants at different growth stages: young (2nd triple leaf formation), middle (pre-flowering period) and old (fruit formation stage).
Each group consisted of 4 plants and a total of 48 pots were used. The experiments were carried out in climate chambers at 28 ± 2 °C, 65 ± 10% relative humidity and a photoperiod of 16/8 (L/D) hours, and a completely randomized design (CRD) was employed.
The infested plants were left for about two weeks and they were carefully brushed from mites. The leaf samples (15 leaf plant-1) were taken from all directions to ensure that the samples are representative.
The macro (phosphorus, potassium, calcium, magnesium) and micronutrient (iron, copper, manganese, zinc, boron) contents were analyzed according to Burt (2004) in the Laboratory of the Department of Soil Science and Plant Nutrition, Faculty of Agriculture, Selcuk University. Analysis of the macro- and micronutrients were performed using an ICP-OES (Agilent 5110).
Plant growth parameters
The collected bean plant samples were washed with tap water, 0.2 N HCl solution, and distilled water, respectively, and then dried with filter papers and placed in a paper bag. The samples were kept in an oven at 70 ºC to dry until they reached a constant weight and their dry weights were recorded for plant dry weight (g).
The plant height (cm) was recorded by measuring their length from the above-ground parts of the beans. The leaf length (cm) was measured from the end of the plant stem to the tip of the leaf. Moreover, all leaves on the plant were counted and recorded.
Statistical Analysis
Data were analyzed using Two-way ANOVA and differences between treatments were determined in the Minitab19 package program. Percentage data were arcsine-transformed before statistical analysis (Zar 1999). The means were compared by the Tukey HSD (Honestly Significant Differences) test (P ≤ 0.05) using SPSS Base 17.0 for Windows (SPSS Inc., USA).
Results
Effect of four Tetranychus urticae infestation densities on macro and micronutrient contents in bean plants
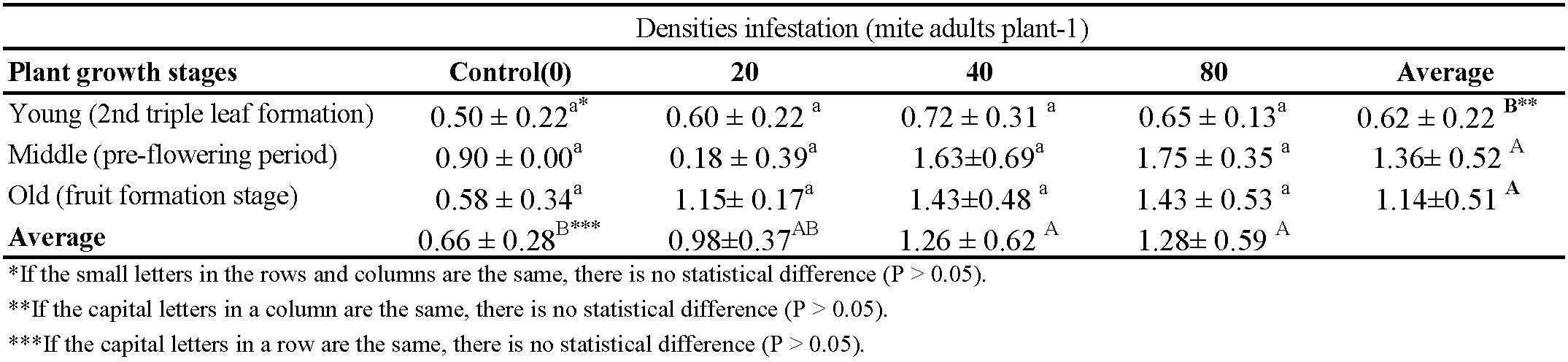
There were significant differences in phosphorus content between different plant stages (P < 0.05) after feeding of T. urticae. However, no significant differences were observed in phosphorus content between the treatments with different mite densities (Table 1).
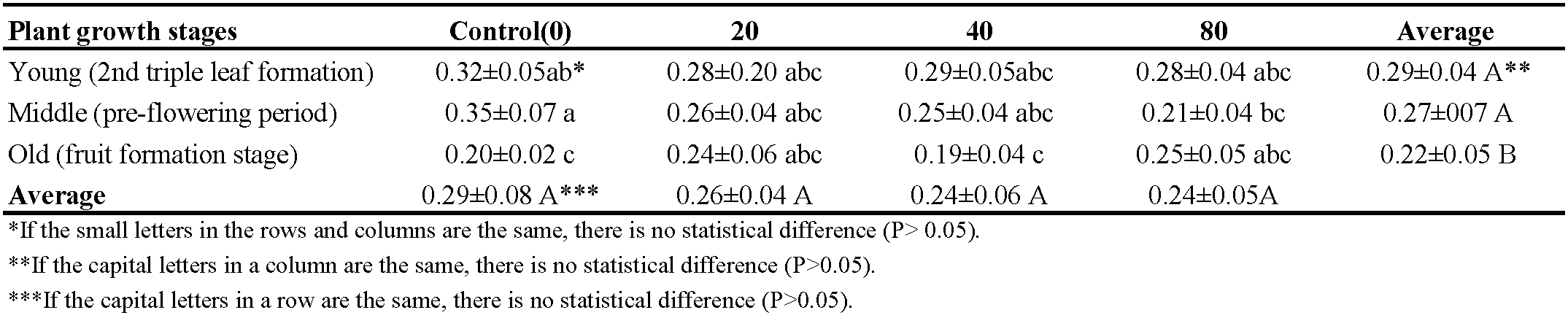

The lowest phosphorus content (0.19%) in the plant was found in the treatment with a density of 40 mites at the old phenological stage of the plant (fruit fotmation), and the highest content (0.35%) was in the control at the middle phenological stage of the plant (pre-flowering period). The phosphorus content was not affected by an increase in the mite density, and no change was observed in its level at the young plant phenological stage (2nd triple leaf formation), but there was a reduction in its content at the middle phenological stage (pre-flowering period). As the phenological stages of T. urticae-infested plants progressed, a concomitant decline in phosphorus content was observed (Table 1).
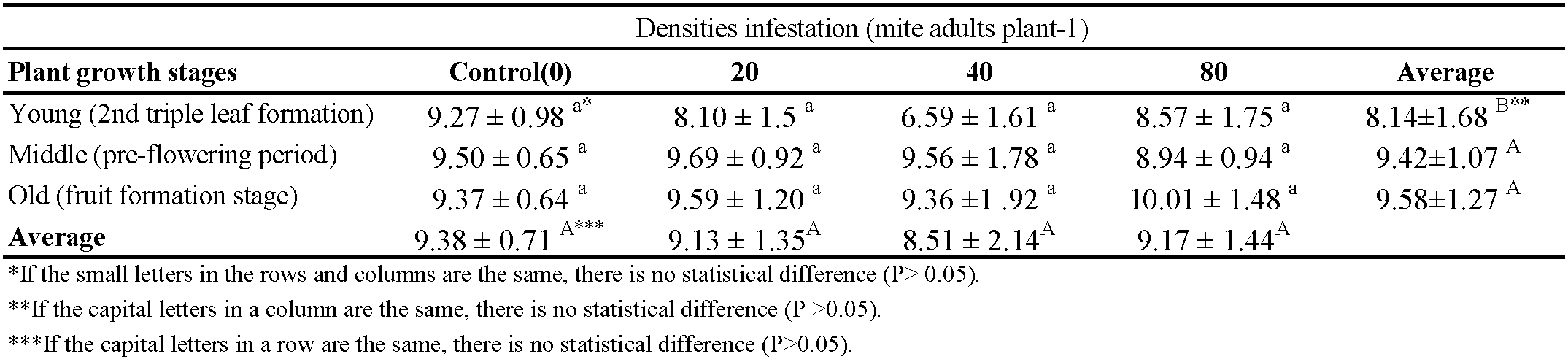
The phenological stages of the plant affected significantly the potassium content (P < 0.05). The lowest potassium content (8.14%) was determined in the young phenological stage. As plant growth increases, mite feeding didn't affect the potassium levels in the plant. Mite feeding reduced potassium levels in the young plant phenological stage. (Table 2).
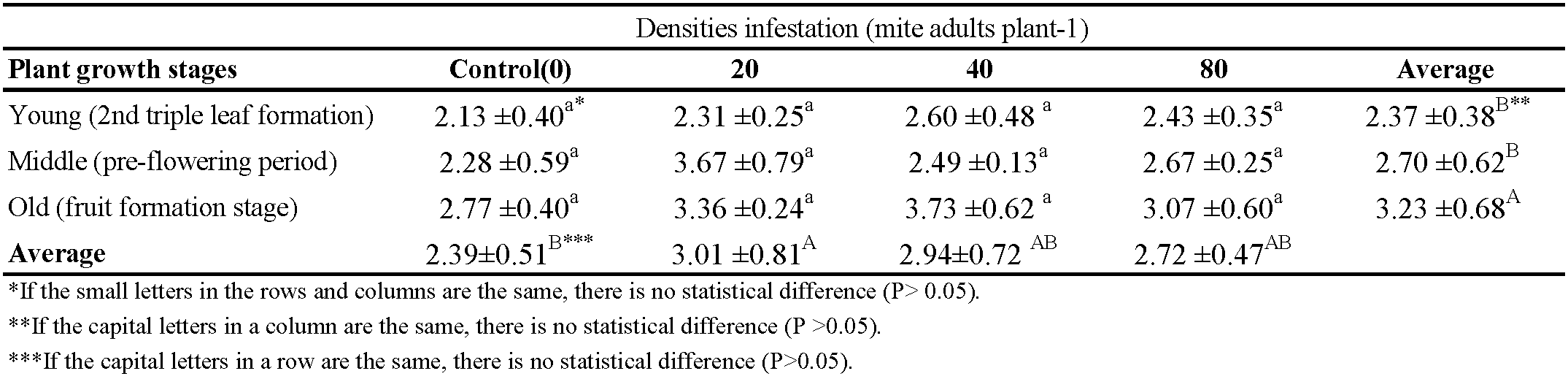
The calcium content in the plant was significantly affected by the plant phenological stages and mite densities (Table 3). The stage of 2nd triple leaf (young stage) contained the lowest calcium level. As can be seen, the plant growth increases, the effect of mites decreased and calcium contents increased. In addition, the increased mite density coincided with a decrease in the calcicum content according to the average of the whole plant stage (Table 3).
There were no effect of mite densities on the content of magnesium in the plant (data not shown).
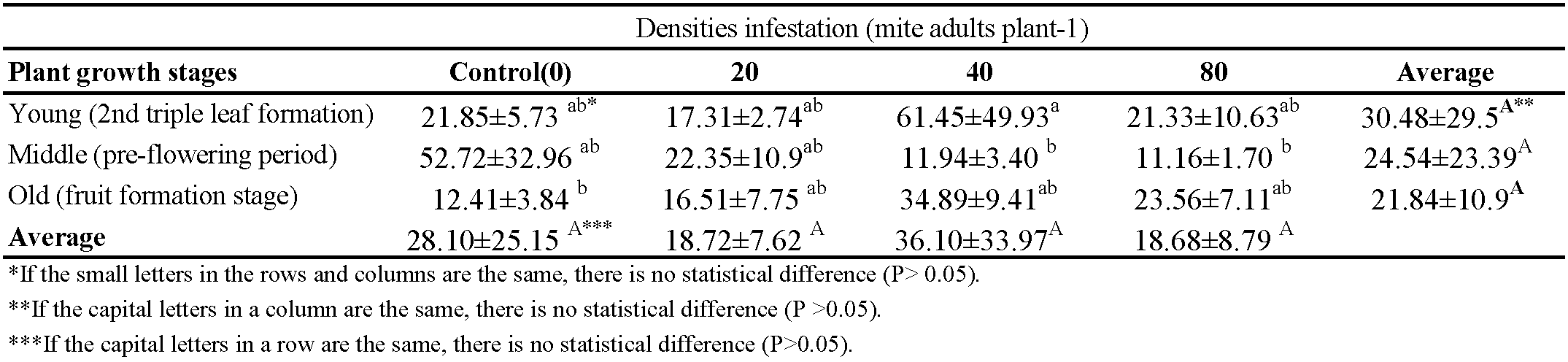
Regarding the effect of mite densities on microelements (iron, manganese, zinc, boron, copper) content in the plant, no differences were observed except for the copper level. The lowest copper content (11.16 ppm) was found in the treatment with a density of 80 mites in the middle stage of the plant, and the highest was 52.72 ppm in the treatment of the control (Table 4).
Effect of different Tetranychus urticae densities on plant growth parameters
An increase in mite density correlated positively with plant dry weight (Table 5), while both plant height and leaf length showed a decrease. There was no significant change in the number of leaves as mite density increased.
Discussion
Our data revealed significant changes in specific macronutrient levels (except for magnesium). The levels of phosphorus and calcium were negatively correlated with T. urticae densities, while potassium correlated positively. Thes results contradict those of Koçak and Erdal (1999), who reported an increase in phosphorus and no change in potassium levels in bean plants infested by T. urticae. These contrasting results could be attributed to several factors, including differences in bean variety, plant growth medium, and mite density.
Divergent outcomes have also been reported by researchers regarding changes in plant nutrients caused by mite feeding on different plant species. For instance, Rasmy et al. (1974) mentioned that the concentrations of nitrogen, phosphorus and potassium decreased with increasing populations of Eutetranychus orientalis feeding on orange plant leaves. Similarly, Kamel et al. (2019) found that potassium and phosphorus contents decreased in the leaves of peas due to T. urticae feeding. Regarding micronutrients, we observed no significant differences in micronutrient contents depending on mite density. This agrees with Koçak and Erdal (1999), who also pointed out that no change in zinc levels in bean leaves infested by T. urticae. However, they found a decrease in iron, copper and an increase in manganese content.
Interestingly, our study showed that the dry weight of the plants increased with an increase in T. urticae density. This coincids with the findings of Rasmy et al. (1974), who reported a similar increase in the dry weight of orange plant leaves infested with E. orientalis. Satar et al. (2020) also observed higher dry matter content in Valencia orange fruits severely damaged by the citrus russet mite Phyllocoptruta oleivora compared to healthy fruits. However, this disagrees with the findings of Koçak and Erdal (1999), who reported no change in dry matter weight in T. urticae-infested bean plants. Furthermore, Rodriguez et al. (1983) found that T. urticae feeding on soybeans at different developmental stages led to a decrease in dry matter, particularly in the young developmental stage. These contrasting results highlight the complex interplay between mite feeding, plant species, and plant growth stages, emphasizing the need for further research to fully understand these interactions.
Conclusion
This study demonstrated that different densities of T. urticae, significantly impacted both macronutrient levels and dry matter content in common bean plants, while having no significant effect on micronutrient levels. One of the most important aspects in crop production is the proper supply of nutrients during the growing stage (Rahbar et al. 2018). Plant nutrient imbalances can reduce resistance to insect and mite damages. For this reason, fertilization programs should be established in order to reduce the damage caused by T. urticae to a certain extent. Our laboratory findings may provide important preliminary information on fertilization in the design of a comprehensive program for management of T. urticae in common bean fields. However, further field studies are necessary to validate these results and to develop practical recommendations. In addition to all these analyzes, the effects of T. urticae on the secondary metabolites in the plant should be determined, and phytochemical analyzes of compounds such as terpenoids, phenolics, alkaloids, flavanoids, cyanogenic glycosides which are involved in the chemical defense mechanism in the plant, should be performed. In this way, it is predicted that the effect of the pest population density and nutrition on the plant can be explained more apparently, and more effective measures can be taken.
Conflict of interest
The authors declare that they have no conflicts of interest.
References
- Abdel-Wali M., Mustafa T., Al-Lala M. 2012. Residual toxicity of abametin, milbemectin and chlorfenapyr to different populations of two-spotted spider mite, Tetranychus urticae Koch, (Acari: Tetranychidae) on cucumber in Jordan. World J. Agric. Sci., 8(2): 174-178.
- Akçin A. 1988. Yemeklik dane baklagiller. Selçuk Üniversitesi Yayınları: 43. Ziraat Fakültesi Yayınları 8. pp. 377. (in Turkish)
- Altieri M.A., Nicholls C.I. 2003. Soil fertility management and insect pests: Harmonizing soil and plant health in agroecosystems. Soil & Tillage Research, 72: 203-211. https://doi.org/10.1016/S0167-1987(03)00089-8
- Anonymous 2023. Characteristics of Yunus-90 bean seed, Available from: https://arastirma.tarimorman.gov.tr/gktaem/Belgeler/Tescilli%20%C3%87e%C5%9Fitlerimiz/Kuru%20Fasulye/yunus-90.pdf
- Atalay E., Kumral N.A. 2013. Tetranychus urticae (Koch) (Acari:Tetranychidae)' nin farklı sofralık domates çeşitlerinde biyolojik özellikleri ve yaşam çizelgeleri, Türkiye Entomoloji Dergisi, 37 (3): 329-34. (in Turkish)
- Burt R. 2004. Soil survey laboratory methods manual. Soil Survey Investigations Report. (No:42). United States Department of Agriculture, Natural Resources Conservation Service, National Soil Survey Center, Lincoln, Nebraska.
- Çobanoğlu S., Kumral N.A. 2014. Ankara, Bursa ve Yalova illerinde domates yetiştirilen alanlarda zararlı ve faydalı akar (Acari) biyolojik çeşitliliği ve popülasyon dalgalanması. Türkiye Entomoloji Dergisi, 38 (2): 197-214. (in Turkish)
- Elma F.N., Alaoğlu Ö. 2008. Konya ilinde peyzaj alanlarındaki ağaç ve çalılarda bulunan zararlı akar türleri ve doğal düşmanları. Türkiye Entomoloji Dergisi, 32(2): 115-129. (in Turkish)
- Erdoğan P. 2016. Kadife çiçeği (Tagetes erecta L.)'nin Tetranychus urticae Koch. (Acari: Tetranychidae)'ye karşı tuzak bitki olarak kullanımı. Akademik Ziraat Dergisi, 6: 109-114. (in Turkish)
- Harris F.A., Fupper G.R., Furr R.E., Dugger P., Richter D. 1998. Potassium fertilizer effect on twospotted spider mite. Proceedings Beltwide Cotton conferences, San Diego, California, USA p: 982-983.
- Kamel M.S., Abou-Zaid A.M., Yasin, S.A. 2019. The Effectiveness of Phytochemical Components and Climatic Factors on Population Fluctuation of the Spider Mite, Tetranychus urticae Koch on Sweet Pea and Pea Crops. Journal of Applied Plant Protection, 8(1): 15-21. https://doi.org/10.21608/japp.2019.59842
- Koçak E., Erdal İ. 1999. İki noktalı kırmızıörümcek [(Tetranychus urticae Koch (Acarina: Tetranychidae)]'nin fasulyenin (Phaseolus vulgaris L) besin kompozisyonu ve kuru madde ağırlığına etkileri. Bitki Koruma Bülteni, 39(3): 151-164. (in Turkish)
- Kytö M., Niemelä P., Larsson S. 1996. Insects on trees: population and individual response to fertilization. Oikos, 75(2): 148-159. https://doi.org/10.2307/3546238
- Mammadova T., Guliyeva L., Moulahoum H., Tok K., Küçükçobanoğlu, Y., Yildiz Aktas L., Zihnioglu F. 2023. Role of phytochemicals and secondary metabolites from Mentha spicata in acetylcholine esterase inhibition for effective pest control of Tetranychus urticae Koch. International Journal of Acarology, 49(7-8): 366-377. https://doi.org/10.1080/01647954.2023.2275754
- Nansen C., Sidumo A.J., Martini X., Stefanova K., Roberts J.D. 2013. Reflectance-based assessment of spider mite "bio-response» to maize leaves and plant potassium content in different irrigation regimes. Computers and Electronics in Agriculture, 97: 21-26. Doi: https://doi.org/10.1016/j.compag.2013.06.007
- Polat H., Kasap İ. 2011. Van ilinde üç farklı fasulye (Phaseolus vulgaris L.) çeşidi üzerinde ikinoktalı kırmızıörümcek, Tetranychus urticae Koch, 1836 (Acari: Tetranychidae)'nin popülasyon gelişimi. Turkish Journal of Entomology 35(1): 145-154. (in Turkish)
- Rahbar M., Fathipour Y., Soufbaf M. 2018. Fertilizer-Mediated Ditrophic Interactions between Aphis gossypii and Cucumber. Journal Agricultural Science and Technology, 20(5): 987- 998.
- Rasmy A.H., Abouaziz A.B., Eltanahy M.M. 1974. Effect of citrus brown mite, Eutetranychus orientalis (Acarina: Tetranychidae), infestation on the N, P, K and pigments of sour orange leaves. Experientia, 30: 1016-1017. https://doi.org/10.1007/BF01938981
- Ribeiro M.G.P.M.M., Micherff Filho M., Guedes I.M.R.., Junqueıra A.M.R., 2012. Efeito da adubação química na infestação do ácaro rajado e na produção do morangueiro. Horticultura Brasileira, Vitória da Conquista, 30(4): 673-680. https://doi.org/10.1590/S0102-05362012000400018
- Rodriguez J.R., Reicosky D.A., Patterson C.G. 1983. Soybean and mite interaction: effects of
- cultivar and plant growth stage, Journal of the Kansas Entomological Society, 56 (3): 320-326.
- Satar S., Tiring, G., Tusun A., Yeşiloğlu T. 2020. The effect of Phyllocoptruta oleivora (Ashmead) (Acari: Phyllocoptidae) on fruit quality of Valencia orange Derim, 37(1): 44-50. https://doi.org/10.16882/derim.2020.591334
- Tehri K., Gulati R., Geroh M. 2014. Damage potential of Tetranychus urticae Koch to cucumber fruit and foliage: Effect of initial infestation density. Journal of Applied and Natural Science, 6(1): 170-176. https://doi.org/10.31018/jans.v6i1.395
- Ülker M., Ceyhan E. 2008. Determination of some agricultural characters of common beans (Phaseolus vulgaris L.) genotypes in central Anatolian ecological condition. Selcuk Journal of Agriculture and Food Sciences, 22(46): 77-89.
- Whalon M., Mota-Sanchez D., Holling-worth R. 2008. Analysis of global pesticide resistance in arthropods. In: Global Pesticide Resistance in Arthropods. CABI: Publisher. p. 5-31. https://doi.org/10.1079/9781845933531.0005
- Zar J.H. 1999. Biostatistical analysis. Prentice-Hall: Publisher. pp: 662



2024-03-05
Date accepted:
2024-11-06
Date published:
2024-11-19
Edited by:
Tsolakis, Haralabos

This work is licensed under a Creative Commons Attribution 4.0 International License
2024 Türkoğlu, Huriye; Çetin, Hüseyin and Elma, Fatma Nur
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)