



Synonymy of the water mite subgenera Orientmomonia and Kondia in the genus Momonia (Momoniidae, Acari): an evaluation based on morphology and molecular data
Abé, Hiroshi  1
; Iwasa, Masahiro A.
2
and Morimoto, Shizuko3
1
; Iwasa, Masahiro A.
2
and Morimoto, Shizuko3
1✉ Biological Laboratory, College of Bioresource Sciences, Nihon University, Kameino 1866, Kanagawa 252-0880, Japan.
2Division of Natural History, Department of Zoological Science, College of Bioresource Sciences, Nihon University, Kameino 1866, Kanagawa 252-0880, Japan.
3Museum of Nature and Human Activities, Hyogo, 6 Yayoigaoka, Sanda, Hyogo 669-1546, Japan.
2024 - Volume: 64 Issue: 2 pages: 353-362
https://doi.org/10.24349/gf51-eshnZooBank LSID: 310A2A99-E973-4B80-B581-EDE029ECED11
Original research
Keywords
Abstract
Introduction
The genus Momonia in the family Momoniidae consists of five subgenera: Momonia, Kondia, Orientmomonia, Paramomonia, and Zelandomomonia (Smit 2020). Among these five subgenera, Kondia was originally established by Sokolow (1926) as a new genus based on a single female specimen of Kondia karelica from Russia; thereafter, this genus was moved to subgeneric rank by Cook (1974). Therefore, this species is currently presented as Momonia (Kondia) karelica. On the other hand, Orientmomonia was established by Pešić (2014) based on a single male specimen of Momonia (Orientmomonia) koreana from South Korea.
Recently, the authors had an opportunity to examine several specimens of Momonia collected from a river in Hyogo and Nagasaki Prefectures in Japan. After a brief morphological examination of these specimens, sympatric male and female individuals were assumed to belong to different subgenera. All males obtained from the collecting site had characteristics of the subgenus Orientmomonia, and all the females had those of the subgenus Kondia. This suggests the possibility that Orientmomonia is synonymous with Kondia. Therefore, we test the hypothesis that Orientmomonia is synonymous with Kondia based on the sympatric male and female specimens in the light of morphology and molecular data.
Material and methods
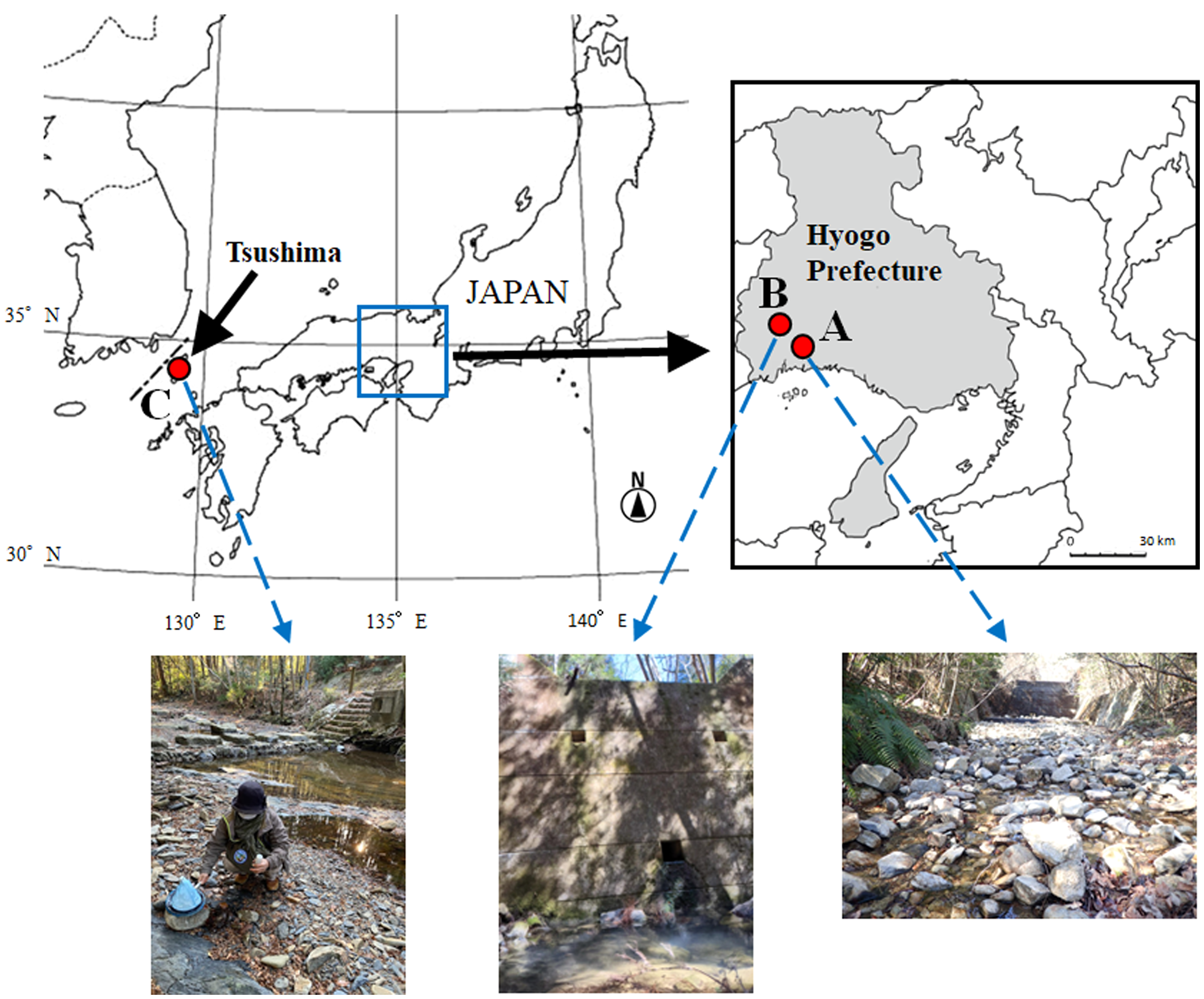
Sympatric male and female Momonia specimens were collected from the Otani River (34°47′02.70″ N, 134°30′24.85″ E) (Fig. 1A), a tributary of the Ibo River, at Ibogawa Town, Tatsuno City, Hyogo Prefecture, Japan, on December 25, 2019; November 19 and 20, 2021; and January 14 and 23, 2022. Allopatric female specimens were collected from the Imotani River (34°50′20.4″ N, 134°27′39.6″ E) (Fig. 1B) at Wakasano Town, Aioi City, Hyogo Prefecture, Japan, on March 3 and 16, 2022, and the Syushi River (34°35′16.8″ N, 129°26′02.4″ E) (Fig. 1C) at Kamitsushima Town, Tsushima City, Nagasaki Prefecture, Japan, on December 31, 2022. The collection sites at the Otani River and Imotani River were puddles of seepage water from the drainage port at the bottom of the erosion control dam. The collection site at the Syushi River was a small pool beside a small stream flowing through the park. The mites were collected by sweeping the pool with a plankton net with a 300 μm mesh size. All specimens were collected by one of the authors, S. Morimoto, and/or the late Mr Kazuhisa Inada (Himeji City).
Morphological investigations
The specimens were preserved in Imamura's fluid (glycerin : tap water : acetic acid = 10 : 9 : 1) (Imamura 1965). They were mounted in glycerin jelly, and the cover slips were sealed with Canada balsam under a stereomicroscope (SZH10, OLYMPUS Corp.). They were observed under a light microscope (BH-2, OLYMPUS Corp.). Photos of unmounted specimens were taken with a digital microscope (VCR800, OMRON Corp.). Illustrations were drawn with the aid of a camera lucida, and measurements were made with an ocular micrometre. In the text, measurements are always given in micrometres (µm).
The body parts were measured in the following format: (1) Idiosoma: ventral length (i.e. total length), from the anterior-most margin of the first coxal plate to the terminal end of the idiosoma; dorsal length, from the anterior-most margin to the terminal end of the idiosoma; width, at the widest point. (2) Anterior dorsal plate, posterior dorsal plate, anterior coxal group (Cx-I–II), total coxal group (Cx-I–IV): length, from the anterior- to posterior-most margins along the longitudinal median axis; width, at the widest point. (3) Seven pairs of platelets of the dorsum (Pl-1–7): length, the longest position of the platelets. (4) Genital field: length, from the anterior to posterior margins of the gonopore in the male, from the pregenital to postgenital plates in the female; width, at the widest point of the gonopore in the male, the widest point of the lateral margins of both genital plates in the female. (5) Genital plate: length, from the anterior- to posterior-most margins of each right or left plate along the longitudinal median axis; width, at the widest point of each right or left plate. (6) Gnathosoma: length, from the anterior- to posterior-most margins of gnathosoma with posterior anchoral process. (7) Chelicera: length, straight dorsal length from the proximal base to the distal end including claw. (8) Palp (P-1–5) and leg segments (I-L-1–6, IV-L-3–6): length, straight dorsal length from the proximal base to the distal end; height, the middle part of segments in I-L-4, I-L-5, and the highest point in I-L-6. (9) L, length; W, width; H, height.
The material is deposited in the Museum of Nature and Human Activities, Hyogo (MNHAH), Sanda, Hyogo, 669-1546, Japan.
Molecular investigations
In total, five sympatric specimens that had been stored in 70% EtOH were used for the current analysis (Table 1). Of them, the entire body of two male and one female specimens (#A, #B, and #C) from the Otani River was completely crushed, and its inner tissues were squeezed by needles. On the other hand, in two female specimens (#G and #H) from the Otani River, a gnathosoma was removed and inner tissues were extracted from the exoskeleton with needles for specimen storage. This was done with extreme care to avoid breaking the exoskeleton (Meguro et al. 2017). After sufficient removal of the EtOH through air-drying treatment, these inner tissues were digested by the pK treatment (2 µL of pK 300 mAU/mL + 50 µL pK buffer, in total) at 37˚C overnight. After the pK treatment, genomic DNA was extracted using Promega Wizard Genomic DNA Purification Kit (#A1120) in accordance with the supplier's instruction.

A fragment approximately 700 bp long of the mitochondrial cytochrome oxidase subunit I (COI) gene region was amplified by the PCR method using primers LCO1490 for the forward direction and HCO2198 for the reverse direction (Folmer et al. 1994). PCR amplification was performed, following the procedure described in Goka et al. (2001) and Meguro et al. (2017). After confirmation of the amplification, the primers were removed by PEG (20% polyethylene glycol + 2.5M NaCl) precipitation, and both strands of the PCR products were sequenced directly and automatically using the BigDye Terminator Cycle Sequencing Kit ver. 3.1 (ABI) and the 3730 Genetic Analyzer (ABI). All sequences were aligned manually using the IdentityX program. The current sequences were deposited in the DNA databases (DDBJ/EMBL/GenBank) with accession Nos. LC794182–LC794186. In addition, we constructed a maximum-likelihood tree with the current sequences and those of nine water mite taxa (Table 1) based on the Tamura-Nei model (Tamura and Nei 1993) that considered all codon positions. Bootstrap analysis was performed (1,000 replicates) using MEGA11 (Tamura et al. 2021). The tick Ixodes ricinus was used as an outgroup (Table 1).
Results
Morphological investigations
As a result of the comparative examination of the diagnostic characteristics of Momonia species obtained in the present study, all male specimens are identified as Momonia (Orientmomonia) koreana Pešić, 2014. On the other hand, all female specimens are identified as Momonia (Kondia) sp. Taxonomic descriptions of these two species are given below.
Momonia (Orientmomonia) koreana Pešić, 2014

Specimens examined — One male, from the Otani River, Nov. 19, 2021, K. Inada leg.; thirty-eight males, the same locality as above, Nov. 20, 2021, and Jan. 23, 2022, K. Inada and S. Morimoto leg.
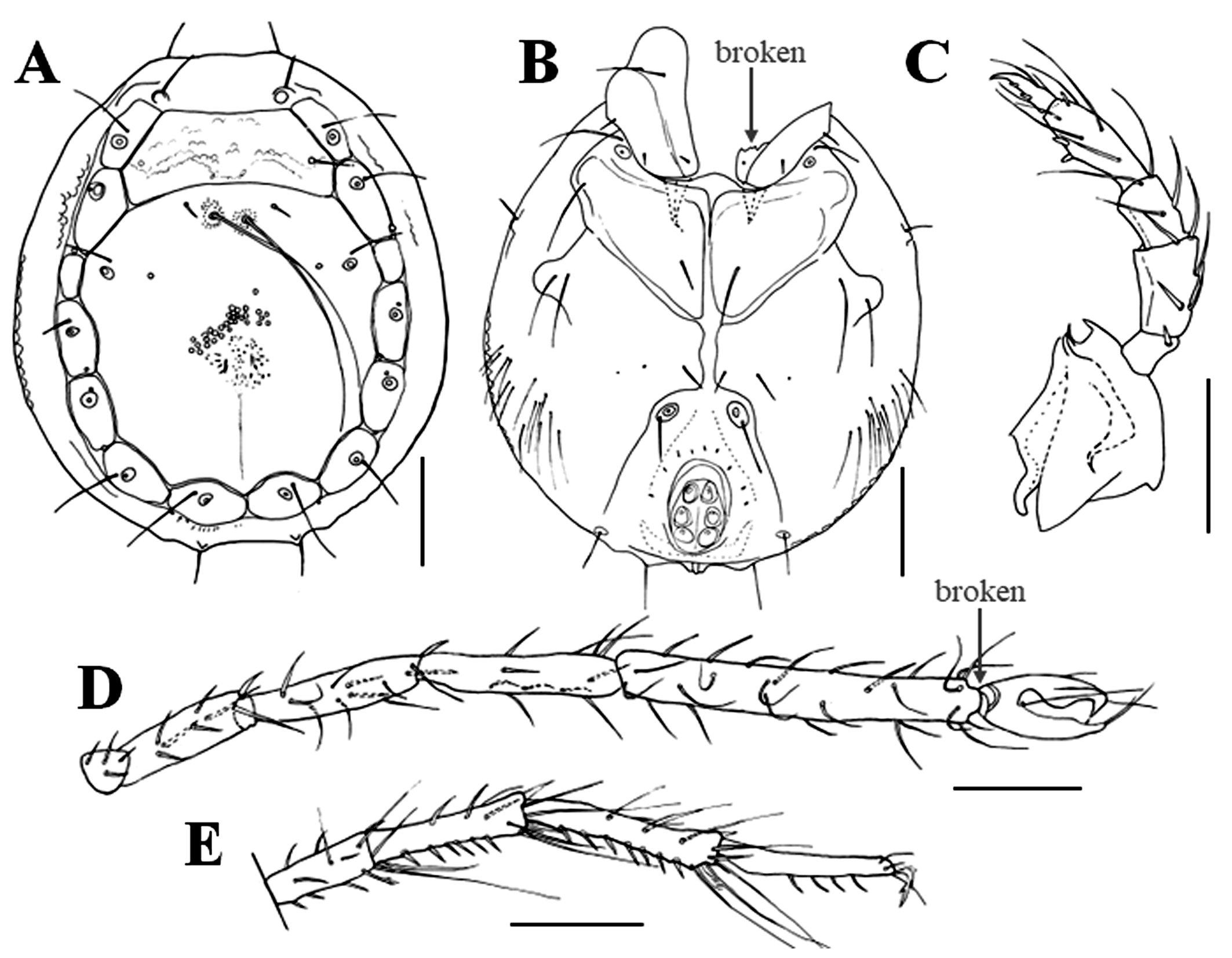
Characteristics — Light green body with dark red eyes (Fig. 2A). Idiosoma oblong and flatback; Pl-1–7 surrounding anterior and posterior dorsal plates; posterior end of anterior dorsal plate with a depression; middle of the anterior margin of the posterior dorsal plate with a pair of long setae (Fig. 3A). Cx-IV surrounding the genital field, gonopore with three pairs of acetabula (Fig. 3B). P-1–5 with 1, 5, 4, 8, 6 setae (including spine and claw) (Fig. 3C); P-4 with two ventral spines and one anterodorsal peg-like spine; distal two filiform setae on P-3 and most filiform setae on P-4 long and slender. I-L-5 nearly three times longer than I-L-6, 1.28–1.41 times higher than I-L-4; ratio of length to height 6.78–7.65 in I-L-5, 1.81–2.27 in I-L-6 (Fig. 3D); III-L-3–5 and IV-L-3–5 each with 1–3, 3–5, 2–3 distal, long swimming setae and sometimes also with plumose spines (Fig. 3E).
Measurements (n = 6) — Idiosoma ventral L/W 472–504/365–410, dorsal L 443–467; anterior dorsal plate L/W 57–65/189–217; posterior dorsal plate L/W 287–303/267–295. Pl-1–7 L: 69–78, 70–90, 33–45, 66–78, 62–78, 66–74, 53–70. P-1–5 L: 25–34, 63–69, 42–46, 69–80, 40–53. Cx-I–II L/W 123–144/193–209. Gonopore L/W 62–77/42–53. Gnathosoma L 111–126. Chelicera L 105–120. I-L-1–6 L: 33–49, 86–115, 144–152, 152–160, 278–303, 94–107; I-L-4–6 H: 29, 37–41, 43–57; I-L-5 L/I-L-6 L ratio 2.59–3.05, I-L-5 H/I-L-4 H ratio 1.28–1.41, I-L-5 L/I-L-5 H ratio 6.78–7.65, I-L-6 L/I-L-6 H ratio 1.81–2.27; IV-L-3–6 L: 94–103, 119–131, 145–152, 123–144.
Remarks — The outstanding characteristic of the male is the presence of seven pairs of platelets surrounding the anterior and posterior dorsal plates. This feature is applicable to the monotypic subgenus Orientmomonia established by Pešić (2014) based on a single male specimen of Momonia (Orientmomonia) koreana Pešić, 2014. Characteristics of the present males accord well with those of M. (O.) koreana, except for a somewhat smaller body size. Length and height ratios of I-L-5 to I-L-6 in the present males almost agree with those in the male of M. (O.) koreana. The present males have a pair of long setae on the anterior margin of the posterior dorsal plate that were not shown in the original figure (Fig. 8A) of M. (O.) koreana by Pešić (2014). However, after confirming this matter with Dr Pešić, it is proven that these long setae are probably present also in M. (O.) koreana. As a result of morphological examination, it can be concluded that the present male is M. (O.) koreana Pešić, 2014.
Momonia (Kondia) sp.
Specimens examined — One female, from the Otani River, Dec. 25, 2019, K. Inada leg.; 27 females, the same locality as above, Nov. 20, 2021, Jan. 14 and 23, 2022, K. Inada and S. Morimoto leg.; one female, the Imotani River, March 3, 2022, Inada leg; two females, the same locality as above, March 16, 2022, Morimoto leg; three females, the Syushi River, Dec. 31, 2022, S. Morimoto leg.
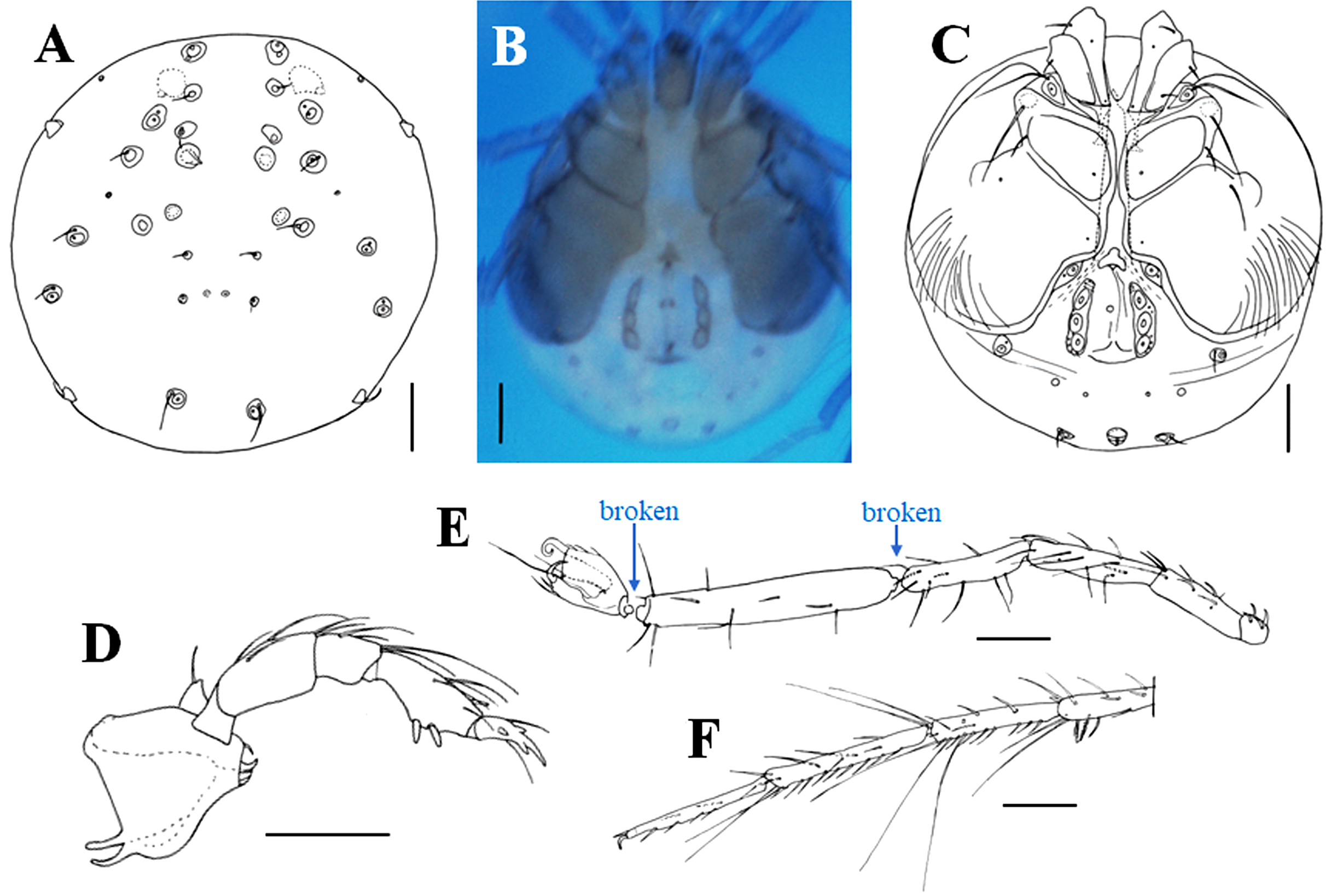
Characteristics — Light green body with dark red eyes (Fig. 2B). Idiosoma soft and round; anterior dorsal portion with a depression and two pairs of round dorsal muscle insertion platelets, anterior pair 31–35 and posterior pair 25 in diameter; posterior dorsal part with a pair of small pores and two pairs of non-glandular setae arranged in a rectangular shape (Fig. 4A). Right and left Cx-I–IV widely separated in the unmounted specimen (Fig. 4B), whereas sometimes closer due to artificial deformation (Fig. 4C); Cx-III with a long seta on anterolateral corner; Cx-IV greatly expanding posteriorly as well as laterally, posterolaterally rounded, with almost 15 setae in posterolateral area. Genital plates elongated bearing three pairs of acetabula. P-1–5 with 1, 5, 4, 7, 6 setae (including spine and claw), respectively (Fig. 4D); P-4 with two ventral spines and one anterodorsal peg-like spine; distal two filiform setae on P-3 and most filiform setae on P-4 long and slender. I-L-5 more than three times longer than I-L-6, 1.58–1.68 times higher than I-L-4; ratio of length to height 5.69–6.37 in I-L-5, 1.31–1.72 in I-L-6 (Fig. 4E); III-L-3–5 and IV-L-3–5 each with 1–3, 3–5, 2–3 distal, long swimming setae and sometimes also with plumose spines (Fig. 4F).
Measurements (n = 6) — Idiosoma ventral L/W 607–779/541–713, dorsal L 525–656. P-1–5 L: 27–41, 78–95, 46–61, 82–105, 50–69. Cx-I–II L/W 135–170/205–270. Cx-I–IV L 443–526. Genital field L/W 160–220/110–143; Genital plate L/W 100–126/29–36. Gnathosoma L 130–162. Chelicera L 126–149. I-L-1–6 L: 41, 98–135, 152–193, 160–201, 312–394, 98–123; I-L-4–6 H: 31–37, 49–62, 57–82; I-L-5 L/I-L-6 L ratio 3.14–3.60, I-L-5 H/I-L-4 H ratio 1.58–1.68, I-L-5 L/I-L-5 H ratio 5.69–6.37, I-L-6 L/I-L-6 H ratio 1.31–1.72; IV-L-3–6 L: 103–148, 156–205, 205–253, 164–230.
Remarks — Main characteristics of the female are the soft body, the presence of two pairs of round dorsal muscle insertion platelets on the anterior half of the dorsum, two pairs of setae on the posterior dorsal portion, long filiform seta on the anterior corner of Cx-III, and widely separated right and left Cx-I–IV. These features are applicable to the monotypic subgenus Kondia originally established by Sokolow (1926). At present, only females of a single species Momonia (Kondia) karelica are known. This species was originally recorded in the Republic of Karelia in the north-western part of Russia. Afterwards, this species was recorded also from Gästrikland on the eastern coast of Sweden by Lundblad (1962). Specific features and illustrations of M. (K.) karelica in Cook (1974) (Fig. 1628) and Smit (2020) (Fig. 3008) probably referred to Lundblad (1962) (Fig. 409) instead of the original description by Sokolow (1926). However, the trouble is that taxonomic features of this species given in Lundblad (1962) are slightly different from those in Sokolow (1926). The main differences based on the illustrations are as follows (features in Lundblad (1962) are given in parenthesis): The anterior end of Cx-I is not protruded forward from the anterior margin of the dorsum (clearly protruded); the posterior two-thirds of the genital plate beyond posteriorly to the level of posterior margin of Cx-IV (half of the genital plate). This cannot exclude the possibility that the female in Lundblad (1962) was not M. (K.) karelica. Therefore, we compared the present females only with M. (K.) karelica in the original descriptions by Sokolow (1926).
The results of morphological comparisons between these two species are as follows (features of M. (K.) karelica are indicated in parenthesis): The posterior pair of the round dorsal muscle insertion platelets are 25 in diameter (12 in diameter); two pairs of dorsal setae on the posterior part of the dorsum are arranged to form a rectangle (inverted trapezoid); the anterior end of Cx-I protruded forward from the anterior margin of the dorsum (not protruded); about one-quarter of the genital plates beyond posteriorly to the level of posterior margin of Cx-IV (more than half of the genital plate); anterior margin of the genital plate with three setae (two setae). Due to the morphological differences mentioned above, it can be concluded that the present female is not M. (K.) karelica.
Molecular investigations
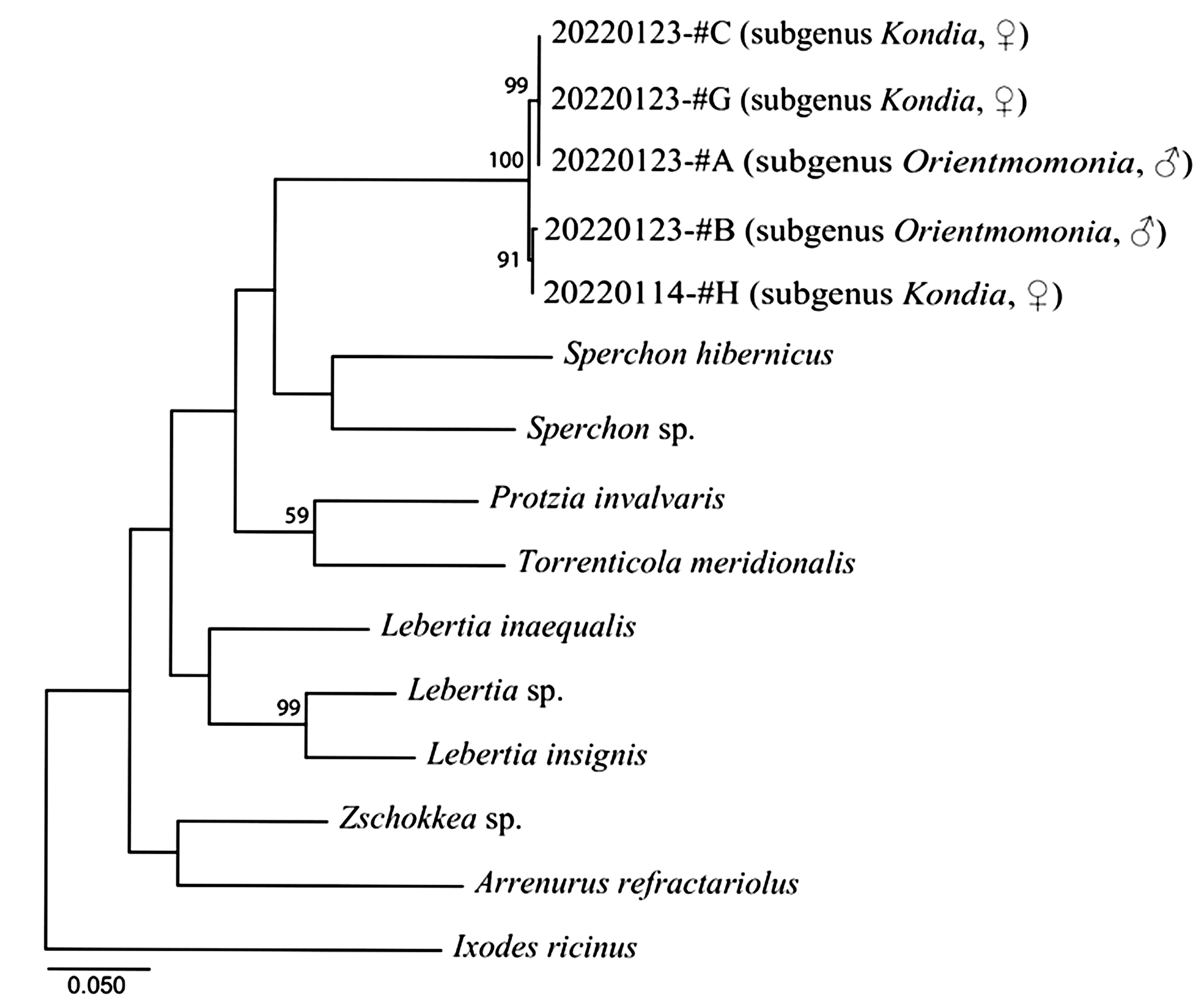
We determined 658 bp length of the COI gene sequences for the current samples. A phylogenetic tree was constructed (lnL = -4380.707, BIC = 9076.316, AICc = 8829.642) with a transition/transversion ratio of 1.076 and most transversions were recognized intergenerically. In the tree, our mite samples form a single clade from the other taxa (Fig. 5). This clade is divided into two subclades as #A-#C-#G and #B-#H and there were four transitions between both subclades. In each subclade, #A, #C, and #G shared the same haplotype and #B and #H carried quite similar haplotypes with only one transversion.
Discussion
As a result of the morphological examination of taxonomic characteristics in the present specimens, it became clear that the male is identified as Momonia (Orientmomonia) koreana Pešić, 2014, and the female is Momonia (Kondia) sp. In addition, considering the differentiation states of the COI gene among the five Momonia samples and the intergeneric relationships in other taxa of the current phylogenetic tree, we can conclude that our five mite samples seem to be conspecific. Namely, the male and female of Momonia species in the present study turned out to be of the same species.
From the results obtained in the present investigations, the subgenus Orientmomonia Pešić, 2014 is regarded as the junior synonym of the subgenus Kondia Sokolow, 1926 according to the Article 23.3.5 in ICZN (1999). Therefore, the taxonomic status of the subgenus Orientmomonia and the diagnosis of the subgenus Kondia are revised as follows.
Genus Momonia Halbert, 1906
[Japanese name: Kawa-momodani zoku]
Subgenus Kondia Sokolow, 1926
[Japanese name: Tamari-momodani azoku]
(Type: Kondia karelica Sokolow, 1926)
Kondia Sokolow, 1926: 32.
Momonia Halbert, 1906: 5; K. Viets, 1926: 195; 1956: 515; Sokolow, 1927: 309.
Momonia (Kondia) Cook, 1974: 371; K. O. Viets, 1987: 531; Gerecke et al., 2016: 370; Smit, 2020: 706.
Momonia (Orientmomonia) Pešić, 2014: 223; Smit, 2020: 706–707. Syn. nov.
Diagnosis — Male: Dorsal shield composed of small anterior and large posterior plates, flanked by seven pairs of platelets. Female: Soft body, two pairs of round dorsal muscle insertion platelets on the anterior half of the dorsum, two pairs of setae on the posterior dorsum, right and left Cx-I–IV widely separated, posterolateral portion of Cx-IV expanded.
In addition, the specific status and diagnosis of Momonia koreana are revised as follows.
Momonia (Kondia) koreana Pešić, 2014 comb. nov.
[Japanese name: tamari-momodani]
Momonia (Orientmomonia) koreana Pešić, 2014: 223–225, Figs. 5D–F, 8A–B, 9A–I; Smit, 2020: 705–707, Figs. 3004, 3009–3011. Syn. nov.
Diagnosis — Males: The anterior margin of the posterior dorsal plate with a pair of long setae. Females: Two pairs of dorsal setae on the posterior part of the dorsum arranged to form a rectangle, the posterior one-quarter of the genital plate beyond posteriorly to the level of the posterior margin of Cx-IV.
Remarks — The sexual dimorphism of the dorsum (male: dorsum covered by plates and platelets vs. female: dorsum mainly soft) is clearly differentiating the subgenus Kondia from Momonia s. str. The subfamily Momoniinae is newly recorded in Japan. According to the current knowledge, the subgenus Kondia in the genus Momonia contains two described species (probably three, as it is rather probable, the female described by Lundblad (1962) is not conspecific with M. karelica): Momonia (Kondia) karelica and M. (K.) koreana. Considering the present distribution of these two species, the subgenus Kondia is probably widely distributed in the Palearctic region.
Acknowledgments
We are very grateful to Dr Pešić (Univ. of Montenegro) for his competent advice on identifying the present male specimens as Momonia (Kondia) koreana. Special thanks are also due to the late Mr K. Inada, who discovered the existence of Momonia species in Hyogo Prefecture in Japan. Finally, we would like to express our heartfelt condolences on the passing of Mr K. Inada.
References
- Cook D.R. 1974. Water mite genera and subgenera. Mem. Amer. Ent. Inst., 21: 1-860.
- Folmer O., Black M., Hoeh W., Lutz R., Vrijenhoek R. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol., 3: 294-299.
- Gerecke R., Gledhill T., Pešić V., Smit H. 2016. 8. Acari: Hydrachnidia III. In: Gerecke R. (ed.), Chelicerata: Acari: III. Süßwasserfauna von Mitteleuropa 7/2-3. Heidelberg, Dordrecht, London, New York: Springer Spektrum, pp. 1-429. https://doi.org/10.1007/978-3-8274-2689-5
- Goka K., Okabe K., Yoneda M., Niwa S. 2001. Bumblebee commercialization will cause worldwide migration of parasitic mites. Mol. Ecol., 10: 2095-2099. https://doi.org/10.1046/j.0962-1083.2001.01323.x
- Halbert J.N. 1906. Notes on Irish Hydrachnida; with descriptions of a new genus and two new species. Ann. Mag. Nat. Hist., 18 (7): 4-12. https://doi.org/10.1080/00222930608562568
- ICZN (International Commission on Zoological Nomenclature). 1999. International Code of Zoological Nomenclature, 4th Edition. London: The International Trust for Zoological Nomenclature; 306 pp.
- Imamura T. 1965. A case of water mites, 4. Collection and experimental techniques. In: Sasa, M. (ed.). Mites, an introduction to classification, bionomics and control of Acarina. Tokyo: University of Tokyo Press; p. 34. [In Japanese]
- Lundblad O. 1962. Die Hydracarinen schwedens. II. Ark. Zool., 14 (1): 215-216.
- Meguro H., Iwasa M.A., Goka K., Abé H. 2017. Molecular phylogenetic relationships of halacarid mites suggest the reevaluation of traditional subfamily classification. Acarologia, 57: 633-641. https://doi.org/10.24349/acarologia/20174182
- Pešić V. 2014. Checklist of the water mites (Acari, Hydrachnidia) of Korea, with description of one new subgenus and two new species. Ecol. Montenegrina, 1: 204-230. https://doi.org/10.37828/em.2014.1.29
- Smit H. 2020. Water mites of the world, with keys to the families, subfamilies, genera and subgenera (Acari: Hydrachnidia). Monogr. Ned. Entomol. Ver., 12: 1-774.
- Sokolow I. 1926. Eine neue Hydracarinen-Gattung aus Russisch-Karelien, Kondia nov. gen. Zool. Anz., 66: 32-40.
- Sokolow I. 1927. Über die Identität der Hydracarinengattung Kondia Sokolow mit Momonia Halbert. Zool. Anz., 69: 308-310.
- Tamura K., Nei M. 1993. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol., 10: 512-526. https://doi.org/10.1093/oxfordjournals.molbev.a040023
- Tamura K., Stecher G., Kumar S. 2021. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol., 38: 3022-3027. https://doi.org/10.1093/molbev/msab120
- Viets K. 1926. Versuch eines Systems der Hydracarinen. Zool. Anz., 69 (7-8): 188-199.
- Viets K. 1956. Die Milben des Süßwassers und des Meeres. Hydrachnellae et Halacaridae (Acari). II. und III. Teil: Katalog und Nomenklator. Jena, Germany: G. Fischer; 870 pp.
- Viets K.O. 1987. Die Milben des Süßwassers (Hydrachnellae und Halacaridae [part.], Acari). II: Katalog. Sonderb. Naturwiss. Ver. Hamburg, 8: 1-1012.



2024-01-22
Date accepted:
2024-03-07
Date published:
2024-03-14
Edited by:
Mąkol, Joanna

This work is licensed under a Creative Commons Attribution 4.0 International License
2024 Abé, Hiroshi; Iwasa, Masahiro A. and Morimoto, Shizuko
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)