



An update on the knowledge about scutacarid mites as phoronts and inquilines
Baumann, Julia  1
1
1✉ Institute of Biology, Department of Biodiversity and Evolution, University of Graz, Universitätsplatz 2, 8010 Graz, Austria.
2024 - Volume: 64 Issue: 1 pages: 3-17
https://doi.org/10.24349/sy16-8iqnOriginal research
Keywords
Abstract
Introduction
Scutacarids (Scutacaridae, Pygmephoroidea, Heterostigmatina) are mostly soil-inhabiting, fungivorous mites which are often associated with various other animal taxa: about half of the more than 800 described species (most recent number published by Rahiminejad et al. (2023)) can be found as phoronts or inquilines of other animals. In my review article from 2018 (Baumann 2018), I gave an overview about all associations between scutacarids and other animal taxa published until May 2017, and the aim of the present work is to update this information by including all new articles released between May 2017 and September 2023 (and the occasional publication that had slipped my attention in my previous review article). Following the structure of Baumann (2018), new information about either new scutacarid species and/or new hosts is given in the text; further reports that simply list new combinations of scutacarid species and hosts and/or new geographical localities are listed only in the Annex. The Annex is ordered according to the hosts, giving information on the associated scutacarid species, the type of association (inquiline, phoront, or both), the geographic range and occasionally other interesting remarks (for example, other known habitats of the mites, like soil or moss).
The online supplementary material consists of a table listing the scutacarid species first and adding the reported host taxa for each species afterwards. While the Annex only contains the new information published between 2017 and 2023, the table in the supplementary material is an updated version of the one from 2018, thus containing all information about scutacarids and their hosts available to date.
Methods
The articles were gained by searching the term ′Scutacaridae′ for the period 2017-2023 in Google Scholar and Web of Science in September 2023, and by scanning the references in the respective articles for publications that I had missed before. In order to maintain clarity and to keep focus on the mites, author names are only provided for scutacarid species but not for their hosts.
Results and Discussion: New information about scutacarid species and their hosts
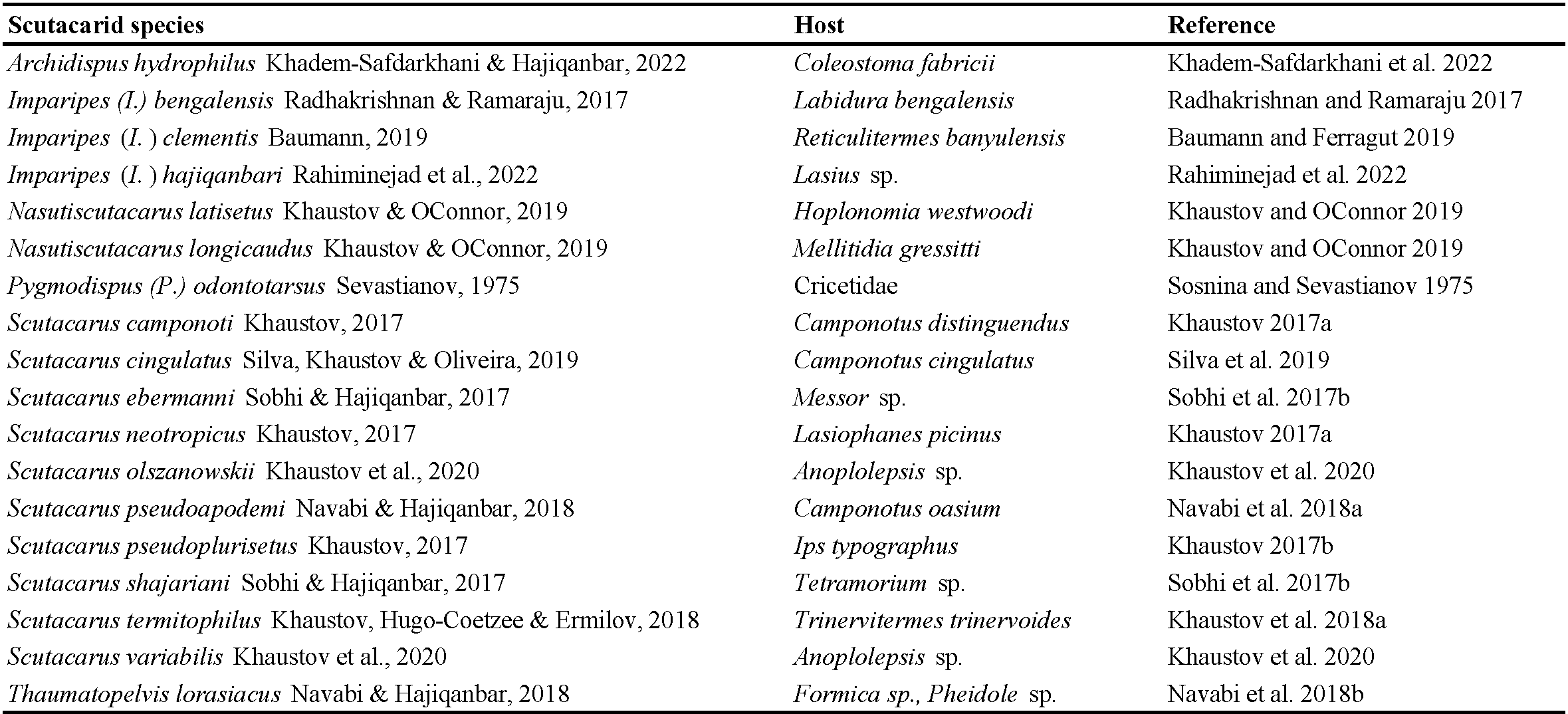
Between May 2017 and September 2023, 27 articles containing information about scutacarids as phoronts or inquilines have been published, dealing with mites from 15 countries (Algeria, Australia, Brazil, Chile, Egypt, India, Iran, Mexico, Papua New Guinea, Russia, South Africa, Spain, Sri Lanka, Ukraine and Uruguay). As result of the present literature study, 17 new scutacarid species described since 2017 and one species already described in 1975 can be added to the scutacarids found in association with other animal taxa (Table 1). Moreover, seven species already known to science have been recorded as phoronts or inquilines for the first time since 2017 (Table 2). Eight animal (sub)families, 18 genera and 28 species have been found as new host taxa for scutacarid mites.


Arachnids (Annex I)
Until now, only two species of Scutacarus had been reported in association with (nemesiid) spiders, and both had been encountered in South America (Baumann 2018). There has now been an interesting new finding of a scutacarid species phoretic on spiders: a member of Imparipes, most probably a new species, attached between coxae III and IV of the Australian cave-inhabiting anamid spider Troglodiplura beirutpakbarai (Marsh et al. 2023).
Hexapoda
Isoptera (Annex I)
Two new scutacarid species associated with termites have been described since Baumann (2018): Scutacarus termitophilus Khaustov, Hugo-Coetzee & Ermilov, 2018 on Trinervitermes trinervoides (Khaustov et al. 2018a), which is the first species of the genus Scutacarus found in association with termites, and Imparipes clementis Baumann, 2019 (Baumann and Ferragut 2019). The latter was found attached to the coxae of Reticulitermes banyulensis (Baumann et al. 2018, Baumann and Ferragut 2019). Termites could never be seen removing attached mites of I. clementis, even when they were self- or allogrooming. Attachment didn′t happen by claws on leg I, as is typical for Scutacaridae (Baumann 2018), but by large empodia on legs II and III (Baumann et al. 2018). This species was thus the first example of a scutacarid without claws on leg I that still can attach to a phoresy host, which indicates that -contrary to previous opinion- all other species without claws I might also be able to perform phoresy using their empodia. The attachment of the mites seemed to be weak as the mites detached after the slightest external disturbance, which might indicate that phoresy is restricted to the inside of the termite nest (Baumann et al. 2018).
Inside the termites' nests, I. clementis was found either on the termites or inside the tunnels that the termites created in dead wood, and they were located preferably near termite carcasses (Baumann et al. 2018, Baumann and Ferragut 2019). Only few scutacarid specimens could be found in the soil surrounding the nests. Imparipes clementis turned out to be easy to rear in the laboratory: lab cultures of the mites thrived on different substrates and larvae and males could be gained and described through those cultures (Baumann and Ferragut 2019). During those rearings, it became also clear that the number of phoretic scutacarids rose when the termite colony was in a bad state (identified as such by an increasing number of dead termites). In one instance, one female of I. clementis was observed performing appetence behavior (Baumann et al. 2018), being now the twelfth scutacarid species reported to show this behavioral trait.
Coleoptera (Annex II and Annex III)
Within Coleoptera, the by far largest number of associated scutacarids can be found in the family Carabidae (Baumann 2018). However, since 2017, only three scutacarid species have been reported from carabid beetles (see Annex II), and only one of these reports is worth mentioning here: Heterodispus turkmenistaniensis Khaustov & Chydyrov, 2005 was encountered in Iran on the beetle Scarites (Scarites) procerus eurytus (Sobhi et al. 2017a), and this is the first report of the carabid subfamily Scaritinae as hosts for Scutacaridae. The authors found H. turkmenistaniensis also on beetles belonging to the family Tenebrionidae, and it was also reported on Scarabaeidae (see below). Until now, this scutacarid species was only known from soil samples.
Two new species of Scutacaridae associated with two other beetle families have been described: Archidispus hydrophilus Khadem-Safdarkhani & Hajiqanbar, 2022 was found attached to Coleostoma fabricii (Hydrophiliidae; see Annex III) on its hosts sternite between the coxae (Khadem-Safdarkhani et al. 2022). This beetle feeds on decomposing material below wet rocks or among grass roots. This material supposedly is an ideal substrate for the growth of fungal mycelia, which in turn are a likely food source for the mite (Khadem-Safdarkhani et al. 2022). In the family Curculionidae (see Annex III), Scutacarus pseudoplurisetus Khaustov, 2017 was described from the wood borer Ips typographus (Khaustov 2017b and Khaustov et al. 2018b), a species which has already been noted as host for two scutacarid species before (Baumann 2018). The total number of scutacarid species associated with I. typographus now went up to five as Khaustov (2017b), and Khaustov et al. (2018b) also reported Scutacarus palustris Rack, 1966, a species also associated with ants and rodents (Baumann 2018), and Scutacarus muscocicola Sevastianov, 1983, which had until now only been reported from moss and litter, on this beetle. All five species associated with I. typographus belong to the genus Scutacarus.
In the family Scarabaeidae, three new genera have been reported as a new host for scutacarids: the genus Adoretus for Heterodispus turkmenistaniensis, the genus Hybalus for Pygmodispus odontotarsus Sevastianov, 1975 (Khaustov and Frolov 2018), and the genus Oryctes for Scutacarus longitarsus (Berlese, 1905) (Rahiminejad et al. 2020a, 2023).
All three scutacarid species appear to be generalists in terms of habitat and/or host range, as P. odontotarsus has already been reported from the nest of a small mammal and from soil, S. longitarsus has been found on dipterans and various families of coleoptera (Baumann 2018 and this publication) and H. turkmenistaniensis on different coleoptera families and from soil (this publication).
Imparipes (Sporichneuthes) intermedius Paoli, 1911, a species until now only known from soil and litter, has now also been encountered on different scarab beetles (Rahiminejad et al. 2020a)
Furthermore, six new beetle families have been identified as hosts for Scutacaridae: Cerambycidae, Dryopidae, Elateridae, Geotrupidae, Lucanidae and Meloidae (see Annex III).
In Cerambycidae, Scutacarus hystrichocentrus Sevastianov, 1983 was encountered on the beetle Paranda caspia by Hajiqanbar and Arjomandi (2019). This beetle attacks dead host plants which had already been infested by wood boring insects, and the authors conclude that the dead wood presumably is a suitable habitat for scutacarid mites. Until now, S. hystrichocentrus was only known as associate of ants of the genus Lasius (see Baumann 2018).
In the family Dryopidae, the genus Dryops has been reported as new host for Scutacarus sphaeroideus Karafiat, 1959 (Rahiminejad et al. 2020b, 2023). About 75% of dryopid species are aquatic (Khagani et al. 2022), but although no nearer information was available, it is most probable that S. sphaeroideus was attached to one of the remaining riparian or terrestrial species.
Beetles of the family Elateridae were identified as new hosts for Archidispus insolitus (Kurosa, 1974) and Imparipes (I.) moderatus Khaustov, 2008. Archidispus insolitus, until now only known from Carabidae (Baumann 2018), has also been found on Meloidae (see below), Tenebrionidae and Scarabaeidae (Rahiminejad et al. 2023). Imparipes (I.) moderatus was encountered on Drasterius bimaculatus by Navabi et al. (2018a). This scutacarid species was until now only known as associate of ants (Baumann 2018). Drasterius bimaculatus adults live on the ground of many different terrestrial habitats including riverside alluvial soils, whereas their larvae are rhizophagous (Gradinarov 2003, Hardersen et al. 2012). The high mobility of the adults as well as the rhizophagous lifestyle of the larvae most likely make these beetles an attractive host for scutacarids.
In Geotrupidae, Geotrupes spiniger was determined as host for Scutacarus longitarsus by Rahiminejad et al. (2020a). This species builds brood chambers that are filled with dung (Klemperer 1979), which most likely facilitates the growth of fungi and thus allows scutacarids to thrive there.
Lucanus ibericus of the family Lucanidae has also been newly reported as host for Scutacarus longitarsus (Rahiminejad et al. 2020a, 2023). The larvae of lucanid beetles live in decayed wood, a suitable habitat for scutacarids.
Finally, the family Meloidae has been listed as a new host for Archidispus insolitus (Rahiminejad et al. 2023). Adult beetles are mostly phytophagous, but their juvenile stages are often associated with solitary hymenopterans and complete their metamorphosis in soil (Ghahari & Campos-Soldini 2019), which could explain the association of the scutacarids with these beetles.
Hymenoptera: Bees (Annex IV)
Scutacaridae don′t play an important role as associates of the honey bee Apis mellifera (family Apidae). Still, the third scutacarid species after Imparipes apicola (Banks, 1914) and Scutacarus acarorum (Goeze, 1780) (Baumann 2018) was now encountered as inquiline of A. mellifera by Abd El Hady and Mahfouz (2021). They found Scutacarus evansi Momen & Curry, 1987 in bee hives in Egypt, albeit only in low abundance.
In different Bombus species (also family Apidae), Scutacarus acarorum can be found as a frequent associate. This mite species generally prefers hibernated queens as phoresy host (Baumann 2018). It has now also been found associated with Bombus bellicosus and B. pauloensis in Uruguay (Revainera et al. 2019), and although authors state that the associated acarofauna in general preferred queens as host, the prevalence of S. acarorum was highest in males of both Bombus species in the respective study. Another species, Imparipes (I.) rafalskii Dastych, 1978, was newly found associated with B. terrestris and also with the genus Andrena of the family Andrenidae (Sobhi et al. 2017a). This scutacarid was until now only known phoretic on the apid Dasypoda plumipes (Baumann 2018).
Nazari et al. (2019) found an Imparipes species phoretic on either the genus Andrena (Andrenidae) or on Antophora (Apidae); the real host is unclear because the text and table 2 of the respective publication are inconsistent with one another. If the host in fact is Antophora, this would be the first report of this genus as host for Scutacaridae.
Until now, only two species of the scutacarid genus Nasutiscutacarus were known, N. ampliatus Beer & Cross, 1960 and N. anthrenae Beer &Cross, 1960, both of which are phoretic on bees of the soil nesting family Halictidae (Baumann 2018). This number was now doubled by Khaustov and OConnor (2019), who described two new scutacarids associated with halictid bees: N. latisetus Khaustov & OConnor, 2019 from Sri Lanka and N. longicaudus Khaustov & OConnor, 2019 from Papua New Guinea. Nasutiscutacarus is characterized by an elongated gnathosoma which implies a highly specific, albeit still unknown, feeding source (Khaustov and OConnor 2019). Since all known species of Nasutiscutacarus are associated with Halictidae, the elongated gnathosoma most likely is an adaptation to special conditions in the hosts' nests. However, other scutacarid genera possessing typical scutacarid gnathosomata can also be encountered in association with Halictidae (Baumann 2018), which indicates that it must be a unique niche within halictid nests that is exploited by Nasutiscutacarus.
Hymenoptera: Ants (Annex V, Annex VI and Annex VII)
Most scutacarids in associations with other animals are associated with ants (Baumann 2018), therefore it is not surprising that 10 of the 18 new phoretic scutacarid species (given in Table 1) also are associated with ants. Scutacaridae belong to the superfamily Pygmephoroidea, and within the superfamily they might be best adapted to the association with ants, as can be concluded based on a study of the myrmecophilous pygmephoroid species in Western Siberia by Khaustov and Tolstikov (2016). The respective work revealed that more than 70% of the found species indeed belonged to the family Scutacaridae (the remaining species belonged to Neopygmephoridae and Microdispidae). The authors also found highly specific mite communities for most ant species, indicating a coevolution between mite lineages and ants.
Dolichoderinae, Ponerinae (Annex V)
In the subfamily Dolichoderinae, two new Tapinoma species have been reported as hosts for Scutacaridae. A not nearer identified Scutacarus species has been found associated with Tapinoma sessile, an invasive species occurring in natural and urban areas all over North America (Uppstrom 2010, Blumenfeld et al. 2021), and Imparipes comatus Mahunka, 1970 has been reported on Tapinoma tauridis from Iran (Sobhi et al. 2017a). However, T. tauridis is currently considered to be a junior synonym of T. erraticum (Bolton 1995, updated online 2020) which has been listed as host for I. comatus before (see Baumann 2018).
Until now, only one ant species (Ponera coarcata) of the subfamily Ponerinae has been known as host for Scutacaridae, and it has already been stated that intensified collection effort of these ants should increase the number of associated mites (Baumann 2018). Uppstrom (2010) effectively reported two not nearer identified Imparipes species and one Scutacarus species from Ponera pennsylvanica from North America, and Lopes et al. (2015) cite a Japanese book by Maruyama et al. (2013) who report a Scutacarus species attached to Brachyponera chinensis.
Rocha et al. (2020) also reported scutacarid mites phoretic on Neoponera villosa from Mexico; however, I was able to study the respective mites in December 2022 and they turned out not to be scutacarids, but members of the morphologically similar neopygmephorid genus Petalomium (Baumann unpublished).
The following new scutacarids have been described as associates of ant species belonging to the subfamily Formicinae (Annex VI): six Scutacarus species Thaumatopelvis lorasiacus Navabi & Hajiqanbar, 2018 and Imparipes hajiqanbari Rahiminejad et al., 2022. Moreover, I. lituanicus Lazauskene & Sevastianov, 1974, which was until now only known from forest litter, has been reported associated with Lasius sp. (Rahiminejad et al. 2022).
The genus Anoplolepsis was reported as host for scutacarids for the first time, specifically for Scutacarus olszanowskii Khaustov et al., 2020 and S. variabilis Khaustov et al., 2020. This case is interesting as the host ants were found in old termite nests of Trinervitermes trinervoides (Khaustov et al. 2020). Another new host genus was Lasiophanes, an endemic genus of Patagonia. Lasiophanes picinus, a terrestrial species that lives in relatively humid soils (Fergnani et al. 2008), was found as host species for S. neotropicus Khaustov, 2017 (Khaustov 2017b).
Thaumatopelvis lorasiacus was not only found on Formicinae, but also phoretic on ants of the genus Pheidole (Rahiminejad et al. 2022), which belongs to the subfamily Myrmicinae (Annex VII). Two more new Scutacarus were described associated with this subfamily: S. ebermanni Sobhi & Hajiqanbar, 2017 and S. shajariani Sobhi & Hajiqanbar, 2017 (Sobhi et al. 2017b).
Khaustov and Tolstikov (2016) and Silva et al. (2019) mentioned the attachment sites of several of the found scutacarid species in their papers (for details, see Annex V - VII). These authors also reported that the scutacarids could be found on workers of various ant species and also on winged ants (that is, sexual forms). In accordance with this finding, Imparipes hajiqanbari was also collected from alates of Lasius sp. (Rahiminejad et al. 2022).
Conclusion/closing remarks
As this review shows, knowledge about the scutacarid fauna associated with other animals has considerably increased through various publications since May 2017. However, almost all papers are restricted to describing new species or reporting associations between mites and their hosts, but rarely investigating further. This means that three of the four ''future perspectives''- behavioural aspects, cryptic species complexes, phoretomorphism, and scutacarid fauna- commented in my previous work (Baumann 2018) still have to be dealt with.
Regarding behavioural aspects, the studies about Imparipes clementis, which is associated with termites (Baumann et al. 2018, Baumann and Ferragut 2019), gave some new insights as they demonstrated that phoretic attachment is possible even without claws on leg I, that the hosts never tried to remove attached mites, that mites can be found in areas inside the termite nests where termite corpses are buried and that the number of phoretic mites increases when a colony is in a bad state (presumably so that the mites have a chance to leave the ''sinking ship'', the moribund nest). There is still a lot of information about behavioral aspects lacking, for example: How exactly is the phoretic behavior initiated, how is it stopped? What influence, if at all present (e.g. sanitary role), do the mites have on their hosts?
There haven′t been any new studies dealing with cryptic species complexes or phoretomorphism in Scutacaridae, but there was a study investigating the influence of the dispersal ability on phenotypic patterns. By comparing populations of a phoretic scutacarid species (with supposed high dispersal ability) with those of a presumed non-phoretic species (with supposed low dispersal ability), it was revealed that the non-phoretic species shows a clear phenotypic population structure compared to the homogenous phoretic species, caused most likely by differences in their gene-flow (Baumann 2021). This result supports the assumption that phoresy does not promote (allopatric) speciation as it, on the contrary, allows for panmixis of geographically separated populations (Seeman and Walter 2023).
Molecular genetic studies could further shed a light on questions regarding the population structures of phoretic scutacarids, possible cryptic species complexes and the mechanisms determining the phoretomorphism which is present in some species. However, molecular genetic analyses including scutacarids are still scarce and the respective studies don′t focus on Scutacaridae but only include them in comprehensive phylogenetic trees or studies on the diversity of acarofauna (e.g. Young et al. 2012, Klimov et al. 2018). The respective publications show that DNA extraction of scutacarids is feasible, but may require special handling (like the application of semidestructive methods) due to the small size of the mites.
ANNEX: LIST OF SCUTACARID SPECIES ASSOCIATED WITH DIFFERENT HOSTS
I. Scutacarid species associated with Ectognatha (excl. Coleoptera and Aculeata)
II. Scutacarid species associated with Carabidae (Hexapoda, Ectognatha, Coleoptera)
III. Scutacarid species associated with Coleoptera excl. Carabidae (Hexapoda, Ectognatha)
IV. Scutacarid species associated with Apoidea (Hexapoda, Ectognatha, Aculeata)
V. Scutacarid species associated with Formicidae excl. Formicinae and Myrmicinae (Hexapoda, Ectognatha, Aculeata, Vespoidea)
VI. Scutacarid species associated with Formicinae (Hexapoda, Ectognatha, Aculeata, Formicidae)
VII. Scutacarid species associated with Myrmicinae (Hexapoda, Ectognatha, Aculeata, Formicidae)
VIII. Scutacarid species associated with Mammalia
The following information will be given for each host in telegraphic style: associated scutacarid species; distribution area; type of association (i = inquiline, ip = inquiline and phoretic, nk = not known, p = phoretic); reference(s) and remarks (if available)
* indicates that taxon is recorded for the first time as host for Scutacaridae
I. SCUTACARID SPECIES ASSOCIATED WITH ARACHNIDA AND ECTOGNATHA (EXCL. COLEOPTERA AND ACULEATA)
Arachnida
Aranaea
Anamidae*: Troglodiplura* beirutpakbarai*
Imparipes sp.; Australia; p; Marsh et al. (2023)
Dermaptera
Forficulidae: Labidura* bengalensis*
Imparipes (I.) bengalensis Radhakrishnan & Ramaraju, 2017; India; p; Radhakrishnan & Ramaraju (2017); on the forewing
Isoptera
Rhinotermitidae: Reticulitermes banyulensis*
Imparipes (I.) clementis Baumann, 2019; Spain; ip; Baumann & Ferragut (2019); males and larvae were gained through laboratory cultures; between termites’ coxae
Termitidae: Trinervitermes trinervoides
Scutacarus termitophilus Khaustov, Hugo-Coetzee & Ermilov, 2018; South Africa; style="margin-left: 2em"ip; Khaustov et al. (2018a); phoretic on workers of termites
“Insecta”
Scutacarus sphaeroideus Karafiat, 1959; Iran; p; Hajizadeh & Hosseini (2022); found on insects attracted by light traps
II. SCUTACARID SPECIES ASSOCIATED WITH CARABIDAE (HEXAPODA, ECTOGNATHA, COLEOPTERA)
Harpalinae
Harpalus sp.
Archidispus minor Karafiat, 1959; Iran; p; Hajiqanbar & Arjomandi (2019)
Pterostichinae
Amara sp.
Archidispus szaboi Mahunka, 1977; Iran; p; Rahiminejad & Hajiqanbar (2020)
Scaritinae*
Scarites* (Scarites) procerus eurytus*
Heterodispus turkmenistaniensis Khaustov & Chydyrov, 2005; Iran; p; Msc thesis of Mortazavi (2010), cited in Sobhi et al. (2017a); new record as phoretic species (so far only recorded from soil under cucumbers)
III. SCUTACARID SPECIES ASSOCIATED WITH COLEOPTERA EXCL. CARABIDAE (HEXAPODA, ECTOGNATHA)
Cerambycidae*
Parandra* caspia*
Scutacarus hystrichocentrus Sevastianov, 1983; Iran; p; Hajiqanbar & Arjomandi (2019)
Curculionidae
Ips typographus
Scutacarus muscocicola Sevastianov, 1983; Russia; p; Khaustov (2017b), Khaustov et al. (2018b); new record as phoretic species (so far recorded from moss)
Scutacarus palustris Rack, 1966; Russia; p; Khaustov et al. (2018b)
Scutacarus pseudoplurisetus Khaustov, 2017; Russia; p; Khaustov (2017b), Khaustov et al. (2018b)
Scutacarus scolyti Mahunka & Moser, 1980; Russia; p; Khaustov (2017b), Khaustov et al. (2018b); attached between the coxae of the beetle
Elateridae*
Archidispus insolitus (Kurosa, 1974); Iran; p; Rahiminejad et al. (2023); under elytra or on mesothorax
Drasterius* bimaculatus*
Imparipes (I.) moderatus Khaustov, 2008; Iran; p; Navabi et al. (2018a)
Dryopidae*
Dryops* sp.
Scutacarus sphaeroideus Karafiat, 1959; Iran; p; Rahiminejad et al. (2020b, 2023)
Geotrupidae*
Geotrupes* spiniger*
Scutacarus longitarsus (Berlese, 1905); Iran; p; Rahiminejad et al. (2020a)
Hydrophilidae
Scutacarus sphaeroideus Karafiat, 1959; Iran; p; Rahiminejad et al. (2020b, 2023)
Coleostoma* fabricii*
Archidispus hydrophilus Khadem-Safdarkhani & Hajiqanbar, 2022; Australia; p; Khadem-Safdarkhani et al. (2022)
Lucanidae*
Lucanus* ibericus*
Scutacarus longitarsus (Berlese, 1905); Iran; p; Rahiminejad et al. (2020a, 2023)
Meloidae*
Archidispus insolitus (Kurosa, 1974); Iran; p; Rahiminejad et al. (2023); under elytra or on mesothorax
Scarabaeidae
Imparipes (Sporichneuthes) intermedius Paoli, 1911; Iran; p; Rahiminejad et al. (2020a); new record as phoretic species (so far collected from forest soil and litter)
Adoretus* persicus*
Heterodispus turkmenistaniensis Khaustov & Chydyrov, 2005; Iran; p; Rahiminejad et al. (2023)
Hybalus* sp.
Pygmodispus odontotarsus Sevastianov, 1975; Algeria; p; Khaustov & Frolov (2018);
Onthophagus sp.
Imparipes (Sporichneuthes) intermedius Paoli, 1911; Iran; p; Rahiminejad et al. (2020a); new record as phoretic species (so far collected from forest soil and litter)
Oryctes* nasicornis*
Scutacarus longitarsus (Berlese, 1905); Iran; p; Rahiminejad et al. (2020a, 2023)
Tenebrionidae
Archidispus insolitus (Kurosa, 1974); Iran; p; Rahiminejad et al. (2023); under elytra or on mesothorax
Blaps mortisaga*
Heterodispus turkmenistaniensis Khaustov & Chydyrov, 2005; Iran; p; Sobhi et al. (2017a); new record as phoretic species (so far only recorded from soil under cucumbers)
Gonocephalum pubiferum
Heterodispus turkmenistaniensis Khaustov & Chydyrov, 2005; Iran; p; Msc thesis of Loghmani (2013), cited in Sobhi et al. (2017a); new record as phoretic species (so far only recorded from soil under cucumbers)
IV. SCUTACARID SPECIES ASSOCIATED WITH APOIDEA (HEXAPODA, ECTOGNATHA, ACULEATA)
Apoidea
Andrena sp. (Andrenidae) or Anthophora sp.* (Apidae)
Imparipes sp.; Iran; p; Nazari et al. (2019); not clear if mites were found on Andrena sp. (as given in the text) or on Anthophora (as given in table 2); Anthophora would be first record of the genus as host for Scutacaridae
Apis mellifera
Scutacarus evansi Momen & Curry, 1987; i (?); Egypt; Abd El Hady & Mahfouz (2021); Soliman & Kandil are given as authors of S. evansi in the respective paper; new record as phoretic species (so far only recorded from grassland on reclaimed peat)
Bombus sp.
Scutacarus acarorum (Goeze, 1780); Iran; p; Hajiqanbar & Arjomandi (2019)
Bombus bellicosus*
Scutacarus acarorum (Goeze, 1780); Uruguay; p; Revainera et al. (2019)
Bombus pauloensis*
Scutacarus acarorum (Goeze, 1780); Uruguay; p; Revainera et al. (2019)
Bombus terrestris
Imparipes (I.) rafalskii Dastych, 1978; Ukraine; p; Sobhi et al. (2017a), citing an article in Russian by Zaloznaya & Khaustov (2007)
Imparipes sp. nr. parapicola Delfinado Baker & Abatiello, 1976; Iran; p(?); Rahiminejad et al. (2023); listed as “parasitic on mining bees”
Andrena sp.
Imparipes (I.) rafalskii Dastych, 1978; Iran; p; Sobhi et al. (2017a)
Hoplonomia* westwoodi*
Nasutiscutacarus latisetus Khaustov & OConnor, 2019; Sri Lanka; p; Khaustov & OConnor (2019)
Mellitidia* gressitti*
Nasutiscutacarus longicaudus Khaustov & OConnor, 2019; Papua New Guinea; p; Khaustov & OConnor (2019)
V. SCUTACARID SPECIES ASSOCIATED WITH FORMICIDAE EXCL. FORMICINAE AND MYRMICINAE (HEXAPODA, ECTOGNATHA, ACULEATA, VESPOIDEA)
Dolichoderinae
Tapinoma sessile*
Scutacarus sp.; USA; p; Uppstrom (2010)
Tapinoma tauridis*
Imparipes (I.) comatus Mahunka, 1970; Iran; p; Sobhi et al. (2017a); the ant species is currently considered a synonym of T. erraticum (Bolton 1995, updated online 2020)
Ponerinae
Brachyponera* chinensis*
Scutacarus sp.; Japan; p; Lopes et al. (2015), citing Maruyama et al. (2013); attached to the head of the ant
Ponera pennsylvanica*
two not further identified Imparipes species; USA; p; Uppstrom (2010)
one not further identified Scutacarus species; USA; p; Uppstrom (2010)
VI. SCUTACARID SPECIES ASSOCIATED WITH FORMICINAE (HEXAPODA, ECTOGNATHA, ACULEATA, FORMICIDAE)
Anoplolepsis* sp.
Scutacarus olszanowskii Khaustov et al., 2020; South Africa; p; Khaustov et al. (2020); ants in an old nest of Trinervitermes trinervoides termites
Scutacarus variabilis Khaustov et al., 2020; South Africa; p; Khaustov et al. (2020); ants in an old nest of Trinervitermes trinervoides termites
Camponotus sp.
Imparipes (I.) ignotus Khaustov & Chydyrov, 2004; Iran; p; Navabi et al. (2018a)
Imparipes (I.) malus Khaustov, 2008; Iran; p; Navabi et al. (2018a)
Camponotus chromaiodes*
Scutacarus sp.; USA; p; Uppstrom (2010)
Camponotus cingulatus*
Scutacarus cingulatus Silva, Khaustov & Oliveira, 2019; Brazil; ip; Silva et al. (2019); females found phoretic on ants, males and larvae (but no females) found in the nest
Camponotus distinguendus*
Scutacarus camponoti Khaustov 2017; Chile; ip; Khaustov (2017a); males found in the nest
Camponotus oasium*
Scutacarus pseudoapodemi Navabi & Hajiqanbar, 2018; Iran; p; Navabi et al. (2018a)
Imparipes (I.) morosus Khaustov, 2008; Iran; p; Navabi et al. (2018a)
Cataglyphis sp.
Scutacarus subquadratus Khaustov & Chydyrov, 2004; Iran; p; Sobhi et al. (2017a)
Formica sp.
Scutacarus avarus Khaustov, 2008; Iran; p; Navabi et al. (2018a)
Thaumatopelvis lorasiacus Navabi & Hajiqanbar, 2018; Iran; p; Navabi et al. (2018b)
Formica spp.
Imparipes sp.; USA; p; Uppstrom 2010
Imparipes (I.) obsoletus Rack, 1966; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) nescius Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) robustus Karafiat, 1959; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus atypicus Karafiat, 1959; Russia; nk (presumably ip); Khaustov & Tolstikov (2016); attached to the anterior surface of ant abdomen
3 not nearer identified Scutacarus spp.; USA; p; Uppstrom (2010)
Scutacarus carsticus Mahunka & Mahunka-Papp, 1980; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus pilatus Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus rotundus (Berlese, 1903); Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus subterraneus (Oudemans, 1913); Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Formica fusca
Scutacarus karafiati Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Formica rufa
Scutacarus (tackei?) suborbiculatus Rack, 1964; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)*Formica sanguinea
Scutacarus pseudospinosus Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Lasiophanes* picinus*
Scutacarus neotropicus Khaustov, 2017; Chile; p; Khaustov (2017a)
Lasius sp.
Imparipes (I.) hajiqanbari Rahiminejad et al., 2022; Iran; p; Rahiminejad et al. (2022); found on alates
Lasius spp.
5 not nearer identified
Imparipes spp.; USA; p; Uppstrom (2010)
Imparipes (I.) histricinus Berlese, 1903; Iran; p; Rahiminejad et al. (2022)
Imparipes (I.) lituanicus Lazauskene & Sevastianov, 1974; Iran; p; Rahiminejad et al. (2022); new record as phoretic species (so far only found in forest litter)
Imparipes (I.) obsoletus Rack, 1966; Russia, Iran; p; Khaustov & Tolstikov (2016), Rahiminejad et al. (2022)
Imparipes (I.) sklyari Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
17 not nearer identified
Scutacarus spp.; USA; p; Uppstrom (2010)
Scutacarus heterotrichus Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus lasiophilus Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016); on lateral parts of ant abdomen
Scutacarus longisetus (Berlese, 1903); Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus pseudospinosus Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016); attached to tibia of leg I of Lasius niger
Scutacarus spinosus Štorkán, 1936¸ Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Lasius alienus
Diveripes sp.; USA; p; Uppstrom (2010); attached to the ant head
Pygmodispus sp.; USA; p; Uppstrom (2010); attached to the ant head
Lasius flavoniger*
Imparipes (I.) sevastianovi Khaustov, 2008; Iran; p; Hajiqanbar & Arjomandi (2019)
Lasius flavus
Imparipes (I.) brevitarsus Ebermann, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus insolitus Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus molnari Mahunka, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016); on lateral parts of ant abdomen
Scutacarus moseri Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus ponticulus Mahunka, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus sibiriensis Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016); attached to the ant head
Scutacarus tutus Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Lasius fuliginosus
Imparipes (I.) brevibasis (Sevastianov, 1983); Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) fuliginosophilus Khaustov, 2016; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) sevastianovi Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus flexisetus Karafiat, 1959; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus heterotrichus Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Lasius niger
Lophodispus irregularis (Mahunka, 1971); Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus carsticus Mahunka & Mahunka-Papp, 1980; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus crinitus Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus ellipticus Karafiat, 1959; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus expectatus Karafiat, 1959; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus hauseri Mahunka, 1977; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus hystrichocentrus Sevastianov, 1983; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus (gratus?) hortobagyensis Mahunka, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus kassaii Mahunka, 1965; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus velutinosus Sevastianov, 1983¸ Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Lasius obscuratus*
Imparipes (I.) placidus Khaustov & Chydyrov, 2004; Iran; p; Sobhi et al. (2017a)
Lasius umbratus
Scutacarus aequalis Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus flexisetosimilis Khaustov, 2016; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus levis Mahunka, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016); new record as phoretic species (so far reported from grass sample)
Scutacarus velutinosus Sevastianov, 1983; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
VII. SCUTACARID SPECIES ASSOCIATED WITH MYRMICINAE (HEXAPODA, ECTOGNATHA, ACULEATA, FORMICIDAE)
Aphaenogaster spp.
2 not nearer identified Imparipes spp.; USA; p; Uppstrom (2010)
4 not nearer identified Scutacarus spp.; USA; p; Uppstrom (2010)Messor sp.
Imparipes (I.) histricinus Berlese, 1903; Iran; p; Sobhi et al. (2017a)
Scutacarus ebermanni Sobhi & Hajiqanbar, 2017; p; Iran; Sobhi et al. (2017b)
Scutacarus remissus Khaustov, 2008; Iran; p; Sobhi et al. (2017a)
Scutacarus shivicki Lazauskene & Sevastianov, 1974; Iran; p; Sobhi et al. (2017a); new record as phoretic species (so far reported from soil and litter)
Messor caducus*
Imparipes (I.) gagati Khaustov, 2008; Iran; p; Navabi et al. (2018a)
Myrmica ruginodis
Imparipes (I.) charkoviensis Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) comatosimilis Metwali, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus ellipticus Karafiat, 1959; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus hystrichocentrus Sevastianov, 1983; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus kassaii Mahunka, 1965; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus latus Karafiat, 1959; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus myrmecophilus Metwali, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus myrmicinus Khaustov, 2015; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus ovoideus Karafiat, 1959; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus pseudospinosus Khaustov, 2008¸ Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus spinosus Štorkán, 1936¸ Russia; nk (presumably ip); Khaustov & Tolstikov (2016); on lateral parts of ant abdomen
Scutacarus subterraneus (Oudemans, 1913); Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Pheidole sp.
Thaumatopelvis lorasiacus Navabi & Hajiqanbar, 2018; Iran; p; Navabi et al. (2018b)
Solenopsis molesta*
2 not nearer identified Imparipes spp.; USA; p; Uppstrom (2010)
Tetramorium sp.
Imparipes (I.) imaginatus Mahunka, 1981; Iran; p; Sobhi et al. (2017a)
Imparipes (I.) lentus Khaustov, 2008; Iran; p; Sobhi et al. (2017a)
Imparipes (I.) malus Khaustov, 2008; Iran; p; Navabi et al. (2018a)
Imparipes (I.) placidus Khaustov & Chydyrov, 2004; Iran; p; Sobhi et al. (2017a)
Imparipes (I.) tenuis Mahunka, 1981; Iran; p; Sobhi et al. (2017a)
Scutacarus shajariani Sobhi & Hajiqanbar, 2017; Iran; p; Sobhi et al. (2017b)
Tetramorium caespitum
Imparipes sp.; USA; p; Uppstrom (2010)
Imparipes (I.) parapicosimilis Metwali, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) histricinus Berlese, 1903; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) imaginatus Mahunka, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) lentus Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Imparipes (I.) hortobagyensis Mahunka, 1981; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Lophodispus irregularis (Mahunka, 1971); Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus claviger (Paoli, 1911); Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
Scutacarus pseudospinosus Khaustov, 2008; Russia; nk (presumably ip); Khaustov & Tolstikov (2016)
VIII. SCUTACARID SPECIES ASSOCIATED WITH MAMMALIA
Cricetidae
Pygmodispus odontotarsus Sevastianov, 1975; Ukraine; i; Sosnina & Sevastianov (1975)
acarologia_4673_SupplementaryReferences.pdf
acarologia_4673_SupplementaryTable.pdf
References
- Abd El Hady M.A.H., Mahfouz H.M. 2021. Species composition and diversity of mites associated with honeybees Apis mellifera L in the Egyptian apiaries. Arch. Agri. Sci. J., 4 (1): 148-155. https://doi.org/10.21608/aasj.2021.182108
- Baumann J. 2018. Tiny mites on a great journey- a review on scutacarid mites as phoronts and inquilines (Heterostigmatina, Pygmephoroidea, Scutacaridae). Acarologia, 58(1): 192-251. https://doi.org/10.24349/acarologia/20184238
- Baumann J. 2021. Patterns of intraspecific morphological variability in soil mites reflect their dispersal ability. Exp. Appl. Acarol., 83: 241-255. https://doi.org/10.1007/s10493-020-00587-y
- Baumann J., Ferragut F. 2019. Description and observations on morphology and biology of Imparipes clementis sp. nov., a new termite associated scutacarid mite species (Acari, Heterostigmatina: Scutacaridae; Insecta, Isoptera: Rhinotermitidae). Syst. Appl. Acarol., 24(2): 303-323. https://doi.org/10.11158/saa.24.2.12
- Baumann J., Ferragut F., Šimić S. 2018. Lazy hitchhikers? Preliminary evidence for within-habitat phoresy in pygmephoroid mites (Acari, Scutacaridae). Soil Org., 90(3): 95-99. https://doi.org/10.25674/mzmh-9h78
- Blumenfeld A.J., Eyer P.-A., Helms A.M., Buczkowski G., Vargo E.L. 2021. Consistent signatures of urban adaptation in a native, urban invader ant Tapinoma sessile. Mol. Ecol., 31(18): 4832-4850. https://doi.org/10.1111/mec.16188
- Bolton, B. 1995. A New General Catalogue of the Ants of the World. Cambridge, MA: Harvard University Press. Updated online 2020 as An Online Catalogue of the Ants of the World, https://www.antwiki.org/wiki/Online_Catalogue_of_the_Ants_of_the_World (accessed 22.06.2023).
- Fergnani P., Sackmann P., Cuezzo F. 2008. Environmental determinants of the distribution and abundance of the ants, Lasiophanes picinus and L. valdiviensis, in Argentina. J. Insect Sci., 8:36. https://doi.org/10.1673/031.008.3601
- Ghahari H., Campos-Soldini MP. 2019. An annotated catalogue of blister-beetles (Coleoptera: Tenebrionoidea: Meloidae) of Iran. Entomofauna, 40/1 (5): 59-138.
- Gradinarov D. 2003. New Natural Hosts of Entomopathogenic Nematodes (Rhabditida: Steinernematidae, Heterorhabditidae) from Bulgaria. Acta Zool. Bulg., 55 (3): 59-64.
- Hajiqanbar H., Arjomandi E. 2019. Heterostigmatic mites (Acari: Trombidiformes: Prostigmata) associated with Coleoptera and Hymenoptera in Mazandaran province, northern Iran. Pers. J. Acarol., 8 (4): 343-352. https://doi.org/10.22073/pja.v8i4.56685
- Hajizadeh J., Hosseini R. 2022. Introduction of mites collected from light traps in Rasht city. Plant Pests Res., 12(1): 1-13. doi: 10.22124/iprj.2022.5602 https://doi.org/10.3389/fpls.2021.783578
- Hardersen S., Toni I., Cornacchia P., Curletti G., Leo P., Nardi G., Penati F., Piatella E., Platia G. 2012. Survey of selected beetle families in a floodplain remnant in northern Italy. Bull. Insectol., 65 (2): 199-207.
- Khadem-Safdarkhani H., Hajiqanbar H., Riegler M., Seeman O., Katlav A. 2022. Two New Phoretic Species of Heterostigmatic Mites (Acari: Prostigmata: Neopygmephoridae and Scutacaridae) on Australian Hydrophilid Beetles (Coleoptera: Hydrophilidae). Insects, 13: 483. https://doi.org/10.3390/insects13050483
- Khaghani R., Vatandoost H., Tashakori G., Jalilian A. 2022. Bio Ecology of Aquatic and Semi-Aquatic Insects Of Order Coleoptera In The World. J. Mar. Sci. Res. Oceanogr., 5 (3): 157-178 https://doi.org/10.33140/JMSRO.05.03.05
- Khaustov A.A. 2017a. Two new species of myrmecophilous scutacarid mites (Acari: Scutacaridae) from Chile. Syst. Appl. Acarol., 22 (1): 115-124. https://doi.org/10.11158/saa.22.1.12
- Khaustov A.A. 2017b. A new species and new records of pygmephoroid mites (Acari: Scutacaridae, Neopygmephoridae, Pygmephoridae) associated with Ips typographus (Coleoptera: Curculionidae: Scolytinae) from Western Siberia, Russia. Int. J. Acarol., 43 (8): 594-602. https://doi.org/10.1080/01647954.2017.1390608
- Khaustov A.A., Frolov A.V. 2018. A new species, new genus and new records of heterostigmatic mites (Acari: Heterostigmata) phoretic on scarab beetles of the subfamily Orphninae (Coleoptera: Scarabaeidae). Zootaxa, 4514 (2): 181-201. https://doi.org/10.11646/zootaxa.4514.2.3
- Khaustov A.A., OConnor B.M. 2019. Two new species of Nasutiscutacarus (Acari: Heterostigmata: Scutacaridae) associated with halictid bees (Hymenoptera: Halictidae) from Papua New Guinea and Sri Lanka. Int. J. Acarol., 44 (8): 386-394. https://doi.org/10.1080/01647954.2018.1529055
- Khaustov A.A., Tolstikov A.V. 2016. The diversity, mite communities, and host specificity of pygmephoroide mites (Acari: Pygmephoroidea) associated with ants in Western Siberia, Russia. Acarina, 24 (2): 113-136. DOI: 10.21684/0132-8077-2016-24-2-113-136 https://doi.org/10.21684/0132-8077-2016-24-2-113-136
- Khaustov A.A., Hugo-Coetzee E.A., Ermilov S. 2018a. A new species of Scutacarus (Acari: Heterostigmatina: Scutacaridae) associated with Trinervitermes trinervoides (Isoptera: Termitidae) from South Africa. Int. J. Acarol., 44 (2-3): 59-67. https://doi.org/10.1080/01647954.2018.1450444
- Khaustov A.A., Klimov P.B., Trach V.A., Bobylev A.N., Salavatulin V.M., Khaustov V.A., Tolstikov A.V. 2018b. Review of mites (Acari) associated with the European Spruce Bark Beetle, Ips typographus (Coleoptera: Curculionidae: Scolytinae) in Asian Russia. Acarina, 26 (1): 3-79. DOI: 10.21684/0132-8077-2018-26-1-3-79 https://doi.org/10.21684/0132-8077-2018-26-1-3-79
- Khaustov A.A., Hugo-Coetzee E.A., Ermilov S. 2020. Two new myrmecophilous Scutacarus (Acari: Scutacaridae) from South Africa. Ann. Zool., 70 (3): 397-407. https://doi.org/10.3161/00034541ANZ2020.70.3.011
- Klemperer H.G. 1979. An analysis of the nesting behaviour of Geotrupes spiniger Marsham (Coleoptera, Scarabaeidae). Ecol. Entomol., 4: 133-150. https://doi.org/10.1111/j.1365-2311.1979.tb00569.x
- Klimov P.B., OConnor B.M., Chetverikov P.E., Bolton S.J., Pepato A.R., Mortazavi A.L., Tolstikov A.V., Bauchan G.R., Ochoa R.. 2018. Comprehensive phylogeny of acariform mites (Acariformes) provides insights on the origin of the four-legged mites (Eriophyoidea), a long branch. Mol. Phylogenet. Evol., 119: 105-117. https://doi.org/10.1016/j.ympev.2017.10.017
- Loghmani A. 2013. Heterostigmatic mites (Acari: Heterostigmatina) associated with insects in some parts of Northeastern Iran [MSc Thesis]. Iran, Tehran: Tarbiat Modares University, 102pp., p. 76.
- Lopes J.M.S., Oliveira A.R., Delabie J.H.C. 2015. Interações formigas/ácaros, com ênfase em ácaros foréticos associados a poneromorfas. In: Delabie, J.H.C. et al., orgs. As formigas poneromorfas do Brasil. Ilhéus, BA: Editus: 375-387. https://doi.org/10.7476/9788574554419.0025
- Maruyama M., Komatsu T., Kudo S., Shimada T., Kinomura K. 2013. The Guests of Japanese Ants. Minamiyana, Japan: Tokai UniversityPress. xii + 208 p
- Marsh J.R., Milner S.J., Shaw M., Stempel A.J., Harvey M.S., Rix M.G. 2023. A Case for Below-Ground Dispersal? Insights into the Biology, Ecology and Conservation of Blind Cave Spiders in the Genus Troglodiplura (Mygalomorphae: Anamidae). Insects, 14: 449. https://doi.org/10.3390/insects14050449
- Mortazavi A. 2010. Prostigmatic mites (Acari:Prostigmata) associated with Coleoptera in some regions of Kerman province, Iran [MSc Thesis]. Iran, Tehran: Tarbiat Modares University, 146pp. p. 98.
- Navabi A., Hajiqanbar H., Mortavazi A. 2018a. New species and records of the genera Scutacarus and Imparipes (Acari: Heterostigmatina: Scutacaridae) associated with Coleoptera and Hymenoptera (Arthropoda: Insecta) from Iran. Zootaxa, 4531 (4): 532-540. https://doi.org/10.11646/zootaxa.4531.4.4
- Navabi A., Hajiqanbar H., Mortavazi A. 2018b. First record of the genus Thaumatopelvis Mahunka, 1973 (Acari: Prostigmata: Scutacaridae) from Asia with description of a new species associated with ants (Hymenoptera: Formicidae). Syst. Appl. Acarol., 23 (2): 360-366. https://doi.org/10.11158/saa.23.2.11
- Nazari S., Monfared A., Nemati A., Azhari S. 2019. A survey on bees (Insecta, Hymenoptera, Apoidea) and their associated mites in Chaharmahal and Bakhtiari province of Iran. J. Insect Biodivers. Syst., 5 (2): 107-120. https://doi.org/10.52547/jibs.5.2.107
- Radhakrishnan V., Ramaraju K. 2017. First record on Imparipes bengalensis sp. nov. (Acari: Scutacaridae) on Labidura bengalensis Dohrn (Forficulidae: Dermaptera) from Tamil Nadu, India. J. Entomol. Zool. Stud., 5 (6): 1606-1611.
- Rahiminejad V., Hajiqanbar H. 2020. New records of mites of the Heterostigmata (Acari: Prostigmata) associated with insects from Golestan Province, northern Iran. Pers. J. Acarol., 9 (3): 233-242. https://dx.doi.org/10.22073/pja.v9i3.61223
- Rahiminejad V., Hajiqanbar H., Talebi A. A. 2020a. An alpha diversity survey of Heterostigmatic mites (Trombidiformes, Prostigmata) phoretic on scarabaeoid beetles in Hycrani forest, northern Iran. Syst. Appl. Acarol., 25 (11): 2033-2046. https://doi.org/10.11158/saa.25.11.9
- Rahiminejad V., Seyedein S., Nadimi A. 2020b. New records of soil dwelling mites of the superfamily Pygmephoroidea (Acari: Heterostigmatina) from northern Iran. J. Insect Biodivers. Syst., 6 (3): 307-315. https://doi.org/10.52547/jibs.6.3.307
- Rahiminejad V., Nadimi A., Seyedein S. 2022. Contribution to the knowledge of the genus Imparipes Berlese (Acari: Heterostigmata: Scutacaridae) associated with ants in Iran. Zootaxa, 5133 (4): 585-593. https://doi.org/10.11646/zootaxa.5133.4.9
- Rahiminejad V., Nadimi A., Afshari, A. 2023. A catalog of heterostigmatic mites (Acari: Trombidiformes) of Iran. Persian J. Acarol., 12 (2): 259-313. https://doi.org/10.22073/pja.v12i2.78237
- Revainera P.D., Salvarrey S., Santos E., Arbulo N., Invernizzi C., Plischuk S., Abrahamovich A., Maggi M.D. 2019. Phoretic mites associated to Bombus pauloensis and Bombus bellicosus (Hymenoptera Apidae) from Uruguay. J. Apic. Res., 5 (3): 455-462. https://doi.org/10.1080/00218839.2018.1521775
- Rocha F.H., Lachaud J.-P., Pérez-Lachaud G. 2020. Myrmecophilous organisms associated with colonies of the ponerine ant Neoponera villosa (Hymenopter: Formicidae) nesting in Aechmea bracteata bromeliads : a biodiversity hotspot. Myrm. News, 30: 73-92. doi: 10.25849/myrmecol.news_030:073
- Seeman O.D., Walter D.E. 2023. Phoresy and Mites: More than just a free ride. Ann. Rev. Entomol., 68: 69-88. https://doi.org/10.1146/annurev-ento-120220-013329
- Silva R.A., Khaustov A.A., Delabie J.H.C., Oliveira A.R. 2019. A new species of Scutacarus and description of larva and male of Petalomium megasolenidiatum Silva, Khaustov & Oliveira (Acari: Heterostigmatina: Neopygmephoridae, Scutacaridae) from Brazil. Syst. Appl. Acarol., 24 (8): 1343-1362. https://doi.org/10.11158/saa.24.8.2
- Sobhi M., Hajiqanbar H., Mortazavi A. 2017a. A contribution to the knowledge of scutacarid mites (Acari: Pygmephoroidea: Scutacaridae) associated with Coleoptera and Hymenoptera (Arthropoda: Insecta) from northwestern Iran. Acarologia, 57 (4): 1103-1111. DOI: 10.24349/acarologia/20174222 https://doi.org/10.24349/acarologia/20174222
- Sobhi M., Hajiqanbar H., Mortazavi A. 2017b. Two new myrmecophilous species of the genus Scutacarus (Acari: Prostigmata: Scutacaridae) with world keys to related species groups. Entomol. Sci., 20: 292-301. https://doi.org/10.1111/ens.12255
- Sosnina E.F., Sevastianov V.D. 1975. Mites of cohort Tarsonemina (Trombidiformes) from the nests of small mammals in the forest zone of the Crimean Mountains. Rev. Entomol. URSS, 1: 202-214.
- Uppstrom K.A. 2010. Mites (Acari) Associated with the Ants (Formicidae) of Ohio and the Harvester Ant, Messor pergandei, of Arizona. MscThesis, The Ohio State University, 228pp. http://rave.ohiolink.edu/etdc/view?acc_num=osu1279217236
- Young M.R., Behan-Pelletier V.M., Hebert P.D.N. 2012. Revealing the Hyperdiverse Mite Fauna of Subarctic Canada through DNA Barcoding. PLoS One, 7 (11): e48755. https://doi.org/10.1371/journal.pone.0048755
- Zaloznaya L.I., Khaustov A.A. 2007. First find of the mite Imparipes (Imparipes) rafalskii (Acari, Scutacaridae) in Ukraine. Vestn. Zool., 41(3): 266 [in Russian]



2023-10-16
Date accepted:
2023-12-01
Date published:
2023-12-14
Edited by:
Pfingstl, Tobias

This work is licensed under a Creative Commons Attribution 4.0 International License
2024 Baumann, Julia
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)