



Unguizetes (Knorozovia) ershovae (Acari, Oribatida, Mochlozetidae), a new oribatid mite subgenus and species from Mexico
Ermilov, Sergey G.  1
and Yurtaev, Andrey A.
2
1
and Yurtaev, Andrey A.
2
1✉ Tyumen State University, Institute of Environmental and Agricultural Biology (X-BIO), Tyumen, Russia.
2Tyumen State University, Institute of Environmental and Agricultural Biology (X-BIO), Tyumen, Russia.
2023 - Volume: 63 Issue: 4 pages: 1187-1196
https://doi.org/10.24349/r0vq-kaq5ZooBank LSID: 7D3FB050-7F63-4EAB-9279-84166B14DC0D
Original research
Keywords
Abstract
Introduction
The oribatid mite family Mochlozetidae (Acari, Oribatida) comprises 13 supraspecies taxa (11 genera and two subgenera) with 79 species having a cosmopolitan distribution collectively. Many representatives of the family are arboreal but some species are registered in soil-litter (e.g., Norton 1983, 1984; Corpuz-Raros and Ermilov 2019, 2020).
Unguizetes Sellnick, 1925, with type species Oribata sphaerula Berlese, 1905, is the largest genus of Mochlozetidae. Its taxonomic revision has been presented by Ermilov (2016). During faunistic study of oribatid mites collected from Mexico, we found a new species belonging to the new subgenus of Unguizetes. The main goal of our paper is to describe Unguizetes (Knorozovia) n. subgen., with U. (K.) ershovae n. sp as type species, based on adults.
Also, we present an identification key to the known genera/subgenera of Mochlozetidae using morphological traits and supraspecies system mostly from the systematic and revision taxonomic papers (e.g., Grandjean 1959, 1960; Norton 1983, 1984; Norton and Behan-Pelletier 2009; Schatz et al. 2011; Ermilov 2016; Ermilov and Corpuz-Raros 2017; Ermilov and Friedrich 2017).
Presently, five Mochlozetidae genera were found in Mexico (Palacios-Vargas and Iglesias 2004; Vázquez-González et al. 2016): Dynatozetes Grandjean, 1960; Mahunkazetes Balogh and Balogh, 1992; Mochloribatula Mahunka, 1978; Mochlozetes Grandjean, 1930; and Uracrobates Balogh and Mahunka, 1967.
Methods
Observation and documentation
For measurement and illustration, specimens were mounted in lactic acid on temporary cavity slides. All measurements are in micrometers. Body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the notogaster; other structures we oriented to avoid parallax errors. Notogastral width refers to the maximum of the ventral plate in ventral aspect and on level of the pteromorphs in dorsal aspect. Setal lengths were measured perpendicular to their long axis, accounting for curvature. Formulas for leg solenidia are given in square brackets according to the sequence genu-tibia-tarsus. Drawings were made with a camera lucida using a Leica DM 2500 transmission light microscope. For SEM microscopy alcohol preserved mites were dusted with silver and scanned with the aid of a JEOL–JSM-6510LV SEM microscope.
Terminology
Morphological terminology used in this paper follows that of Grandjean (Grandjean 1959, 1960), Norton (1983, 1984), Ermilov (2016, 2017); also Norton (1977) for leg setal nomenclature and Norton and Behan-Pelletier (2009) for overview are used.
Abbreviations
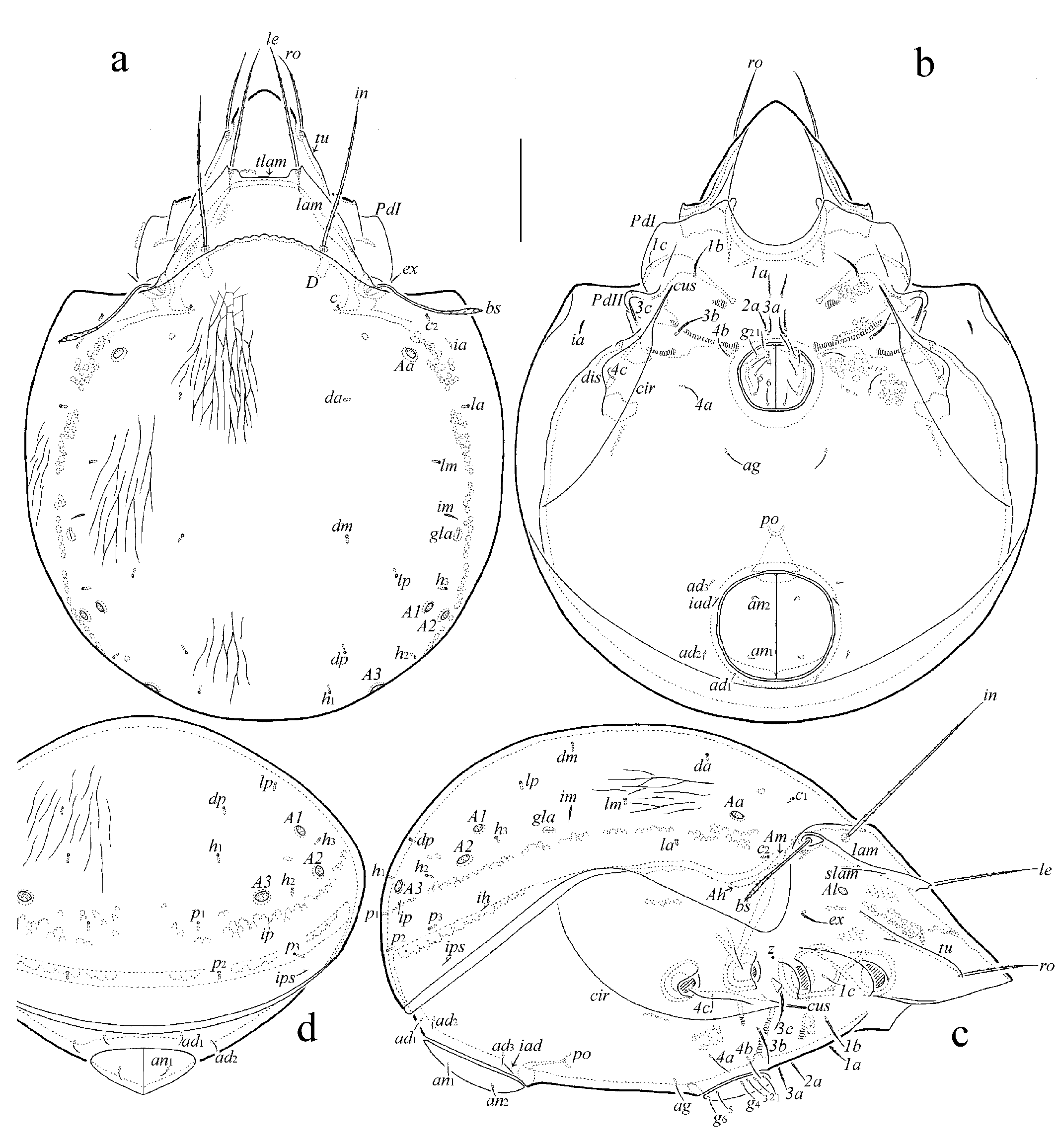
Prodorsum: lam = lamella; tlam = translamella; slam = sublamella; Al = sublamellar porose area; tu = tutorium; ro, le, in, bs, ex = rostral, lamellar, interlamellar, bothridial, and exobothridial setae, respectively; D = dorsophragma. Notogaster: c, da, la, dm, lm, dp, lp, h, p = notogastral setae; Aa, A1, A2, A3 = porose areas; ia, im, ip, ih, ips = lyrifissures; gla = opisthonotal gland opening. Gnathosoma: a, m, h = subcapitular setae; or = adoral seta; d, l, cm, acm, ul, su, lt, vt, inf, sup = palp setae; ω = palp solenidion; cha, chb = cheliceral setae; Tg = Trägårdh's organ. Epimeral and lateral podosomal regions: 1a, 1b, 1c, 2a, 3a, 3b, 3c, 4a, 4b, 4c = epimeral setae; Am, Ah = humeral porose areas; PdI, PdII = pedotecta I, II, respectively; cus = custodium; dis = discidium; cir = circumpedal carina; z = aperture of supracoxal gland. Anogenital region: g, ag, an, ad = genital, aggenital, anal, and adanal setae, respectively; iad = adanal lyrifissure; po = preanal organ. Legs: Tr, Fe, Ge, Ti, Ta = trochanter, femur, genu, tibia, and tarsus, respectively; pa = porose area; ω, σ, φ = solenidia; ɛ = famulus; d, l, v, ev, bv, ft, tc, it, p, u, a, s, pv, pl = setae.
Taxonomy
Family Mochlozetidae Grandjean, 1960
Genus Unguizetes Sellnick, 1925, p. 473
Type species: Oribata sphaerula Berlese, 1905, by original designation, p. 171
Unguizetes (Knorozovia) n. subgen.
ZOOBANK: 82B48A2A-3D97-4E3F-991F-A8AAF25ACEB6 ![]()
Type species: Unguizetes (Knorozovia) ershovae n. sp.
Adult — With character states of Unguizetes (see Ermilov 2016) but with 14 pairs of notogastral setal alveoli and well developed, complete dorsosejugal suture.
Remarks — The new subgenus is similar to the nominate subgenus in many morphological traits, but differs from the latter by the subgeneric traits (14 pairs of notogastral setal alveoli; dorsosejugal suture strong, complete versus 10 pairs of notogastral setal alveoli; dorsosejugal suture interrupted or indistinct medially).
Also, the new subgenus is similar to Gephyrazetes (Gephyrazetes) Hirauchi, 1999 in the presence of 14 pairs of notogastral setae and complete dorsosejugal suture, but differs from the latter by the notogastral setae represented by alveoli (versus setae medium-sized, setiform), the localization of notogastral porose area A1 in dorsomedial position (versus A1 in lateroperipheral position), long tutorium (versus short), and in having six pairs of genital setae (versus four or five pairs).
Etymology — The subgenus is named after Yuri Valentinovich Knorozov (1922–1999), a brilliant Russian scholar in the American studies, who deciphered the Mayan hieroglyphic writing in the middle of the 20th century. Knorozov was a historian, ethnographer, linguist, a staunch supporter of systemic and interdisciplinary research, the creator of the theory of ethnic semiotics and the theory of communication as the basis of collective activity. He repeatedly visited Guatemala and Yucatan, studying the culture and the origin of Native American languages, especially the Mayan. In addition, the zoo environment, which is an important part of the ethno-cultural context, has always been a special interest of Yu.V. Knorozov.
Unguizetes (Knorozovia) ershovae n. sp.
ZOOBANK: DA26D698-E8B0-4FE5-AAE3-8669ED54D294 ![]()
(Figures 1–4)



Material examined — Holotype (female) and five paratypes (three males and two females): Mexico, 20°41′N, 87°03′W, Quintana Roo, Municipio de Solidaridad, vicinities of Playa del Carmen, leaf litter under trees and bushes in secondary semi-evergreen tropical forest (date and collector unknown; collection of the Tyumen State University Museum of Zoology, Tyumen, Russia; Gashev et al. 2005).
Type deposition — The holotype is deposited in the collection of the Senckenberg Museum of Natural History, Görlitz, Germany; five paratypes are deposited in the collection of the Tyumen State University Museum of Zoology, Tyumen, Russia. All specimens are preserved in 70% solution of ethanol with a drop of glycerol.
Diagnosis — Body length: 585–690. Notogaster distinctly striate, partially forming reticulate ornamentation. Cusp of lamella with lateral tooth; translamella straight, comparatively broad; tutorium without cusp, directed to insertion of rostral seta. Rostral, lamellar and interlamellar setae long, setiform, barbed; ro shortest, in longest; bothridial seta long, with slightly developed, fusiform, barbed head. Dorsosejugal suture undulate. Pteromorph well developed. Notogastral porose areas rounded; A1 and A2 located close to each other. Fourteen pairs of notogastral setae as alveoli. Palp setae lt′, lt″, vt′, vt″ on tarsus and l′, l″ on genu short. Custodium long, narrowly triangular. Discidium broadly triangular. Epimeral, genital and aggenital setae medium-sized to comparatively short, setiform, slightly barbed; anal and adanal setae minute, setiform, roughened. Adanal lyrifissure located close and slightly diagonal to anal plate.
Description of adult — Measurements – Body length: 690 (holotype), 585, 600, 630 (male paratypes), 675, 690 (female paratypes); notogaster width (level of pteromorphs): 510 (holotype), 435, 450, 465 (male paratypes), 495, 510 (female paratypes); ventral plate width: 480 (holotype), 420, 435, 435 (male paratypes), 465, 480 (female paratypes).
Integument (Figs 1a, 1c, 1d, 3c, 3d, 4b) – Body color brown. Body and legs covered by thin layer of gel-like cerotegument; lateral side of body partially with microgranulate cerotegument. Surface densely microfoveolate (visible in dissected specimens under high magnification, 10 × 100); lamella entirely slightly striate (Fig. 3c); notogaster distinctly longitudinally striate (Fig. 4b), forming in anterior part reticulate ornamentation (Fig. 3d).
Prodorsum (Figs 1a, 1c, 3a–3d, 4a) – Rostrum broadly rounded. Lamella (including cusp) about 1/2 length of prodorsum, with short cusp having distinct lateral tooth; translamella straight, comparatively broad; tutorium about 1/2 length of prodorsum, lamelliform, without cusp. Sublamellar porose area oval (9–11 × 6–7). Rostral (75–82), lamellar (120–127) and interlamellar (165–176) setae setiform, barbed; ro inserted on tutorial end; bothridial seta (109–116) with long stalk and short, slightly developed, fusiform, barbed head; exobothridial seta (37–45) setiform, thin, barbed. Dorsosejugal porose area not observed.
Notogaster (Figs 1a, 1c, 1d, 3a, 3b, 4a) – Dorsosejugal suture strong, undulate. Pteromorph broadly rounded laterally. Four pairs of rounded porose areas (11–15); A1 and A2 located comparatively close to each other, both in lateral position but A1 anteromedial to A2. Fourteen pairs of notogastral setae represented by alveoli. Opisthonotal gland opening and all lyrifissures distinct.
Gnathosoma (Figs 2a–c) – Subcapitulum size: 150–161 × 97–101; subcapitular (a, h: 22–26; m: 41–49) and adoral (11) setae setiform, thin, barbed. Palp length: 116–120; setation: 0–2–1–3–9(+ω); setae (lt), (vt) on tarsus and (l) on genu short; postpalpal seta (6) spiniform, roughened. Chelicera length: 172–187; setae (cha: 52–56; chb: 32–34) setiform, barbed.
Epimeral and lateral podosomal regions (Figs 1b, 1c, 4a) – Epimeral setal formula: 3–1–3–3; all setae (3c: 37–45; 1b, 3b: 34–41; 1a, 2a, 3a, 4a, 4b: 26–30; 1c, 4c: 13–15) setiform, thin, slightly barbed; 1c, 4a, 4b, 4c thinner than others. Humeral porose areas Am and Ah elongate oval, slightly visible (Fig. 1c). Custodium long, narrowly triangular. Discidium broadly triangular. Circumpedal carina long, distally fused to custodium.
Anogenital region (Figs 1b, 1c) – Genital and aggenital setae (22–26) setiform, thin, slightly barbed; anal and adanal setae (11) setiform, thin, roughened. Adanal lyrifissure located close and slightly diagonal to anterior half of anal plate, posterior to ad3. Marginal porose area not observed.
Legs (Figs 2e, 2d, 4a, 4c) – Generally, typical for Unguizetes (e.g., Ermilov 2016, 2017). Median claw distinctly thicker than lateral claws, all slightly barbed dorsally; each lateral claw with small tooth ventrodistally. Dorsoparaxial porose area on femora I–IV, trochanters III, IV, ventrodistal porose area on tibia I–IV and proximoventral porose area on tarsi I–IV well visible. Formulas of leg setation and solenidia: I (1–5–3–4–20) [1–2–2], II (1–5–3–4–15) [1–1–2], III (2–3–1–3–15) [1–1–0], IV (1–2–2–3–12) [0–1–0]; homology of setae and solenidia indicated in Table 1.


Etymology — The species is named in honor of Galina Gavrilovna Ershova, a world-renowned Russian historian, anthropologist and epigrapher specializing in the study of the ancient civilizations, cultures, and languages of the New World. She is a disciple of Yu.V. Knorozov and currently continues to successfully develop his scientific ideas.
Discussion
Subías (2022, online version 2023) includes 10 genera and three subgenera in Mochlozetidae. He considers the genus Drymobatoides Jacot, 1936, with additional subgenus Rykella Balogh, 1962 as a representative of the family Drymobatidae and supported an independence of the subgenus Unguizetes (Calugarella) Balogh and P. Balogh, 1992.
The main family trait of Drymobatidae is the presence of both porose areas and saccules on the notogaster (see Balogh and Balogh 1984; Norton and Behan-Pelletier 2009). The representatives of Drymobatoides have only porose areas; therefore, we support the placement of the genus within Mochlozetidae. Also, earlier, we (Ermilov 2016; Ermilov and Corpuz-Raros 2017) explained the reasons why we do not support subgeneric statuses of Calugarella and Rykella.
Key to the known supraspecies taxa of Mochlozetidae
1. Posterior margin of notogaster triangularly narrowed or truncate
...... 2
— Posterior margin of notogaster broadly rounded
...... 3
2. Posterior margin of notogaster triangularly narrowed
...... Uracrobates (Uracrobates) Balogh and Mahunka, 1967
— Posterior margin of notogaster truncate
...... Uracrobates (Parauracrobates) Ermilov and Martens, 2015
3. All notogastral setae medium-sized, setiform; dorsosejugal suture complete
...... 4
— All notogastral setae minute, needle-form or represented by alveoli; dorsosejugal suture usually interrupted medially, rarely, complete
...... 6
4. All or some notogastral porose areas shortly band-like; notogastral porose area A1 in dorsomedial position; translamella absent
...... Nesiotizetes Jacot, 1934
— All notogastral porose areas rounded/oval; notogastral porose area A1 in lateroperipheral position; translamella present
...... 5
5. Notogaster with 14 pairs of setae
...... Gephyrazetes (Gephyrazetes) Hirauchi, 1999
— Notogaster with 10 pairs of setae
...... Gephyrazetes (Oligogephyrazetes) Subías, 2019
6. Notogaster with 20–50 pairs of minute porose areas; dorsosejugal suture complete
...... Drymobatoides Jacot, 1936
— Notogaster with less than 20 pairs of well-developed porose areas; dorsosejugal suture usually interrupted medially, rarely, complete
...... 7
7. Notogastral porose area Aa divided, represented by two areas
...... 8
— Notogastral porose area Aa not divided, represented by one area
...... 11
8. Genital plate with five setae; all notogastral setae minute, needle-form
...... 9
— Genital plate with six setae; all notogastral setae represented by alveoli
...... 10
9. Translamella absent; tutorium present; pteromorph developed; notogaster usually with two pairs of posteroperipheral porose areas
...... Mochlobates Norton, 1984
— Translamella present; tutorium absent; pteromorph reduced; notogaster usually with more than two pairs of posteroperipheral porose areas
...... Mochloribatula Mahunka, 1978
10. Both part of notogastral porose area Aa distant from each other, one part in dorsomedial position, the other part in lateroperipheral position; all notogastral porose areas rounded/oval
...... Mahunkazetes Balogh and Balogh, 1992
— Both part of notogastral porose area Aa located close to each other, in lateroperipheral position; all or some notogastral porose areas shortly band-like
...... Mochlozetes Grandjean, 1930
11. Notogaster with five to eight pairs of porose areas, of them, three to six pairs in posteroperipheral position; translamella absent
...... Dynatozetes Grandjean, 1960
— Notogaster with four pairs of porose areas, of them, two pairs (A2, A3) in posteroperipheral position; translamella usually present, rarely, absent
...... 12
12. Notogaster with 14 pairs of setae; dorsosejugal suture complete
...... Unguizetes (Knorozovia) n. subgen.
— Notogaster with 10 pairs of setae; dorsosejugal suture interrupted medially
...... 13
13. Notogastral porose area A1 in dorsomedial position
...... Unguizetes (Unguizetes) Sellnick, 1925
— Notogastral porose area A1 in lateroperipheral position
...... Podoribates Berlese, 1908
Acknowledgements
We thank Dr. Julia Baumann (University of Graz, Graz, Austria) and two anonymous reviewers for valuable comments; and Nikita Shulaev (Tyumen State University, Tyumen, Russia) for SEM micrographs. This research was partially supported by the cooperative agreement No. FEWZ-2021-0004 from the Russian Ministry of Science and Higher Education.
References
- Balogh J. 1962. Mission zoologique de l'I.R.S.A.C. en Afrique Orientale (P. Basilewsky et N. Leleup, 1957). LXXV. - Acari, Oribates. Ann. Mus. Roy. Afr. Centr. in-8°, Zool., 110: 90-131.
- Balogh J., Balogh P. 1984. A review of the Oribatuloidea Thor, 1929 (Acari: Oribatei). Acta Zool. Hung., 30: 257-313.
- Balogh J., Balogh P. 1992. The oribatid mite genera of the World. Vol. 1. Budapest: Hung. Nat. Mus. Press. 263 p.
- Balogh J., Mahunka S. 1967. New oribatids (Acari) from Vietnam. Acta Zool. Acad. Sci. Hung., 13: 39-74.
- Berlese A. 1905. Acari nuovi. Manipulus IV (Acari di Giava). Redia, 2: 154-176.
- Berlese A. 1908. Elenco di generi e specie nuove di Acari. Redia, 5: 1-15.
- Corpuz-Raros L., Ermilov S.G. 2019. Catalogue of oribatid mites (Acari: Oribatida) from the Malay Archipelago. Zootaxa, 4716: 1-240. https://doi.org/10.11646/zootaxa.4716.1.1
- Corpuz-Raros L., Ermilov S.G. 2020. Catalogue of oribatid mites (Acari: Oribatida) from Continental Southeast Asia. Zootaxa, 4893: 1-216. https://doi.org/10.11646/zootaxa.4893.1.1
- Ermilov S.G. 2016. Contribution to the knowledge of the oribatid mite genus Unguizetes (Acari, Oribatida, Mochlozetidae). Syst. Appl. Acarol., 21: 1281-1299 https://doi.org/10.11158/saa.21.10.1
- Ermilov S.G. 2017. Supplementary description of Unguizetes clavatus Aoki, 1967 (Acari, Oribatida, Mochlozetidae) on the basis of Vietnamese specimens. Acarina, 25: 97-107. https://doi.org/10.21684/0132-8077-2017-25-2-97-107
- Ermilov S.G., Corpuz-Raros L. 2017. New data on the Philippine oribatid mite fauna, with a contribution to knowledge of the genus Drymobatoides (Acari, Oribatida, Mochlozetidae). Acarologia, 57: 295-308. https://doi.org/10.1051/acarologia/20174156
- Ermilov S.G., Friedrich S. 2017. Contribution to knowledge of the oribatid mite genus Dynatozetes (Acari, Oribatida, Mochlozetidae). Spixiana, 40: 23-28. https://doi.org/10.11158/saa.23.4.6
- Ermilov S.G., Martens J. 2015. The oribatid mite genus Uracrobates (Acari, Oribatida, Mochlozetidae). Syst. Appl. Acarol., 20: 188-194. https://doi.org/10.11158/saa.20.2.5
- Gashev S.N., Elifanov A.V., Tolstikov A.V. 2005. Zoological Museum of the Tyumen State University as the oldest center for studying biodiversity in the Tyumen District. Bul. Tyumen State Univ., 3: 177-189.
- Grandjean F. 1930. Oribates nouveaux de la région Caraїbe. Bul. Soc. Zool. France, 55: 262-284.
- Grandjean F. 1959. Sur le genre Mochlozetes Grandj. 1930 (Oribate). Acarologia, 1: 452-474.
- Grandjean F. 1960. Les Mochlozetidae n. fam. (Oribates). Acarologia, 2: 101-148.
- Hirauchi Y. 1999. Some new taxa of the family Ceratozetidae (Oribatida) from the Tateyama Mountains, Centr. Japan. J. Acarol. Soc. Japan, 8: 103-116. https://doi.org/10.2300/acari.8.103
- Jacot A.P. 1934. Some Tyroglyphina (Sarcoptiformes) of the Marquesas Islands. Bul. Bernice P. Bishop Mus., Honolulu, 114: 211-238.
- Jacot A.P. 1936. Undescribed Mossmites from Mauritius. Ann. Mag. Nat. Hist. (Ser. 10), 17: 393-402. https://doi.org/10.1080/00222933608655134
- Mahunka S. 1978. Neue und interessante Milben aus dem Genfer Museum XXIV. First contribution to the fauna of the Dominican Republic (Acari: Oribatida). Redia, 61: 551-564.
- Norton R.A. 1977. A review of F. Grandjean's system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal D.L. (Ed.). Biology of oribatid mites. Syracuse: SUNY Col. Environ. Sci. Forestry. pp. 33-61.
- Norton R.A. 1983. Redefinition of Mochloribatula (Acari: Mochlozetidae), with new species, recombinations, and notes on plant associations. Acarologia, 24: 449-464.
- Norton R.A. 1984. Notes on Nathan Banks' and Henry Ewings' species of Mochlozetidae (Acari: Sarcoptiformes) with the proposal of a new genus. Acarologia, 25: 397-406.
- Norton R.A., Behan-Pelletier V.M. 2009. Oribatida. In: Krantz G.W., Walter D.E. (Eds). A Manual of Acarology (TX). Lubbock: Texas Univ. Press. Chapter 15. pp. 430-564.
- Palacios-Vargas J.G., Iglesias R. 2004. Oribatei (Acari). In: Bousquets J.L., Morrone J.J., Ordoñez O.Y., Fernández I.V. (Eds). Biodiversidad, Taxonomía y Biogeografía de Artrópodos de México: Hacia una síntesis de su conocimiento. México, D.F.: Facul. Cien., Univ. Nac. Autón. México. Vol. IV. pp. 431-468.
- Schatz H., Behan-Pelletier V.M., OConnor B.M., Norton R.A. 2011. Suborder Oribatida van der Hammen, 1968. In: Zhang, Z.-Q. (Ed.). Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148: 141-148. https://doi.org/10.11646/zootaxa.3148.1.26
- Sellnick M. 1925. Javanische Oribatiden. Treubia, 6: 459-475.
- Subias L.S. 2019. Nuevas adiciones al listado mundial de ácaros oribátidos (Acari, Oribatida) (14ª actualización). Rev. Ibér. Aracnol., 34: 76-80.
- Subías L.S. 2022. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles). Monogr. Electr. S.E.A., 12: 1-538.
- Subías L.S. 2023. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles), 18ª actualización: 1-540. Available from: http://bba.bioucm.es/cont/docs/RO_1.pdf (accessed February 2023).
- Vázquez-González M.M., May Uicab D.A., Alamilla-Pastrana E.B. 2016. Riqueza específica y biodiversidad de Cozumel, Quintana Roo, México. Teoría y Praxis, 19: 137-171. https://doi.org/10.22403/UQROOMX/TYP19/07



2023-06-22
Date accepted:
2023-11-09
Date published:
2023-11-15
Edited by:
Baumann, Julia

This work is licensed under a Creative Commons Attribution 4.0 International License
2023 Ermilov, Sergey G. and Yurtaev, Andrey A.
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)