



The tribes Kampimodromini Kolodochka and Typhlodromipsini Chant & McMurtry (Mesostigmata: Phytoseiidae) in the Serra do Espinhaço, Brazil, with a key to the Brazilian species of Typhlodromips De Leon
Ferragut, Francisco  1
and Navia, Denise
2
1
and Navia, Denise
2
1✉ Instituto Agroforestal Mediterráneo. Universitat Politècnica de València. Camino de Vera, s/n. 46022 Valencia, Spain.
2CBGP, INRAE, CIRAD, Institut Agro, IRD, Univ Montpellier, Montpellier, France.
2022 - Volume: 62 Issue: 4 pages: 1217-1253
https://doi.org/10.24349/mi5x-xnneZooBank LSID: 65746D20-782A-4753-8FB6-1937C723D12D
Original research
Keywords
Abstract
Introduction
This paper represents the second contribution to the phytoseiid fauna of the Serra do Espinhaço (Espinhaço range) in eastern Brazil, after the first paper dealing with species of the genus Neoseiulus Hughes (Ferragut and Navia, 2022). The objective is to report on the collected phytoseiid mites of the genera Neoparaphytoseius Chant & McMurtry and Proprioseius Chant in the tribe Kampimodromini Kolodochka, and species of the genus Typhlodromips De Leon in the tribe Typhlodromipsini Chant & McMurtry.
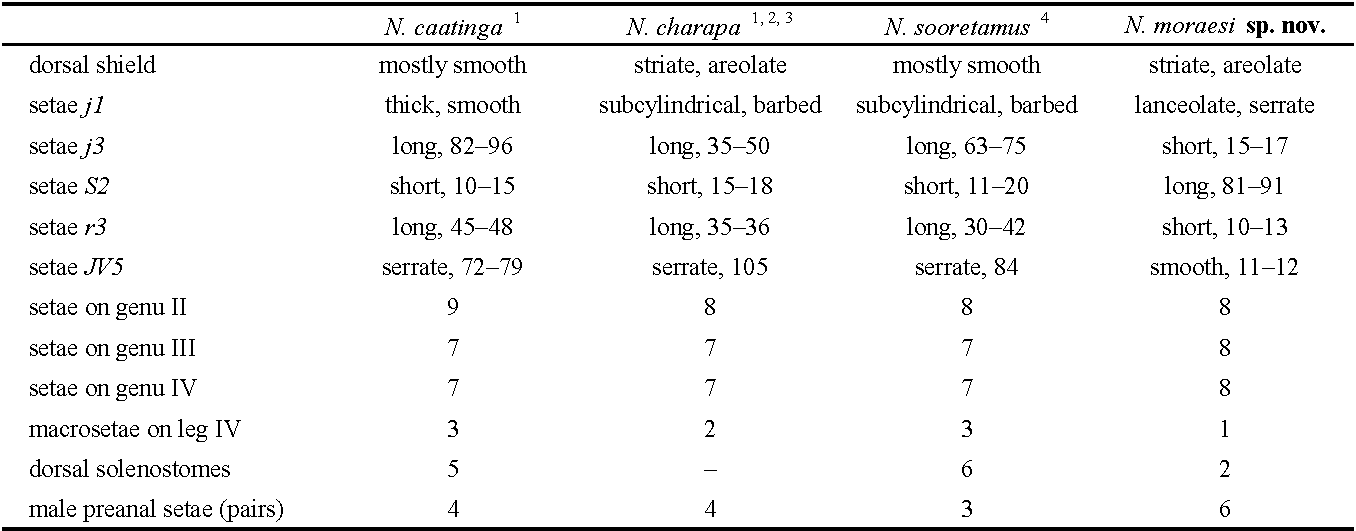
The tribe Kampimodromini is poorly represented in Brazil, where only six species have been reported from the 238 valid species described or cited in the country, three belonging to the genus Proprioseius, two to Neoparaphytoseius, and one to Paraphytoseius Swirski & Schechter (Demite et al. 2022). The genus Neoparaphytoseius was created by Chant and McMurtry (2005a) to accommodate Amblyseius sooretamus El-Banhawy (original designation), a peculiar species restricted to Brazil. Inaccuracies in the original description led to Chant and McMurtry to include the genus in the subtribe Paraphytoseiina by having a supposed indentation or notch in the lateral margins of the dorsal shield, at the level of setae s4. Later, Jiménez et al. (2014) examined the holotype, paratypes and additional specimens collected in Brazil, concluding that the indentation does not exist, and only a shallow concavity can be observed. They redefined the genus and suggested its inclusion within the subtribe Kampimodromina. During 30 years the genus was considered monotypic, but recently two additional species from Perú and Brazil were added (Jiménez et al. 2014; Silva et al. 2021). We described here a new species of Neoparaphytoseius, N. moraesi Ferragut sp. nov., distinctly different from the known species by the strongly tuberculate dorsal setae j1, s4, S2, Z4 and Z5, which provide the appearance of some Proprioseius, by the different length and morphology of the dorsal setae and caudoventral setae JV5, and by having most of them covered by a very characteristic and dense layer of crystals. We also fully redescribe the female and describe for the first time the male of Proprioseius gibbus Moraes & Denmark, a very peculiar species only known from the state of Bahia in Brazil, which displays one of the strangest habitus within the Phytoseiidae.
Compared with other Amblyseiinae, the genus Typhlodromips, from the tribe Typhlodromipsini, is a relatively morphological homogeneous group with a general appearance Neoseiulus-like, multidentate cheliceral fixed digits, macrosetae at least on legs III (not always obvious) and IV, and a characteristic, marked and rounded concavity in the lateral margins of the dorsal shield, at the level of setae R1, which gives the dorsal view of some species a peanut-like appearance. Twenty-three species have been reported from Brazil (Demite et al. 2022) which accounts 23.5% of the world species of the genus. Species distinction is not easy in some cases, as they have been recognized based mostly on morphological continuous characters, like the length of dorsal setae and leg macrosetae. Furthermore, old original descriptions are often not very helpful as they include few characteristics and illustrations are poor and do not show the complexity of phytoseiid morphology. We suspect that a considerable proportion of synonymies exist among the described species. The examination of the four species found in this study allow us to give a full complementary description of T. ariri Gondim Jr. & Moraes and T. japi Lofego, Demite & Feres females. We also provide a full redescription of T. constrictatus (El-Banhawy) describing for the first time the structure of the female insemination apparatus and the male of the species, and we propose the new species T. ovametapodalis Ferragut sp. nov., distinctive by having an oval-shaped posterior metapodal shield. Finally, we also provide a key to the separation of the Brazilian species of the genus.
Material and methods
Field surveys were conducted between January 2010 and January 2012 from native and spontaneous vegetation along the Serra do Espinhaço (Espinhaço Range) in the Brazilian states of Bahia and Minas Gerais. Information on the climatic, edaphic, and vegetation characteristics of the studied area can be found in Ferragut and Navia (2022).
Phytoseiid mites were collected by the authors and extracted from plants by examining the leaves under a stereomicroscope or by beating parts of the plant on a fine sieve. Specimens were mounted and clarified in Heinze-PVA medium and stored in an oven at 50 °C until total clarification. After clarification the phytoseiids were examined at 400x and 1000x magnification using differential interference contrast (DIC) in a compound microscope Nikon Eclipse Ni-U (Nikon Corporation, Tokyo, Japan). Illustrations were performed on a graphic tablet using Concepts software (https://concepts.app ![]() ). Images were captured with a digital camera Nikon DMX 1200 (Nikon Corporation, Tokyo, Japan) and edited in LightRoom CC and Photoshop CC (© Adobe Systems Inc.). Measurements of morphological traits were obtained with the software NIS-Elements D 3.1 and are given in the text in micrometers (μm). Measurements of the main morphological characters were made as follows: dorsal shield length from the anterior to posterior shield margins along the midline, and its width between lateral margins at the level of setae s4; length of peritremes from the stigma opening to the apex; length of the genital shield from the anterior margin of the hyaline surface to the posterior margin of the shield; ventrianal shield length as the distance between anterior and posterior margins along the midline; ventrianal shield width was measured between the lateral margins of the shield at the level of setae ZV2 and at level of anus between the margins of the shield and measuring at the level of the sockets of perianal setae; the cheliceral fixed digit length was measured from dorsal lyrifissure to the tip; the movable digit from basal articulation to tip of the digit. In the description of the new species, each measurement in females corresponds to the holotype (in bold) followed by the average and the ranges (in parentheses) corresponding to all specimens studied (holotype + paratypes). In the males and immatures of the new species and in the immatures, males and females of already known species, measurements correspond to the average of all examined specimens followed by the respective ranges in parentheses if measurements were variable.
). Images were captured with a digital camera Nikon DMX 1200 (Nikon Corporation, Tokyo, Japan) and edited in LightRoom CC and Photoshop CC (© Adobe Systems Inc.). Measurements of morphological traits were obtained with the software NIS-Elements D 3.1 and are given in the text in micrometers (μm). Measurements of the main morphological characters were made as follows: dorsal shield length from the anterior to posterior shield margins along the midline, and its width between lateral margins at the level of setae s4; length of peritremes from the stigma opening to the apex; length of the genital shield from the anterior margin of the hyaline surface to the posterior margin of the shield; ventrianal shield length as the distance between anterior and posterior margins along the midline; ventrianal shield width was measured between the lateral margins of the shield at the level of setae ZV2 and at level of anus between the margins of the shield and measuring at the level of the sockets of perianal setae; the cheliceral fixed digit length was measured from dorsal lyrifissure to the tip; the movable digit from basal articulation to tip of the digit. In the description of the new species, each measurement in females corresponds to the holotype (in bold) followed by the average and the ranges (in parentheses) corresponding to all specimens studied (holotype + paratypes). In the males and immatures of the new species and in the immatures, males and females of already known species, measurements correspond to the average of all examined specimens followed by the respective ranges in parentheses if measurements were variable.
The generic concepts are those proposed by Chant and McMurtry (2003, 2005a). Idiosomal setal nomenclature follows that of Lindquist and Evans (1965) as adapted by Rowell et al. (1978) for the dorsal surface and Chant and Yoshida-Shaul (1991) for the ventral surface of the idiosoma. Leg chaetotaxy follows Evans (1963) and Rowell and Chant (1979). Nomenclature for dorsal solenostomes and poroids is that proposed by Athias-Henriot (1975) updated by Beaulieu and Beard (2018). Terminology for the morphology of the spermatodactyl follows that of Beard (2001). World distribution of each species is based on Demite et al. (2022).
Holotype and some paratypes of the new species and specimens of other species were deposited at the Mite collection of the ESALQ-USP (Escola Superior de Agricultura ''Luiz de Queiroz″, Universidade de São Paulo, Piracicaba, State of São Paulo, Brazil). Other specimens of the species collected were deposited in the Laboratory of Acarology, Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Spain, and the Mite collection of the Centre de Biologie pour la Gestion des Populations (UMR CBGP), Montferrier sur Lez, France.
Results
Subfamily Amblyseiinae Muma, 1961
Tribe Kampimodromini Kolodochka, 1998
Genus Neoparaphytoseius Chant & McMurtry, 2003
Neoparaphytoseius moraesi Ferragut sp. nov.
ZOOBANK: 1021AC24-7E6F-4148-BA6C-EC357DCA540F ![]()
Type material
Female holotype, ten females, four males, and two deutonymph paratypes on Anemia villosa (Anemiaceae); road between Mucugê and Andaraí, Chapada Diamantina, rupestrian grasslands, 12°53′49″S, 41°18′58″W, 1125 m asl (above sea level); 19 January 2012.
Holotype, seven females, and one male paratypes deposited in the Mite collection of the ESALQ-USP (Escola Superior de Agricultura ''Luiz de Queiroz″, Universidade de São Paulo, Piracicaba, State of São Paulo, Brazil). Other specimens in the Laboratory of Acarology, Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Spain, and the Mite collection of the Centre de Biologie pour la Gestion des Populations (UMR CBGP), Montferrier sur Lez, France.
Diagnosis
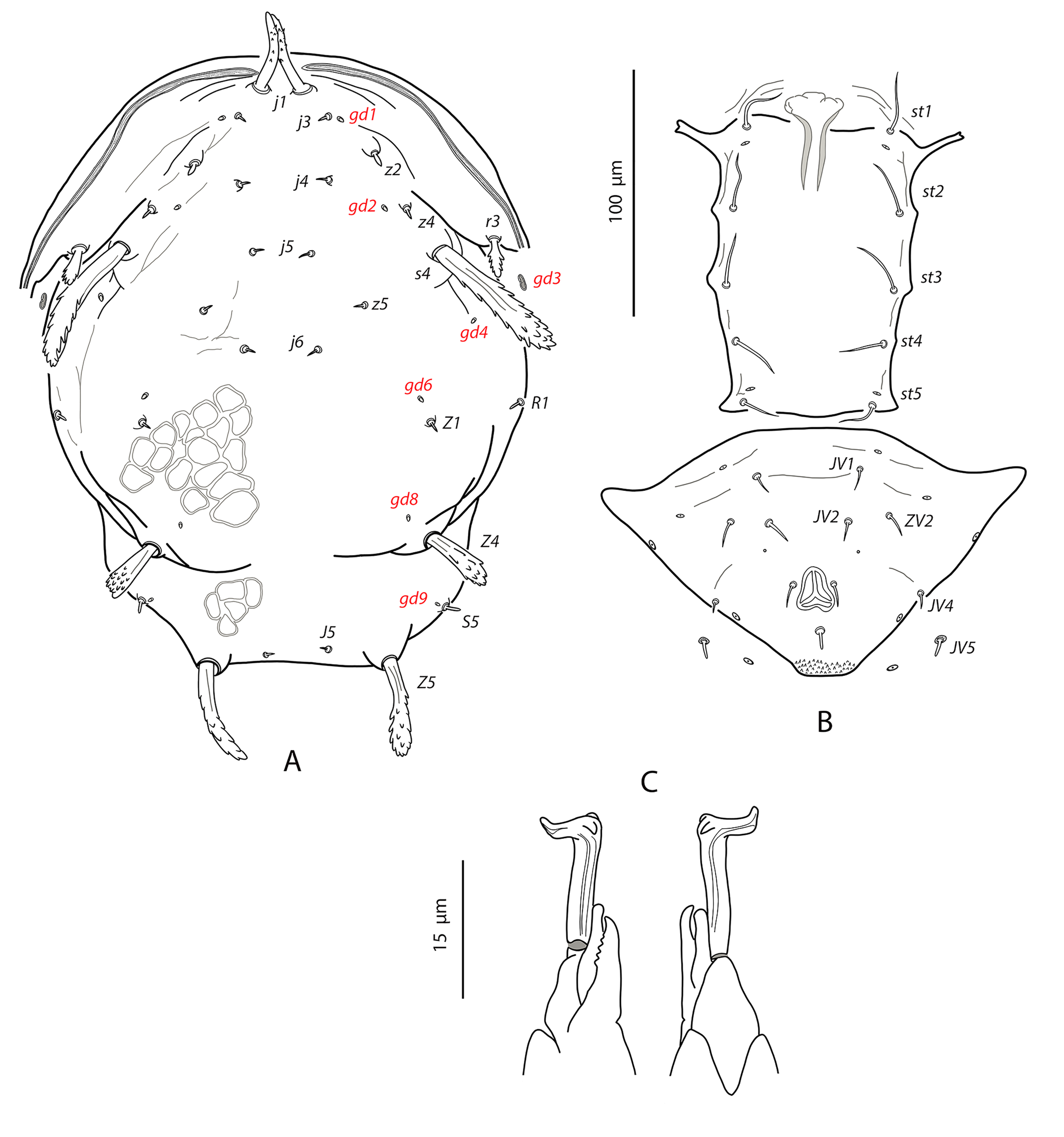
Neoparaphytoseius species with female dorsal shield 363–386 μm long, having lateral irregular margins between setae s4 and Z4; dorsal opisthosomal surface elevated, almost forming a transverse crest at the level of setae Z4; ornamentation striate-reticulate with two large areas of areolate cells between setae j6–Z4 and anterior to insertions of Z5–J5. Dorsal setae of different aspect and size, setae j1 lanceolate and serrate, setae s4, S2, Z4 and Z5 thick, strongly serrated and longer; other short or very short; all setae placed on tubercles and covered by a dense layer of crystals. Sublateral setae r3 and R1 in variable position, sometimes on the soft integument other on the dorsal shield. Peritremes reaching the sockets of setae j1. Ventrianal shield pentagonal and smooth; setae JV1 and ZV2 in marginal position. Setae JV5 short, smooth, and blunt. Calyx hemispherical or shallow bowl, slightly asymmetrical; atrium incised longitudinally. Tectum subtriangular; number of cheliceral teeth variable; fixed digit with one subapical tooth and eight to ten additional teeth, movable digit with two or three teeth. Genua II and III with eight setae: 2 2/1, 2/0 1; macroseta on basitarsus IV knobbed and with crystals, 63–74 long. Male ventrianal shield with six pairs of preanal setae. Spermatodactyl L-shaped.
Description
Female
(n=12) (Figures 1–3)
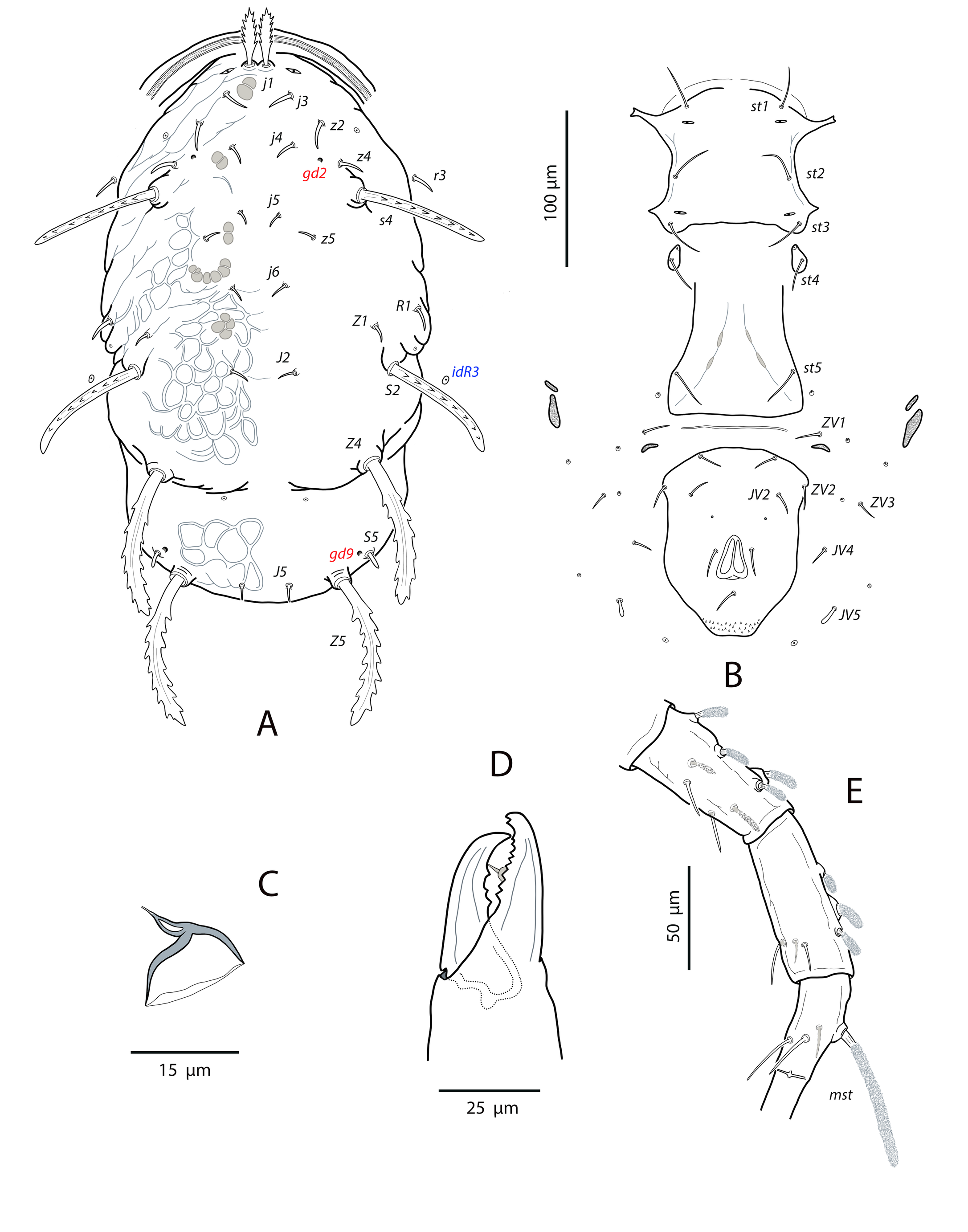
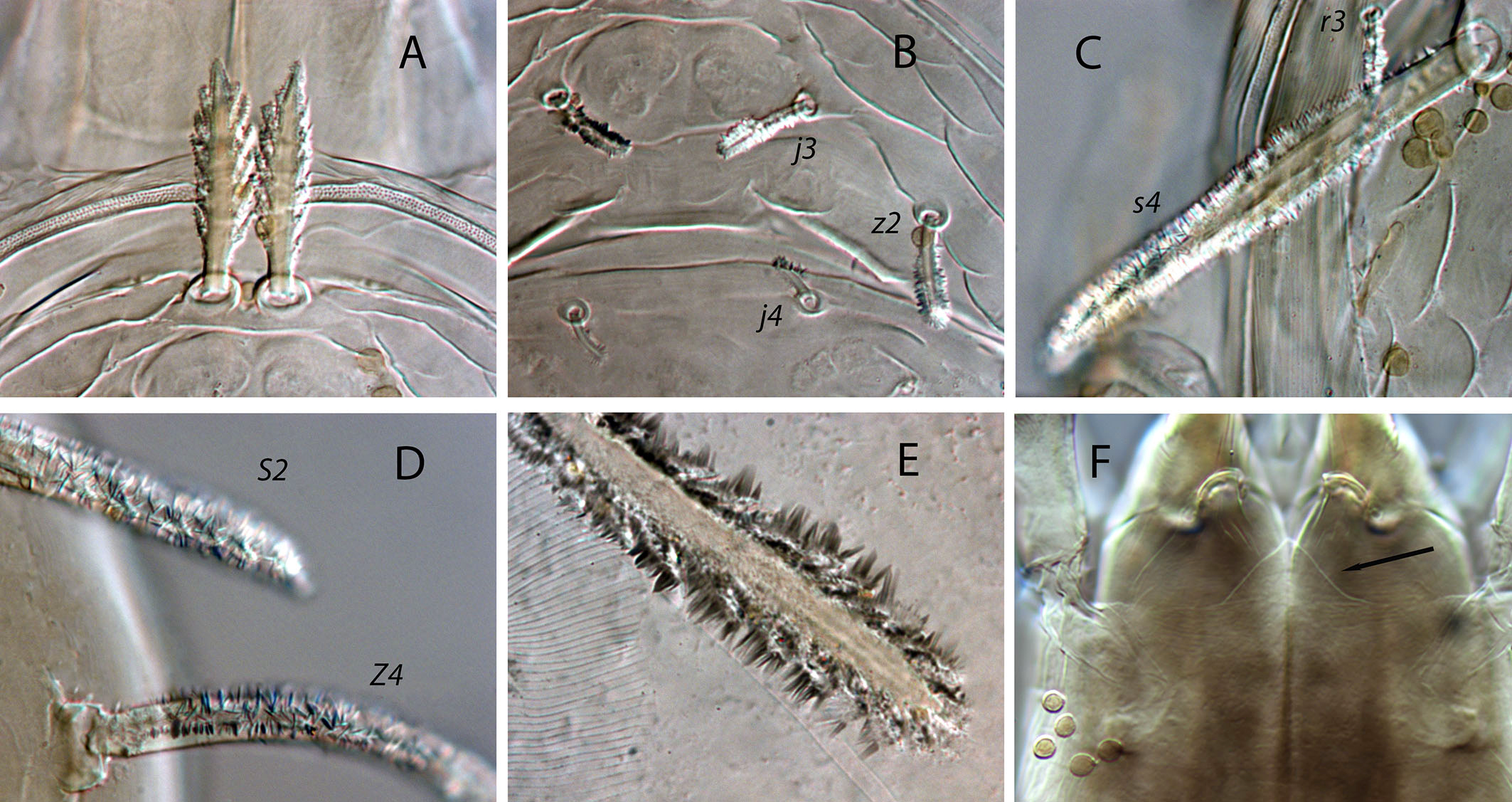
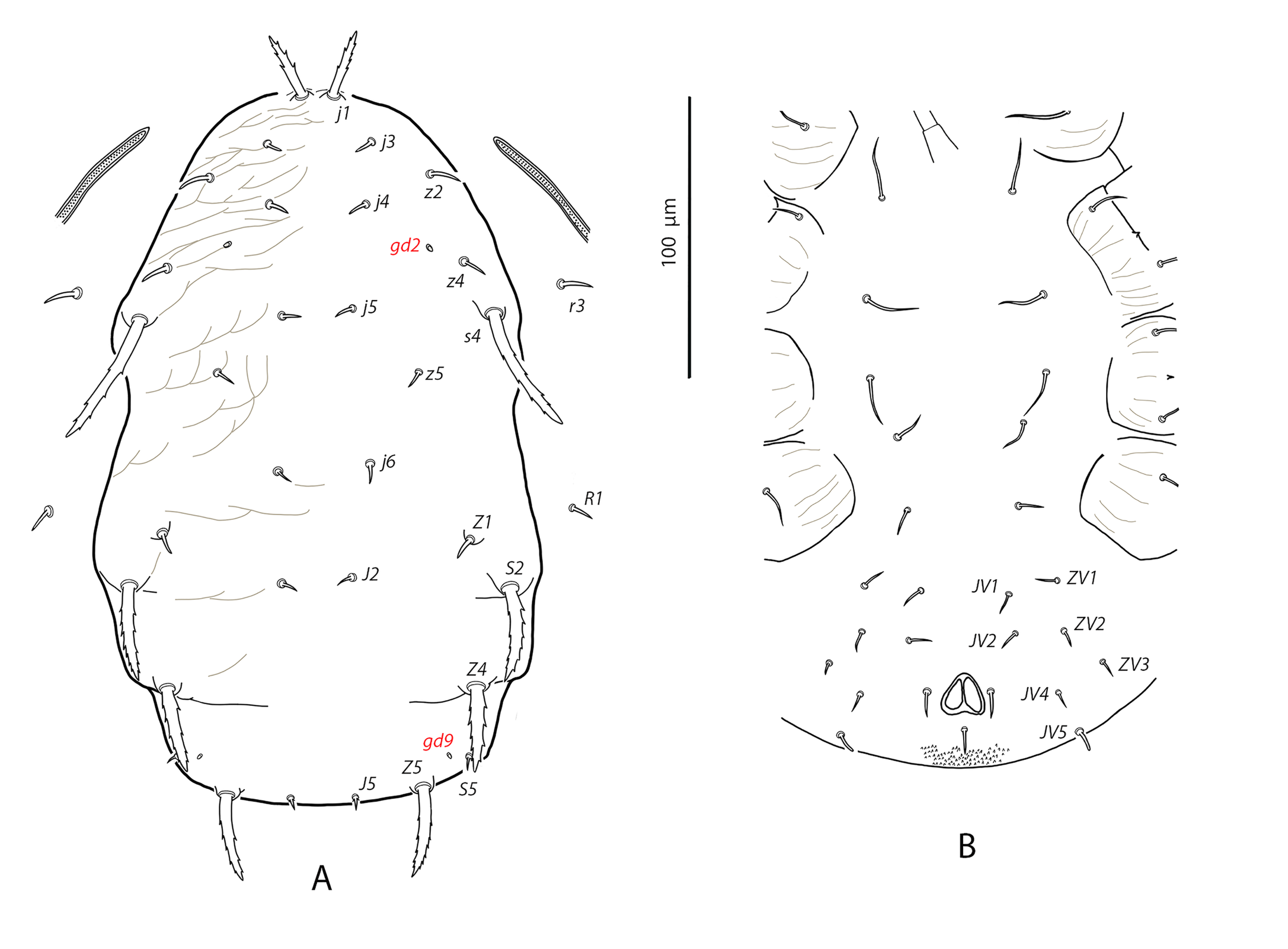
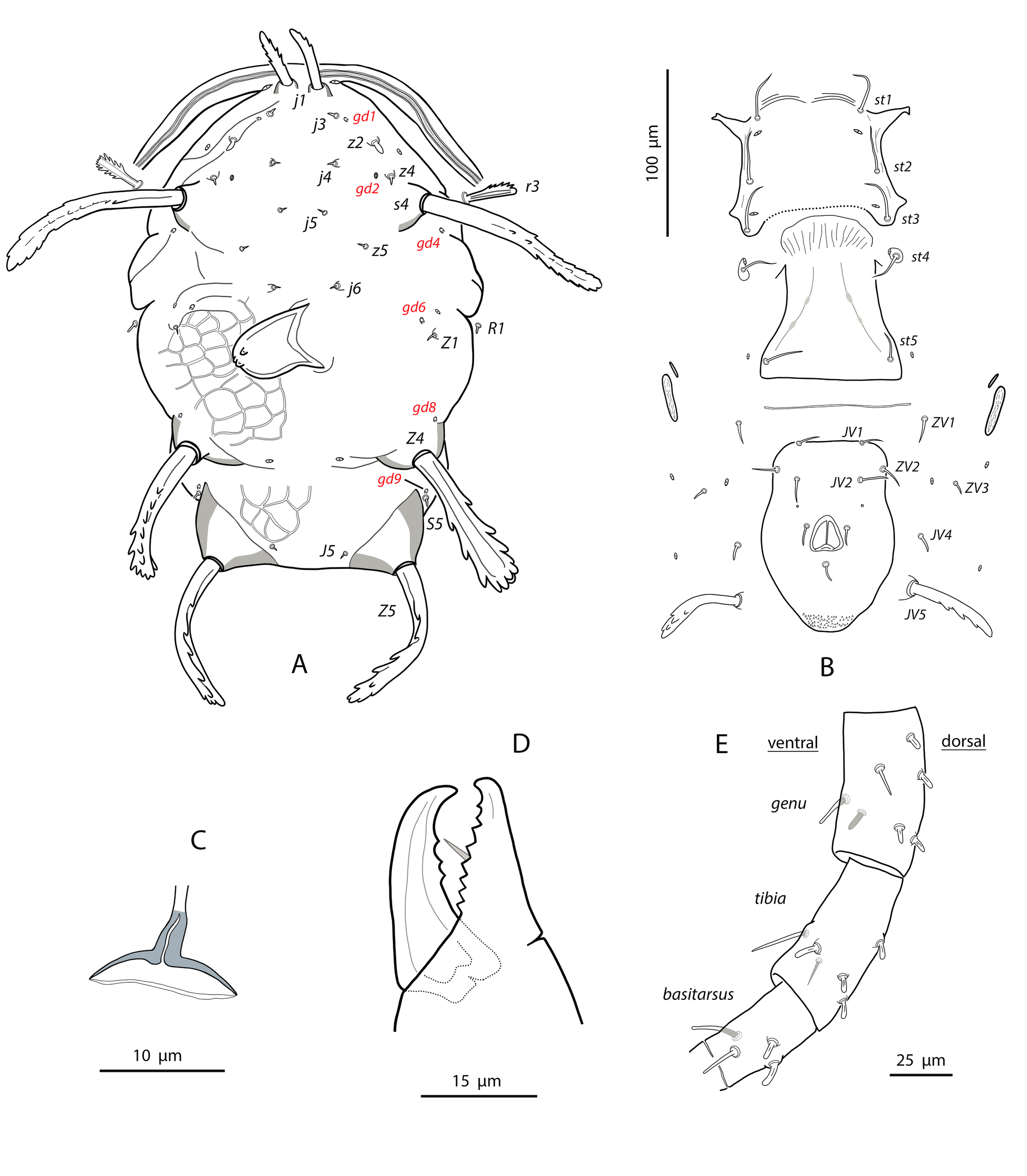
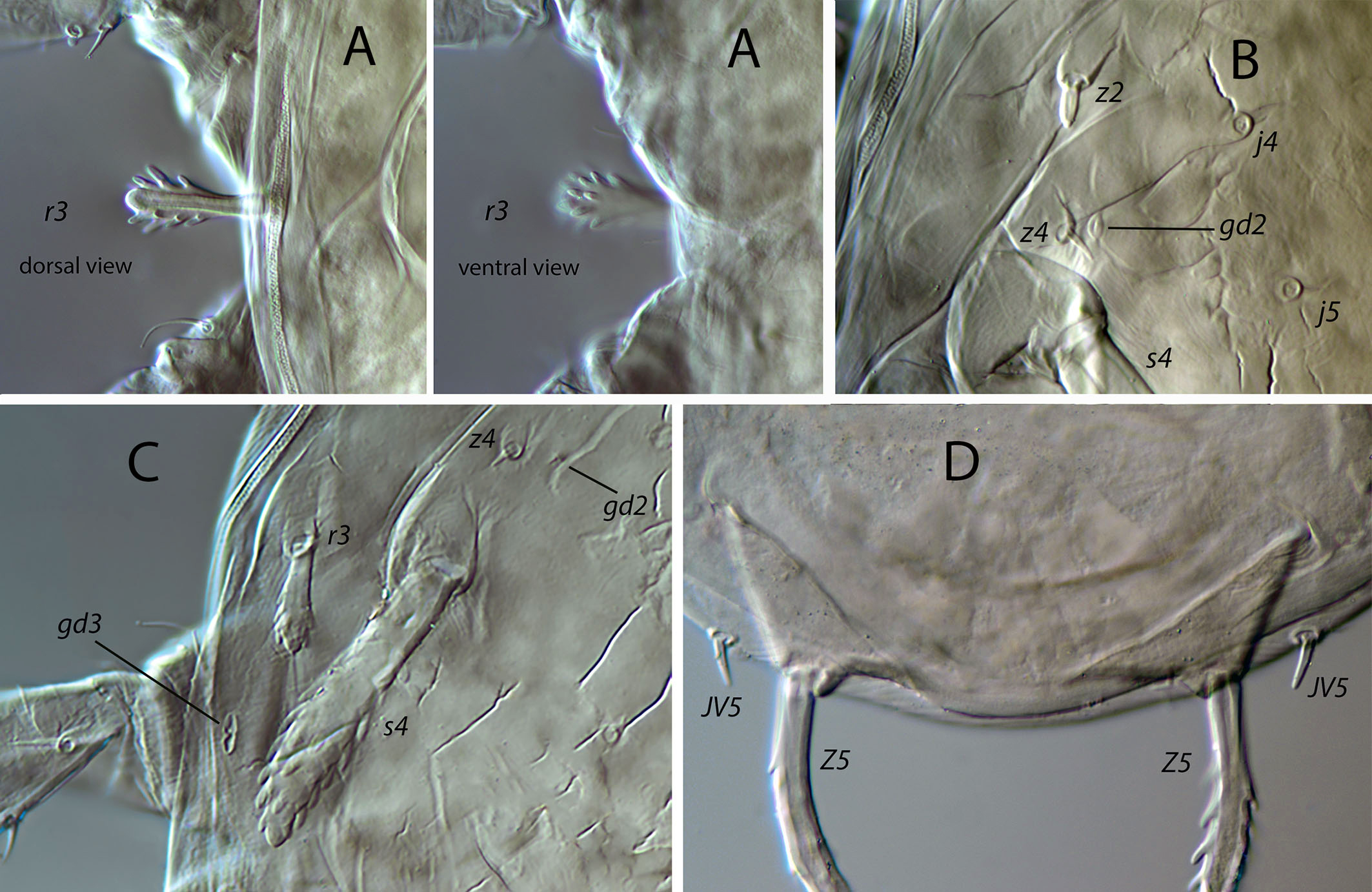
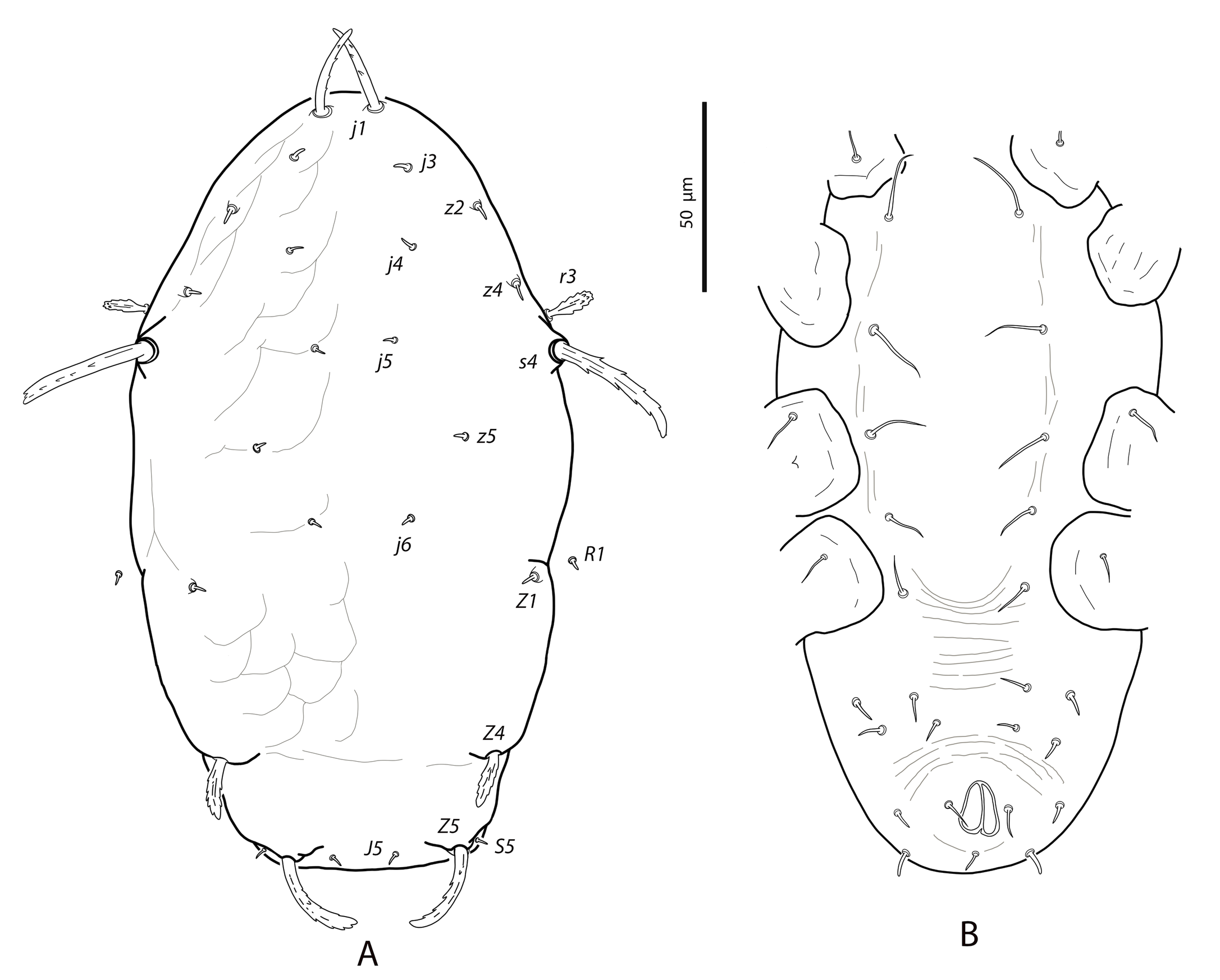
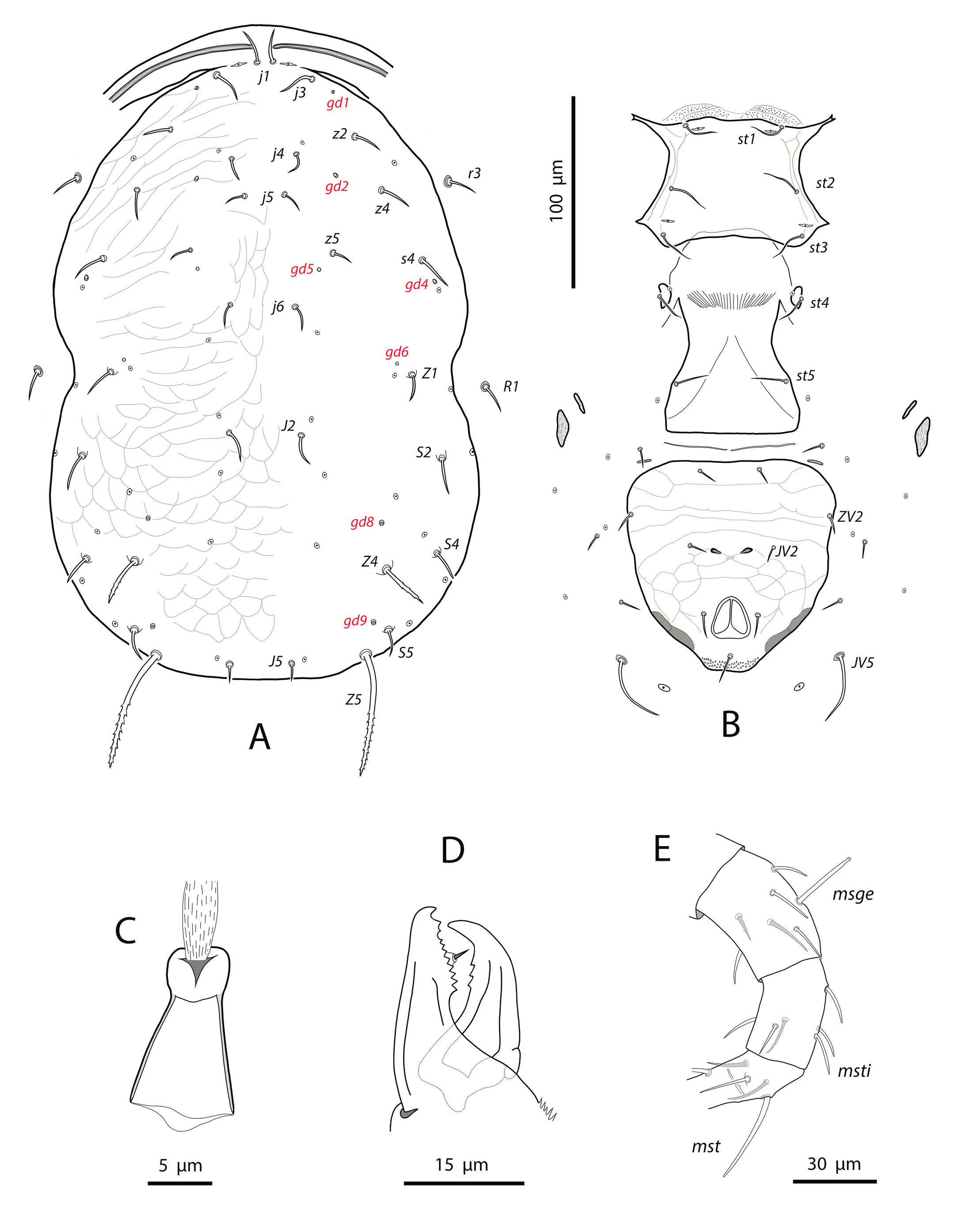
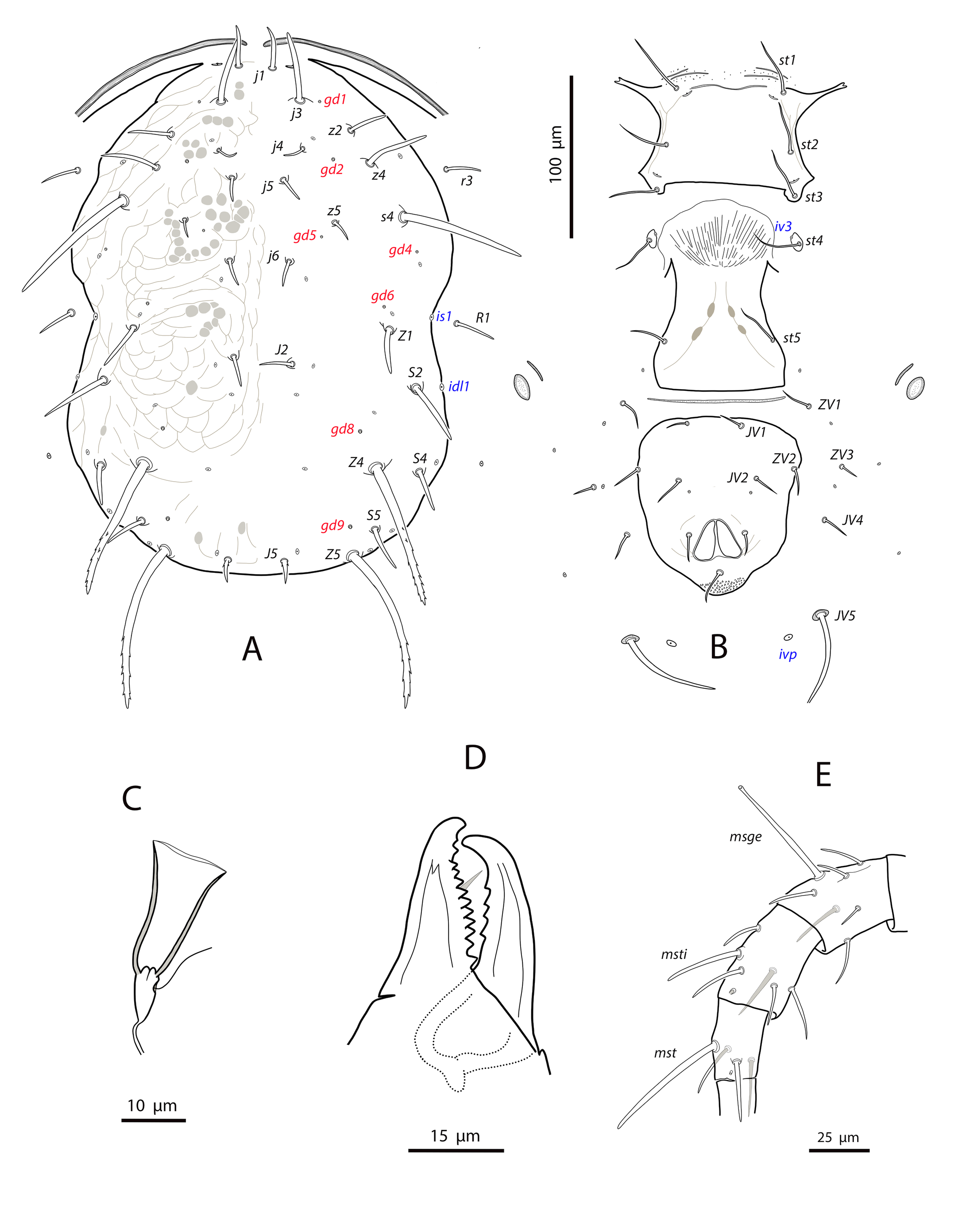
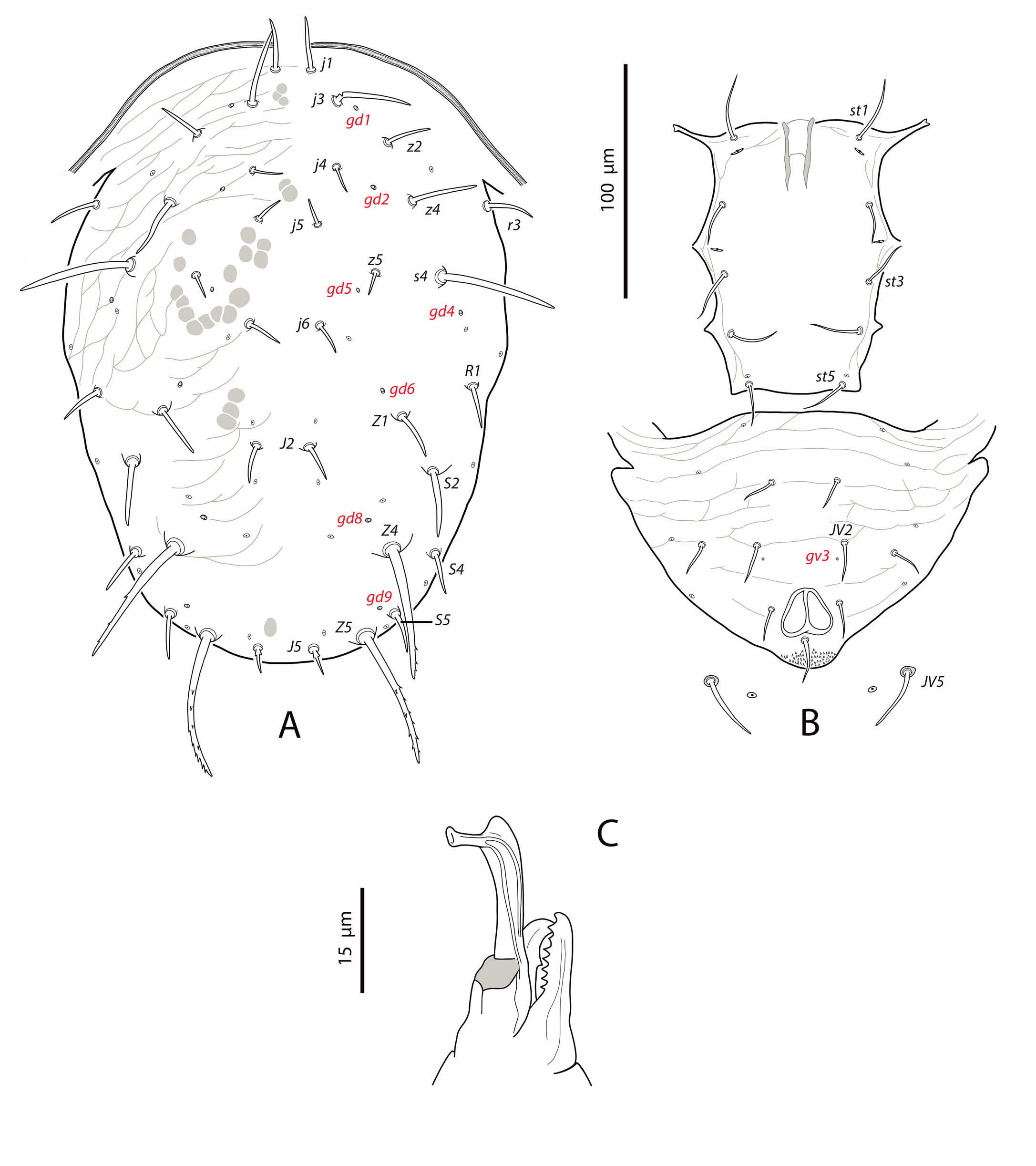
Dorsal and lateral idiosoma — (Fig. 1A). Dorsal shield suboval, wider at the level of setae Z1 or between setae R1–S2; 370, 374 (363–386) long; 204, 200 (180–208) wide at level of setae s4; 237, 239 (230–250) wide at level of setae Z1. Lateral margins irregular, with a conspicuous and rounded indentation between Z1–S2, at the level of Z4, and sometimes at the level of s4, but without representing true incisions or notches. Shield well sclerotized, irregularly striated and reticulated; anterolateral striae between setae j1 and S2, dorsomedial region of the podonotum almost smooth; opisthonotum with two large areas of areolate cells, between setae j6–Z4 and anterior to insertions of Z5–J5; almost smooth between the areolate areas. Opisthonotum with eight pairs of setae and absence of setae S4. Dorsal setae of different length and appearance; setae j1 lanceolate and serrate, setae s4, S2, Z4 and Z5 thick, strongly serrated and longer; the remaining short o very short, smooth and blunt, except J5, basally barbed. All setae placed on tubercles, which are strong and protrude from the shield in the longest setae. These longest setae, and sometimes the shorter, are almost completely covered by small crystals, difficult to remove from the setae (Figs. 2A, 2E). Setae j1 38, 41 (36–49), j3 15, 16 (15–17), j4 8, 9 (8–10), j5 7, 7 (7–8), j6 8, 8 (7–8), J2 8, 9 (8–11), J5 9, 8 (7–9), z2 15, 16 (15–18), z4 13, 13 (10–17), z5 8, 7 (6–8), Z1 11, 9 (8–11), Z4 107, 103 (96–108), Z5 109, 106 (99–117), s4 85, 86 (82–90), S2 86, 87 (81–91), S5 11, 10 (8–11). Sublateral setae r3 and R1 placed indistinctly on the soft integument or on the dorsal shield; 42% of females with both pairs on the shield, 58% with one member of those setae off the shield; on the holotype both r3 are off the shield and both R1 on the shield, on the other females only one seta r3 or R1 is off the shield while the remaining three setae are placed on the shield. Length of r3 10, 12 (10–13); of R1 9, 10 (8–11).
Dorsal adenotaxy and poroidotaxy difficult to discern due to the ornamentation and the presence of crystals around the setae; at least the solenostomes gd2 and gd9 can be identified; gd9 close and anteroparaxial to setae S5. Poroid idR3 present. Peritremes terminating at the level of setae j1; 231, 229 (219–235) long; peritrematal grove narrow, with two or three lines of microvilli; solenostome gd3 present in marginal position on the peritrematal shield.
Ventral idiosoma — (Fig. 1B). Base of tritosternum 13, 13 (12–14) wide; plumose laciniae reaching the sixth row of deutosternal denticles, 36, 35 (33–38) long; total length of tritosternum 108, 106 (102–109). Sternal shield 86, 91 (86–96) long, 87, 89 (86–93) wide; distance st1–st3 75, 78 (75–81), distance st2–st2 79, 79 (78–81). Anterior margin poorly sclerotized; posterior margin more or less straight. Setae st4 on piriform metasternal plates. Epigynal shield 130, 130 (126–136) long; distance st5–st5 81, 79 (76–82). Narrow sclerotization between epigynal and ventrianal shields; elongated and curved microsclerites between setae ZV1 and the anterolateral angles of the ventrianal shield. Ventrianal shield pentagonal and smooth, 117, 120 (115–126) long, 101, 107 (101–113) wide at level of ZV2, 81, 85 (81–89) wide at level of anus; anterior margin convex, laterals concave posterior to setae ZV2. Three pairs of preanal setae. Preanal pores minute and punctiform, posteroparaxial to setae JV2; 44, 45 (41–50) distant each other. Four pairs of setae (ZV1, ZV3, JV4, JV5) and five pairs of poroids surrounding the ventrianal. Setae JV5 short, smooth, and blunt, 12, 12 (11–13); as short as ZV3 and JV4 and shorter than the preanal setae. Anterior metapodal plate bacillar 19, 18 (17–20); posterior metapodal elongate 34, 32 (31–34) long.
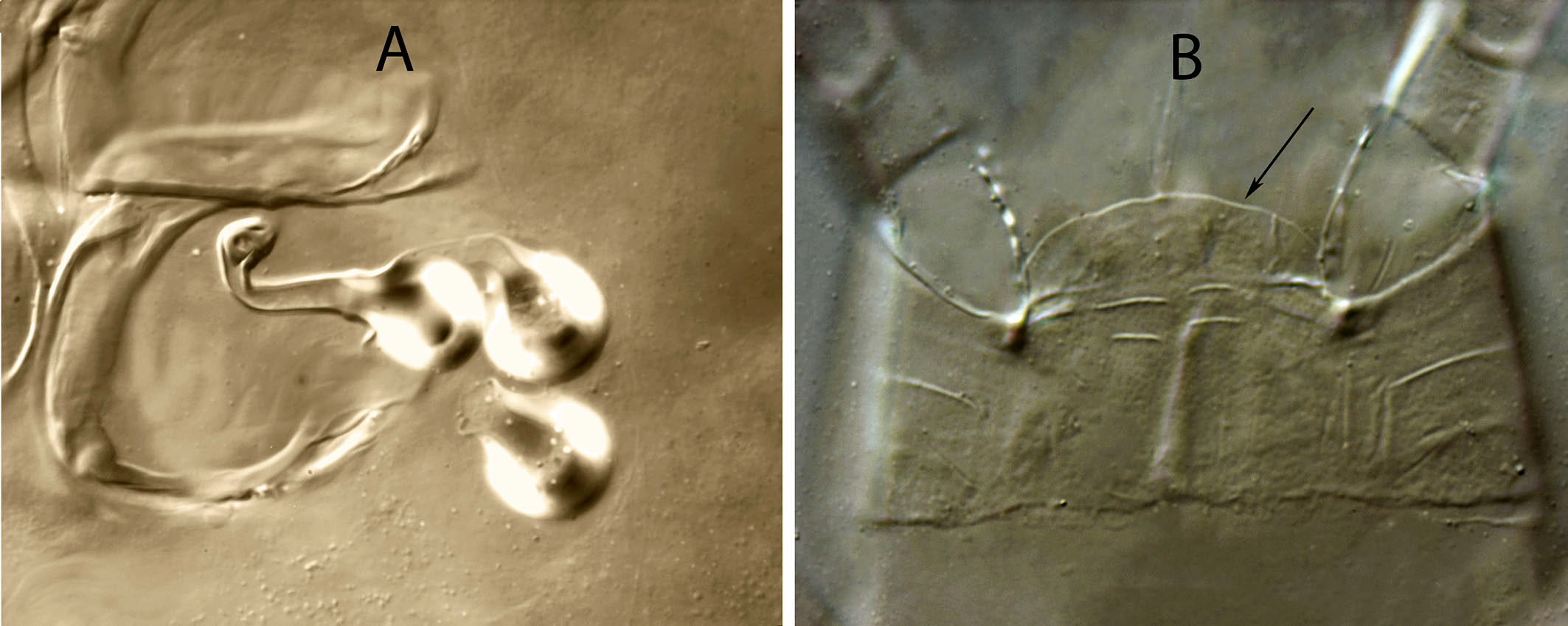
Spermathecal apparatus — (Fig. 1C). Major duct short, thin and tubular, gradually widening near the atrium; atrium incised longitudinally; calyx hemispherical or shallow bowl, asymmetrical, with one arm longer than the other, 11, 10 (8–11) deep, 15, 14 (13–15) wide at the junction with the vesicle.
Gnathosoma and chelicerae — (Fig. 1D). Gnathotectum subtriangular, forming an almost 90-degree angle (86 degrees in the holotype) (Fig. 2F). Deutosternal groove with seven rows of two denticles placed on lateral margins of each row; groove base laterally dentate. Corniculi straight and subparallel, length 36, 36 (34–38). Setae h1 24, 28 (24–31), h2 23, 21 (19–23), h3 23, 23 (22–23), pc 30, 28 (26¬–30). First cheliceral segment 38, 48 (38–54) long; second segment 109, 105 (96–110) long. Fixed digit 40, 40 (38–42), with one subapical tooth and eight to ten additional teeth; differences in the number of teeth were observed even in the same specimen. Movable digit 39, 40 (38–41) long with two or three teeth, again differences in this patter can be found in the same female. Palps 131, 136 (130–141) long.
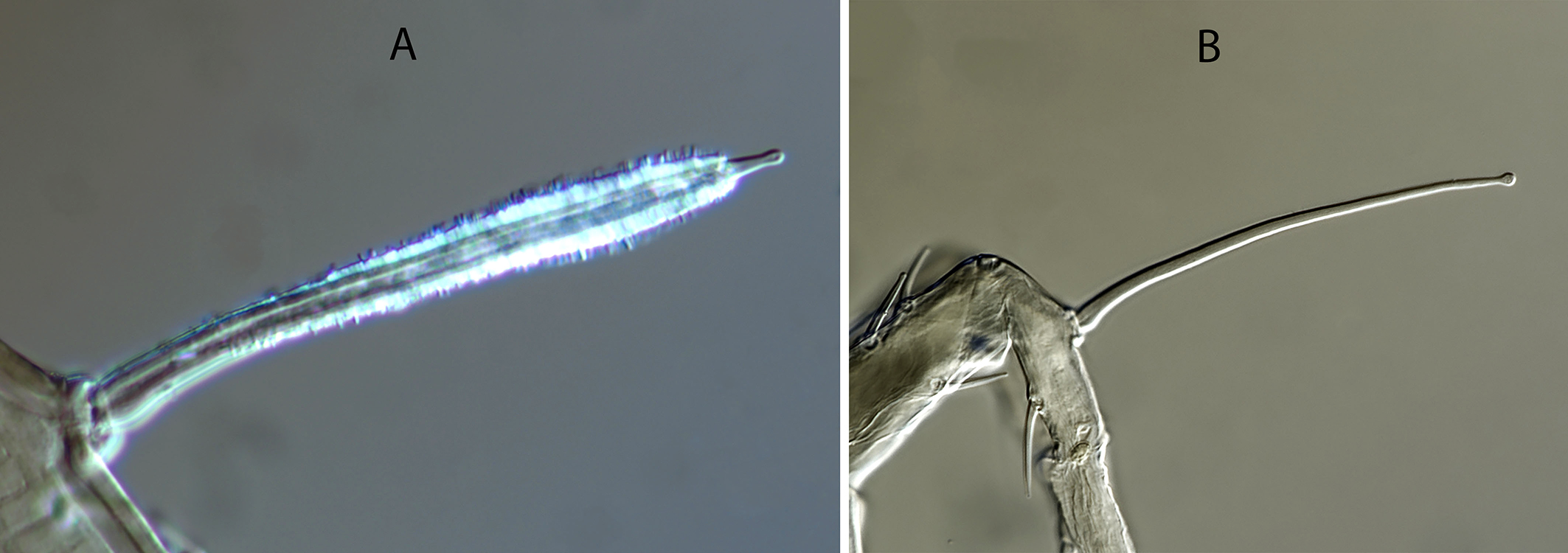
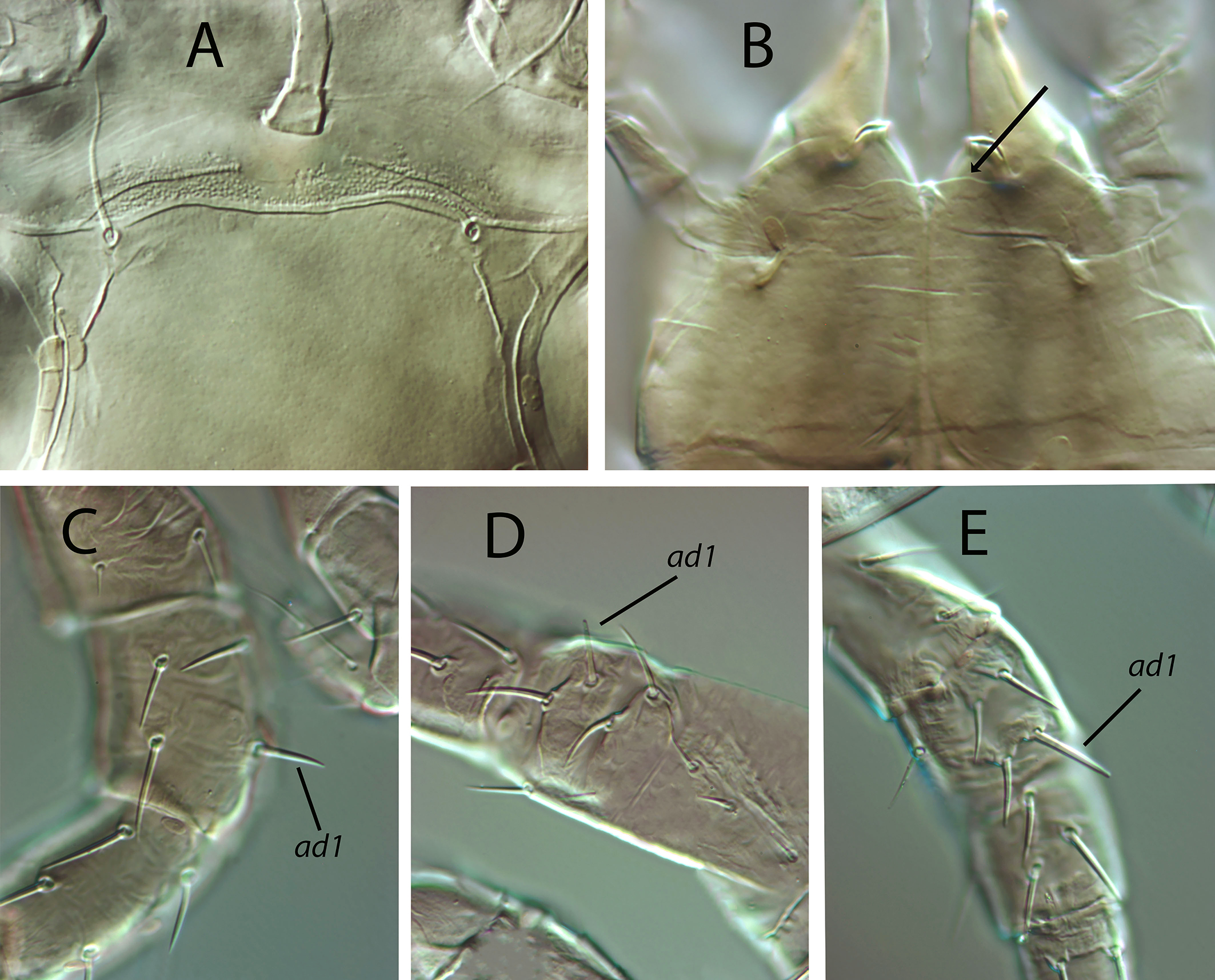
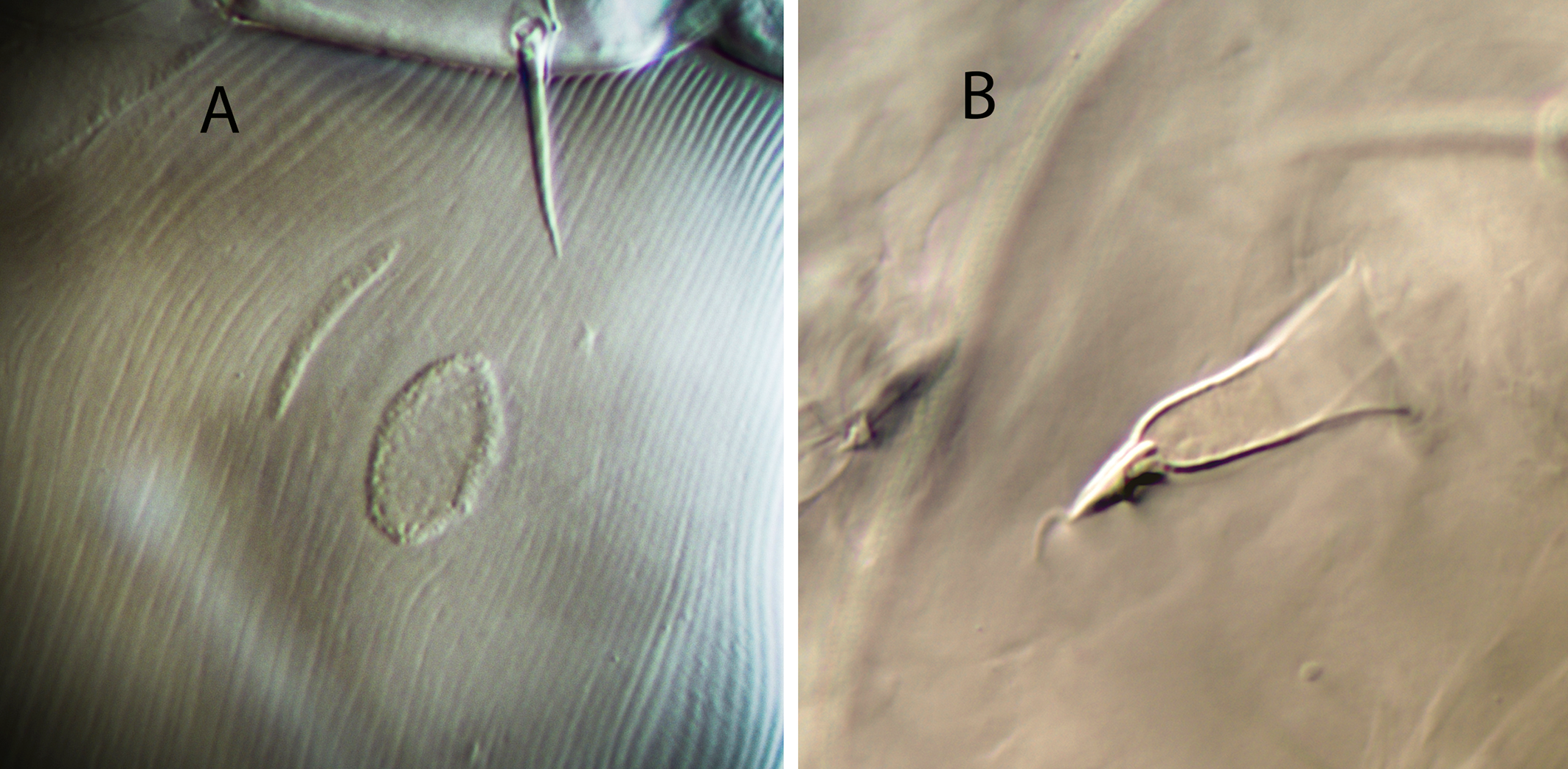
Legs — (Figure 1E). Leg I 368, 378 (368–389); leg II 306, 312 (295–322); leg III 332, 333 (303–352); leg IV 546, 547 (521–571). Setae on legs variable in morphology; in general, dorsal setae on all the segments are shorter, thicker and with crystals, while lateral and ventral setae are longer, setaceous and without crystals. Most of dorsal setae arising from tubercles. Setal formula for legs I–IV (coxae to basitarsi) 2-5-12-10-10-0; 2-5-10-8-7-4; 2-5-6-8-7-4; 1-5-6-8-6-4. Macroseta on basitarsus IV knobbed and always with crystals, at least on the distal two-thirds of its length (Fig. 3A); 70, 69 (63–74) long. Genua II and III with eight setae; setal formula of both segments 2 2/1, 2/0 1. A pore-like structure in the distal part of femur IV, between seta ad1 and the distal margin of the segment.
Male
(n=4) (Figure 4)
Dorsal idiosoma — (Fig. 4A). Dorsal shield 291 (282–301) long, 182 (176–184) wide. Ornamentation of the shield similar to that of female. Adenotactic and poroidotactic pattern not discernible; gland opening gd3 between the peritrematal and dorsal shields. Peritrematal shield fused to dorsal shield at the level of setae r3; peritremes extending forward to the insertion of setae j1; microvilli aligned in two rows. Nineteen pairs of dorsal setae; setal morphology similar to that of female, all dorsal setae smooth except j1, s4, S2, Z4 and Z5, strongly serrated and J5 basally barbed. Dorsal setae arising from tubercles, more developed in the longer setae. Setae j1 32 (30–34), j3 15 (13–16), j4 8 (8–9), j5 7 (7–8), j6, J2 8 (7–8), J5 5 (5–6), z2 16 (14–17), z4 10 (9–11), z5 6 (6–7), Z1 8 (7–9), Z4 48 (44–51), Z5 56 (54–60), s4 58 (56–60), S2 47 (46–50), S5 7 (6–8), r3 12 (12–13), R1 10 (8–11).
Ventral idiosoma — (Fig. 4B). Sternogenital shield smooth except for longitudinal striae on the lateral margins; 152 (147–152) long, 70 (68–71) wide at the level of setae st2, 52 (48–54) wide at level of setae st5. Five pairs of setae and two pairs of poroids on the shield. Posterior margin straight. Ventrianal shield subtriangular, anterior margin convex; 121 (116–124) long, 188 (182–201) wide at anterior corners. Shield with transversal striae on the anterior part. Six pairs of preanal setae and four pairs of poroids; preanal setae of different length, ZV1 twice as long as ZV2 and JV4; ZV1 18 (16–19), JV1 14 (13–14), ZV2 9 (9–10), JV2 11 (10–13), ZV3 7 (7–8), JV4 9 (8–10); setae JV4 in marginal position and aligned with perianal setae. Preanal solenostomes minute and punctiform, posteroparaxial to setae JV2, 40 (39–41) separated from each other. Setae JV5 smooth and blunt 10 (9–10) long; poroids ivp between setae JV5.
Chelicerae and spermatodactyl — (Fig. 4C). Fixed digit 22 (22–23) long, with five–six teeth; movable digit 23 (21–24) long, with one tooth. Spermatodactyl L-shaped; shaft straight, 18 (17–18); heel prominent; foot long 13 (12–13), with lateral and acute projection; toe turned up.
Legs — Leg chaetotaxy and setal formula of genua II and III as in female. One smooth and blunt macroseta on basitarsus IV, 50 (46–53) long, covered by crystals in all the specimens examined.
\subparagraph{Deutonymph}
(n=2) Figure 5
Dorsal and lateral idiosoma — (Fig. 5A). Dorsal shield elongate, 251 (250–251) long, 115 (113–116) wide. Shield surface striated in the anterolateral region, weakly ornamented on the opisthonotum and smooth in the dorsomedian area. Gland openings difficult to discern, only gd2 and gd9 appear to be present. Sixteen pairs of dorsal setae, all smooth and short, except j1, s4, S2, Z4 and Z5, which are longer, very thick, serrated and tuberculated, and J5 very short and basally barbed. Serration of the longer setae less developed than in female, with less barbs or digitations and smaller in size. Lengths of setae: j1 23 (22–24), j3 8, j4 8 (7–8), j5, j6 7 (6–7), J2 6, J5 4, z2 11 (10–11), z4 9 (8–9), z5 6 (5–6), Z1 7, Z4 33 (30–35), Z5 36 (35–36), s4 46 (44–47), S2 35 (33–36), S5 5. Sublaterals r3 and R1 on the interscutal soft cuticle; r3 10 (9–10), R1 7 (6–7). Crystals absent on the dorsal setae. Peritremes extending beyond setae z2.
Ventral idiosoma — (Fig. 5B). Tritosternum 80 (79–81) long, stalk-like base 13 (12–13) long by 11 wide. Coxisternal region with five pairs of setae, poroids not visible in the examined specimens. Seven pairs of setae around to the anal opening (JV1, JV2, JV4, JV5, ZV1, ZV2, ZV3) in addition to the perianal and postanal setae. Preanal pores could not be observed. Primary metapodal shield 14 (13–14) long. Setae JV5 short, blunt and smooth, 7 (6–7) long.
Gnathosoma and chelicera — Anterior margin of tectum domed-shaped. Deutosternal groove with eight rows of denticles. Fixed digit of chelicerae 23 (22–23) long, with eight teeth; movable digit 24 long, with three teeth.
Legs — With the same chaetotaxy as in females. None of the leg setae are covered by crystals. Macroseta on basitarsus IV very long, 78 (75–80), smooth and knobbed (Fig. 3B).
Taxonomic relationships
The habitus of N. moraesi sp. nov. presents remarkable differences with the other species in the genus. On the dorsal shield, the longer and stouter setae in the remaining species are j1, j3, s4, Z4 and Z5, while in the new species j3 is short and setaceous and, on the contrary, setae S2 are very long, stout and serrate. The dorsal plane also appears to be different. In the new species the opisthosoma is clearly elevated at the level of setae Z4, forming an almost complete transversal crest. Posterior to the crest the shield descends abruptly to recover the horizontal plane. This particular morphology can be easily observed on females in profile position before the montage. This character has not been described for the other species.
On the ventral plane, the shape of the ventrianal shield and the relative position of some preanal setae are original in the genus. The ventrianal shield is almost pentagonal with a ratio length/width of 1.2. In the other species this shield is more elongate, almost vase-shaped, with a ratio L/W of 1.7 in N. caatinga (Silva et al. 2021), 1.4 in N. sooretamus (El-Banhawy 1984; Jiménez et al. 2014), and 1.3–1.5 in N. charapa (Jiménez et al. 2014; Souza et al. 2021). Setae JV2 in the previously described species are inserted well-posterior to the insertions of setae ZV2; in the new species, however, setae JV2 are placed anteriorly on the shield, merely posterior to the sockets of ZV2, giving the impression of being almost transversally aligned on some specimens. Finally, JV5 is consistently long, thick and serrate in the other species, while it is short, smooth and knobbed in N. moraesi sp. nov. Morphological differences among all the species are summarized in Table 1.

At a glance, the most striking character is the presence in the adults of birefringent crystals grow on the dorsal setae of idiosoma and legs. These formations cover between 50–90% of setal surface and consist of cylindrical or faceted shapes crowded organized (see Figs. 2 and 3). The function or biological significance of these structures is unknown but the crystals are probably the result of setal secretions, as an internal structure assimilable to a tube can be seen in the naked part of the setae.
Etymology
The new species is dedicated to Prof. Dr. Gilberto José de Moraes for his contributions to the knowledge of phytoseiids and other plant and soil mites and for being the teacher and mentor of an entire generation of Brazilian acarologists and researchers from all over South and Central America, Africa and Europe.
Genus Proprioseius Chant, 1957
Proprioseius gibbus Moraes & Denmark, 1999
Proprioseius gibbus Moraes and Denmark, 1999: 99
Specimens examined
Two females and one male on Acritopappus sp. (Asteraceae); Morro do Pai Inácio, Palmeiras, Bahia, Chapada Diamantina; 12°27′26″S, 41°28′26″W, 1105 m asl; 14 January 2010. Seventeen females, four males, one deutonymph on Acritopappus sp. (Asteraceae); track Lençóis-Morro do Pai Inácio, Lençóis, Bahia, Chapada Diamantina, 12°29′49″S, 41°26′34″W, 871 m asl; 15 January 2010.
Collected specimens deposited in the Mite collection of the ESALQ-USP (Escola Superior de Agricultura ''Luiz de Queiroz″, Universidade de São Paulo, Piracicaba, State of São Paulo, Brazil), Laboratory of Acarology, Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Spain, and the Mite collection of the Centre de Biologie pour la Gestion des Populations (UMR CBGP), Montferrier sur Lez, France.
Geographical distribution
Endemic Brazilian species. It has only been found in the state of Bahia.
Complementary description
Female
(n=12) (Figures 6–8)
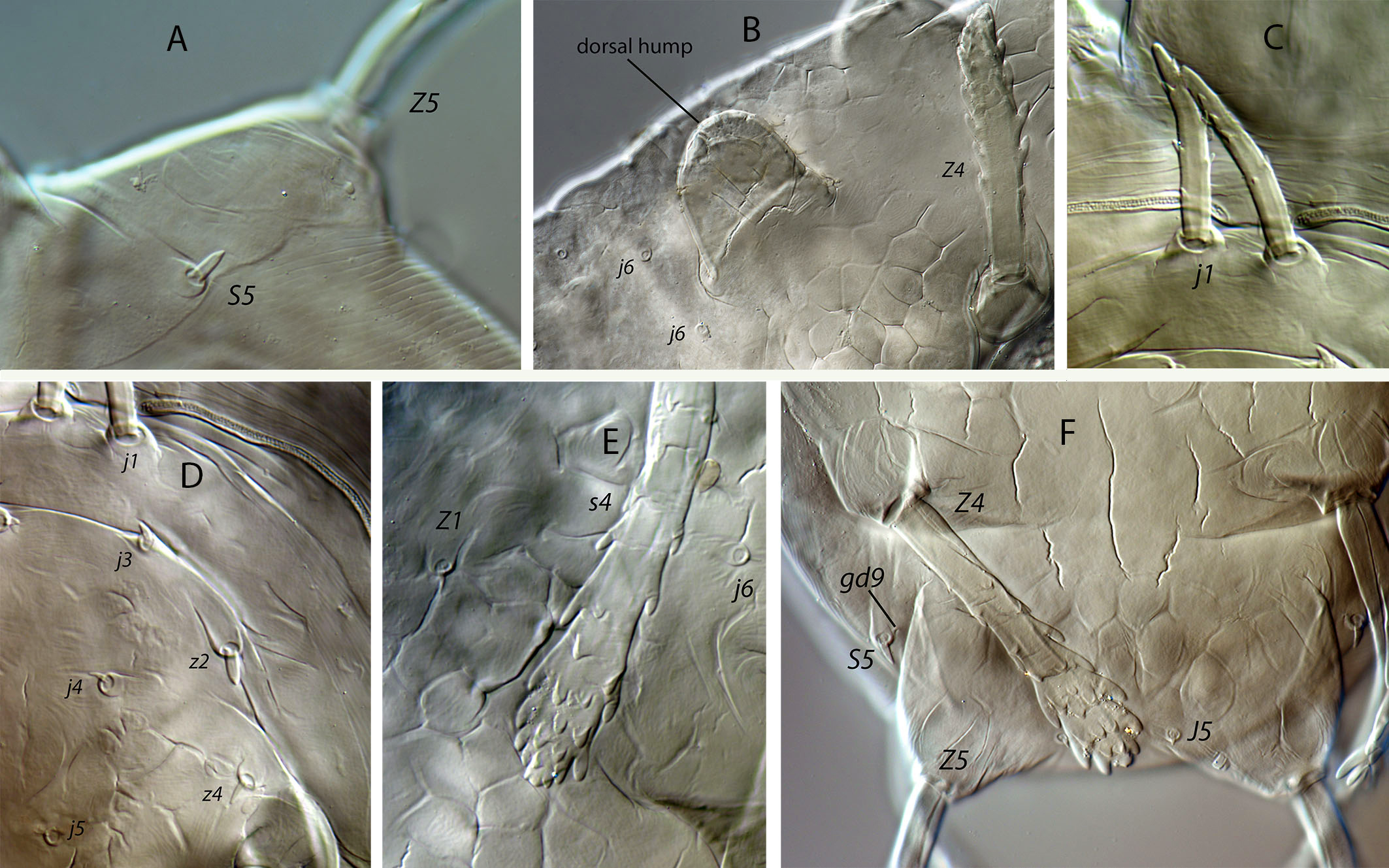
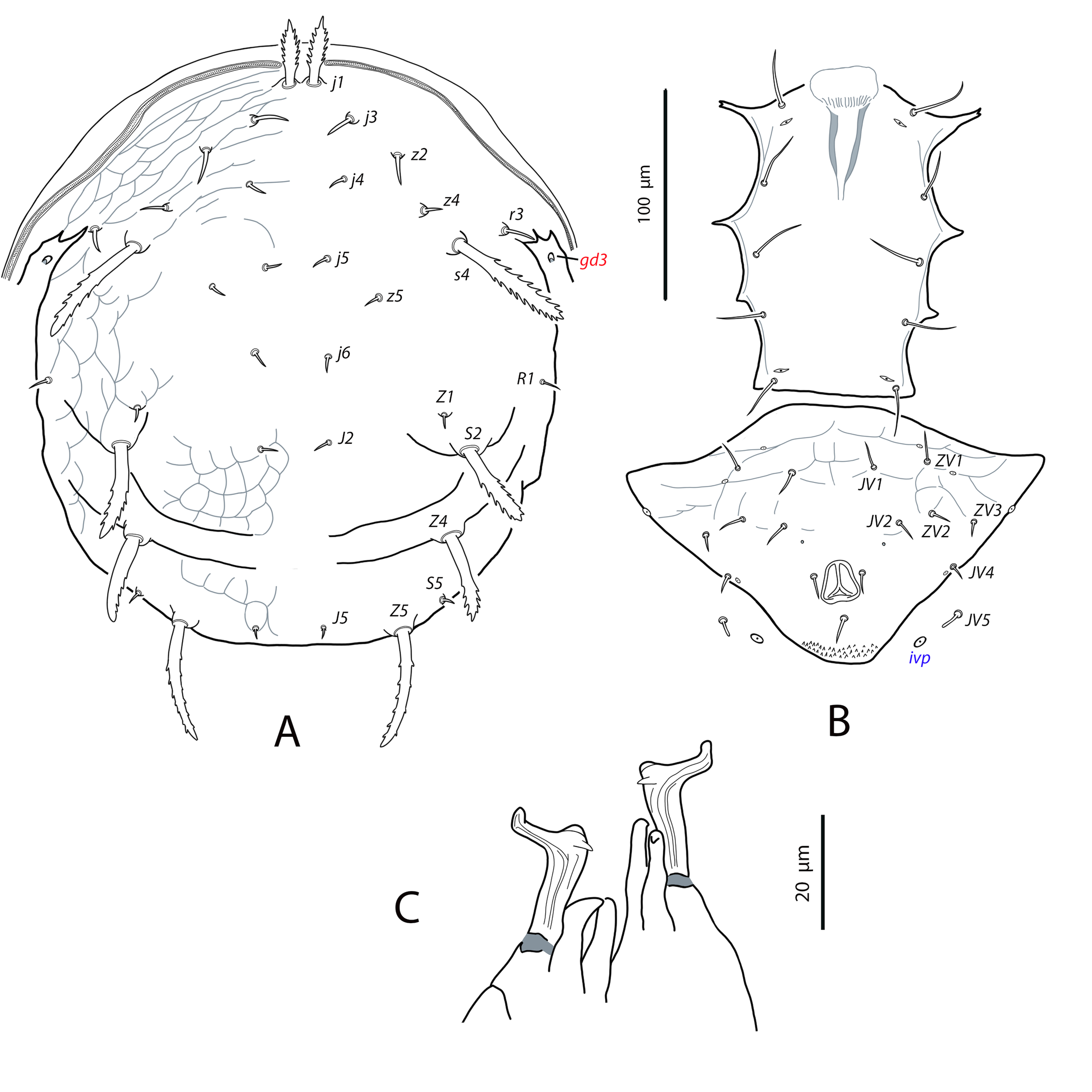
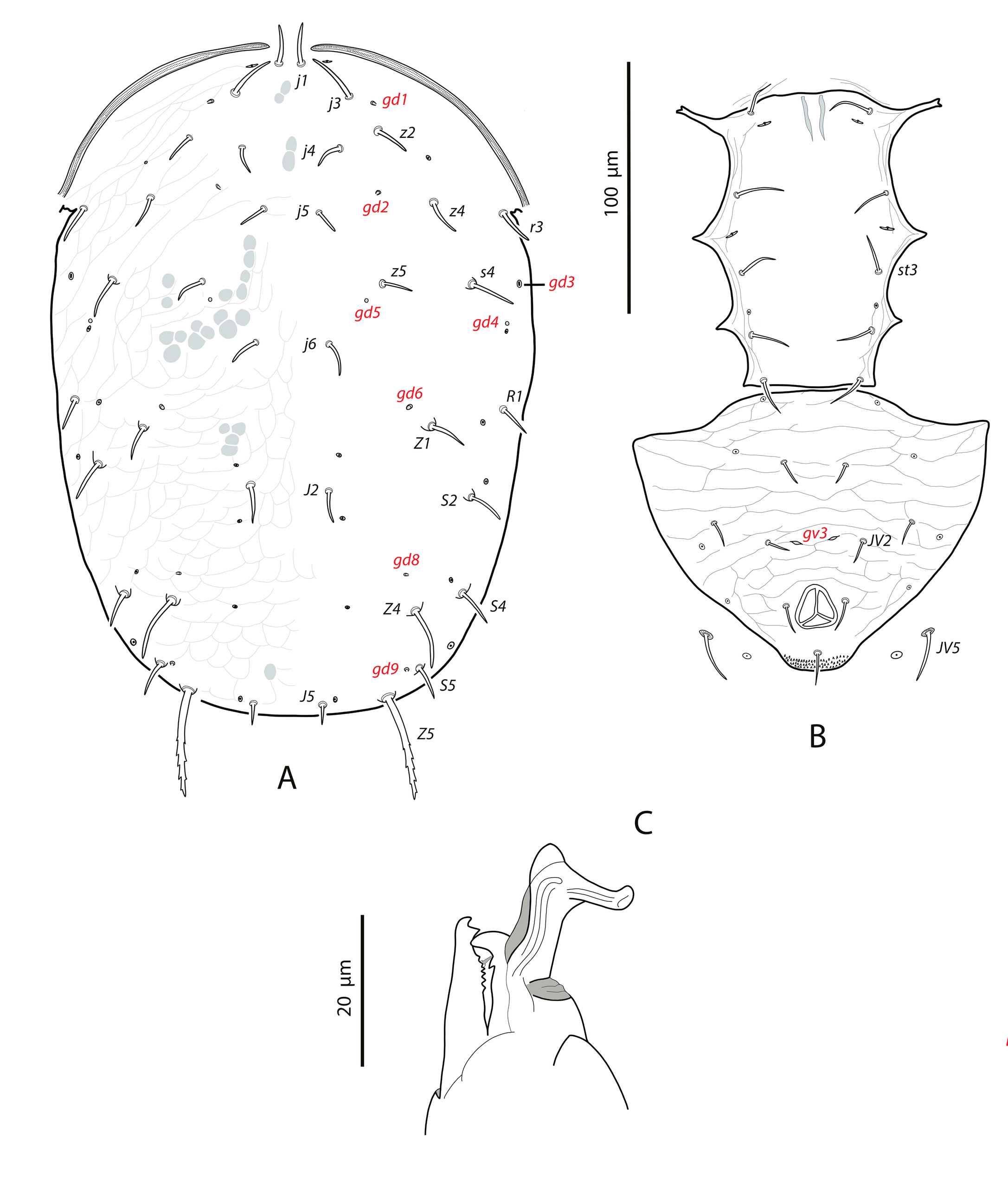
Dorsal and lateral idiosoma — (Fig. 6A). Dorsal shield fusiform, wider at the level of setae j6; 305 (294–317) long, 152 (144–174) wide at level of setae s4, 178 (172–194) wide at level of setae Z1. Lateral margins of the shield sinuous and irregular, sometimes with a small indentation behind setae z2; there is no indication of strong indentation or notch at the level of s4, but the lateral margin is constricted anterior to setae R1. On the posterior part the shield is elevated because of the development of the hypertrophied tubercles from which the setae Z5 arise (Fig. 7A). Dorsal surface irregular, rugose, striated anterolaterally, almost smooth on the mid podonotum, with distinct patches of large pentagonal or reniform cells between setae Z1–Z4 and between the tuberculate bases of setae Z5. On the middle of the dorsum, just behind setae j6, there is a characteristic structure resembling a hump with three terminal digitiform projections; 50 (40–58) long, 35 (33–38) wide at the base (Fig. 7B). Opisthonotum with five pairs of setae (Z1, S5, Z4, Z5, J5). Dorsal setae of different length and morphology; setae j1, s4, Z4 and Z5 thick, strongly lobulated and longer; the remaining short or very short and smooth. Setae j1 broad cylindrical, apically rounded and with 8–10 digitiform barbs (Fig. 7C); setae j3, Z1 and J5 very short and hook-like; setae j4–j6 short and spine-like; setae z2 broad finger-like (Fig. 7D); setae S5 finger-like (Fig. 7A); setae s4 the longest on the shield, very broad, club-shaped and with 20–25 well developed lobulate barbs, most of them concentrated on the terminal club (Fig. 7E); setae Z4 similar in appearance to those s4 (7F); Z5 without club, with more developed lobes. All setae placed on tubercles, which are strongly developed at the base of s4, Z4 and Z5; the tubercle of Z5 is subpyramidal and erect in alive specimens, 28 (23–32) high, 56 (51–62) wide at the base. Length of setae j1 37 (35–42), j3 5 (4–7), j4 3 (3–4), j5 4 (4–5), j6 4 (3–4), J5 5 (4–5), z2 7 (6–8), z4 5 (4–5), z5 4 (4–5), Z1 6 (5–6), Z4 84 (79–89), Z5 95 (91–99), s4 104 (101–108), S5 7 (6–9). Sublateral setae r3 and R1 placed on the soft integument and very different in size and shape; r3 broad, club-shaped, asymmetrical, smooth dorsally, with 5–6 lateral digits, 10–12 ventral digits and a rounded apex, 25 (23–28) long (Fig. 8A); R1 short, smooth, finger-like, 6 (6–7) long.
Dorsal adenotaxy and poroidotaxy difficult to see because of the irregular surface and the small size of pores and poroids. Six pairs of solenostomes (gd1, gd2, gd4, gd6, gd8, gd9); pore gd1 closest to j3 than to z2; gd2 very near to the base of z4, fundibuliform and externally lip-shaped; posteroantiaxial and near the tubercle of s4 (Fig. 8B); gd9 anterior and adjacent to S5. Peritremes thin and sinuous, reaching the insertions of setae j1, 179 (170–186) long; with a characteristic open curve in the distal part; peritrematal grove narrow, with two lines of small microvilli.
Ventral idiosoma — (Fig. 6B). Base of tritosternum 11 (10–13) wide; total length of tritosternum 80 (78–84). Sternal shield 68 (65–72) long, 81 (77–86) wide; distance st1–st3 63 (61–64), st2–st2 72 (69–75). Anterior and posterior margins poorly sclerotized, difficult to see; posterior margin most probably straight or slightly concave. Setae st4 on ovoid metasternal plates. Epigynal shield 100 (96–103) long; distance st5–st5 70 (66–73). Narrow sclerotization between epigynal and ventrianal shields. Ventrianal shield pentagonal and smooth, 112 (97–118) long, 69 (62–77) wide at level of ZV2, 74 (71–76) wide at level of anus; anterior margin straight between setae JV1; anterior corners between JV1 and ZV2 rounded; laterals smoothly convex posterior to setae ZV2. Three pairs of short preanal setae; insertions of setae JV2 slightly posterior to the level of ZV2; usually setae JV1 and ZV2 in marginal position. All the setae on the shield setiform, except the postanal, which is finger-like. Preanal pores very small and punctiform, posteroantiaxial to setae JV2; 38 (35–40) distant each other. Four pairs of setae (ZV1, ZV3, JV4, JV5) and five pairs of poroids surrounding the ventrianal shield. Setae ZV1 and ZV3 pointed, JV4 blunt, JV5 with the same aspect than those longer dorsal setae, on tubercles, 62 (56–65). Anterior metapodal plate short and narrow 7 (6–8); posterior metapodal elongate 21 (18–23).
Spermathecal apparatus — (Fig. 6C). Major duct tubular; atrium subconical; calyx hemispherical, thin walled, 6 (5–6) deep, 11 (9-13) wide at the junction with the vesicle.
Gnathosoma and chelicerae — (Fig. 6D). Length of gnathosoma 170–179; width 79–82. Gnathotectum dome-shaped. Corniculi straight and subparalell, length 36, 36 (34–38). Setae h1 17–18, h2 18–20, h3 21–23, pc 25¬–28. Deutosternal groove with six rows of two lateral denticles, except the distal septum, which has three, two lateral, one median denticles. Short cheliceral digits; fixed digit 21–23, with eight to nine teeth; movable digit 24–26, with three or four teeth. Palps 97 (93–112) long.
Legs — (Fig. 6E). Leg I 287 (274–302); leg II 239 (228–251); leg III 231 (205–247); leg IV 397 (388–425). The legs bear a combination of longer, setiform and pointed setae and shorter, finger-like or hooked blunt setae. In general, dorsal setae on the segments are shorter and finger-shaped, lateral setae are a bit longer and blunt, and ventral setae are longer, setiform and pointed. Setal formula for legs I–IV (coxae to basitarsi) 2-5-12-10-10-0, 2-5-10-8-7-4, 2-5-6-7-7-4, 1-5-6-7-6-4. Genu II with eight setae (2 2/1, 2/0 1), genu III with seven setae (1 2/1, 2/0 1). No macrosetae on leg IV, but dorsal setae on femur, tibia and basitarsus are short, thick and digitiform.
Description
Male
(n=4) (Figures 8–9)
Dorsal idiosoma — (Fig. 9A). Dorsal shield fusiform, 241 (234–246) long, 162 (156–170) wide; lateral margins irregular and sinuous; peritrematal shield fused to the dorsal plate at the level of setae r3. Ornamentation of the shield similar to that of female but less accentuated. The same adenotaxy and poroidotaxy than in females (six pairs of solenostomes, with the absence of gd5); gland opening gd3 hypertrophied, elliptical, on the margin of dorsal shield or on the soft integument (Fig. 8C). Dorsal hump posterior to setae j6 absent. Nineteen pairs of dorsal setae; setal morphology similar to that of female, but shorter in the case of the thick and serrated j1, s4, Z4 and Z5. Most of the dorsal setae and the sublateral r3 placed on tubercles. Setae j1 29 (26–32), j3 5 (4–5), j4, j5, j6 4 (4–5), J5 6 (5–7), z2 5 (4–5), z4 6 (5–7), z5 3 (3–4), Z1 5 (5–6), Z4 24 (21–27), Z5 39 (38–40), s4 50 (46–53), S5 5 (5–6), r3 15 (13–16), R1 5 (4–7). Peritremes very narrow, sinuous, with the same layout described for the female; 161 (154–169) long, extending forward to the insertion of setae j1; one or two rows of microvilli.
Ventral idiosoma — (Fig. 9B). Sternogenital shield smooth, with five pairs of setae and two pairs of poroids; 124 (122–127) long, 69 (66–72) wide at the level of setae st3. Distance st1–st3 62 (60–64), st2–st2 45 (42–48). Anterior margin concave, posterior margin slightly convex. Ventrianal shield subtriangular and smooth or with weak anterior striae; 97 (94–99) long, 152 (148–154) wide at level of anterior corners. Four pairs of preanal setae (JV1, JV2, ZV2, JV4) and four pairs of poroids; setae JV4 in marginal position. Preanal pores minute and punctiform, posteroantiaxial to setae JV2, 34 (33–38) separated from each other. Setae JV5 short, smooth and finger-like (Fig. 8D); 8 (7–9) long.
Chelicerae and spermatodactyl — (Fig. 9C). Fixed digit 17 (16–18) long, with seven–eight teeth (one subapical plus six–seven aligned); movable digit 18 (17–18) long, with one strong tooth. Spermatodactyl L-shaped; shaft straight, 15 (14–15); heel prominent; foot long 9 (9–10), with lateral projection; toe turned up.
Legs — Leg chaetotaxy, setal formula of genua II and III, and combination of short and longer setae as in female. Without macrosetae on leg IV.
Deutonymph
(n=1) Figure 10
Dorsal and lateral idiosoma — (Fig. 10A). Dorsal shield elongate, 208 long, 114 wide. Shield surface irregular without the characteristic reticulate patches of the adults, mostly smooth, with anterolateral striae. Dorsal hump absent. Same adenotaxy as in females. Fourteen pairs of dorsal setae, all smooth and minute, except the tuberculate j1, s4, Z4 and Z5, which are longer, thick and serrated, repeating the same pattern as in females, but with setae Z4 shorter compared to s4 and Z5. Serration of the longer setae more reduced than in females; j1 with six barbs; s4 with about 20 barbs; Z4 with 8–10 barbs; Z5 with 14 projections. Lengths of setae: j1 24, j3 4, j4–J5 3, z2–z4 4, z5–Z1 3, Z4 18, Z5 29, s4 41, S5 3. Sublaterals r3 and R1 on the interscutal membrane; r3 cylindrical, with 8 barbs, 17; R1 smooth, 3. Peritremes reaching the base of setae z2.
Ventral idiosoma — (Fig. 10B). Coxisternal region with five pairs of setae. Six pairs of setae around to the anal opening in addition to the perianal and postanal setae. Setae JV5 short and smooth, 8 long. Preanal pores not discernible.
Gnathosoma and chelicera — Deutosternal groove with eight rows of denticles. Fixed digit of chelicerae 18 long, with eight teeth; movable digit 17 long, with two teeth.
Legs — With the same chaetotaxy as in females. Macroseta on basitarsus IV differentiate, 14 long, curved, smooth and knobbed.
Remarks
Proprioseius gibbus exhibits one of the strangest habitus in the entire family Phytoseiidae. A unique feature of this species is the existence of a well-developed hump in the middle of the dorsal shield (less developed but also present in the Brazilian relatives P. aculeatus Moraes & Denmark (Moraes and Denmark, 1999) and P. retroacuminatus Zacarias & Moraes (Zacarias and Moraes, 2001)). The base of this structure is somewhat flexible and the hump is oriented in different positions in the mounted specimens, but is erect and plump in live females. Nothing is known about the possible function of the structure, but given that only adult females have it, we can hypothesize that could be a reservoir of fatty cells, proteins or other nutrient substances used by the female to promote the development of oocytes or the oogenesis during the oviposition period. Significant variation of hump size has not been observed. No ducts or structures assimilated to glands are visible inside of or at the base of the hump.
Furthermore, the posterior part of the dorsal shield is elevated and the insertions of the non-tuberculate setae J5 are anterior to the sockets of the tuberculate setae Z5. This characteristic shape results from the strong protruding tubercles from which the dorsal setae Z4 and, particularly, Z5 arise.
Interestingly, there is a sexual dimorphism in the development of setae JV5. This seta is long, thick, flexible, and serrate in the female; on the contrary, it is very short, thin, rigid, smooth, and finger-like in the male.
Setal sizes and other morphological traits in the collected females are similar to those presented in the original description, except for the shorter setae z2, s4 and Z4 (about 10% shorter in our specimens).
Tribe Typhlodromipsini Chant & McMurtry, 2005
Genus Typhlodromips De Leon, 1965
Typhlodromips ariri Gondim Jr. & Moraes
Typhlodromips ariri Gondim Jr. and Moraes, 2001: 82
Specimens examined
Ten females on Clethra scabra (Clethraceae); forest clump next to Juquinha monument, road MG-010 between Cardeal Mota and Conceição do Mato Dentro, Serra do Cipó, Minas Gerais; 19°15′17″S, 43°33′10″W, 1354 m asl; 18 August 2011.
Collected specimens deposited in the Mite collection of the ESALQ-USP (Escola Superior de Agricultura ''Luiz de Queiroz″, Universidade de São Paulo, Piracicaba, State of São Paulo, Brazil), Laboratory of Acarology, Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Spain, and the Mite collection of the Centre de Biologie pour la Gestion des Populations (UMR CBGP), Montferrier sur Lez, France.
Geographical distribution
Brazilian endemic species restricted so far to São Paulo state. This is the first record in Northeast Brazil.
Complementary description
Female
(n=10) (Figure 11)
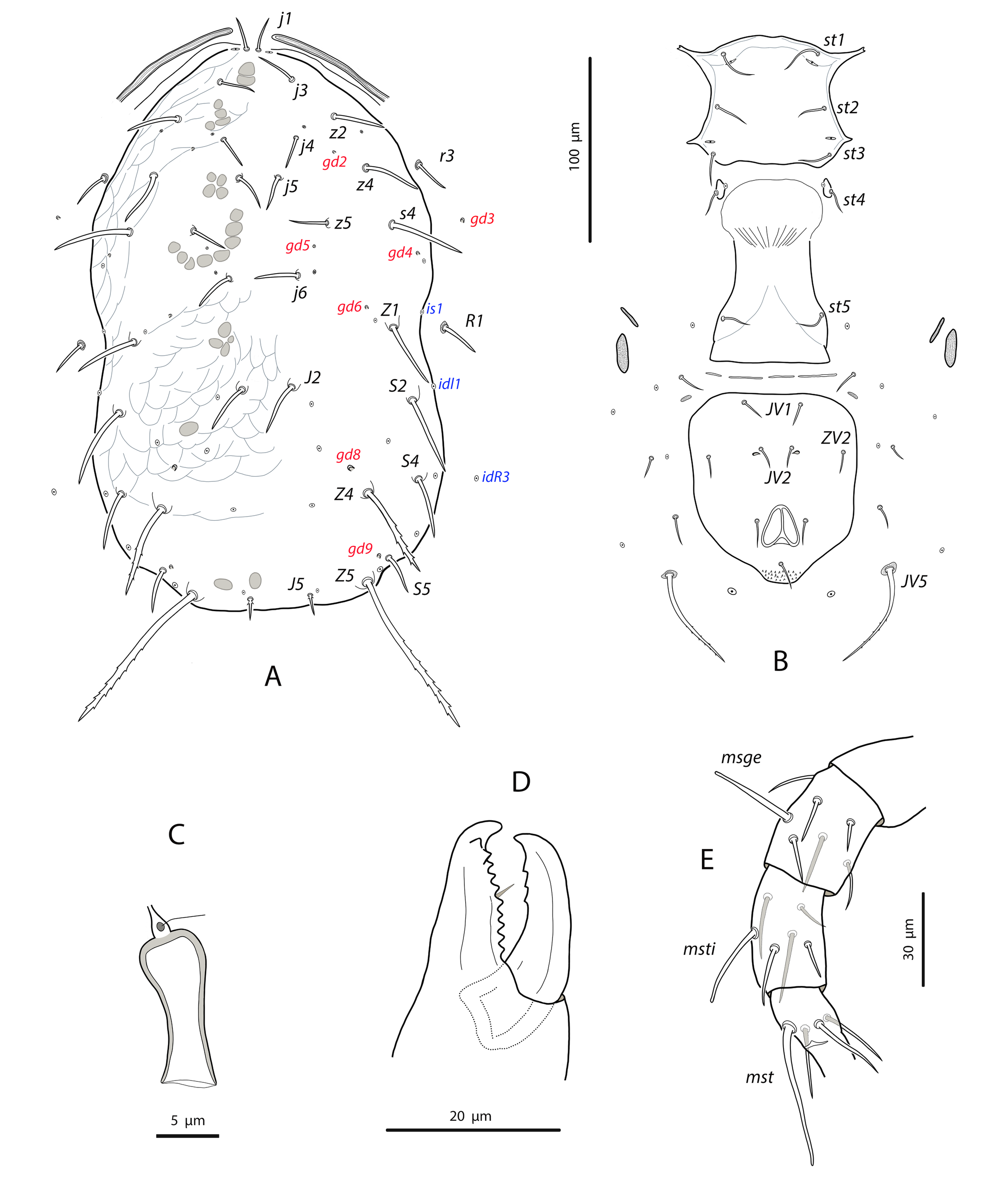
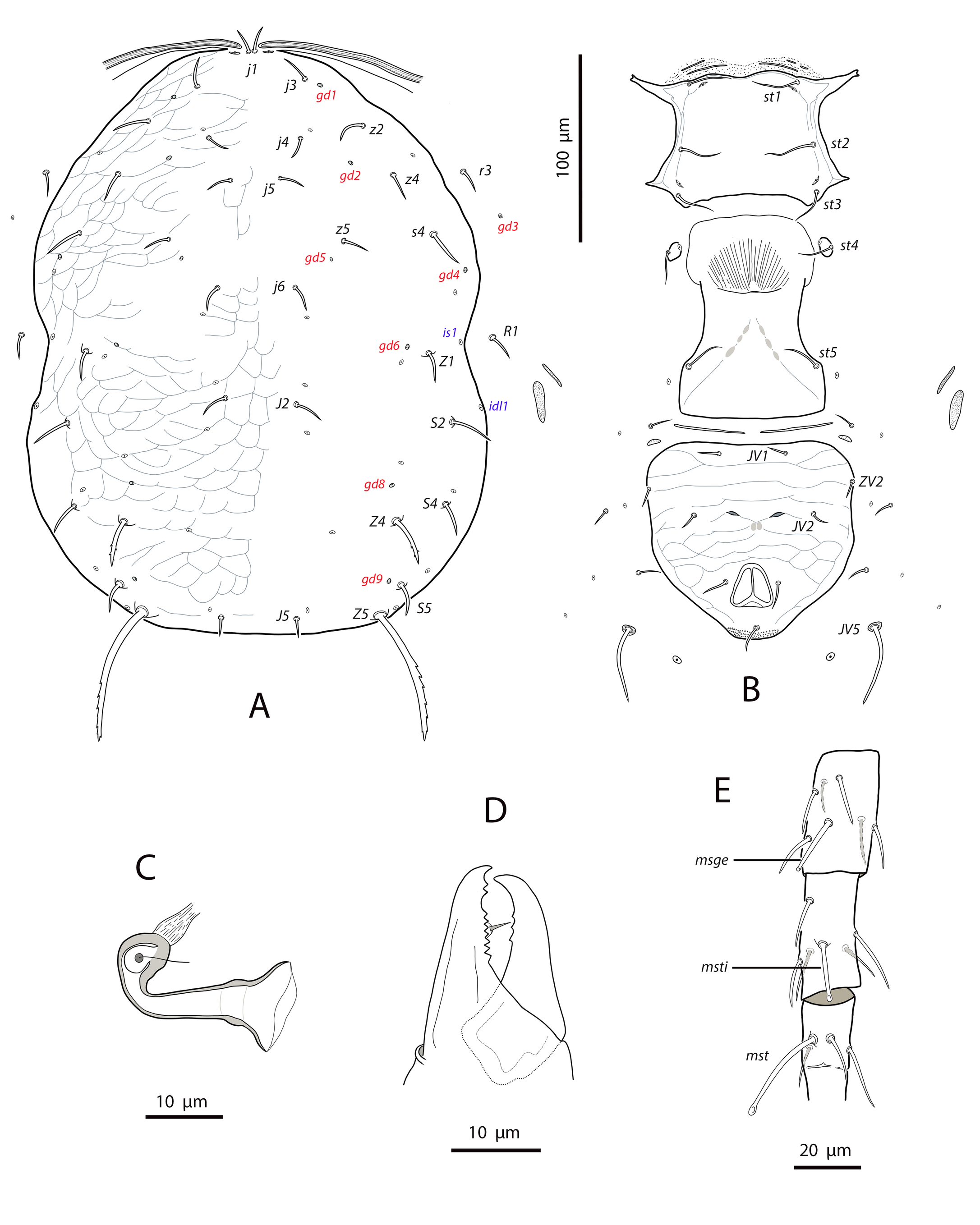
Dorsal and lateral idiosoma — (Fig. 11A). Dorsal shield suboval, 296 (287–304) long, 170 (165–176) wide; posterolateral margins at the level of setae S5 concave. Shield surface striated anterolaterally, forming several closed cells, weakly striate-reticulate on the remaining podonotum, distinctly imbricate between setae Z1 and Z4, almost smooth posterior to Z4. Seventeen pairs of dorsal setae, being s4, Z1, S2, Z4 and Z5 the longest; all smooth and blunt, except Z4 and Z5 serrate on the distal part (about 8–10 barbs in Z4, 16–18 barbs in Z5) and J5 proximally barbed. Setae j1 18 (17–19), j3 23 (22–25), j4 16 (16–17), j5 17 (16–18), j6 21 (20–22), J2 25 (24–28), J5 9 (9–10), z2 25 (22–28), z4 27 (27–28), z5 17 (16–18), Z1 31 (28–32), Z4 44 (40–47), Z5 86 (80–90), s4 38 (36–41), S2 39 (38–41), S4 28 (27–30), S5 22 (20–23). Most of dorsal setae arising from tubercles. Six pairs of dorsal solenostomes, gd2, gd4, gd5, gd6, gd8 and gd9; gland openings small and punctiform, except gd8 funnel-shaped and gd9 crescentic; pore gd6 difficult to see in some females; gd5 posteroparaxial to setae z5, gd9 next to and anteroparaxial to S5. Sixteen pairs of dorsal poroids; is1 and idl1 in marginal position. Peritremes reaching almost the bases of setae j1; 171 (168–177) long; groove with only two rows of microvilli. Sublateral setae subequal, placed on small microsclerites; r3 18 (17–19), R1 17 (17–18). Solenostome gd3 placed on the striated integument between the peritrematal and dorsal shields, at the level of setae s4. Poroids idR3 at the level of setae S4.
Ventral idiosoma — (Fig. 11B). Base of tritosternum 9 (6–10) wide; plumose laciniae reaching the fourth row of deutosternal denticles, 66 (60–70) long, including the fused proximal section, 29 (26–34). Sternal shield subquadrate; 65 (64–65) long, 66 (64–69) wide. Distance setae st1–st3 56 (55–57); st2–st2 58 (57–61). Anterior margin of the shield convex, posterior margin straight or slightly convex. Surface mostly smooth, with a few striae near the anterior and lateral margins; with three pairs of setae and two of poroids. Setae st4 on isolated platelets. Epigynal shield 107 (101–112) long, 67 (66–68) wide at level of posterior corners; distance between setae st5 56 (54–58). Ventrianal shield broad pentagonal, wider than epigynal; anterior corners rounded; 102 (101–104) long, 92 (91–95) wide at the level of setae ZV2 and 83 (78–85) at the level of anus; distance between preanal pores 21 (19–22). Shield evenly smooth, bearing three pairs of setae and a pair of crescentic pores; setae JV1 and ZV2 separated from the shield margin; preanal pores posterior or posteroantiaxial and adjacent to the insertions of setae JV2. Four pairs of setae and five pairs of poroids on the caudoventral integument around the shield; setae JV5 barbed and placed on oval sclerites, 55 (52–59) long. Small and elongate microsclerites near the anterior corners of the shield. Anterior metapodal shield narrow and elongate, 14 (13–15), posterior metapodal elliptic, 19 (17–21).
Spermathecal apparatus — (Fig. 11C). Major duct thin, inconspicuous in the proximal part; atrium small, in a conic structure at the junction of major duct and the base of calyx; calyx tubular with the proximal part wider than the distal part, 12 (12–13) long, 5 (5–7) wide at the base, 3 (3–4) wide distally.
Gnathosoma —(Fig. 11D). Length 143–149; width 80–89. Gnathotectum convex, margin regular and smooth. Corniculi 28–31 long, slightly divergent. Setae h1 15–18, h2 17–18, h3 14–15, pc 20¬–22. Deutosternal groove with seven rows of two denticles placed on the lateral margins of each row. First cheliceral segment 22–25 long, 28–29 wide; second segment 62–67 long, 26–28 wide. Fixed digit of chelicera 25 (24–27) long, with two subapical teeth and a comb of 8–9 aligned teeth; movable digit 26 (25–28), with three denticles. Palps 86–92 long.
Legs — (Fig. 11E). Genua II and III with 7 setae; setal formulae 2 2/0, 2/0 1 and 1 2/1, 2/0 1, respectively. The erect seta ad1 on tibia III can be considered as a macroseta, as it is clearly longer than the remaining seta on the segment, 19 (18–22); macroseta on genu III, 18 (17–19) long. Leg IV with three blunt macrosetae, erect macroseta on genu 27 (25–29); macroseta on tibia 27 (25–28), on basitarsus, 44 (42–45).
Remarks
Typhlodromips ariri can be easily differentiated from related species by the following features. (1) the tubercles from which most of the dorsal setae (mainly opisthosomal setae) arise, and the noticeable length of setae Z5; (2) the shape of the ventrianal shield and the position of preanal pores; (3) the shape of the spermathecal calyx, and (4) the concavity in the lateral margin of the dorsal shield at the level of setae S5. The last character is very original in the genus Typhlodromips and is reminiscent of many species in the genus Typhlodromalus Muma (Chant and McMurtry, 2005b).
The specimens collected are morphologically similar to those of the original description. Only dorsal setae Z1, Z5, s4, and macrosetae on leg IV are slightly longer. The presence of tubercles at the bases of the dorsal opisthosomal setae was not mentioned in the original description.
Typhlodromips constrictatus (El-Banhawy, 1984)
Amblyseius constrictatus El-Banhawy, 1984: 134
Neoseiulus constrictatus Moraes et al., 1986: 76
Typhlodromips constrictatus Chant and McMurtry, 2005a: 325
Specimens examined
Fourteen females, five males on an unidentified Lauraceae; forest clump next to Juquinha monument, road MG-010 between Cardeal Mota and Conceição do Mato Dentro, Serra do Cipó, Minas Gerais; 19°15′17″S, 43°33′10″W, 1354 m asl; 18 August 2011.
Collected specimens deposited in the Mite collection of the ESALQ-USP (Escola Superior de Agricultura ''Luiz de Queiroz″, Universidade de São Paulo, Piracicaba, State of São Paulo, Brazil), Laboratory of Acarology, Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Spain, and the Mite collection of the Centre de Biologie pour la Gestion des Populations (UMR CBGP), Montferrier sur Lez, France.
Geographical distribution
Previously known from Brazil, collected in the states of Bahia, Espírito Santo and Minas Gerais.
Complementary description
Female
(n=10) (Figures 12–13)
Dorsal and lateral idiosoma — (Fig. 12A). Dorsal shield peanut-shaped, with a conspicuous constriction at level of setae Z1; 322 (306–337) long, 202 (194–210) wide at the level of s4, maximum width of the podoscutum 221, maximum width of opisthoscutum 227, 196 wide at the level of constriction. Dorsal surface reticulate; elongate cells covering anterolateral margins, weak median cells between setae j4 and J2, scale-like cells covering the entire opisthoscutum. Dorsal setae smooth and pointed, except Z4 and Z5, which are conspicuously barbed in the distal part (6¬–8 barbs in Z4, about 18 strong barbs in Z5) and J5 basally barbed. All the dorsal setae on the opisthosoma, except J2 and J5, are placed on tubercles. Setae j1 18 (16–18), j3 17 (16–19), j4 11 (11–12), j5 11 (10–12), j6 12 (12–13), J2 15 (14–17), J5 9 (9–10), z2, z4 14 (13–16), z5 11 (10–12), Z1 16 (15–17), Z4 25 (23–26), Z5 61 (60–62), s4 18 (16–19), S2 18 (16–18), S4, S5 16 (15–17). Sublateral setae r3 and R1 subequal, 14 (13–15) and situated on small sclerites. Dorsal shield with seven pairs of solenostomes and sixteen pairs of poroids. Solenostomes gd4, gd5 and gd6 minute, difficult to see; gland opening gd9 anteroparaxial to setae S5. Peritremes 197 (187–203) long, extending forward to setae j1; internal groove with two lines of microvilli.
Ventral idiosoma — (Figs. 12B, 13A). Presternal area with well-developed granulate ornamentation (Fig. 13A). Sternal shield 66 (62–71) long, 78 (75–82) wide¬. Distance st1–st3 61 (60–61), distance st2–st2 67 (64–69). Anterior margin concave medially; posterior margin straight or slightly concave; longitudinal striae near the lateral margins. Sockets of setae st1 on the anterior margin of the shield; setae st4 placed on almost hemispherical platelets off the shield. Epigynal shield 107 (105–111) long, 66 (62–70) wide at posterior corners; distance st5–st5 59 (57–60). Transversal thin sclerotization between epigynal and ventrianal shields; elongate microsclerites on the soft integument, near the anterior margins of the ventrianal shield. Anterior (secondary) metapodal shield 15 (14–15) long, posterior (primary) 21 (19–22) long. Ventrianal shield broad pentagonal, wider than the epigynal shield; anterior and lateral margins straight; 107 (99–114) long, 105 (100–113) wide at level of ZV2, 81 (79–87) wide at level of anus. Surface of the shield striated anteriorly, reticulated posterior to setae JV2. Preanal pores crescentic, situated between setae JV2 or somewhat posteroparaxial to the insertions of those setae; distance between pores 18 (18–19). Four pairs of setae and five pairs of poroids surrounding the shield. Setae JV5 smooth, distally blunt, on sclerites, 40 (39–41) long.
Spermathecal apparatus — (Fig. 12C). Major duct broad and membranous; atrium well-developed, as wide as the base of calyx, incised longitudinally in most of its length; calyx short tubular, diverging smoothly through the vesicle, 9–10 long, 8 wide next to vesicle.
Gnathosoma —(Figs. 12D, 13B). Gnathosomal length 149–161; width 86–91. Margin of gnathotectum more or less straight, smoothly undulated (Fig. 13B). Setae h1 22–24, h2 21–22, h3 23–25, pc 26–28. Deutosternal groove with six rows of two denticles placed on lateral margins of each row. Fixed digit of chelicera 26–29 long, with two subapical teeth and a comb of eight–nine teeth; movable digit 23–26, with three teeth. Palps 101–106 long.
Legs (Figs. 12E, 13C–E). Genu II with seven setae (2 2/0, 2/0 1); genu III with seven setae (1 2/1, 2/0 1); tibia I with ten setae (2 2/1, 2/1 2). Macrosetae on legs II and III hardly to differentiate; only those on leg IV are clearly recognizable. Macroseta on genu IV 24 (23–25), on tibia IV 15 (14–18), on basitarsus IV 34 (33–36).
Description
Male
(n=5) (Figure 14)
Dorsal idiosoma — (Fig. 14A). Dorsal shield 261 (256–265) long, 171 (166–179) wide. Ornamentation of the shield similar to that of female. Adenotactic and poroidotactic pattern as in females; gland opening gd3 present, oval button-shaped, between setae s4 and the shield margin. Peritrematal shield fused to dorsal shield at the level of setae r3; peritremes 169 (165-173) long, extending forward to the bases of setae j1; microvilli aligned in two rows. Nineteen pairs of dorsal setae, all smooth except Z5, barbed in the distal half. Most of setae on tubercles, tubercles more accentuated at the base of opisthosomal setae. Setae j1 15 (15–16), j3 18 (17–19), j4 11 (10–12), j5 10 (10–11), j6 11 (10–13), J2 14 (13–14), J5 8, z2 13 (12–14), z4 14 (13–15), z5 10 (9–11), Z1 14 (12–16), Z4 22 (20–24), Z5 43 (41–47), s4 16 (15–16), S2 16 (15–18), S4 16 (13–18), S5 14 (13–15), r3 12 (10–13), R1 13 (12–14).
Ventral idiosoma — (Fig. 14B). Sternogenital shield smooth except for several longitudinal striae on the lateral margins of the shield, 114 (111–116) long, 65 (61–67) wide at the level of setae st3. With five pairs of setae and three pairs of poroids. Posterior margin of the shield straight or slightly convex. Ventrianal shield subtriangular, anterior margin convex; 109 (103–111) long, 137 (131–141) wide at anterior corners. Shield entirely reticulate, with three pairs of preanal setae and four pairs of poroids. Crescentic solenostomes gv3 between setae JV2, 15 (14–16) separated from each other. Setae JV5 smooth, 22 (20–23) long, placed on platelets; poroids ivp between setae JV5.
Chelicerae and spermatodactyl (Figure 14C). Fixed digit 21 (20–22) long, with a subapical tooth and five to six teeth aligned; movable digit 20 (20–21) long, with one tooth. Spermatodactyl L-shaped; shaft straight or slightly curved, 14–15; combined length of foot plus heel 13–14; toe turned up. Vellum present along the shaft.
Legs —. Leg chaetotaxy and formula of genua II and III as in female. Macrosetae blunt to knobbed, that on genu IV 17 (15–17), on tibia IV 15 (14–15), on basitarsus IV 28 (26–30) long. Macrosetae on genu II and III difficult to distinguish from the rest of setae on the segment.
Remarks
The generic placement of T. constrictatus has been modified several times. Originally described as Amblyseius constrictatus by El-Banhawy, it was moved to Neoseiulus in the first catalogue of the family Phytoseiidae by Moraes et al. (1986), and finally transferred to the genus Typhlodromips by Chant and McMurtry (2005a). The later authors emphasized that species in Neoseiulus and Typhlodromips are morphologically similar in many aspects, but Typhlodromips can be separated by the presence of macrosetae on other legs in addition to leg IV and by the multidentate fixed digit of chelicera. However, the existence of macrosetae on genua II and III is not so evident in many cases, and T. constrictatus is a good example (see Fig. 13 D–E). Macrosetae on legs II and III from many Typhlodromips species are not longer than the remaining setae on the segment (sometimes are shorter) and can only be differentiate by subtle details, as their greater thickness and the end of the structure, which is blunt or knobbed in contrast with the other pointed setae. This fact was already discussed by Beard (2001) in her review of the Australian Typhlodromips.
The original description of the species by El-Banhawy was relatively complete, but the shape of the spermatheca could not be seen, as the calyx was facing up in the mounted specimens. Later, Chant and McMurtry (2005a) divided the genus Typhlodromips into five species groups based on the presence/absence of setae J1 and the morphology of the spermatheca; by this reason they could not include T. constrictatus in any of the proposed species groups. According to the spermathecal apparatus described here and the absence of setae J1, T. constrictatus can be placed in the ariri species group. This species has been reported several times in Brazil since its description in 1984 (Lofego, 1998; Silva et al. 2010; Nuvoloni et al. 2015); nevertheless, these publications do not include information on the female spermatheca and only Lofego (1998) shows an illustration of its morphology, which is coincident with that we describe in this study.
Length of dorsal setae in the examined specimens is similar to that of the original description, except for the setae j1 and macrosetae on leg IV, which are somewhat longer, and coincides with those provided by Lofego (1998) for specimens from Bahia state.
Typhlodromips japi Lofego, Demite & Feres
Typhlodromips japi Lofego et al., 2011: 112.
Specimens examined
Six females on Solanum lycocarpum (Solanaceae); area of cerrado, Cardeal Mota, Serra do Cipó, Minas Gerais, 19°18′25″S, 43°36′26″W, 1015 m asl; 18 August 2011.
Collected specimens deposited in the Mite collection of the ESALQ-USP (Escola Superior de Agricultura ''Luiz de Queiroz″, Universidade de São Paulo, Piracicaba, State of São Paulo, Brazil), and the Laboratory of Acarology, Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Spain.
Geographical distribution
Brazilian species restricted up to now to the states of São Paulo and Rio Grande do Sul.
Complementary description
Female
(n=6) (Figures 15–16)
Dorsal and lateral idiosoma — (Fig. 15A). Dorsal shield suboval; lateral waist at the level of setae Z1 very pronounced; lateral margins regular. Shield mostly reticulate, with elongate cells; ornamentation weak or absent in the median area between setae j1–j3 and between insertions of setae j4–j6; imbricated cells between setae j6–J2 and posterior to the latter setae. Measurements of the shield 309 (300–322) long, 183 (174–194) wide. Seventeen pairs of short, smooth and pointed setae, except Z4 and Z5, the longest on the shield and distally barbed; setae J5 barbed basally; setae Z5 blunt. At least Z1, Z4, S2, S4 and S5 placed on tubercles. Measurements of setae j1 15 (11–18), j3 16 (14–18), j4 10, (9–11), j5 11 (10–12), j6 12 (11–12), J2 13 (12–14), J5 10 (9–10), z2 14 (13–16), z4 14 (13–14), z5 11 (10–11), Z1 13 (11–14), Z4 23 (22–24), Z5 67 (63–71), s4 18 (16–19), S2 18 (17–20), S4 16 (14–18), S5 15 (14–16). Seven pairs of dorsal solenostomes; gland pore gd9 mesad to the insertions of S5. Sixteen pairs of poroids, with is1 and idl1 in marginal position. Peritremes extending forward to level of setae j1; 180 (177–183) in length; internal groove with two longitudinal lines of microvilli. Sublateral setae similar in length; r3 14 (12–15), R1 14 (11–16); solenostome gd3 on the interscutal membrane, somewhat anterior to the level of setae s4.
Ventral idiosoma — (Fig. 15B). Base of tritosternum 9 (8–9) wide; plumose laciniae reaching the sixth row of deutosternal denticles, 64 (63–66) long, including the fused proximal section. Presternal area granulate. Sternal shield longer than wide; 60 (58–61) long, 72 (70–74) wide; anterior margin concave medially, posterior margin moderately concave; shield smooth, with striae at the base of anterior arms and along the laterals, with three pairs of setae and two pairs of poroids; distance st1–st3 56 (55–57), distance st2–st2 61 (59–62). Setae st4 on ovoid sclerites. Epigynal shield smooth; 102 (101–104) long, 65 (61–69) wide at posterior corners; distance st5–st5 59 (59–60). Ventrianal shield broad pentagonal, 103 (97–107) long; 102 (96–104) wide at level of ZV2; 75 (73–77) wide at level of anus. Anterior margin of the shield slightly concave between insertions of setae JV1, anterolateral corners rounded, lateral margins straight or very slightly concave, narrower at the level of preanal pores. Shield transversally striate anterior to preanal pores, reticulate between preanal pores and anus. Three pairs of short preanal setae; insertions of JV1 separated from the shield margin, ZV2 inserted just on the margin; JV2 at the level of or slightly anterior or slightly posterior to preanal pores. Preanal pores crescentic, distance between them 21 (20–23). Elongate microsclerites near the anterolateral corners, on the soft integument; four pairs of setae and five pairs of visible poroids around the shield. Setae JV5 smooth, blunt, on oval sclerites, 38 (33–41) long. Anterior metapodal shield elongate and very thin, 16 (14–19) long; posterior metapodal elliptical, 20 (19–21).
Spermathecal apparatus — (Figs. 15C, 16A). Major duct membranous, almost as wide as the proximal part of calyx; atrium globular, vacuolated, 6 long, 6–7 wide; calyx long tubular, 23 (22–24) long per 9 (8–9) wide, with parallel arms, only flaring in the distal part, near the vesicle. Calyx curved or angled at the level of the first proximal third in all the females examined, forming an angle of up to 90 degrees.
Gnathosoma —(Figs. 15D, 16B). Length 155–159; width 68–79. Tectum convex, with more or less regular margins (Fig. 16B). Setae h1 17–19, h2 12–14, h3 17–19, pc 20¬–22. Deutosternal groove with seven rows of two denticles placed on the anterolateral margins of each row; the first and second proximal septa bear two and one additional lateral denticles, respectively, while in the seventh septum the denticles are separated from the laterals. First cheliceral segment 23–25 long, 21–24 wide; second segment 63–64 long, 18–20 wide. Fixed digit of chelicera 25 (23–27) long, with two subapical teeth and a comb of seven–nine aligned teeth; movable digit 23 (21–24), with three denticles. Palps 93 (91–95).
Legs — (Fig. 15E). Tarsus I tanned. Genua II and III with seven setae; setal formulae 2 2/0, 2/0 1 and 1 2/1, 2/0 1, respectively. Macroseta on genu III slightly knobbed, 12 (11–13) long. Blunt-knobbed macrosetae on leg IV; on genu 17 (16–18), on tibia 16 (15–16), on basitarsus 31 (28–33) long.
Remarks
According to data of this study the geographical distribution of T. japi has been extended northward, until the state of Minas Gerais. This species is most similar to T. corniformis Moraes, Barbosa & Castro and T. constrictatus. There are minimal differences in setal length, size of dorsal and ventral shields, cheliceral dentition and leg chaetotaxy between T. japi and T. cornifornis, and only the structure of the atrium in the spermatheca appears to be different, enlarged and vacuolated in the former species and inconspicuous in the latter. The eventual synonymy between these species needs to be evaluated in more depth. The external habitus of T. constrictatus is also similar to that of T. japi, with equivalent lengths of setae and other characters, but the shape of the spermatheca, short subconical in the former and curved elongated in the latter, is clearly discriminant and serves to differentiate unequivocally both species.
Typhlodromips japi appears to have a promiscuous mating system. The vesicles of all the females were filled with several spermathophores, in a number from two to seven (Fig. 16A). There were no big differences between the number of spermatophores inside the right and left vesicles in the same female, and the average number of those packets of sperm was 5.25 per vesicle. All the females showed the calyx curved in the same orientation and angled at the same point; that is why we consider this a characteristic morphological feature in the species rather than an artefact due to the mounting process.
The studied females are morphologically similar to those of the species description collected in São Paulo, and the setal measurements fit the original description and data provided later by Gonçalves et al. (2015) and Rocha et al. (2015) for specimens from Rio Grande do Sul.
Typhlodromips ovametapodalis Ferragut sp. nov.
ZOOBANK: 9E7DBABE-7205-41C8-8D07-209B8B230803 ![]()
Type material
Female holotype, seven females paratypes and one male paratype on Paralychnophora bicolor (Asteraceae); pedestrian grasslands, track to Cachoeira de Fumaça, Vale do Capão, Chapada Diamantina, Bahia, 12°36′17″S, 41°28′19″W, 1321 m asl; 12 January 2010.
Holotype, three females and one male paratypes deposited in the Mite collection of the ESALQ-USP (Escola Superior de Agricultura ''Luiz de Queiroz″, Universidade de São Paulo, Piracicaba, State of São Paulo, Brazil). Other paratypes in the Laboratory of Acarology, Instituto Agroforestal Mediterráneo, Universitat Politècnica de València, Spain, and the Mite collection of the Centre de Biologie pour la Gestion des Populations (UMR CBGP), Montferrier sur Lez, France.
Diagnosis
Typhlodromips species with female dorsal shield 311–327 μm long; mostly reticulate, striate laterally, almost smooth in the posterior region. Dorsal setae relatively thick and blunt; all smooth, except Z4 and Z5 distally barbed and J5 proximally barbed. All the dorsal setae placed on tubercles. Setae j3 1.6 times longer than j1; s4 2.7 times longer than Z1; S4 and S5 subequal; Z5 1.1 times longer than Z4. Seven pairs of dorsal solenostomes. Peritremes extending in between the insertions of j1. Ventrianal shield subpentagonal, smooth, with three pairs of preanal setae and a pair of punctiform solenostomes posteroparaxial to setae JV2; setae ZV2 in marginal position. Anterior metapodal shield thin and elongate, posterior oblong or suboval. Spermathecal atrium conical, as wide as the calyx base; calyx elongated, subconical or saccular. Fixed digit of chelicera with two subapical teeth plus ten aligned teeth; movable digit with four teeth. Genu II with eight setae (2 2/1, 2/0 1); genu III with seven setae (1 2/1, 2/0 1). Macrosetae on leg III and IV smooth and blunt, except that on genu IV, which is knobbed. Macrosetae on genu and tarsus IV subequal, longer than that on tibia IV. Male ventrianal shield subpentagonal, transversally striated, with three preanal setae and punctiform solenostomes posteroparaxial to setae JV2. Setae JV2 and ZV2 almost transversally aligned. Fixed cheliceral digit with seven teeth, movable digit with one robust tooth. Spermatodactyl L-shaped.
Description
Female
(n=8) (Figures 17–18)
Dorsal and lateral idiosoma — (Fig. 17A). Dorsal shield peanut-like in shape, as broad at the level of setae s4 as at the level of S2, distinctly constricted at level of setae R1. Shield length 315, 317 (311–327); 193, 193 (182–206) wide at level of s4. Surface evenly imbricate except in the laterals of the opisthosoma, where is smooth or striate. Seventeen pairs of dorsal setae, all blunt and smooth except Z4 and Z5, which are barbed and J5 basally barbed. All the dorsal setae are set on tubercles, the most developed in the base of the longest setae. Setae j3 1.6 times longer than j1; s4 2.7 times longer than Z1; S4 and S5 subequal; Z5 1.1 times longer than Z4. Measurements of dorsal and sublateral setae: j1 28, 26 (25–28), j3 44, 42 (41–44), j4 13, 14 (12–16), j5 17, 14 (12–17), j6 14, 14 (12–16), J2 19, 19 (16–21), J5 11, 12 (11–12), z2 24, 22 (21–24), z4 38, 35 (31–38), z5 13, 12 (10–13), Z1 25, 24 (22–26), Z4 81, 80 (77–82), Z5 88, 88 (86–90), s4 65, 65 (64–67), S2 35, 34 (28–36), S4 24, 23 (21–25), S5 23, 22 (20–24). Seven pairs of punctiform dorsal solenostomes and sixteen pairs of poroids; poroids is1 and idl1 placed on the margin of the shield. Pore gd1 antiaxial to j3; gd5 posteroparaxial to z5; gd9 paraxial to setae S5. Sublaterals similar in length; r3 22, 21 (20–22), R1 19, 20 (18–21). Peritremes 202, 207 (204–214) long, extending forward to the level of j1; internal groove with two longitudinal lines of microvilli.
Ventral idiosoma — (Fig. 17B). Base of tritosternum 11, 11 (10–11) wide; total length of tritosternum 94, 92 (89-94); plumose laciniae 45, 44 (43–46) long. Presternal area smooth or weakly granulate (holotype), with two pairs of thin and transversal sclerites. Sternal shield wider than long; 61, 63 (60–66) long, 79, 79 (72–82) wide; anterior margin concave, posterior margin straight between the posterolateral projections. Shield mostly smooth, with lateral striae, three pairs of setae and two pairs of poroids; distance st1–st3 62, 63 (61–64), distance st2–st2 70, 69 (63–73). Setae st4 and poroids iv3 on metasternal shields. Epigynal shield smooth, 112, 111 (108–116) long, 79, 73 (66–79) wide at posterior corners; distance st5–st5 67, 67 (66–68). Transversal sclerotization between epigynal and ventrianal shields. Ventrianal shield broad pentagonal, 107, 111 (107–116) long; 96, 94 (88–98) wide at level of ZV2; 80, 79 (75–84) wide at level of anus. Anterior margin straight, anterior corners rounded, laterals somewhat concave posterior to the insertions of setae ZV2. Setae JV1 placed close to the anterior margin, setae ZV2 marginal. Surface of the shield smooth, sometimes with perianal striae. Preanal pores punctiform, posterior or slightly posteroparaxial to setae JV2; 41, 40 (36–43) distant each other. Four pairs of setae (ZV1, ZV3, JV4, JV5) and five pairs of poroids (four poroids ivo and ivp) surrounding the ventrianal shield. Setae JV5 blunt, smooth, and placed on sclerites, 58, 58 (53–61). Metapodal plates similar in length but differing in shape; anterior (secondary) narrow and bacillar 16, 16 (15–18) long, posterior (primary) oblong 15, 16 (15–17) long, 9, 9 (8–9) wide (Fig. 18A).
Spermathecal apparatus — (Fig. 17C, 18B). Major duct thin, tubular; atrium conical 7–9 long, as wide as the calyx base; calyx elongated, subconical or saccular, 20, 19 (18–20) long, 13, 12 (11–15) wide at the junction with the vesicle.
Gnathosoma —(Fig. 17D). Length 192–203; width 105–109. Tectum not discernible. Setae h1 24–27, h2 22–23, h3 24–27, pc 26–28. Deutosternal groove with seven septa with two denticles each placed on the anterolateral margins, except the first proximal septa which bears four lateral denticles and the second proximal with three anterolateral denticles. First cheliceral segment 41–45 long, 41–44 wide; second segment 93–99 long. Fixed digit of chelicera 30, 32 (29–33) long, with two subapical teeth and a comb of ten aligned teeth; movable digit 32, 31 (30–34), with four teeth. Palps 124, 120 (118–125).
Legs — (Fig. 17E). Leg I 339, 341 (333–346); leg II 289, 290 (283–292); leg III 282, 283 (273–¬290); leg IV 376, 379 (369–387). Genu II with eight setae (2 2/1, 2/0 1); genu III with seven setae (1 2/1, 2/0 1). Macrosetae on leg III and IV; msge III 27, 25 (23–27), msge IV 49, 48 (44–49), msti IV 33, 31 (30–33), mst IV 54, 49 (45–54). All macrosetae are smooth and blunt, that on genu IV sometimes knobbed.
Male
(n=1) (Figure 19)
Dorsal idiosoma — (Fig. 19A). Dorsal shield 256 long, 181 wide. Ornamentation similar to that of female, although less accentuated. Adenotactic pattern as in females, but openings minute; solenostome gd3 not discernible in the examined specimen. Peritrematal shield fused to dorsal shield anterior to the level of setae r3; peritremes 181 long, terminating at the level of setae j1. Nineteen pairs of dorsal setae, all pointed or blunt and smooth, except Z4 and Z5 barbed and J5 basally barbed. All setae arising from tubercles, more accentuated in the opisthosomal setae. Setae j1 22, j3 32, j4 10, j5, j6 12, J2 15, J5 10, z2 19, z4 27, z5 9, Z1 21, Z4 56, Z5 60, s4 47, S2 26, S4 18, S5 20, r3, R1 18.
Ventral idiosoma — (Fig. 19B). Sternogenital shield smooth with longitudinal striae along the lateral margins, 115 long, 72 wide at the level of setae st3; distance st1–st3 59, st5–st5 39. With five pairs of setae and two pairs of poroids. Posterior margin of the shield slightly convex. Ventrianal shield subtriangular, fused to parapodal shields, anterior margin convex; 109 long, 161 wide at anterior corners. Surface striate-reticulate. With three pairs of preanal setae and four pairs of poroids. Solenostomes gv3 small and punctiform, posterior and slightly paraxial to the sockets of setae JV2, 31 separated from each other. Setae JV5 smooth, blunt, and placed on sclerites, 28 long.
Chelicerae and spermatodactyl — (Fig. 19C). Fixed digit 21 long, with one subapical tooth and a comb of six teeth; movable digit 19 long, with one strong tooth. Spermatodactyl L-shaped; shaft slightly curved; combined length of foot and heel 20; heel prominent, lateral projection present.
Legs —. Leg segment chaetotaxy and formula of genua II and III as in female. Macroseta on genu III barely differentiate (13 long); macroseta on genu IV knobbed, 28; macrosetae on tibia and tarsus IV blunt, 22 and 38 long, respectively.
Taxonomic relationships
Typhlodromips ovametapodalis sp. nov. belongs to the ariri species group. Within this group several species have a saccular calyx and a broad pentagonal ventrianal shield, like T. alpicola (Ehara), T. gimanthus Beard, T. ojibwa (Chant & Hansell), and T. tee (Schicha). However, all these species have setae s4 noticeably shorter (between 30–85% shorter); setae Z5 distinctly longer than Z4 (ratio Z5/Z4 between 1.3–2.2, while in the new species is 1.1); macrosetae on tibia IV longer than on genu IV (except T. tee); dorsal shield with the central area smooth (except T. alpicola); seven instead of eight setae on genu II (except T. objibwa), and none of them has the dorsal shield setae placed on tubercles (Chant & Hansell, 1971; Ehara, 1982; Schicha, 1983; Beard, 2001).
The most similar described species is T. pompeui Gonçalves & Ferla from southern Brazil. Typhlodromips pompeui differs from the new species for having: i) a bigger size, with the dorsal shield 25% longer; ii) dorsal setae not placed on tubercles; iii) setae j3 and Z4 45% shorter; setae S5 70% shorter; setae Z5 23% longer; iv) three pairs of dorsal solenostomes instead of seven in the new species, v) seven setae on genu II instead of eight; vi) preanal pores crescentic instead of punctiform; vii) posterior metapodal shield elongated, no suboval (Gonçalves et al. 2015).
Etymology
The species name refers to the characteristic shape of the posterior metapodal shield, oblong or suboval instead of the more usual elongate shape. It is a compound Latinised noun derived from the words ova meaning ''egg'' and metapodalis referring to the isolated posterior metapodal plate.
Key to females of the Brazilian species of the genus Typhlodromips De Leon
A. Calyx of spermatheca shallow, dish-shaped (coincident with the culmulus species group of Chant and McMurtry, 2005a)
...... 1
B.Calyx of spermatheca relatively short, 4–10 μm, tubular or subconical
...... 2
C. Calyx of spermatheca longer, 10–20 μm, tubular or saccular, usually flaring distally (coincident in part with the ariri species group)
...... 4
D. Calyx of spermatheca elongate, 18–42 long, broad tubular, fundibuliform (funnel-shaped), trumpet-like or slender tubular (coincident with the lugubris species group)
...... 11
E. Major duct in the spermatheca broad membranous, bladder-like, calyx saccular, 13–15 long (matching with the bladderae species group)
...... T. salvadorii Gonçalves & Ferla
1. Genu II with six setae
...... T. mangleae De Leon
— Genu II with seven setae
...... T. pederosus (El-Banhawy) (Setal measurements do not exist for this species in the literature)
2. Calyx of spermatheca tubular or slightly conical
...... 3
— Calyx of spermatheca distinctly conical or funnel-shaped
...... T. fordicey (De Leon), T. cananeiensis Gondim Jr. & Moraes, T. igapo Nuvoloni, Lofego, Rezende & Feres
(On the bases of the original descriptions, re-descriptions and illustrations it is virtually impossible to distinguish between T. fordicey, T. cananeiensis and T. igapo. Differences in setal lengths and other features are minimal if not non- existent. The taxonomical status of these species needs to be re-evaluated with a detailed examination of the respective types).
3. Dorsal shield mostly smooth; preanal pores adjacent to setae JV2; setae Z4 43– 47
...... T. theobromae Souza, Oliveira & Gondim Jr
— Dorsal shield mostly reticulate; preanal pores well-separated from setae JV2; setae Z4 23–26
...... T. constrictatus (El-Banhawy)
4. Preanal setae ZV2 placed posteriorly to setae JV2; angle formed by the lines between setae JV1–JV2 and setae JV2–ZV2 greater than 90°; preanal pores posterior and adjacent to setae JV2
...... T. ariri Gondim Jr. & Moraes
— Preanal setae ZV2 placed anteriorly to setae JV2; angle formed by the lines between setae JV1–JV2 and setae JV2–ZV2 less than 90°
...... 5
5. With a neck between the atrium and calyx of the spermatheca; preanal pores anteroparaxial to JV2
...... T. pallinii Gonçalves, Silva & Ferla
— Without a neck between atrium and calyx; preanal pores posteroparaxial to setae JV2
...... 6
6. Setae s4 and S2 subequal (17–18)
...... T. jucara Gondim Jr. & Moraes
— Setae s4 about 1.5–2.2 times longer than S2
...... 7
7. Setae Z5 shorter, 60–80 long
...... 8
— Setae Z5 longer, 90–120 long
...... 9
8. Setae j3 24 long
...... T. sabaculus Denmark & Muma
— Setae j3 39–44 long
...... T. sinensis Denmark & Muma
9. Genu II with eight setae; dorsal setae on tubercles
...... T. ovometapodalis Ferragut sp. nov.
— Genu II with seven setae; dorsal setae not placed on tubercles
...... 10
10. Setae s4 stout; setae Z5 about 90–92 long
...... T. robustisetus Moraes, Barbosa & Castro
— Setae s4 setiform; setae Z5 110–120 long
...... T. pompeui Gonçalves & Ferla
11. Calyx of spermatheca broad tubular or funnel-shaped, 10–21 long
...... 12
— Calyx of spermatheca trumpet-like or slender and tubular
...... 16
12. Calyx tubular, usually curved and flaring distally
...... 13
— Calyx fundibuliform or funnel-shaped
...... 14
13. Spermatheca with a conspicuous, nodular and enlarged atrium; setae Z4 21–25 long
...... T. japi Lofego, Demite & Feres
— Atrium of the spermatheca inconspicuous; setae Z4 27–32 long
...... T. corniformis Moraes, Barbosa & Castro
14. Dorsal setae s4, S2, Z4, Z5 with pointed tips; on leg IV only macrosetae are knobbed
...... T. furcus Lofego, Demite & Feres
— Dorsal setae s4, S2 (sometimes), Z4, Z5 with expanded tips; leg IV with other knobbed setae in addition to macrosetae
...... 15
15. Genu II with seven setae; major duct thin tubular, with an incomplete ring encircling the duct
...... T. baculiductus Ferragut & Carvalho
— Genu II with ten setae; major duct broad, without ring around the duct
...... T. paramilus Nuvoloni & Lofego
16. Spermathecal calyx 18–24 long, widening gradually from atrium to vesicle; preanal pores anteroparaxial to setae JV2
...... T. amilus De Leon
— Spermathecal calyx 28¬–42 long, slender and tubular, flaring abruptly at the junction with the vesicle; preanal pores between and in transversal line with setae JV2
...... 17
17. Dorsal shield evenly reticulate; calyx of spermatheca 28–30 long
...... T. dentilis (De Leon)
— Dorsal shield smooth in the dorsomedian and posterior regions; calyx of spermatheca 40–42 long
...... T. angustus Guanilo & Moraes
Legal requirements
The biological material was collected under the Brazilian government official authorization conceded to D. Navia by Chico Mendes Institute for Biodiversity Conservation, Ministry of Environment (permanent collection permit No. 20650–1).
Acknowledgements
To Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq), Brazil, for granting the fellowship to the second author (DN) (PP, Process N° 3121322017–4).
References
- Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini. II. Le relevé organotaxique de la face dorsal adulte (Gamasides, Protoadéniques, Phytoseiidae). Acarologia, 27: 20-29.
- Beard J.J. 2001. A review of Australian Neoseiulus Hughes and Typhlodromips De Leon (Acari: Phytoseiidae: Amblyseiinae). Invert. Taxon., 15: 73-158. https://doi.org/10.1071/IT99017
- Beaulieu F., Beard J.J. 2018. Acarine biocontrol agents Neoseiulus californicus sensu Athias-Henriot (1977) and N. barkeri Hughes (Mesostigmata: Phytoseiidae) redescribed, their synonymies assessed, and the identity of N. californicus (McGregor) clarified based on examination of types. Zootaxa, 4500: 451-507. https://doi.org/10.11646/zootaxa.4500.4.1
- Chant D.A. 1957. Descriptions of some phytoseiid mites (Acarina, Phytoseiidae). Part I. Nine new species from British Columbia with keys to the species of British Columbia. Part II. Redescriptions of eight species described by Berlese. Can. Entomol., 89: 289-308. https://doi.org/10.4039/Ent89289-7
- Chant D.A., Hansell R.I.C. 1971. The genus Amblyseius (Acarina: Phytoseiidae) in Canada and Alaska. Can. J. Zool., 49 (5): 703-758. https://doi.org/10.1139/z71-110
- Chant D.A., McMurtry J.A. 2003. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae): Part II. The tribe Kampimodromini Kolodochka. Intern. J. Acarol., 29: 179-224. https://doi.org/10.1080/01647950308684331
- Chant D.A., McMurtry J.A. 2005a. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae) Part VII. Typhlodromipsini n. tribe. Intern. J. Acarol., 31: 315-340. https://doi.org/10.1080/01647950508683673
- Chant D.A., McMurtry J.A. 2005b. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae) Part VI. Tribe Euseiini n. tribe, subtribes Typhlodromalina n. subtribe, Euseiina n. subtribe and Ricoseiina n. subtribe. Intern. J. Acarol., 31: 187-224. https://doi.org/10.1080/01647950508684424
- Chant D.A., Yoshida-Shaul E. 1991. Adult ventral setal patterns in the family Phytoseiidae (Acari: Gamasina). Intern. J. Acarol., 17: 187-199. https://doi.org/10.1080/01647959108683906
- De Leon D. 1965. Phytoseiid mites from Puerto Rico with descriptions of new species (Acarina: Mesostigmata). Fla. Entomol., 48: 121-131. https://doi.org/10.2307/3493102
- Demite P.R., Moraes G.J. de, McMurtry J.A., Denmark H.A., Castilho R.C. 2022. Phytoseiidae Database. Available from: www.lea.esalq.usp.br/phytoseiidae (last access on 24 June 2022).
- Ehara S. 1982. Two new species of phytoseiid mites from Japan (Acari: Phytoseiidae). Appl. Entomol. Zool., 17 (1): 40-45. https://doi.org/10.1303/aez.17.40
- El-Banhawy E.M. 1984. Description of some phytoseiid mites from Brazil (Acarina: Phytoseiidae). Acarologia, 25: 125-144.
- Evans G.O. 1963. Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bull. British Mus. (Nat. Hist.) Zool., 10: 275-303. https://doi.org/10.5962/bhl.part.20528
- Ferragut F., Navia D. 2022. The genus Neoseiulus Hughes (Mesostigmata: Phytoseiidae) in the Espinhaço Range, a great reservoir of biodiversity in Brazil. Zootaxa, 5120: 523-542. https://doi.org/10.11646/zootaxa.5120.4.4
- Gondim Jr M.G.C., Moraes G.J. de 2001. Phytoseiid mites (Acari: Phytoseiidae) associated with palm trees (Arecaceae) in Brazil. Syst. Appl. Acarol, 6: 65-94. https://doi.org/10.11158/saa.6.1.11
- Gonçalves D., Cunha U.S., Bampi P.M., Moraes G.J. de, Ferla N.J. 2015. Phytoseiid mites (Acari: Mesostigmata) from Araucaria Forest of the State of Rio Grande do Sul, Brazil, with new records and descriptions of four new species. Zootaxa, 4032: 569-581. https://doi.org/10.11646/zootaxa.4032.5.6
- Jiménez S., McMurtry J.A., Moraes G.J. de 2014. Description of a new species of Neoparaphytoseius Chant & McMurtry (Acari: Mesostigmata: Phytoseiidae) from Peru, with a redefinition of the genus. Zootaxa, 3841: 293-300. https://doi.org/10.11646/zootaxa.3841.2.8
- Kolodochka L.A. 1998. Two new tribes and the main results of a revision of Paleartic phytoseiid mites (Parasitiformes, Phytoseiidae) with the family system concept [in Russian]. Vestnik Zoologii, 32: 51-63.
- Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina. Mem. Entomol. Soc. Can., 47: 1-64. https://doi.org/10.4039/entm9747fv
- Lofego A.C. 1998. Caracterização morfológica e distribuição geográfica das espécies de Amblyseiinae (Acari: Phytoseiidae) no Brasil. M. Sc. Dissertation, Universidade de São Paulo, Brasil, 123 pp.
- Lofego A.C., Demite P.R., Feres R.J.F. 2011. Two new species of Typhlodromips (Acari: Phytoseiidae) from Brazil. Intern. J. Acarol., 37: 110-115. https://doi.org/10.1080/01647954.2010.502905
- Moraes G.J. de, Denmark H.A. 1999. The genus Proprioseius Chant (Acari: Phytoseiidae), with descriptions of two new species from Brazil. Syst. Appl. Acarol., 4: 97-102. https://doi.org/10.11158/saa.4.1.14
- Moraes G.J. de, McMurtry J.A., Denmark H.A. 1986. A catalog of the mite family Phytoseiidae. References to taxonomy, synonymy, distribution and habitat. EMBRAPA - DDT, Brasilia, Brazil, 353 pp.
- Muma M.H. 1961. Subfamiles, genera, and species of Phytoseiidae. Flor. Stat. Mus. Bull., 5: 267-302.
- Nuvoloni F.M., Lofego A.C., Castro E.B., Feres R.J.F. 2015. Phytoseiidae (Acari: Mesostigmata) from rubber tree crops in the State of Bahia, Brazil, with description of two new species. Zootaxa, 3964: 260-274. https://doi.org/10.11646/zootaxa.3964.2.6
- Rocha M.S., Silva G.L., Silva J.O., Freitas E.M. Ferla N.J. 2015. Phytoseiid mites (Acari: Phytoseiidae) in the northern coastal region of the Rio Grande do Sul State, Brazil. Neotrop. Biodivers., 1: 22-35. https://doi.org/10.1080/23766808.2015.1052230
- Rowell H.J., Chant D.A. 1979. Observations on the ontogeny of setae in the family Phytoseiidae (Acarina: Gamasina). Can. J. Zool., 57: 670-682. https://doi.org/10.1139/z79-080
- Rowell H.J., Chant D.A., Hansell R.J.C. 1978. The determination of setal homologies and setal patterns on the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata). Can. Entomol., 110: 859-876. https://doi.org/10.4039/Ent110859-8
- Schicha E. 1983. New species, new records, and redescriptions of phytoseiid mites from Australia, Tahiti and the African region (Acari: Phytoseiidae). Intern. J. Entomol., 25 (2-3): 103-126.
- Silva E.A., Reis, P.R., Zacarias M.S., Marafeli P.P. 2010. Fitoseídeos (Acari: Phytoseiidae) associados a cafezais e fragmentos florestais vizinhos. Ciênc. Agrotec., Univ. Fed. Lavras, 34: 1146-1153. https://doi.org/10.1590/S1413-70542010000500010
- Silva L.R.A., Silva E.S., Marticorena J.L.M., Moraes G.J. de 2021. A new species of Neoparaphytoseius (Acari: Mesostigmata: Phytoseiidae) from Brazil, with a review of the genus. Zootaxa, 4985: 235-244. https://doi.org/10.11646/zootaxa.4985.2.6
- Souza R.B., Duque M.P., Amorim S.A., Cavalcante A.C.C., Guesdon I.R., Demite P.R. (2021) First report of the phytoseiid mite Neoparaphytoseius charapa Jiménez, McMurtry & Moraes, 2014 (Acari: Phytoseiidae) to Brazil, with notes on the occurrence of the genus. Entomol. Commun., 3: ec03052. https://doi.org/10.37486/2675-1305.ec03052
- Zacarias M.S., Moraes G.J. de 2001. Two new species of phytoseiid mites (Acari: Phytoseiidae) from the State of São Paulo, Brazil. Syst. Appl. Acarol., 6: 95-100. https://doi.org/10.11158/saa.6.1.12



2022-09-26
Date accepted:
2022-12-05
Date published:
2022-12-09
Edited by:
Tsolakis, Haralabos

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Ferragut, Francisco and Navia, Denise
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)