



Paedomorphosis and sexuality in Eulohmanniidae (Acari, Oribatida): surprising diversity in a relictual family of oribatid mites
Norton, Roy A.  1
and Ermilov, Sergey G.
2
1
and Ermilov, Sergey G.
2
1State University of New York, College of Environmental Science and Forestry, Syracuse, New York, USA.
2Tyumen State University, Institute of Environmental and Agricultural Biology (X-BIO), Tyumen, Russia.
2022 - Volume: 62 Issue: 4 pages: 989-1069
https://doi.org/10.24349/p0b0-usvsZooBank LSID: D92ACB1E-F31E-42DE-96BB-BF01623A5298
Original research
Keywords
Abstract
`Es handelt sich jedenfalls um eine sehr primitive Art' (Willmann 1931, p. 96).'Eulohmannia ribagai ... est une relique, la relique la plus isolée que nous connaissions' (Grandjean 1969, p. 149).
Introduction
The oribatid mite genus Eulohmannia, in the monobasic Eulohmanniidae, has been almost unmistakable since its nearly simultaneous discovery by Antonio Berlese (1910) and Ivar Trägårdh (1910; under the junior synonym Arthronothrus). An elongated, tubular form and waist-like cervical collar that is associated with a conspicuous, telescoping sejugal (protero-hysterosomatic) articulation, as well as an unusual pale orange-yellow color in mature adults, give them a facies unique among mites (Fig. 1C). Berlese (1910) proposed Eulohmannia as a subgenus of Lohmannia, with Lohmannia (Eulohmannia) ribagai Berlese, 1910 as type species. Not long after (1916b), he elevated Eulohmannia to genus rank, without discussion.
To date, nearly all distributional records have been attributed to the type species, E. ribagai (Berlese, 1910), or its supposed subspecies and synonyms. For the most part, Eulohmannia exhibits a Laurasian distribution (Hammer and Wallwork 1979): members have been reported from across temperate and boreal regions of the Nearctic and Palaearctic realms (Marshall et al. 1987; Mahunka and Mahunka-Papp 1995) and are unknown from the Southern Hemisphere (Subías 2004). Perhaps the most unusual records include those of E. ribagai from Iran (Akrami 2015) and subtropical Zhejiang Province in southeastern China (Chen et al. 2010), and of an undetermined species (`Eulohmannia sp.') from Egypt (Wafa et al. 1964; Tadros 1975).
Morphological uniqueness, some traits viewed as primitive, and a paucity of species have led to Eulohmannia being treated as an isolated evolutionary relict (Willmann 1931; Grandjean 1954a, 1969). This is reflected in its redundant but widely accepted classification as the sole genus in Eulohmanniidae Grandjean, 1931 and Eulohmannioidea Grandjean, 1931. The latter was first recognized as the superfamily Eulohmannoidea (sic) by Grandjean (1969), but since all family-group names are considered concurrent, its date is 1931.
Eulohmannia has two other special attributes. Its members typically are part of the euedaphic microarthropod fauna: they inhabit fine humus found beneath leaf litter or epigeic mosses in forests (Riha 1951; Rajski 1967; Lebrun 1971; Lions 1978; Ito 1986; Beck and Woas 1991), and both arctic (Hammer 1952) and alpine (Schweizer 1956) tundra. Having watched live individuals, Lebrun and Wauthy (1981) noted how their narrow, yet articulated body form is advantageous in small pore spaces. While there are many literature records, these mites probably are under-reported, since faunal surveys typically focus on more superficial soil layers. Second, like many other euedaphic mites, E. ribagai has been considered a'parthenogenetic'—more precisely thelytokous—species (Grandjean 1941a; Norton and Palmer 1991). Thelytoky has not been proven experimentally but virtually all adult specimens examined for gender and reported in the literature have been female; to our knowledge, there has been only one, uncertain report of a male (Grandjean 1956b). Consequently, we were surprised to encounter males in what appear to be sexual populations from northwestern North America (see below).
Our general purpose is to characterize the morphological development of these mites and to reveal some unexpected diversity. Specifically, we have five goals. (1) First, we summarize and discuss development in E. ribagai. Some ontogenetic traits have been examined in the literature, primarily by F. Grandjean (see Norton and Ermilov 2014), but many aspects have remained unknown and are the focus of new observations based on material from New York, supplemented by specimens from Alaska, Canada, Sweden, Austria, Germany, Russia (Far East) and China. (2) Other named and as-yet unnamed species of Eulohmannia are discussed, including one that appears to be sexual, and we offer a new diagnosis for the genus. (3) We propose Paedolohmannia metzi n. gen., n. sp. based on adults and juveniles from the western USA. This surprising mite does not develop the anal segment and also appears to be sexual. (4) A modified diagnosis of Eulohmanniidae follows, summarizing information from the literature and new observations herein. (5) Finally, we review the systematic relationships of Eulohmanniidae (and its monobasic superfamily) based on morphological and molecular evidence.
Material and methods
Specimens
The provenance of examined specimens is given below, under the respective species. Adults and juveniles were sorted from stored alcohol-preserved samples that derived either from Berlese-funnel extracts or from aqueous washes of humus and mineral soil following the flotation procedures of Kethley (1991). Juveniles were easily associated with adults from the same sample, since the unique facies changes little during development. In no case were adults of a second species of Eulohmanniidae present in the respective sample. Sources and depositories for specimens include the following: CNC – the Canadian National Collection of Insects, Arachnids and Nematodes, Agriculture and Agri-Food Canada, Ottawa, Ontario, Canada; RNC – the personal collection of Roy A. Norton, Syracuse, New York, USA; TSUMZ – the Tyumen State University Museum of Zoology, Tyumen, Russia; UAM – the University of Alaska Museum, Fairbanks; USNM – the National Museum of Natural History, Smithsonian Institution, Washington, DC, USA (mite collections housed with the U.S. Department of Agriculture collections in Beltsville, Maryland).
Preparation and documentation
Most observations and data are from specimens temporarily mounted in cavity slides in a medium of lactic acid diluted with water (2:1; Grandjean 1949a). Dissected mouthparts, legs and fragments of body regions requiring close study were in some instances permanently mounted in Hoyer's medium for observation and photography with oil-immersion lenses. Coarse sagittal sections were made with a hand-held razor blade fragment, on cleared specimens temporarily fixed to a slide with mounting medium. Musculature was viewed on uncleared or lightly cleared specimens temporarily mounted in glycerine; visualization was improved as needed using polarized light (Grandjean 1971).
Drawings were made with a camera lucida using a Leica DM 2500 transmission light microscope. Other observations and light photography employed bright-field, polarized, and Nomarski (DIC) illumination using a Nikon Eclipse E800 compound microscope. Light micrographs were obtained, usually as image stacks, with an AmScope MU800 digital camera. Image stacks were combined using the Helicon Focus Pro (v. 5.0) suite; the stacks varied widely in number of individual images, usually only several for highly magnified (1000 x) images and 15-30 for lower magnifications. As needed, images were adjusted with Adobe Photoshop (CS3) for contrast and color balance. For SEM microscopy alcohol preserved mites were coated with gold and scanned using a TESCAN Mira3 LMU SEM microscope.
Terminology and conventions
Morphological terminology is mostly that of F. Grandjean: see Travé and Vachon 1975 for references, Norton 1977 for leg setal nomenclature and Travé et al. (1996) or Norton and Behan-Pelletier (2009) for overview. Terms are translated from French (Hammen 1980) but Grandjean's original abbreviations and figure notations are usually retained. Paired structures are described in the singular unless noted otherwise. Throughout, there are references to numbered Remarks; each reference is parenthetic, in the form'(R1, R2, etc.).' Due to the broad, soft sejugal articulation and potential for telescoping, a fully contracted adult may have only 90-94% of its fully distended length. Because specimens exhibited the full range of distension, we standardized measurements to represent an intermediate level in which the telescoping articulation was discounted, as follows: body length was measured as the sum of notogastral (gastronotal in juveniles) and prodorsal length, with the latter measured only to the tip of the rostrum—i.e., the projecting parts of the gnathosoma were not included as they also varied in distension. Width refers to the maximum hysterosomal width in dorsal aspect. Measurements of specific structures or distances are given either as a single number meant to be representative of an average-sized individual, or an estimated range taken from a small sample of several individuals. Measured structures were oriented to prevent foreshortening errors.
Setal and solenidial formulas represent counts per segment for appendages (from leg I to IV; famulus included for tarsus I). Disjunctions (offsets) of pseudosymmetrical pairs of tarsal setae are characterized by identifying the more distal seta (Grandjean 1958b); we use the shorthand formula of Norton and Fuangarworn (2015) in which the anterior (ʹ) and posterior (ʺ) setae are indicated by respective letters a and p, with o (null) indicating no noticeable disjunction, and combinations (e.g. ao, po) indicating weak and variably absent disjunctions; x indicates that the pair is absent or incomplete on a particular tarsus. Epimeral setation is given as the number of pairs per podosomal segment (I-IV).
Common abbreviations and notations
Instars (abbreviations used primarily in telegraphic text and for parenthetic data): La – larva; Pn – protonymph; Dn – deutonymph; Tn – tritonymph; Ad – adult. Also with adjectival forms (e.g., Pnal).
Prodorsum. Setae: ro, le, in, bs, exa, exp – rostral, lamellar, interlamellar, bothridial and exobothridial (anterior, posterior) setae, respectively. Other structures: bo – bothridium; col – cervical collar; mu – mucro; pbg – postbothridial groove; PD – prodorsum; rph – rostrophragma; sej – sejugal articulation.
Notogaster, gastronotum. Setae: c-row (c1, c2, c3, cp); d-row (d1, d2); e-row (e1, e2); f2; h-row (h1, h2, h3); ps-row (ps1, ps2, ps3). Other structures: ia, im, ip – anterior, middle, posterior lyrifissures, respectively; ih, ips – lyrifissures associated with setal rows h and ps, respectively; na – plicature band between notogaster and aggenital region; ncx – narrow scissure between notogaster and coxisternum; NG – notogaster.
Coxisternum and lateral podosoma. Setae: eI – supracoxal seta; 1a, 1b, 1c, 2a, 3a, 3b, 3c, 3d, 4a, 4b, 4c – setae of epimeres I–IV (notations for epimere IV setae use the modification of Norton and Franklin 2018); Structures: ap.1, ap.2, ap.3 – apodemes 1, 2, 3, ap.sj – sejugal apodeme; ap.st – sternal apodeme; Cl – Claparède's organ; cot – cotyloid fossa of chelicera; cpc – podocephalic canal; co – cotyloid wall of rudimentary acetabulum; CX – coxisternum; g4 – gland opening above leg IV; pdc – prodorsal carina; vc – vertical contour marking anterior extent of cervical collar.
Anogenital region. Setae: ad1, ad2, ad3, ad4 – adanal setae; ag – aggenital seta; an1, an2 an3, an4 – anal setae; bpv – plicature band between adanal segment and notogaster (`border of the ventral plate'); psi – inguinal seta of row ps; x, y – questionable neotrichous setae; ψ, τ – eugenital setae of unpaired and paired lobes, respectively. Other structures: AD – adanal plate or segment; AN – anal plate or segment; ap.pa – preanal apodeme; GEN – genital plate; iad, ian – adanal, anal lyrifissure, respectively; t.pdv, t.mdv – tendon insertions for dorsoventral muscles; Vp – posterior genital papilla (`verruca').
Gnathosoma. Setiform organs: a, m – anterior, middle seta of gena; h – hypostomal seta of mentum; sup, inf, d, l, acm, cm, ul, vt, lt – palp setae; ep – postpalpal seta; ω – palp tarsal solenidion; chb – cheliceral seta. Other structures: αf – abaxial (antiaxial) fissure of rutellar manubrium; br – rutellar brush (= ciliary comb); CH – chelicera; en – line of attachment for cheliceral frame; G – gena; H – mentum; LL – lateral lip; l.or – lamellated organ; LS – labrum; op′ – adaxial oncophysis; RU – rutellum; scl – embedded sclerite of labrum.
Legs. Setiform organs: σ, φ, ω – solenidia of genu, tibia and tarsus, respectively (with numeric subscript if relevant); e – famulus of tarsus I; d, l, v – dorsal, lateral, ventral setae of whorl, respectively; bv, ev – basal trochanteral setae; a, c, ft, it, m, p, pl, pv, s, tc, u – tarsal setae; dv – vestige of seta d. Segments: Tr, Fe, Ge, Ti, Ta – leg trochanter, femur, genu, tibia, tarsus, respectively. Parentheses around leg setal notations denote the two members of a pseudosymmetrical pair on a given leg segment, rather than a true bilateral pair (unless otherwise indicated); when denoted separately, prime and double-prime (′, ″) distinguish the seta on the anterior and posterior face, respectively.
Internal. Muscles: m.adv, m.mdv, m.pdv – dorsoventral muscles of hysterosoma (anterior, middle posterior, respectively); m.pr – proterosomal retractor-adjustor muscles. Gut: cae – midgut caecum; cln – colon; fb – food bolus; fp – fecal pellet; pfb – food bolus precursor; pco – postcolon; ven – ventriculus.
Others defined in text as needed.
Systematic context
Throughout, we follow the classification and rankings of oribatid mites presented by Schatz et al. (2011) unless noted otherwise. Our view of general phylogenetic relationships is that of Norton (1998), which is largely consistent with that of Haumann (1991) and Weigmann (2006), except the latter authors do not include the Astigmata within Oribatida. We adopt Haumann's name Novoribatida to include Parhyposomata, Mixonomata, Nothrina and Brachypylina; with reference to the opisthonotal gland, these sometimes are referred to as the'glandulate' oribatid mites (setting aside the question of Astigmata). Some names are variably used in these works. Most important, Desmonomata has two very different contexts, so we avoid its use herein; we use Brachypylina as equivalent to Grandjean's Circumdehiscentiae, i.e., the'higher' oribatid mites.
Ontogeny of Eulohmannia ribagai Berlese, 1910
(Figs 1–15; all based on New York specimens unless noted)
Background
The most extensive written description of E. ribagai remains that of Trägårdh (1910; as Arthronothrus biunguiculatus) but it deals only with the adult. Other treatments are brief, vary in characters used, and also vary in whether particular characters are applied to the family, genus or species diagnoses, since the higher taxa have been long considered monobasic. Adults identified as Eulohmannia ribagai have been illustrated more than a dozen times, based on specimens from around the Holarctic. The figures (and associated text if present) suggest significant variation, especially in body size and the number and size of various body setae (see R1, R5, R12), which might have led Balogh and Mahunka (1983) to suggest the existence of Palaearctic subspecies. As explained below, some purported variation may not be real, but we suspect the name E. ribagai currently represents a species group. Accumulating molecular evidence has exposed cryptic species in other groups of oribatid mites, both sexual and thelytokous (Heethoff et al. 2007; Schäffer et al. 2019; Lienhard and Krisper 2021; Pfingstl et al. 2021).
Juveniles of E. ribagai were partly described by Grandjean (1939c), based on material from Brittany, France. This remains an essential reference, but it lacks overall figures of the body, gnathosoma and legs, and it reduces leg setation to numerical formulas. Suzuki (1979) presented a dorsal habitus image of a nymph from Japan, purportedly belonging to E. ribagai, but there are reasons to doubt the species identification (see below).
Below, we offer a narrative description of the ontogenetic development from larva to adult. The prelarva is unknown, as we have not had E. ribagai in culture and we have never found a prelarva within a female (see below).
Material examined
Our data, figures, and discussions are based primarily on samples from a population in central New York State (Cortland and Onondaga counties) that is morphologically consistent with studied Palaearctic specimens and with the carefully prepared figures of Lebrun and Wauthy (1981) and Weigmann (2006). The provenance of supplementary material is detailed below. All examined adults were female, except in samples from northwestern North America (see below). Unless indicated otherwise, specimens are in the personal collection of the first author, with a selection of each instar from Cortland Co., NY in the TSUMZ.
USA — New York: Cortland Co., Tully, Heiberg Memorial Forest, 42°46.19′N, 76°04.61′W, humus and upper soil in young hardwood forest, R.A. Norton col., 29-v-2017 (~80 Ad, 11 Tn, ~80 La); same, 2-vii-2017 (4 Ad, 2 Tn, 6 Dn, 14 Pn); same, 01-viii-2017 (8 Tn, 12 Dn, 3 Pn); same, J. Cianciolo col., 17-vi-1999 (3 La); Onondaga Co., Clark Reservation St. Park, Glacier Lake basin, south slope, 42°59.65′N, 76°05.42′W, R.A. Norton col., from deep humus on north-facing talus slope, in Acer, Betula alleghaniensis forest, 26-vi-2002 (11 Ad 1 La); same, 4-x-2002 (4 Ad); same 7-vii-2009 (5 Ad); same, 02-xi-2016 (39 Ad, 4 Tn, 1 Dn); St. Lawrence Co., Cranberry Lake Biological Station, Barber Island, 26-vii-1983, R.A. Norton col., from dense mat under Lycopodium obscurum, in beech forest (1 Ad). Alaska: Fairbanks, University of Alaska Campus, West Ridge, 14-iv-2021, R. Andrews col., from soil-litter in boreal forest (8 Ad: 6 females, 2 males, 4 undetermined, UAM).
Canada — Alberta: Kananaskis Country, Fortress Mountain, 12-vii-1983, V. Behan-Pelletier col., from litter under semi-prostrate alpine fir in alpine area (2 Ad, CNC). Newfoundland: Gross Morne National Park, Berry Hill area, near Rocky Harbour, 29-vii-1976, E. Lindquist col., from spruce-fir litter above seashore (1 Ad, CNC); 2 mi. N of Eddieʹs Cove, 11-viii-1976, E. Lindquist col. from crowberry-bilberry mat and litter above seashore (1 Ad); St. Anthony, 12-viii-1976, E. Lindquist col., from moss, grass, herbs and substrate by seepage, lighthouse area (1 Ad, CNC). New Brunswick: Kouchibouguac National Park, 19-vi-1978, R. Cope col., from den in mixed woods (1 La, CNC). Nova Scotia: Cape Breton Highlands National Park, Clyburn Brook, 7-ix-1983, V. Behan-Pelletier col., from red oak litter at base of very old tree on rocky slope (1 Ad, CNC); same, but Pleasant Bay, 7-ix-1983, from thick Fagus litter (1 Pn, 1 Tn); same but Mica Mountain, Barren, 12-ix-1983, from Arctostaphylos, Ledum, Vaccinium and Alnus litter (1 Ad). Northwest Territories: Reindeer Station, edge of Caribou Hills, 68°42′N, 134°07′W, 8-vii-1987, R.A. Norton col., from litter, roots under Spiraea and birch (Betula papyrifera) (~ 7 cm to permafrost) (10 females, 8 males; 2 Tn). Ontario: Lanark Co., Tennyson, 14-vi-1970, B. Stewart col., from moss on rock in deciduous bush (2 Ad, CNC).
Europe — Austria: Vorarlberg, Vandans, Lüner See, southern slope of Seekopf, 2030 m a.s.l., 47°03′12.2″N, 9°44′37.3″E, 30-vi-2010, I. Schatz col., from sieving alpine grassland and cushion plants (1 Tn, 1 Pn); Vorarlberg, Rheintal valley, Batschuns near Rankweil, 590 m a.s.l., 47°16′45.3″N, 9°39′51.1″E, H. Schatz col., from soil in pasture grass with roots (2 Tn); Salzburg, Badgastein, Stubner Alm, on base of Stubnerkogel, ~1815 m a.s.l., 47°07′N, 18°07E, vii-1977, H. Schatz col., from soil in cultivated pasture near timberline (1 Tn). Germany: Saxony, District of Görlitz, Ostritz, Neißetal, 1967, H.D. Engelmann col., from deciduous forest (11 Ad; see Engelmann 1972); Bremen, Teufelsmoor, 53°16ʹN, 8°54ʹS, 3-xii-1987, H. John col., from litter under ferns and scattered birch in coal mining area (1 Tn). Sweden: Torne Lappmark, Abisko, 3-vii-1975, A. Edler col., from'reindeer moss' (Cladonia lichen) on stone and clay (1 Ad); Skåne, Traneröds Bog, 30-viii-1974, A. Edler, col., substrate unknown (7 Ad).
Asia — Russia: Far East, Khabarovsk Territory, Bikin District, 9 km SSE Boitsovo (46°49′N, 134°23′E), 4-ix-1991, R.A. Norton col., from fern litter, rhizomes in moist ravine under Picea litter (1 Ad); same, Khabarovsk District; Bolshekhekhtsirsky State Nature Reserve, mixed forest, 48.275656 135.047959, 17-ix-1988, V. Behan-Pelletier col., from Sorbus and Pinus koraiensis litter (8 Ad, CNC); same, from litter under dead fallen tree (1 Ad, 1Tn); same, edible mushrooms under Betula (1 Ad). China: Beijing Administrative Zone, Men Tou Gou District, Donling Mountain, ca 1200 m a.s.l., 39°58′N, 115°26′E, 6-x-1997, R.A. Norton col. from moist litter at edge of small stream in Populus, Juglans forest, with much herbaceous litter (1 Ad).
Description of ontogeny
Dimensions — In the New York population, the various instars (n = 10 of each) had a range of total length × maximum width as follows: La 340–369 × 116–132; Pn 427–448 × 126–149; Dn 494–543 × 145–182; Tn 553–660 × 165–204; Ad (all females) 679–752 × 209–223. Most examined adults and juveniles from other studied populations were within these ranges (R1).
Facies and proportions — The elongated, almost cylindrical form of the body develops gradually (Figs 1-4), by changing proportions (proterosoma more than 3/4 length of hysterosoma in La but less than 2/3 in Ad), especially by relative elongation of the hysterosoma (slightly more than 1.5 times its maximum width in La but about twice its width in Ad). In all instars the basal region of the proterosoma comprises a ring-like cervical collar (col) that inserts into the hysterosoma when the mite is fully contracted, but the collar gradually becomes more waist-like during ontogeny. Also in all instars, the paraprocts are oriented almost vertically, such that defecation occurs posteriorly, rather than ventrally. This seems advantageous for a species inhabiting narrow pore spaces in soil.
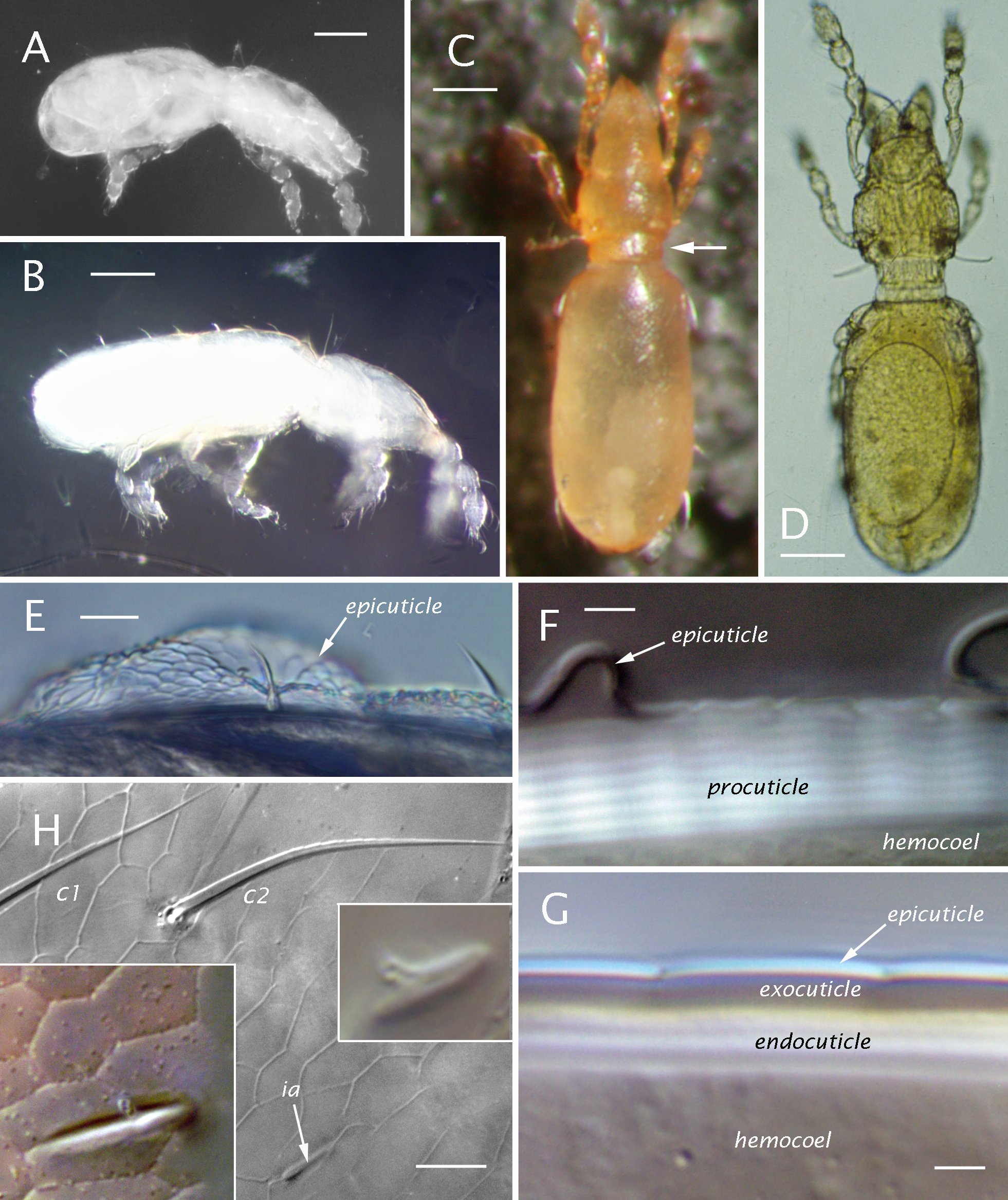
Integument (Figs 1, 2) — Juvenile instars are colorless (Fig. 1A, B); mature adults are light orangish-yellow when living (Fig. 1C), with teneral and long-preserved specimens being paler, straw-colored (Fig. 1D). Except for articulations and appendages, the epicuticle (epiostracum of Grandjean 1956b) is reticulated in all instars by tessellating, 5- or 6-sided (rarely 4) polygons that are almost flat but circumscribed by sharp, depressed lines. Mostly the polygons are rather regular in form and size (honey-combed, with width usually 10-13 um in Ad) but locally they can be smaller or more elongated (Figs 1H, 2, 7, 8B). In juveniles, but not adults, the epicuticle commonly detaches with lactic acid treatment (Fig. 1E, F), leaving the underlying procuticle without surface pattern. With light microscopy several types of inner cuticle can be distinguished. The general body cuticle of juveniles seems somewhat leathery and elastic, but to characterize it as hardened in some manner—as did Grandjean (1969;'chitinized') and Woas (2002;'sclerotized')—seems inappropriate. In polarized light or strong DIC illumination there is distinct layering and glowing in the procuticle (Fig. 1F; see R8), which is unaffected by clearing (lactic acid). In the adult, differentiated body sclerites have a relatively thin, dense layer under the epicuticle that we assume is an exocuticle (Fig. 1G), which causes adult cuticle to be more rigid than that of juveniles, though still relatively elastic, deformable. The melanization typical of adult oribatid mite exocuticle is not apparent. Beneath the adult exocuticle only several, relatively thick underlying endocuticular layers are distinctly visible in light microscopy. Electron micrographs show the thinner external layers in adult cuticle (Alberti et al. 1981, their Fig. 7A; 1997, their Fig. 1A). Pore canals are unusually dense and branched in sclerites of the adult, but ultrastructure of juvenile cuticle has not been studied. Segments of appendages (Figs 7D, 11F) have thick, dense exocuticle in all instars and no endocuticle layers noticeable in light microscopy. Most articulations, such as those of appendages and the sejugal articulation, which allows telescoping of protero- and hysterosoma, lack both the epicuticular reticulation and the distinct procuticular layering (Fig. 5E-G); by contrast, the main articulations between hysterosomal plates of the adult, which seem less supple, show the layering (Fig. 9E).

Prodorsum (Figs 2-4) — In all instars, the outline of the prodorsum is roughly ovate in dorsal view. Its outline broadens at mid-length, more noticeably in the adult than in juveniles, such that the prodorsum is widest just posterior to seta in. A distinct post-bothridial groove (pbg; Fig. 2E) dorsally delimits the cervical collar (col), which is overlapped by the hysterosoma when the mite is fully contracted. Overall, the collar integument has the general form of reticulation (Figs 2E, 5H) but on the ventral surface in the adult (not juveniles) the anterior part usually has elongated parallel ridges (Fig. 8B). The rostral tectum is well-developed in all instars, with a distinct rostrophragma (rph; Fig. 5D), but it is relatively short and effaces laterally, leaving the gnathosoma partly exposed. The rostral margin is not extended anteriorly by a distinct limb. A medial mucro (mu) is present in all instars, which can appear like a rounded lobe when seen obliquely. Seen flat, the projection forms a small equilateral triangle in the larva and a more elongated mucro—set off by a pair of shallow notches—in nymphs and adult (Fig. 2D). Otherwise, the rostral margin is nearly smooth or weakly scalloped. In the adult, a narrow, solid marginal band lacks the reticulate pattern (Fig. 2D, black arrow); on each side it meets a raised carina (pdc), which runs posteriorly, effacing below seta exp (Figs 2B, 5H).

Prodorsal setae are similar in all instars (Figs 3A, 4C). Except for the bothridial seta they are simple, smooth (or nearly so) and attenuate. Setae exp and le are smallest, in is longest, and exa and ro are intermediate in length (Table 1). Pair ro are nearly adjacent, separated by 2-3 alveolar diameters, and insert immediately behind the medial mucro (Fig. 2D). Pair le are distinctly shorter than their mutual distance in all our material. Seta exa has a consistent, unusual position anterior to and well removed from the bothridium.
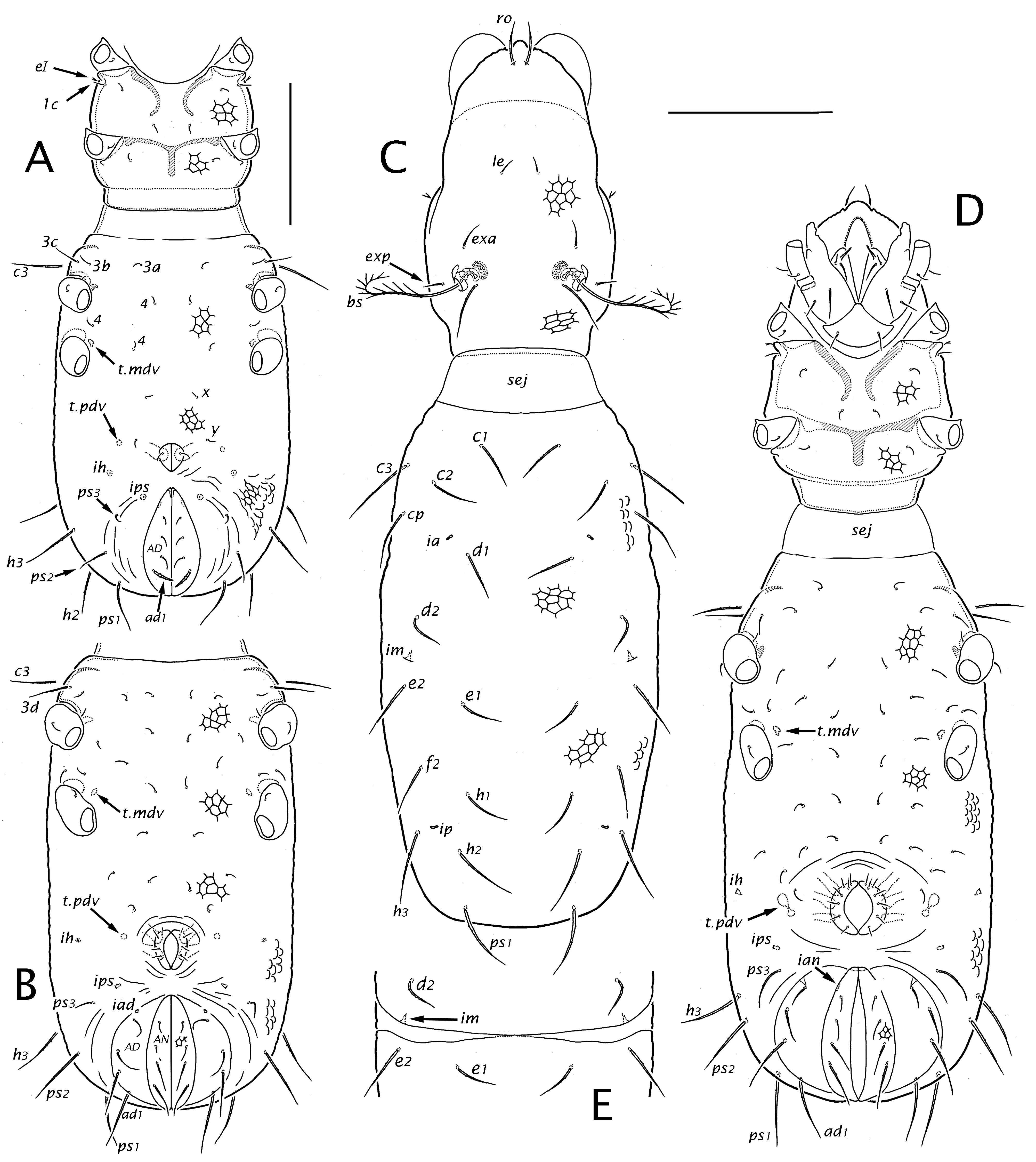
In all instars the bothridial seta (bs) arches dorsolaterally, then curves ventrad; it is thin, distally attenuate, and conspicuously pectinate on the outer curvature, with 11 to 13 long dorsal tines (alternate tines slightly divergent), and often with minute barbs in a separate row closer to the inner curvature; the basal quarter to third may appear glabrous but often has similar minute, inconspicuous barbs. The bothridium opens on a low elevation and has a slightly raised rim (Fig. 2E); there are two internalized chambers in the larva and three in nymphs and adult (Fig. 5A, C), with at least the most internal chamber having several distinct raised rings. As described in detail by Grandjean (1939b) there are two porose saccules that invaginate from the bothridial wall in the narrow, curved region between the inner chamber and the setal insertion (R2). In the larva and nymphs, the two saccules are similar (Fig. 5B), but in the adult one saccule is elongated and flattened (Fig. 5C).

Digestive system (Fig. 6) — The structure of the gastrointestinal tracts was studied only superficially, in glycerine preparations of uncleared specimens, but its general structure does not appear to vary during ontogeny. The esophagus leads to a barrel-like, thick-walled ventriculus (ven). From the posterolateral region of the ventriculus arise a pair of conspicuous, egg- to sausage-shaped midgut caeca (cae) that occupy much of the lateral region of the hysterosoma posterior to the ventriculus; like the hysterosoma in general, their proportions change during ontogeny, from about twice as long as wide in the larva to about three times in the adult. The ventriculus opens posteriorly to a tubular colon (cln) that is separated by a constriction from a similarly shaped postcolon (pco).

Hysterosomal dorsum (Figs 2-4, 7, 9) — In juveniles, the gastronotum is not clearly circumscribed from the ventral regions; the reticulated cuticular pattern continues around the circumference of the mite between the level of the legs and the paraproctal region, broken in nymphs only by the genital valves. Depending on treatment, the gastronotum of some juveniles may show a transverse crease at mid-length, but this is an artifact (Fig. 4E; R3).
A notogaster becomes defined in the adult with the appearance of two paired articulations. Using Grandjean's (1956b) terminology and notations, one is band na, a substantial, weakly curved plicature band of flexible cuticle separating the notogaster (NG) from the aggenital region (AG) of the uniform hysterosomal venter (Fig. 7H); it runs posteriorly from near the insertion of leg IV to merge with the similarly curved ventral plicature band (bpv) articulating the notogaster and adanal plates (Fig. 9A). The effect is to create an obtuse angle at the meeting of these curves such that the notogaster projects slightly into the space between genital and adanal plates from each side. This unusual form has been the source of incorrect observations and confusing terminology (R4).

The second, anterior articulation (Fig. 7C; ncx) delimits the notogaster from the coxisternum; it is a very narrow, inconspicuous lateral scissure that probably allows only slight flexing. Scissure ncx begins at the sejugal articulation in the humeral region, where it marks a change in the nature of that articulation: dorsal to ncx the notogastral margin bears a narrow but distinct tectum in adults (ngt; Fig. 5E, F)—only weakly defined on the juvenile gastronotum—that overhangs the cervical collar when the mite is fully contracted, while ventral to ncx no tectum exists (Fig. 5G). The scissure runs posteriorly, passing over the insertion of leg III where it bends ventrad, as if to pass behind leg IV; but it effaces in the vicinity of gland opening g4 (Fig. 7C), such that ncx and na do not meet. In some instances, epicuticular reticulations may align so as to give a false impression of a longer scissure (Fig. 7D), but the notogaster is consistently fused to the coxisternum in the short intervening distance. Based on all adults we examined, regardless of provenance, the statement by Grandjean (1956b) and implication by Hammen (1959), that band na continues anteriorly to the sejugal articulation, is incorrect.
There is no evidence of an opisthonotal gland or its opening (gla) in any specimen we studied. The ontogenetic appearance of lyrifissures follows the usual pattern, with ia, im, ip, ips and ih occupying the adult notogaster. Lyrifissures are slit-like in all instars (Fig. 1H); only for ih of the larva and ips of the protonymph—i.e., when these lyrifissures first appear—do they have a narrowly elliptical cupular form (Fig. 11B). Lyrifissure ia has a slightly more lateral position in the larva than in later instars (cf. Figs 3A, 4C); ip also shifts position, being posteroventral to seta f2 in the larva but dorsal to f2 in nymphs and adult. Both ih and ips make the usual small migrations seen in acariform mites with the addition of paraproctal segments (in Pn and Dn, respectively). In the adult, lyrifissure ips comes to lie close to and parallel to the notogastral margin, a short distance anterior to the angular projection, but the position of ih is more variable. Grandjean (1956b, his Fig. 1A) and Lebrun and Wauthy (1981, their Fig. 2) showed ih with an orientation similar to that of ips but slightly more anterior on the notogastral margin. In our material ih is sometimes more removed from the margin, and oriented more vertically.
In the larva gastronotic setae are minutely, asymmetrically barbed, but in later instars they are smooth, or nearly so (Fig. 7A, B); while they may become proportionally thinner during ontogeny their size relative to the body decreases somewhat, concomitant with body elongation (cf. Figs 3A, 4C). Absolute sizes are given in Table 1. The chaetome of the adult notogaster has been reported differently in the literature (R5), but in all our material, regardless of provenance, there are 15 pairs, with no indication of setal vestiges to indicate which seta is absent from a holotrichous (16 pairs) chaetome. The gastronotic chaetome of juveniles is consistent with that of the adult, i.e., no setae are added or deleted from the gastronotum other than the usual anamorphic addition of segment PS and its setae in the protonymph: so, there are 12-15-15 pairs in the larva, nymphs and adult. The larval count assumes an absence of inguinal seta hi and the notations applied to the dorsal setation assume the missing seta is f1 (R6).


Lateral podosoma and coxisternum (Figs 2-4, 7, 8) — In all instars, the prodorsum and the epimeral region of the proterosoma merge without clear separation. This is easily seen in the uninterrupted cuticle of the cylindrical cervical collar. Anterior to the collar two linear structures appear to intervene between the prodorsum and coxisternal epimeres, but neither is an articulation. The more dorsal is carina pdc, noted above (Fig. 2B, D). The other is the podocephalic canal (cpc; Figs 2B, 3C), which runs from the gnathosomal articulation posteriorly, just above trochanter II, to the groove (pbg) delimiting the cervical collar. Development of the podocephalic canal and its associated glands was illustrated and described in detail for E. ribagai by Grandjean (1939b, 1939c, 1968, 1971; see also Hammen 1982; Alberti and Coons 1999). Our observations are consistent with his, including the presence of a small separate gland opening (g4) just above the insertion of leg IV in nymphs and adult (Fig. 7C, D). Juveniles have no thickened, rib-like lateral'nervure' such as that found in some other mixonomatans (Grandjean 1966, Norton and Sidorchuk 2014). Claparède's organ of the larva (Figs 3C, D, 8A) was described in detail by Grandjean (1939c, his Fig. A): it is elongated and clavate, with a piriform head and a stalk having three or four distal annuli that seem to allow bending.
The propodosomal part of the coxisternum is a single unit, lacking distinct articulation between or within epimeres I and II. While there are no borders per se, the epimeres appear delineated more or less into four regions in transmitted light by the complement of coxisternal apodemes, which is similar in all instars. On the propodosoma, apodemes 1 (ap.1) and 2 (ap.2) are large, thin vertical lamina (Figs 3C, 4A, D; illustrated only in cross section). Pair ap.1 are strongly cupped posterolaterally; they are well separated medially in the larva, but their curved medial portions become noticeably closer in nymphs and adult. Pair ap.2 meet medially but are there deflected straight posteriad to form what can be considered a subunit: the bilayered sternal apodeme (ap.st), which ends at the strong vertical contour (vc) marking the edge of the constricted cervical collar (Fig. 8B, C). Extrinsic leg musculature that attaches to these apodemes (partly seen in Fig. 8C) does not change noticeably during ontogeny. The articulations of trochanters I, III and IV with the body are fully exposed, as in most macropyline taxa, but trochanter II appears partially recessed, protected anteriorly by a cuticular fold. This fold is indistinct in juveniles but in the adult it seems to form a rudimentary acetabulum, with the cotyloid wall (co) formed by a strong declivity posterior to ridge r2 (Fig. 8C, D; cf. Grandjean 1952b, his Fig. 1C).

Epimeres of the metapodosoma likewise have no distinct borders in any instar, and epimere IV merges seamlessly with the hypertrophied aggenital region behind it. Two pairs of apodemes exist in all instars, associated with epimere III (no apodeme is associated with epimere IV in any instar); these are relatively small, extending only slightly into the hysterosoma and therefore are inconspicuous, especially in juveniles. One, which we consider the sejugal apodeme (ap.sj) as it invaginates from the presumed anterior margin of epimere III, is posteroventrally cupped. It is most easily distinguished in the adult, at the front of the hysterosoma just below scissure ncx (Fig. 7E, F) and in dorsoventral view it tapers posteriorly to efface at the level of seta 3d (Fig. 7G). Part of the extrinsic musculature of leg III (Fig. 8C) inserts on its posteroventral face. In lateral view, fascicles of the anterior dorsoventral muscle (m.adv) may seem directed toward ap.sj (Fig. 6D), but almost certainly they insert on the centrally located endosternite (not illustrated; see Akimov and Yastrebstov 1991, their Fig. 3) from above. The second is a small apodeme 3 (ap.3) projecting internally from the body wall just anterior to the insertion of trochanter III; it is a simple vertical lamina associated with leg musculature and in all instars it is visible by transparency in dorsoventral view (Figs 3C, 7G, 8C).
In nymphs a pair of strong, conspicuous dorsoventral muscles (m.mdv) originate medial to gastronotic seta d2 (Fig. 6A) and each inserts via tendon just medial to the leg IV insertion (t.mdv; Figs 4A, B, D). The larva, with its proportionally shorter hysterosoma, lacks m.mdv but the posterior dorsoventral muscles (see below) have an analogous middle position (cf. Fig. 3C; t.pdv). The adult also lacks m.mdv (Fig. 6C), perhaps due to different force requirements associated with the presence and organization of sclerites.
Setation of the propodosomal venter is normal for oribatid mites, with most setae being smooth or weakly barbed (lengths given in Table 1); in all instars epimeres I and II have three and one pairs, respectively. Seta 1c has the usual scale-like form in the larva (Fig. 8A), covering the tip of Claparède's organ when the latter is retracted, and is a simple seta in later instars. In all instars, supracoxal seta eI is inserted posterodorsal to leg I; it is small, with an isodiametric stem and distal bifurcation of two acuminate tines. The symmetry of the tines and relative proportion of tines to stem varies within populations, but the most equal tine: stem ratios were seen in some NY specimens (Fig. 8G, bottom), while proportionally short tines were more often seen in European specimens (e.g., Fig. 8J, top; see Fujikawa 2014), and in northern Canada (Fig. 8I, bottom). Tines sometimes were broken (Fig. 8H, bottom) and in one case a tine was abnormally short and bulbous.
On the metapodosoma, the setation of epimere III also appears to have a development normal for oribatid mites: in all studied specimens, there are two pairs in the larva, three in the protonymph and four in subsequent instars (R11). The setation of epimere IV is complicated by a progressive neotrichy that appears to begin with the first formation of the epimere in the protonymph, where there are three pairs in the vicinity of legs IV (Fig. 4A, all marked'4'); this contrasts with the usual complement in oribatid mites, where the protonymph has a single pair of epimere IV setae (Grandjean 1934c). The two pairs near the genital aperture (x, y) probably do not belong to epimere IV (R11, R13). In subsequent instars, setae are added to the region of epimere IV but the absence of epimeral borders makes the number equivocal (Fig. 4B, D).
Genital-aggenital region (Figs 3, 4, 9) — Beginning with its appearance in the protonymph, the small genital aperture is positioned unusually far posteriorly, at about two-thirds the length of the hysterosoma and removed by its length, or less, from the anal aperture. Collectively, the valves have an oval or slightly obovate outline, without marginal tecta. The aperture length increases during ontogeny from less than a third that of the anal aperture (as seen in ventral view, so slightly foreshortened) to about half the anal aperture length in tritonymph and adult (cf. Fig. 4A, D). In the adult, a pair of parenthetic arms of the sclerotized aggenital region partially envelop the genital aperture but do not close behind it; the end of an arm often is partially or fully separated (usually asymmetrically) as a small island-like sclerite (Fig. 9A; Grandjean 1956b, his Fig. 1A). Behind the genital plates is a transverse band of unsclerotized cuticle that merges laterally with articulations bpv and na, and like them it shows layered procuticle in polarized light (Fig. 9A, E). Lateral to the genital valves (and in a similar position in the larva) tendons of the paired posterior dorsoventral muscles (t.pdv) insert via minute hardened projections (one per muscle fiber) at a location halfway between the midline and the lateral contour (Figs 3A, 4A, B, D). The muscles (m.pdv; Figs 6B, C, 9C) originate in the region ventral to notogastral setae f2 and h3, so probably function in controlling hemocoel pressure. There is a single pair of tendons in the larva, proto- and usually the deutonymph, but two adjacent tendons and insertion points in the tritonymph and adult. When two are present, their insertions may be separate or connected (Fig. 9B). In the adult, these lie in tandem along the margin of the aggenital plate (Fig. 9A).

Setae in this region are relatively small (Table 1), attenuate, smooth or weakly barbed. Those of the genital valves follow an ontogenetic formula (Pn to Ad) of 1-4-7-9. We encountered no variation in this development in juveniles, or in females from Nearctic or European populations, but literature reports of the adult genital setation vary significantly (R12), and we have noted variation (7-9 pairs) in studied males (see below). Setation in the aggenital region develops as part of the increasing ventral plate neotrichy, and no aggenital setae can be unequivocally identified. Allowing for the posterior displacement of the genital aperture, setae x and y of the larva and protonymph (Figs 3C, 4A) might be precocious aggenital setae (R13), but subsequently they become lost in the increasing overall neotrichy of the ventral plate (Fig. 4B, D). Collectively for epimere IV and the aggenital region, there are about 9-10 setae on each side in the deutonymph, 14-15 in the tritonymph and 16-21 in the adult. While this neotrichy has been called a'plethotrichy' (Grandjean 1956b), the setae seem mostly ordered in nearly symmetrical pairs, rather than being randomly placed or'chaotic' (see Hammen 1980).
The female ovipositor (Fig. 9E, F) is short and lacks coronal (k) setae, but the three distal lobes have a normal setation of six pairs (Grandjean 1956a; Ermilov 2011), all of which appear to be eupathidial. The unpaired ventral (posterior) lobe bears the long seta ψ1 (28-32) and the shorter ψ2 (12-15); the paired dorsal (anterior) lobes have four short pairs, τ1 (12-13) and the slightly shorter (10-11) τ2, τ3 and τ4. Based on males from apparently sexual populations in northwestern North America (Reindeer Station and Fairbanks; see below), the small spermatopositor (Fig. 9D) occupies less of the genital vestibule than does the ovipositor, and seen ventrally it is elliptical, ~ 20-25 long (relative to 65-70 for genital plates). The soft cuticle at its base is not plicate but otherwise the structure is similar in form to that of Perlohmannia (Grandjean 1958a, his Fig. 3D, E). The spermatopositor is difficult to study due to small size and obscuring internal structures, but two examples were relatively clear. There are seven pairs of setae, all small (6-8) and apparently eupathidial. A conspicuous feature is that the two pairs of short ψ setae are closely adjacent in a curving transverse row, most easily located by their alveoli (Fig. 9D). The five other pairs are longitudinally arranged, but the posterior pair often are difficult to find and seem to be at a slightly more proximal level on the structure; the latter may be a single pair of remaining k setae (absent from the ovipositor), with the other four representing τ1-τ4. As in Perlohmannia, there is a weakly defined sclerotized support near the central midline.
Paraproctal region (Figs 3, 4. 11) — Since the anal aperture is nearly terminal throughout ontogeny, the paraprocts—segments PS, AD, AN respectively in La, Pn, Dn—develop in a rather idealized form, as a series of parenthetic valves that are not distorted by the usual strong'caudal bend' of acariform mites. In each instar, there is a small, vertical preanal apodeme (ap.pa) extending from the anterior end of the paraproctal valves (Fig. 9A, E) to which muscles from the genital valves insert; these are the'anoprogenital muscles' of Grandjean (1971) or'constrictors of the genital valve' of Akimov and Yastrebstov (1991). Segment PS has four pairs of setae in the larva, but when it becomes part of the gastronotum in the protonymph there are only three pairs. We follow Grandjean's (1949b) interpretation, that the most anterior ps seta is inguinal (psi = ps4), and is lost in the protonymph. Paraproctal setae are attenuate and smooth or weakly barbed, with length in a particular row increasing posteriorly (Table 1). There are two exceptions. Seta ps1 is conspicuously thickened and barbed in the larva (Figs 3C, 11B, C), but is thin, attenuate in later instars. The setation of segment AD develops similarly—ad1 is thickened and acute in the protonymph (Figs 4A, 11C), but normal in later instars—except that it bears four pairs of setae in nymphs and adult (R14). Segment AN also has four pairs of setae from the time it forms. Lyrifissure ips is absent when the segment is paraproctal but appears in the protonymph, when the segment is incorporated into the gastronotum. Lyrifissures iad and ian also are delayed one instar, appearing in the deuto- and tritonymph, respectively. In the adult, iad and ian are inconspicuous, lying on the lateral margin of the plate close to the anterior seta (Fig. 11A); for ian often the canal is easier to see than the small slit. Grandjean (1956b; his Fig. 1A) did not illustrate ian, but it is present in all Palaearctic adults and tritonymphs that we examined.
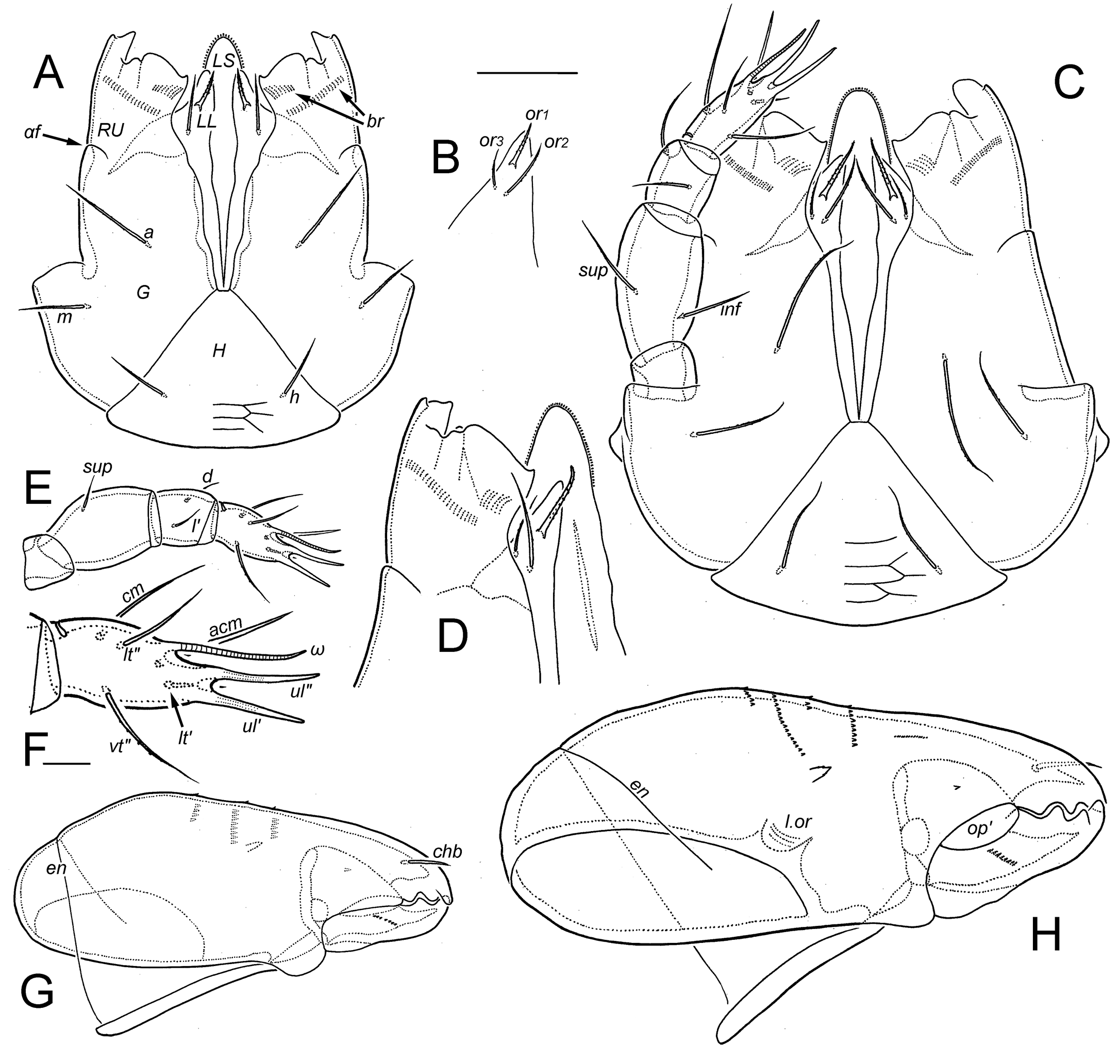
Gnathosoma (Figs 10, 11; Alberti and Coons 1999, their Fig. 92B) — Other than absolute size (see Table 2) there are few changes to the gnathosoma during development. In all instars the subcapitulum is noticeably longer (~1.3×) than wide (slightly flattened in figures) and distinctly stenarthric. The mentum (H), with transversely elongated reticulation, forms a relatively small, equilateral triangle that occupies little more than a third the subcapitular length. Each gena (G) is smooth, elongated and gradually tapering distally. The rutellum (RU)—nearly vertical in its natural position—is about half the length of the gena, from which it is clearly demarcated dorsally and laterally (manubrial line αf). The rutellum is about as long as wide and is atelobasic, leaving the adoral lips exposed in ventral view. The distal margin includes a strong, thumb-like lateral process ending in two unequal cusps, a short but strong, pigmented tooth close to its base, and a flat cutting edge occupying the medial half, with a small point at the medial end (Fig. 10A, C, D). The dorsal face of the rutellum has several fine carinae running proximally from the distal margin, and in all instars there are two oblique rutellar brushes (ciliary combs; br): a short distal brush with long cilia near the medial side, and a second at mid-length that is parallel with the first but longer and with smaller cilia. The labrum (LS) has a typical form, narrowly triangular but distally rounded in all instars, supported by a conspicuous pair of narrow, strut-like embedded sclerites (Fig. 11D; scl). The distal quarter has a row of narrow, contiguous denticles around its margin, with a second row dorsally near the tip; four inconspicuous transverse rows of narrow denticles are distributed along its dorsal face (Fig. 11E), and the ventral face has several fine transverse grooves. The lateral lips lack dorsal cilia or denticles and we observed no distinct ventral sclerite. The larva has two pairs of adoral setae: or1 usually tapers only distally and appears thickened by coarse barbs, or2 is acicular to acuminate and nearly smooth. In the protonymph or3 is added, similar to or2 but shorter, and both setae are acuminate to attenuate, with inconspicuous barbs. Setae of the hypostomal mentum (h) and gena (a, m) all are similar: thin, attenuate, smooth or with few minute barbs. The postpalpal seta (ep) is isodiametric and distally forked (Fig. 15C), generally similar to supracoxal seta eI, but depending on viewing angle the branches can be superimposed (Fig. 11F); rarely one branch is minute or even absent.

The palp has four segments in all instars: the femur and (glabrous) genu are fully fused, lacking articulating cuticle, vestigial suture, or even change of thickness (Fig. 11F) to mark their juncture. Femoral seta inf forms in the protonymph; otherwise, the setal complement is unchanging, with nymphs and the adult having the formula 0–[2+0]–2–7(+ ω). Setal forms are shown on Figs 10 (C, E, F) and 11 (F, G). On the tarsus, pair (ul) are eupathidial in all instars, terminal and distally directed, inserted almost in tandem vertically. The unpaired acm is semi-erect, a normal, acicular seta in the larva (Fig. 10F), but in nymphs and adult it is eupathidial. There is no evidence of seta su (= sul) or its vestige in any instar. Tarsal solenidion ω is relatively thin, long, ceratiform, nearly straight and directed distally, closely parallel to seta ulʺ (Fig. 11G).

In all instars the chelicera has a typical, chelate-dentate form (Fig. 10G, H). The adaxial face of the cheliceral body is emarginated in its proximal half and there is a distinct paraxial oncophysis (opʹ) and lamellated organ (l.or). The edge of l.or can extend distally, producing a line that can be misinterpreted as belonging to Trägårdh's organ, which is absent. Slightly above the center of the adaxial face there usually is a single small spine with minute teeth on its sides or distally, and above that are 3-4 vertical rows (differing in length) of small, uniform denticles; the movable digit has an oblique row of uniform, minute denticles at about mid-length on the abaxial face. The cheliceral frame attachment (line en) is somewhat distal, such that about a quarter of the chelicera is inserted through the body wall. Cheliceral seta chb inserts dorsolaterally in the distal third of the fixed digit—relatively short, acicular and nearly smooth; cha is absent in all instars.

Legs (Figs 12-15) — Legs are relatively short, with leg I (the longest) averaging 0.38-, 0.41-, and 0.44-times body length for larva, nymphs and adult, respectively. Proportional leg length changes slightly during development. Legs I-III average 1: 0.89: 0.98 in the larva; in nymphs, leg IV starts relatively small, but becomes proportionally larger, equaling leg I in the adult (legs I-IV average 1: 0.80: 0.85: 0.75 in Pn, 1: 0.80: 0.85: 0:85 in Dn, 1: 0.80: 0.85: 0:98 in Tn, 1: 0.85: 0.83: 1 in Ad). Leg form changes little during ontogeny: most notable are (a) a slight elongation of femur I, (b) development of a more pronounced proximal stalk on tarsus and tibia I, and (c) relative elongation of femur IV, which is about equal in length to the trochanter when formed in the protonymph, but about 1.3-1.4 longer in the adult. Throughout development tarsus I is swollen, compared to other tarsi. In all instars segments may have slight cuticular undulations, particularly ventrally, as well as a microsculpture of short, dense vermiform striae (Fig. 15M, N). There are no porose areas, and the so-called'genual pore' (Grandjean 1940) is absent in all instars.
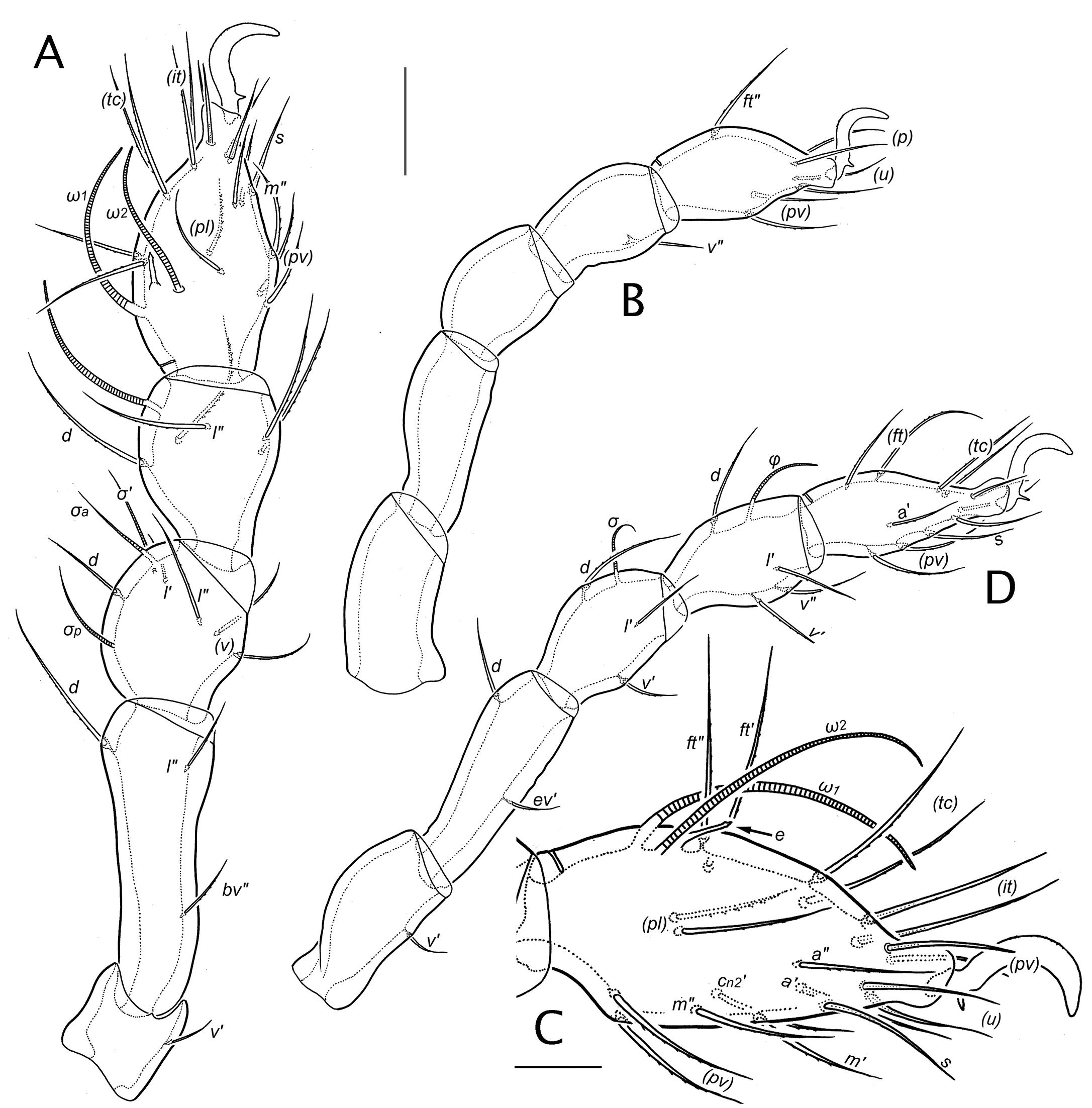
Juveniles have a monodactylous pretarsus; the empodium is a strong claw with a pair of rows of minute barbs on its dorsal curvature and a distinct, straight, ventral spine close to its base (Fig. 15A). In the adult, the empodium regresses, being represented by two minute, blunt, tandem, basally-fused spines emerging from the basilar piece (Fig. 15B; see also Alberti and Coons 1999, their Fig. 112D) that perhaps correspond to the claw and basal spine in juveniles; the pair of strong lateral claws are equal in size, with weak barbs on the dorsal curvature. There are no condylophores.
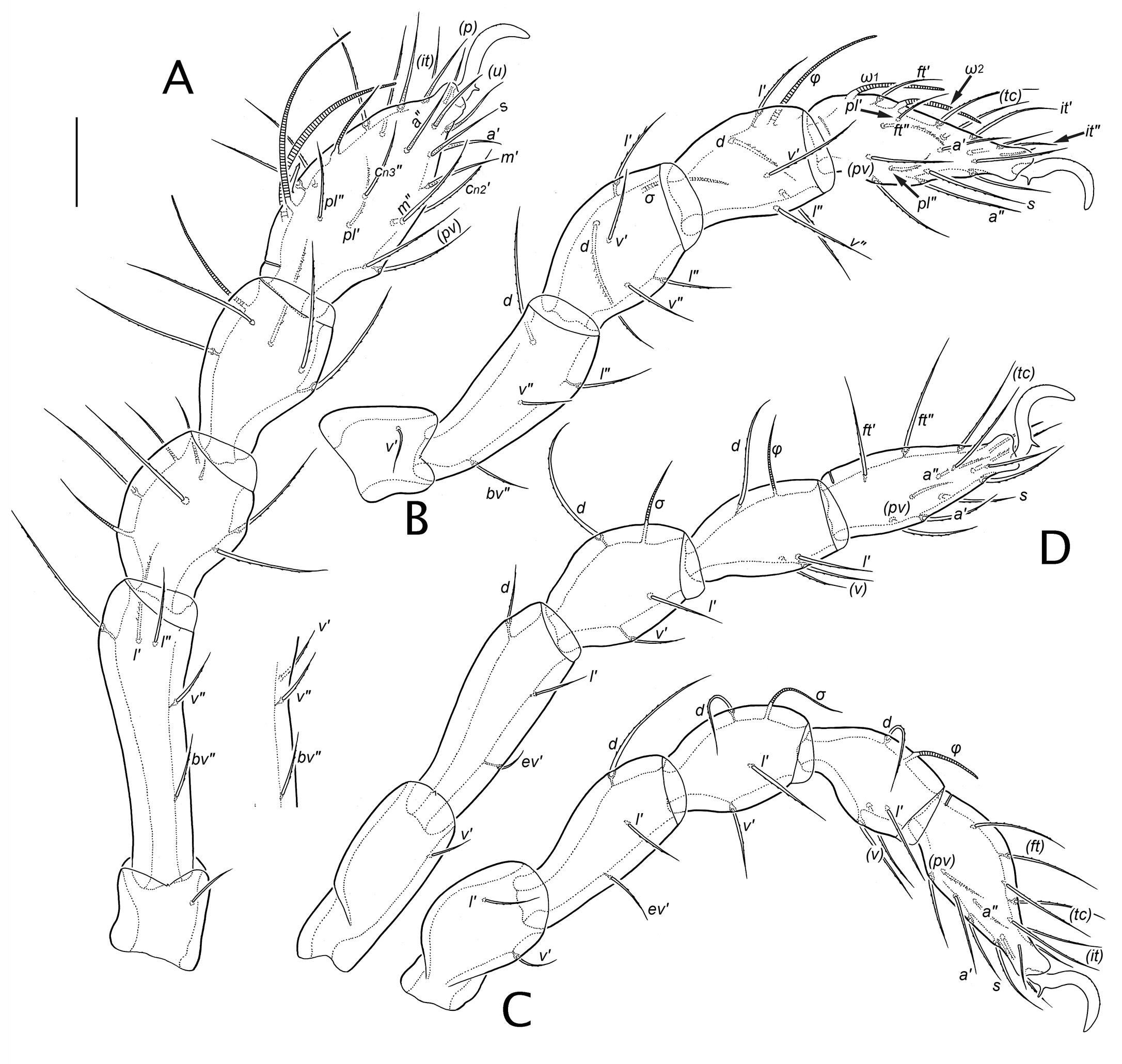
In general, normal (non-eupathidial) leg setae are relatively thin, attenuate and with small, inconspicuous barbs; on tarsus I adaxial setae of the c-row, particularly c1ʹ, usually are more distinctly and densely barbed (Fig. 15M). Numerical formulas for the chaetome of each instar are given in Table 3, and the homologies identified in Table 4. Unusual or otherwise notable features include the following. Femora. On femora I and II, lateral setae exhibit strong vertical displacement (basculation), with lʹ higher and lʺ lower (almost at level of vʺ) than the typical lateral position. Genua. On genu I–III, seta d is represented in the larva by an alveolus with a minute setal vestige (dv; Figs 12B, 15G); the seta is fully formed in all subsequent instars (R15). On genu I, seta lʹ is small and coupled with solenidion σʹ in all instars (see below). Tibiae. The verticil on tibia I has five setae—d, (l), (v)—lacking primitive seta cʺ. Seta vʺ of tibia IV is present in the protonymph (R16). Tarsi. From the larva, primitive seta mʺ is present on tarsus I, with mʹ forming in the deutonymph (R17). The iteral pair forms on tarsi I–III but develops differently on each leg (R18). From the larva, tarsus I has the usual primilateral pair (pl). On tarsus II, adaxial seta plʹ forms in the larva, but not plʺ; however, a lateral seta does form on the posterior face in the tritonymph, and we interpret this as a delayed primilateral seta (R19). Three setae of the primitive c-row form on tarsus I (R20), in successive instars. Discounting the posterior lateral seta of tarsus II, no proximal accessory setae of l- or v-rows form on any tarsus.

On tarsi, disjunctions (offsets) of the pseudosymmetrical pairs of setae are consistent across our material and also through ontogeny, whenever the particular pair is present. For tarsi I-IV, the disjunctions are: (ft) = o-p-p-p; (tc) = po-po-p-p; (it) = ao-po-p-x; (p) = o-o-p-p; (u) = o-o-p-p; (a) = ao-a-p-p; (pl) = a-x-x-x; (pv) = ao-o-p-p; (m) = a-x-x-x; (c1) = ao-x-x-x. This differs much from the simple pattern common to most Brachypylina and Nothrina, where posterior disjunction is the rule except for the primiventral pair (pv), which have anterior disjunctions (Grandjean 1958b, 1960; Wauthy and Fain 1991). It also differs from the pattern of positional analogy, as seen in the mixonomatan family Collohmanniidae, where except for pair (ft) disjunctions are mostly adaxial (Norton and Sidorchuk 2014).

Compared to most oribatid mites, tarsus I is moderately rich in eupathidia—probable taste receptors (Alberti 1998)—with a total of 10 in the adult. Transformation from normal to eupathidial form takes place at different times (Table 5; R21): the proral setae (p) are eupathidial in all instars, but most eupathidia transform one instar after they first appear [(u) and s in Pn, (it) in Dn, mʹ in Tn]. Only the fundamental antelateral pair breaks this pattern, with aʹ transforming in the deutonymph and aʺ in either the tritonymph or adult (R21). The famulus of tarsus I (Fig. 15D-F) is similar in all instars: strongly proclinate, bacilliform (~10-12 in adult), weakly curved, with a weakly formed conical head; vague annular rugosity is often visible.

The solenidial complements of tarsi and tibiae are typical of early-derivative oribatid mites, with adult formulas (legs I-IV) of 3-2-0-0 and 1-1-1-1, respectively, while that of the genu, 3-1-1-1, is exceptional (R22). The developmental aspects of these complements, shown in Table 4, was thoroughly discussed and compared by Grandjean (1964c). The shape of particular solenidia is essentially constant through ontogeny (Figs 12-14): those of genua are attenuate (piliform) while those of tibiae and tarsi (except piliform ω3 on tarsus I and φ on tibia IV) taper only slightly, or at least remain noticeably thickened distally (ceratiform); none are flagellate. There is no coupling of a solenidion with seta d on any genu or tibia: d inserts well proximal to the respective solenidion in all postlarval instars. Beginning in the larva, genu I seta lʹ is imperfectly coupled to σʹ, with adjacent but separate insertions (Figs 12B, 15G; R23).

Biological notes
Phenology — It was not our goal to study phenology in E. ribagai, but repeated sampling at the principal location in central New York (Heiberg Forest; see Material Examined) allows some tentative generalization. Each sample (May, July, August) comprised about 5 kg of fine humus and upper soil, pooled from 4-6 dispersed subsamples. Not all individuals were removed from the Berlese-funnel extracts for study, but the sorting effort was roughly equal for each collection. There was a dramatic drop-off in extracted numbers after May, but there were identifiable, pulsed changes in the relative abundance of instars which seem to indicate a phenological pattern.
The May collection was dominated by nearly equally large numbers of adults and larvae, with a few tritonymphs and no proto- or deutonymphs. Nearly all examined adults were gravid, all but two of these with a single large egg; these two females had a second, smaller egg. A small ad hoc June collection at the same site in an earlier year included three larvae, suggesting that recruitment lasts throughout the spring. In the sparse July collection no larvae were represented, suggesting that the reproductive pulse had passed; all other instars were present, but protonymphs were dominant. The equally sparse August sample contained neither adults nor larvae, and deutonymphs were the dominant instar. Collectively, we could examine only 12 adults from New York and eastern Canada that were collected from June to September (various years), and none was gravid. A relatively large November sample from central New York (Clark Reservation) comprised mostly adults, with a few older nymphs: in a subsample of 25 females, about half (13) were gravid and in several of these the eggs were relatively small, as if in the process of formation.
The overall impression is of a one-year generation time, characterized by: overwintering primarily as adults (gravid or not) and some tritonymphs; oviposition in the spring, with juvenile development during spring, summer and early autumn; and the formation (but not oviposition) of eggs in those adults that developed early enough in the fall to acquire sufficient resources. A one-year development does not always indicate univoltinism (Norton 1994 and cited references) but we have no reason to suggest otherwise for E. ribagai.
In a broad synecological study of oribatid mites in Belgium, Lebrun (1971) also concluded E. ribagai has a one-year generation time, but some other interpretations differed from ours. His data derived from two years of monthly soil samples in an oak forest, where E. ribagai was almost exclusively found in the mull (tending to moder) humus layer. His data did not discriminate the juvenile instars, nor was gravidity of adults determined. In each year, after a winter with zero collected individuals, juveniles appeared in his samples one month before adults, and he considered this to be evidence that eggs were the overwintering life stage. But since adults reached peak abundance soon after they appeared in his samples, it seems unlikely that only eggs overwintered. Perhaps adults and older nymphs overwintered in a lower soil horizon, with nymphs responding more quickly to rising temperatures than adults. After the peak spring density of juveniles, their numbers fell to zero in early- to mid-autumn.
Some of the juvenile decline seen both in Belgium and New York would be expected as they developed into adults, but in each case the autumn decline also was true of adults. Losses to predation could partly account for seasonal declines, but it is more likely that abiotic changes—soil moisture, temperature—affect movements of E ribagai into and out of the sampled humus layer. Existing information suggests that E. ribagai is especially intolerant of dry conditions (Riha 1951; Rajski 1967), so dryness in late summer and early autumn might send them below sampling depth; Rajski (1961, 1967) found especially juveniles to be abundant in deeper layers. The subsequent cold conditions of later autumn and winter might keep them there until an upward migration in spring.
The fact that in May larvae are abundant in Heiberg Forest, yet nearly all adults were gravid, suggests that adults are iteroparous, usually laying each large egg before forming another. Despite carrying a single egg at a time, they seem to be able to multiply rather quickly compared to many other oribatid mites. In a small, semi-natural rearing experiment under favorable spring conditions, Lebrun (1971) found that five adults had produced'numerous' larvae and nymphs after a single month.
Eggs — The unusually large egg (Fig. 1D; 126-179 long, 107-146 wide) has abundant resources but in no instance did we see evidence of advanced embryological development (`uterine development') within the female. This is consistent with the massive egg having to squeeze through the unusually small genital aperture at oviposition, which would seem impossible if significant development had progressed. Egg retention to the prelarval instar is widespread in macropyline oribatid mites, including many mixonomatans (Norton 1994; Norton and Sidorchuk 2014), but known examples have a relatively large genital aperture.
Apparent sexual populations — Eulohmannia ribagai has been considered a thelytokous (parthenogenetic) species since Grandjean (1941a) found that all 17 adults he studied (presumably from France, but no provenance was given) were female. Later (Grandjean 1956b) he found a single possible male—a damaged specimen—and expressed some doubt about his earlier conclusion. This seems odd since even in 1941 he had recognized the existence of rare, spanandric males in populations of some other thelytokous oribatid species. The few available cytological and genetic studies of spanandric males in thelytokous oribatid mites suggest they are nonfunctional, at least in Nothrina (Taberly 1988; Palmer and Norton 1992).
We know of no reports of male E. ribagai since 1956 and found none among studied specimens from New York, Eastern Canada, Europe, or Khabarovsk, Russia. It was therefore surprising to examine samples from two populations in northwestern North America that contained undoubted males but that otherwise were similar to other studied populations. Of 18 adults from Reindeer Station in the Northwest Territories of Canada, nearly half (8) were males, and of eight adults from the vicinity of Fairbanks, Alaska two were males. Equal frequency of sexes, allowing for sampling variation, is typical of sexual oribatid mites (Luxton 1981, Cianciolo and Norton 2006), and the male frequency seen at Reindeer Station would be a representative value for a modest sample of a sexual species. If they are indeed conspecific with thelytokous E. ribagai, these populations would raise at least two questions.
What could explain the apparent pattern? Geographic patterns in reproductive mode have not been proven for any oribatid mite. The several suggestions in the literature relate to species generally thought to be sexual, but with one or a couple reports of population samples that were female-biased (reviewed by Norton and Palmer 1991). By contrast, the pattern in E. ribagai, with thelytoky being the widespread mode, seems consistent with the classical arthropod model of'geographic parthenogenesis', with relictual sexual populations in glacial refugia (e.g., Suomalainen 1950). As part of a Beringian refugium, interior Alaska (Fairbanks population) was unglaciated during the Quaternary (Matthews 1974; Shafer et al. 2010), though the glacial history of the Mackenzie Delta (Reindeer Station population) was complex (MacKay 1974). More fine-grained sampling and genetic studies of northwestern populations should be illuminating, particularly since other sexual eulohmanniid species are now known to exist (R29).
Feeding biology — Eulohmannia ribagai is a particle-feeding oribatid mite that is essentially restricted to the humus layer under leaf litter or mosses, but there is little helpful information in the literature concerning its diet. Surveys of oribatid mite feeding modes (e.g., Schuster 1956; Luxton 1972; Siepel and de Ruiter-Dijkman 1993) do not mention E. ribagai and Lebrun (1971) seemed equivocal when characterizing its food. He listed it as a non-specialized feeder (`panphytophage' of Luxton 1972) in his summary Table 13; elsewhere (p. 83) he noted that its'alimentary regime' was not known with certainty; and finally (p. 147) he assumed it was a'microphytophage' (mycophage or bacteriophage) like other humicolous species. The most excentric proposal was by Schweizer (1957) who, based on some features of the gnathosoma and an incorrect assessment that the anal aperture is small, speculated that E. ribagai is exclusively carnivorous.
Within humus, the main food resources might be: (a) the substrate itself i.e., finely comminuted litter fragments (presumably already processed by other soil biota) and any associated bacterial films; (b) plant roots (or moss rhizoids); and (c) fungal hyphae and spores. Our ad hoc examination of gut contents showed that food boli and fecal pellets are relatively uniform, regardless of collection date, provenance, or instar (Fig. 16A, B, E). They include mostly brown fragments of plant structural material, with discrete structures—e.g., fungal hyphae and possible bryophyte spores (Fig. 16C, D)—being rare enough to be considered accidental, or at least not a focus of feeding.
The chelicerae of E. ribagai seem more robust than would be necessary for ingesting pre-comminuted fragments. Kaneko (1988), who used the width: length ratio of the movable digit to distinguish feeding modes, found fragment feeders and microphytophages to have ratios of 60% or less. That of E. ribagai is over 70%, well in the range of species that need powerful chelicerae to feed on more intact higher plant debris (`macrophytophages') or are opportunistic, unspecialized feeders. The strong chelicerae seem appropriate for removing tissue fragments from living or dead roots, but it is unlikely that E. ribagai is a root specialist. The adults placed in a small, semi-natural rearing chamber by Lebrun (1971) rapidly produced offspring yet presumably the mites had no access to roots (he did not describe the precise food materials provided).
Despite their strong chelicerae, we suspect that the fragments forming the bulk of gut contents in E. ribagai are pre-comminuted, litter-derived particles, and that associated bacterial films provide the principal nutritional resource. This would explain the necessity of high environmental moisture (Riha 1951; Rajski 1967) in a mite with a well-developed epicuticle (see above). It would also help explain the rapid reproduction observed by Lebrun, since such a diet would have higher nutritional value than that of typical litter- or wood-feeding oribatid mite species with extended generation times (Norton 1994, 2007). On the other hand, the range of habitats from which E. ribagai has been collected is quite broad and includes moor and coniferous forest soils that must have more acid conditions, which would not be amenable to high bacterial densities. The application of modern tools, such as fatty-acid analysis and stable isotope fingerprinting (Pollierer and Scheu 2021), to a humus-specialist like E. ribagai in various habitats, would be illuminating.

Based only on preliminary observations of dead specimens, the formation of food boli from ingested particles seems unusual. The colon and postcolon often contain a normal-appearing food bolus and fecal pellet, respectively (Fig. 16D-F); these compact, pigmented structures are visible by transparency even in uncleared specimens. The food accumulates in the ventriculus (Fig. 16A), but the development of a bolus seems unique. In uncleared or slightly cleared specimens, a ventriculus that entirely lacks discernable food particles can contain a single large, well-circumscribed, featureless globule that appears composed of a thick, gelatinous material (as in Fig. 22G). We do not believe this material is ingested (e.g., as a biofilm), since: (a) in the approximately dozen times we have seen it, the globule did not vary noticeably in size; and (b) a mite with unspecialized mouthparts, like E. ribagai, would not be able to scrape a surface without also obtaining particulate matter. We hypothesize that this globule is internally secreted and functions as a food bolus precursor (pfb) since it seems to accumulate food particles within its volume. In one case, in which an unusually small bolus reached the anterior end of the colon, some of the gelatinous material remained (Fig. 16C), but usually it was not discernable beyond the ventriculus. If true, this mechanism of bolus formation differs from that described for Ceratozetes by Woodring and Cook (1962; see also Alberti and Coons 1999), in which a bolus becomes well-formed only after it enters the colon and receives a peritrophic membrane. We do not know if E. ribagai also forms such a membrane.
Parasites — Recording parasitism was not a goal, and our observation methods were inappropriate for this, but the typical parasites of oribatid mites—gregarines, microsporidians, helicosporidians (Purrini and Bukva 1984)—often can be detected despite moderate clearing of specimens. We found no gut gregarines but did find one adult mite with a high density of what we interpret as spores of microsporidians (Fig. 16F). We could not precisely identify the host tissue, due to clearing, but probably it was a nearby caecum: these sporozoans are most often associated with the gut wall (including caeca), sometimes the gonads (Purrini and Weiser 1981).
Nomenclatural issues
Two other names have been considered synonyms of E. ribagai or wrongly assigned to Eulohmannia; these are discussed below.
Arthronothrus biunguiculatus Trägårdh, 1910 — Berlese's (1910) brief description of Eulohmannia ribagai, based on specimens from moss in Tiarno, Italy (see Castagnoli and Pegazzano 1985), was published on 9 February 1910. Trägårdh's much more extensive treatment of the same species, based on material from a (presumed underground) bumblebee nest in Säkok, Sweden, was published on 30 June 1910. Both the genus and species names are synonyms, as first noted by Berlese (1916a), with Berlese's names having precedence.
Iburidania bipectinata — In an early treatment of the Japanese oribatid mite fauna, Aoki (1959) introduced the names Iburidania bipectinata and the family Iburidaniidae but attributed them to K. Kishida, who did not present them in the Latin alphabet. Aoki clearly considered Iburidaniidae to be a junior synonym of Eulohmanniidae, and Iburidania a junior synonym of Eulohmannia. Fujikawa (1991) listed'Iburidania Kishida in Aoki, 1959' as a nomen nudum and in a later synopsis (Fujikawa et al. 1993) listed'Iburidania bipectinata Kishida in Aoki, 1959' as a junior synonym of Eulohmannia ribagai; this latter concept was accepted in a review of Berlese's oribatid mites by Mahunka and Mahunka-Papp (1995). Ohkubo (2002) carefully analyzed the identity of Kishida's mite and instead considered it a species of Epilohmannoides (Epilohmanniidae) not Eulohmannia. Further, he considered the names Iburidania and Iburidania bipectinata to be unavailable under the International Code of Zoological Nomenclature (ICZN), since Kishida himself had provided no Latinized binomen and did not have the intention of proposing new taxa. Ohkubo also believed the names could not be attributed to Aoki (1959), since Kishida's descriptive information was not in a formal publication. However, even if these points could be interpreted differently the names still are unavailable, since Aoki initially treated Iburidania and Iburidaniidae as junior synonyms of preexisting valid names: according to Article 11.6 of the ICZN, a name cannot be first proposed as a junior synonym. Since they are unavailable, the species, genus and—by extension—family names should not be listed in synonymy (e.g., see Subías 2004, 2022).
Other species and diagnosis of Eulohmannia
In a footnote, Balogh and Mahunka (1983) indicated that'Palaearctic forms' considered to represent E. ribagai might eventually be found to represent subspecies. They offered no explanation, but if one considers the published illustrations and descriptions purported to represent E. ribagai, such a comment is not surprising. The available information varies notably in characters such as body size (see R1) and the number of setae on the notogaster (R5), coxisternum (R11) and genital plates (R12). Representation of the pectinate bothridial seta (sensillus) is particularly variable in the literature. As examples, the number of tines is reported as seven by Schweizer (1956), eight by Bulanova-Zachvatkina (1960); 10 by Berlese (1910); 12-13 by Trägårdh (1910) and Hammer (1952); 9-15 by Aoki (1975), 13-15 by Balogh (1943), and 16-19 by Willmann (1931). Also, the size of the tines varies in illustrations, though much of the variation might be accounted for by viewing angle; e.g., in Berlese's (1910) original figure both the strong curvature of bo and the length of its tines are disguised by projection in the dorsal aspect. Whether any of these quantitative variations represent different species-group taxa cannot be determined without more thorough study that also includes molecular data, but Eulohmannia certainly is not monobasic. Below we discuss other species known to us, as well as species wrongly attributed to Eulohmannia.
Eulohmannia bifurcata Fujikawa, 2014, new status

Although it was proposed as a subspecies (Eulohmannia ribagai bifurcata Fujikawa, 2014), we believe there are sufficient reasons to distinguish this mite from E. ribagai and elevate it to species rank. It is known only from type specimens, which were derived from two separate collections in the east-central region of Honshu Island, Japan. One of us (R.A.N.) borrowed the five specimens—all female and all slide-mounted—from the National Museum of Natural History (Tokyo). The holotype (13642; Fig. 16G) from Tochigi Prefecture is strongly crushed (probably intentionally) and used as the source for Fujikawa's Figs. 3-5; the captions to these figures wrongly refer to it as a paratype. The medium (unidentified) is deteriorating somewhat, and fibers are embedded, one of which transects the specimen. The four paratypes from Fukushima Prefecture are whole-mounted, apparently in a resin-based medium; there are two each on slides 13643 and 13644, mixed in a slurry with about 20 specimens of other mite species. Below, we use arbitrary designations (A, B) for the paratypes on each slide: 13643A (Fig. 17A) is a ventrolateral mount, not illustrated by Fujikawa but exhibiting some features that are difficult to see on other specimens; 13643B is a dorsal mount shown in her Fig. 2A; 13644A (Fig. 17E) is a ventral mount shown in her Fig. 2B; 13644B is a dorsal mount, not illustrated. They are not well cleared. Her measurements (length 571-629) suggest that E. bifurcata is slightly smaller than E. ribagai, but the paratypes are contracted; using our method (summing protero- and hysterosoma), they are 630-670.
Differences with E. ribagai — Fujikawa identified traits that seemed to clearly distinguish E. bifurcata from E. ribagai; some were highly relevant to classification and phylogenetic inference, but they were not discussed in this light. The most surprising of these—the purported presence of supracoxal seta eII—could have been perceived as an atavistic return of an ancient trait. Several others—including opisthosomal glands and chelicerae having Trägårdh's organ and seta cha—would have been unique plesiomorphies within the family. As discussed below, we consider these traits and several other to reflect errors in observation or interpretation. However, differences in six traits seem sufficient to recognize this mite as a distinct species. States in E. ribagai (given in parentheses) are based on our studied material from all provenances. Leg setation does not differ from that of adult E. ribagai, including the distribution of eupathidia (see Tables 3-5).
- Epimere III has five pairs of setae (vs. four pairs). This is consistent in all five type specimens. Setae 3a, 3b, 3c, and 3d are similar and rather short, but the most lateral (3e) is long and conspicuous (resembling 3d of E. ribagai). Fujikawa seems to have confused 3e with the similarly-sized notogastral seta c3. She omitted 3e from the ventral figure (her Fig. 2D) and in the dorsal figure (her Fig. 2A) 3e is mistakenly labeled'c3' on the left side, where the actual c3 is not drawn—it curves artificially toward the sejugal articulation and is made inconspicuous by following the notogastral margin in dorsal view. On the neotrichous hysterosomal venter, the setation of epimere IV is inseparable from that of the aggenital region, as in E. ribagai.
- A small vertical ridge (r1) posterior to leg I, distinct in both lateral (Figs 16H, 17D) and dorsoventral views (Fig. 17F), extends ventrad from supracoxal seta eI to the leg insertion (vs. ridge absent; cf. our Fig. 8C).
- The posterior arms of the aggenital sclerite encompassing the genital aperture (Fig. 18A) are broadly rounded posteriorly (vs. distinctly tapered posteriorly; our Fig. 9A; Grandjean 1956b, his Fig. 1A). Fujikawa illustrated this region in three figures, but none is entirely accurate. Her Fig. 2B shows the arms merging behind the genital aperture of a paratype, but they do not; details in this region easily can be misinterpreted due to the state of contraction and cuticular folding. In her Fig. 4C the adanal and anal plates of the holotype are accurately drawn—what she considers the'anterior anal locking-piece' is the preanal apodeme—but the aggenital region is wrongly depicted. Her Fig. 4D shows an intact aggenital region in the holotype; the broad arms are drawn correctly but the small, separate sclerites are omitted, and the lines behind the aperture are not interpretable. This latter figure must have been made before the holotype was strongly crushed, since the region is now split longitudinally. The holotype is dorsally mounted, but with DIC illumination the small'island-like' sclerite posterior to each broad arm is visible by transparency (Fig. 18A).
- Femur IV is about 1.5-1.7 times the length of trochanter IV (vs. 1.3-1.4 times).
- Seta acm of the palp tarsus is a normal seta in the adult (vs. eupathidial; cf. Figs 16M, 11G). Since acm usually transforms to a eupathidium early in ontogeny (the protonymph in E. ribagai) we consider this a neotenic trait. However, the palp could be studied closely only on the holotype, so other examples are needed to confirm this difference.
- Distal leg segments (genu-tarsus) are noticeably sculptured (vs. smooth or with only weak undulations). Fujikawa (her Fig. 5A) identified a small boss distal to tarsus I seta tcʹ that does not exist in E. ribagai, but other sculpturing is more extensive and conspicuous. The ventrodistal cuticle of genu and tibia of legs I and II is strongly undulating (Fig. 18B, C) and the distal three segments of all legs have an oblique declivity on the adaxial face (Fig. 18D, arrows) that emphasizes the basal stalk and gives the impression of a rudimentary, weakly-defined retrotectum.

Additionally, setae on the dorsum of E. bifurcata, particularly prodorsal seta le and notogastral setae, are relatively longer than in most E. ribagai. In E. bifurcata, le is slightly longer than the mutual distance of the paired alveoli, whereas in our studied material of E. ribagai they are distinctly shorter than their mutual distance, often only half that length. However, some literature figures of E. ribagai show le equal to that distance (e.g., Balogh 1943; Kunst 1971). The notogastral setae of E. bifurcata are relatively long, reaching the insertion of nearby setae, or nearly so. In E. ribagai they are generally shorter, typically reaching only about halfway to nearby setal insertions (e.g., Lebrun and Wauthy 1980; Weigmann 2006), but in specimens from New York (Fig. 2C) and Ukraine (Sergienko 1994) they are intermediate in length.
Equivocal or incorrect distinctions — Fujikawa's Table 1 includes seven traits that she believed distinguished the two taxa. Two of these are variable traits of limited value. She characterized the genital setation of E. bifurcata as 7-8 pairs and that of E. ribagai as 8-10. However, the latter species can have as few as seven (R12). While not an absolute difference, E. ribagai usually has nine pairs, compared to eight on those genital plates of E. bifurcata that are most clearly visible. She also characterized supracoxal seta eI as having minute distal tines in E. ribagai, but in some populations they are as long as in E. bifurcata (Fig. 8; R9). A third trait, epimeral setation, was incorrectly characterized for both species, though they do in fact differ (see above, #1). The other four purported differences result, in our opinion, from mistakes in observation or interpretation, as explained below. In each case the relevant specimens were examined at magnifications up to 1500×, under all available types of illumination.
Notogastral setae. Fujikawa indicated that E. ribagai has 16 pairs of notogastral setae, while E. bifurcata could have either 15 or 16. We view both statements as incorrect. There are consistently 15 pairs in adult E. ribagai (see above, and R5). Her suggestion of variation in E. bifurcata relates to the presence or absence of a seta labeled'h3' in her Fig. 2D, which appears to be inserted near her'h2' (our h3), lateral to the lyrifissure that she incorrectly labeled'ih' (see below). She indicated in the text (p. 8) that'h3' was absent from only one side of one specimen, though it is absent from one side of both her Figs. 2A (13643B) and 2D (13644A). We could closely examine the two illustrated regions where'h3' was shown, and each specimen had only one seta in this vicinity: her'h2' (our h3; cf. Fig. 17 G, H). The same is true of paratype 13644B (not illustrated). Paratype 13643A could be studied only on one side, but it clearly shows the presence of a single h-seta in this region (Fig. 17B). This region of the crushed holotype could be studied closely on each side, and there is a single seta. It is unequivocal that the notogastral setation of E. bifurcata comprises 15 pairs, identical to that of E. ribagai.
Opisthonotal glands. Fujikawa's Fig. 2A shows a small circle closely posterior to notogastral seta'h1' (our h2), which she labeled gla, the opisthonotal gland opening. This would be a unique position for gla, well posterior of its usual position somewhere near seta f2. Using all available lighting at 1500× we examined the dorsally mounted paratype (13643B) from which her Fig. 2A was drawn and found no gland aperture. On the left side of the paratype it seems clear that she drew a small bead-like structure (in our Fig. 17H); this may be some artifact of the mounting process or preservation, but it is subsurface and certainly not gla. We could not identify a structure that would have suggested a gland opening on the right side of 13643B, or on either side of the other dorsally mounted paratype (13644B).
Trägårdh's organ. Fujikawa (her Fig. 4E, drawn inverted) illustrated a well-developed Trägårdh's organ on a chelicera of the holotype, but it appears on the abaxial (= antiaxial) face, as shown by the presence of seta chb; this is an impossible position for Trägårdh's organ, which extends from the body wall on the inner (adaxial, paraxial) face of the chelicera (e.g., Hammen 1968). Both chelicerae of the crushed holotype have their abaxial faces up (Fig. 16J), and the adaxial face is visible by transparency; we see no structure on either side of either chelicera that could be identified as Trägårdh's organ. No views were possible on the intact paratypes.
Supracoxal seta eII. Fujikawa indicated that E. bifurcata differed in having three pairs of supracoxal setae (e, eI, eII) compared to only eI in E. ribagai. Clearly, she was mistaken about'e', the postpalpal seta (ep), since it always exists in E. ribagai (e.g., our Fig. 3D). Seta eII indeed does not exist in E. ribagai, but she seemed unaware of the rarity of this metameric homologue of eI. A few taxa of Prostigmata, including Anystidae, Bdellidae and some Parasitengona, as well as Opilioacarida (Hammen 1980; Kethley 1990), possess eII and it can be considered plesiomorphic in those groups. But to our knowledge it is not known to occur in any endeostigmatid or sarcoptiform mite (Grandjean 1939a, 1954a).
Fujikawa included eII in two figures but showed it at two different locations. Her Fig. 2D (ventral view, paratype 13644A) shows the seta bilaterally, on the projecting posterolateral corner of epimere II and distinctly posterior to trochanter II. But when the surface is in focus clearly there is no seta here, on either side (Fig. 17F). We believe she misinterpreted a gland duct as a setal alveolus: the duct of the posterior podocephalic gland (Grandjean 1939b, his Fig. 2; gl.m) is round in cross section (d.gp; Fig. 17F insert) and is found in that exact location. The other figure is her Fig. 4A, which is an inverted lateral view (ventral at top) showing details of the podocephalic region on the left side of the crushed holotype (13642). The cuticle in the relevant posterior region of the canal is broken, but her illustration of this region—which seems unchanged since 2014—wrongly suggests that the cracks are internal structures (cf. Fig. 16H). In this figure, she drew eII dorsal to trochanter II—between it and the podocephalic canal (cpc)—but we found no seta there, only a minute, pale mark of uncertain nature (* in our Fig. 16H). Paratype 13643A provides a clear and intact view of this region: seta eI and all three gland openings (cf. Hammen 1982, his Fig. 15D) are clearly visible, but there is no seta eII, or any other structure at its purported location.
Other notable errors — Four other traits described for E. bifurcata differ conspicuously from those of E. ribagai, but they were not included in her differentiation table.
Notogastral lyrifissures. According to Fujikawa's illustrations and interpretations, E. bifurcata would clearly differ from E. ribagai in the distribution of three typical notogastral lyrifissures: ip, ih, and ips. In adult E. ribagai (Grandjean 1956b; Lebrun and Wauthy 1981) —as well as other species discussed below— the lyrifissures have consistent locations similar to those in many other oribatid mites. Lyrifissure ia is posterior to seta c2; im is posterior to d2; ip is either posteromedial or lateroventral to f2; and both ih and ips lie near the notogastral margin, visible only in lateral or ventral aspect. Fujikawa (2014) did not include lyrifissures in her illustrations of E. ribagai, but five were indicated for E. bifurcata (her Fig. 2A, C). She showed ia and im in their usual places and a third pair posteromedial to f2 — the latter is labeled ih but clearly is ip, which has been associated with segment F (Grandjean 1939d). Therefore, the anterior three pairs are distributed exactly as in E. ribagai. Of the remaining two, one pair purportedly lies on the posterior contour of the notogaster in dorsal view (her Fig. 2A, wrongly labeled ip), medial to setal pair ps1. We studied this region of paratype 13643B, from which the figure was made, and found no lyrifissure there: on the left side there is a small linear artifact (** in Fig. 17H); on the right side the notogaster is damaged (cracks not shown in her figure) and nothing resembling a lyrifissure could be found. The remaining lyrifissure purportedly lies between setae'h2' (our h3) and ps3 and is labeled ips in her Fig. 2B. Her figure was drawn from paratype 13644A, but we found no lyrifissure on either side at that location (cf. Fig. 17G).
Since both ih and ips are theoretically located at the end of their respective setal row (Grandjean 1939d), the positions she ascribed to either of these lyrifissures would be incongruent. In fact, ih and ips of E. bifurcata have the same locations as in E. ribagai, lying in tandem along the border of the notogaster, anterior to seta ps3. They can be seen in paratype 13643A, which shows this region clearly (Fig. 17C) and also in the crushed holotype (Fig. 18A).
Cheliceral setae cha. Fujikawa did not discuss the setation of the chelicera in E. bifurcata, but her Fig. 4E of the holotype shows two setae. Seta chb inserts distally on the abaxial face of the fixed cheliceral digit, as in all instars of E. ribagai (e.g., Fig. 10H). Seta cha, which does not exist in any instar of E. ribagai, is drawn as a small, barbed seta inserted low on the abaxial face, such that the axis of rotation for the movable digit would precisely pass through the purported setal alveolus. This would be a unique location: seta cha exists in most oribatid mites, but it is inserted either on the dorsal midline or high on the adaxial face. We could not find cha on either chelicera of the holotype and believe that Fujikawa misinterpreted the cotyloid fossa of the movable digit (cot, Fig 16J; see Grandjean 1947, his Fig. 2B) as a setal alveolus. We believe that she misinterpreted the edge of the thin articulating cuticle (Fig. 16J, insert) as a seta, but we cannot explain the illustration of fine barbs in her Fig. 4E.
Rutellar brush. The rutellum of E. ribagai bears two independent brushes on its dorsal face, one shorter than the other and differently oriented (e.g., Fig. 10D). In her Fig. 3E, taken from the holotype, Fujikawa illustrated the rutellum of E. bifurcata as having a single brush, but there are two (Fig. 16K), formed as in E. ribagai.
Empodial vestige. Fujikawa described the pretarsus of E. bifurcata as'homobidactyl' and illustrated this for tarsus I of the holotype (her Fig. 5A), where no vestige of an empodial claw was shown. However, such a vestige does exist on all legs that are oriented for favorable viewing (e.g., Fig. 16I, L). It has the same tandem, bidentate form as in E. ribagai (cf. 15B).
Another possible species from Japan?
Suzuki (1979) studied a protonymph from Japan that he thought represented E. ribagai but that differs markedly from the nymphs studied and described by us. His Fig. 2 shows two large, distinct sclerites on the gastronotum, and these, as well as the prodorsum, are punctate or have small, well-spaced foveolation, rather than the general reticulation found in all instars of E. ribagai. Otherwise, the facies are like those of E. ribagai nymphs, and the rostrum has a similar mucro and adjacent notches.
Suzuki referred to Aoki's (1975) drawing of E. ribagai to represent the adult. However, Aoki (his Figs 1, 2) illustrated a mite with lamellar setae and setae of the neotrichous ventral plate that are significantly longer than in all E. ribagai studied by us. Aoki clearly illustrated a complete encompassing of the genital aperture by the aggenital region of the ventral plate; this does not occur in E. ribagai though some published illustrations have been wrong or vague about this detail. No cuticular reticulation was shown or described by Aoki, but this pattern can be indistinct depending on preparation and observation methods—e.g., it has been omitted from several figures of E. ribagai (e.g., Kunst 1971; Weigmann 2006). Enough discrepancy exists to warrant reexamining Japanese mites ascribed to E. ribagai and to investigate their juveniles.
Two undescribed species from Asia
A large sexual species from Russia — We believe a population referred to herein as Eulohmannia. sp. A (Fig. 18E), from Aborigen, represents an undescribed species closely related to E. ribagai. Examined material is from the subarctic Kolyma Highlands of Magadan Oblast, Russian Far East. Like populations of E. ribagai from the Pacific Northwest, this species almost certainly is sexual, and it inhabits an area with a complex glacial history (Bar and Clark 2012). We studied a series of prepared slides from the CNC, with the following data: Aborigen, southern parts of Bolshoi Annachag Mountain, vii-1979, A. Fjellberg col. (4 males, 3 females); same, along Aufeis River, 25-vii-1979, A. Fjellberg col. (1 female); Aborigen Alpine Study Area, Saddle site, 23-vii-1979, V. Behan col. (5 females, 4 males, 1 La, 1 Pn, 2 Dn, 2 Tn); same, Aborigen Transect T1S4, 25-vii-1979, V. Behan col. (1 female). While having the same body facies as E. ribagai, and a similar pretarsal empodium, these mites differ in significant ways. First, with an average total length of 800 µm (n = 19) adults are distinctly larger (R1). The range (718-854) overlaps the values for E. ribagai slightly, but the smallest of the Aborigen mites are male (718-776). Leg IV has a proportionally smaller trochanter (femur ~2× length of trochanter, vs 1.3-1.4 in E. ribagai; cf. Figs 2A, 18E), trochanter III bears three setae (vs. two), and the setation of tarsus I differs in several ways, particularly with regard to setae of the c-row and eupathidia (R20, R21). With the two larger samples being nearly half males (Fig. 18G), the mite is probably sexual. By contrast, our Eulohmannia material from Khabarovsk (also Russian Far East, but in the temperate south) is typical of E. ribagai in all traits, including size (650-756 µm, n = 9), leg traits, and the absence of males.
A paedomorphic species from Kashmir — We have studied four adults from a population in the Kashmir Valley (the Indian-administered union territory of Jammu and Kashmir), referred to herein as Eulohmannia sp. B, that certainly represent an undescribed species. Collection data are: Kashmir, Dachigam National Forest (near Srinagar), 20-viii-1986, R.A. Norton col., from litter at base of elm tree (Ulmus wallachiana) in mixed forest. The body (Fig. 17D; ~ 680 long) appears almost indistinguishable from that of adult E. ribagai, except for lacking the notches lateral to the medial rostral mucro. But all specimens have monodactylous leg pretarsi. The fully developed empodial claw has a strong spine-like process (Fig. 18I), like that of E. ribagai juveniles (cf. Fig. 15A). Leg setation has some juvenile attributes also (R20, R21) but there is no doubt these are adults as the ovipositor is distinct in each, and one has a fully developed egg (Fig. 18H). This is a clear example of neoteny, leading to paedomorphosis.
Equivocal or misplaced species
Gehypochthonius antonii Lombardini, 1962 — After its original description, based on material from Italy, this species seems not to have been mentioned again in the literature for three decades. Bernini et al. (1995) pointed out this oversight and noted that type material does not exist, but they did not discuss its possible identity. Subías (2004) listed G. antonii as a species inquirendum but in a recent update (Subías 2022) he included it in Eulohmannia, without discussion. Based on Lombardini's original description and figures, traits of G. antonii that are consistent with those of Eulohmannia (see below) include the following: (1) a facies described as slender (though not markedly so, based on his Fig. 1); (2) a prodorsum with an apparent posterior constriction (cervical collar) having longitudinal ridges on its ventral face; (3) a yellow coloration; (4) a strongly pectinate bothridial seta; (5) a lack of borders or apodemes associated with epimeres III and IV; (6) genital valves that are displaced far posteriorly; (7) a subterminal anal aperture; (8) an apparent parabolic scissure passing posterior to the genital valves and running toward the insertion of leg IV; (9) bidactylous pretarsi; and (10) a four-segmented palp. Collectively, this is a substantial set of traits that can be found in no known oribatid mite other than Eulohmannia, and the size is consistent with that of E. ribagai adults. But several other traits are incongruent with Eulohmanniidae. These include: (11) a hysterosomal dorsum with two transverse scissures (`sutures'); (12) three pairs of setae on epimeres III, IV; (13) only three pairs of genital setae; (14) only two pairs of distinct, relatively large aggenital setae, i.e., without an aggenital neotrichy of numerous small setae; (15) only three pairs each of anal and adanal setae; and (16) cheliceral digits that are, proportionally, strikingly large. Traits 11-16 cast doubts about the classification of this mite in Eulohmannia. Add to this the fact that Eulohmannia was well-known as a distinctive genus in 1962, having been included and illustrated in all general treatments of oribatid mites (e.g., Sellnick 1928; Willmann 1931; Baker and Wharton 1952; Balogh 1961) and it seems unlikely that Lombardini would not have recognized the genus. In this light, and in the absence of type material, it seems most reasonable to continue listing Gehypochthonius antonii as a species inquirendum.
Eulohmannia skrjabini Bashkirova, 1958 — The name of this mite, collected from southeastern Russia, was considered a junior synonym of Epilohmannia cylindrica (Berlese, 1905) (Epilohmanniidae) by Bulanova-Zachvatkina (1975); probably it was a new proposal, though she gave no such indication. This synonymy almost certainly is incorrect, though it has been maintained in various reviews and checklists, such as Marshall et al. (1987), Panʹkov et al. (1997), Ryabinin and Panʹkov (2002) and, most recently, Behan-Pelletier and Lindo (2019). Bashkirova's (1958) illustration (her Fig. 1) quite clearly shows the distinctive morphology of the family Perlohmanniidae. Rafalski (1966) thought it might be a junior synonym of Perlohmannia dissimilis Hewitt, 1908; Balogh and Mahunka (1983) treated it as a species inquirendum in Perlohmannia; and it was recombined to Perlohmannia (Perlohmannia) skrjabini (Bashkirova, 1958) by Subías (2004). In a recent overview of Perlohmanniidae, Ayyildiz et al. (2016) recombined the name to Hololohmannia skrjabini (Bashkirova, 1958), but it remains known only from the poor original description and was considered a species inquirendum by Subías (2022).
New diagnosis of Eulohmannia
Based on our studies of E. ribagai and the other species discussed above, we offer the following revised diagnosis.
With characters of Eulohmanniidae (see below). Most body integument with epicuticular reticulation of sharply defined, depressed lines. Rostral tectum with medial mucro, with or without small emargination on either side; seta exa about twice length of le, or more. Lyrifissure ip posterior or posterodorsal to seta f2 . Small sejugal apodeme present, on which some extrinsic muscles of trochanter III insert. Gland g4 opening above insertion of leg IV. Anal segment with typical ontogeny, forming in deutonymph with setae present. Setae ps1 of larva and ad1 of protonymph thickened, barbed, unlike other setae of segment. Pretarsi of adult legs usually with large lateral claws, empodial claw minute, bidentate; rarely (Eulohmannia sp. B from Kashmir) lateral claws absent, empodial claw fully formed, hook-like. Palp without vestige of articulation between fused femur and genu. Males present or absent, i.e., with sexual or thelytokous species or populations.
Paedolohmannia n. gen.
ZOOBANK: 2486996C-711A-42D3-ACA1-8D3DB9BA53CD ![]()
Diagnosis — With characters of Eulohmanniidae (see below). Body cuticle indistinctly colliculate, elevations outlined by fine, sparse punctation; mostly without reticulation of sharply defined, depressed lines. Rostral tectum with deep medial emargination, without mucro; seta exa less than twice length of le. Lyrifissure ip lateroventral to seta f2 . Sejugal apodeme absent; all extrinsic muscles of trochanter III insert directly on epimere III surface. Gland g4 opening not observed. Anal segment absent from all instars: adanal segment paraproctal in nymphs and adult. Setae ps1 of larva and ad1 of protonymph not hypertrophied, similar to other setae of segment. Pretarsi of adult with large lateral claws; empodial claw minute. Palp with vestige of articulation between fused femur and genu, in form of shallow crease. Males frequent.
Type species — Paedolohmannia metzi n. sp.
Etymology — The genus name is based on a combination of the Latinized Greek paidos (child) with the root lohmannia, which is used in numerous names for early- to middle-derivative oribatid mites. It reflects the paedomorphic nature of the type species and is considered feminine.
Justification — We justify this new genus proposal based primarily on the complete suppression of the anal segment (AN), including its associated setae and lyrifissures. While suppression of AN is widespread in Prostigmata (Kethley 1990), it is rare among Oribatida and Paedolohmannia may represent only the second example. Most oribatid mites have suppressed the primitive peranal segment, which is retained only by several members of Parhyposomata and Enarthronota (Grandjean 1939d; Norton and Fuangarworn 2015), but we believe some previously proposed losses of AN in Oribatida are incorrect, or are at least equivocal and differently derived, as explained below.
The independence of AN and the adanal segment (AD) is lost in various groups where fusions occur between cuticular components, i.e., the anal and adanal plates, with common and obvious examples being members of Phthiracaroidea. But in most instances, there is evidence that the segment itself is not lost; this might be clear from the unsclerotized paraprocts of juveniles, or in the continued presence of anal setae or their vestiges. In several genera of the enarthronote family Lohmanniidae (Torpacarus, Javacarus, Euryacarus) no clear anal plate is discernible and no unequivocal anal setae are present. These represent the culmination of trends in plate fusion and setal reductions that are seen throughout the family, as indicated by the presence of clear intermediate states (Grandjean 1950b, Balogh 1961). Grandjean (1950b) suggested that segment AN is absent in Torpacarus, but we disagree. We interpret the small fifth seta that forms anteriorly on the paraprocts of the deutonymph (Bischoff de Alzuet 1971) as the anterior anal seta; in some other lohmanniids this seta is positioned at the anterior end when the plate is clearly present (e.g., Haplacarus rugosus Mahunka, 1987).
The trend toward loss of independence of the anal plates from adanal plates, and the regression or loss of anal setae, seems to have been established early in the evolution of Hypochthonioidea, to which Lohmanniidae belong (Norton 2010). But in most instances the segment itself probably does not disappear. In Eohypochthonius (Neoatrichosus), for example, the anal segment clearly appears in the deutonymph, even though the anal plate is vestigial in the adult (Fernandez 1984). This trend suggests the existence of a long-term selective pressure in Hypochthonioidea, which may relate to reducing articulations that are vulnerable to predators, or that represent unnecessary flex points which reduce the efficiency of hydrostatic control.
The suppression of AN in Paedolohmannia seems fundamentally different: normal segmental addition is abruptly curtailed and the neotenic retention of a protonymphal segmentation results in paedomorphosis. The hypochthonioid family Psammochthoniidae provided the first clear oribatid mite example (Fuangarworn and Norton 2013) and Paedolohmannia represents the second.
Paedolohmannia metzi n. sp.
ZOOBANK: AA349558-661E-465D-A4BF-B0088E33CD32 ![]()
Diagnosis
As for genus.
Etymology
The species epithet is a genitive honoring the memory of the late Dr. Louis J. Metz, a soil scientist at the USDA Southeast Forest Experiment Station (Research Triangle Park, North Carolina, USA). He was an important influence and benefactor in the early career of the first author, and also collected the type series of specimens.
Adult
Dimensions — Total length of female (n = 7) 631–698, male (n = 9) 621–660; maximum width of female 170–209, male 166–198. Length of notogaster: ~ 2.0–2.2 times width and ~1.4–1.7 times prodorsal length (including cervical collar).

Integument — Color creamy to pale yellow in preserved specimens. Colliculate surface sculpturing of contiguous, very low mounds (Fig. 19A, C) having polygonal outlines (nearly invisible in transmitted light except at strong angle and high contrast). Punctation (pit diameter usually ~1) in shallow valleys between mounds (Fig. 19F) more conspicuous than mound itself in transmitted light (Fig. 22H, I). Underlying cuticular porosity well developed on prodorsum, less conspicuous or not discerned in other regions.
Prodorsum — Cervical collar narrow (~ 60% of maximum prodorsal width), with numerous parallel longitudinal ridges, especially strong ventrally (Figs 19C, 22J). Cuticle tesselate posterolateral to seta ro (Fig. 22B), with sharply defined depressed lines (similar to that of Eulohmannia), reticulation width 8-12. Medial indentation of rostrum roughly trapezoidal. Setae ro (32–36), le (20–24), in (41–49), exa (32–36) and exp (16) attenuate, nearly smooth (Figs 19A, 20A). Bothridial seta (73–82) sub-pectinate, with 6 to 8 (rarely 10) relatively short tines (length mostly 8-10) on outer curvature in distal part, alternate tines slightly divergent; glabrous basal part occupying up to half setal length. Concentric ridges of inner two bothridial chambers conspicuous (Fig. 22D); porose saccules strongly dimorphic (Fig. 22E): length of elongated saccule up to 3× diameter of round saccule.

Notogaster (Figs 19A, 20A, B, 21A, B) — Widest in middle third, sub-elliptical, such that lateral region of epimere III and seta 3d usually visible in dorsal view. All 15 pairs of notogastral setae relatively short (d1, e1, h1: ~20; p3: 20–24; others: 28–36), attenuate, without barbs. Lyrifissures ia, im, ip 13-16; ip positioned low on notogaster, well ventral to seta f2 and best discerned in lateral view (Fig. 21A). Lyrifissures ih, ips 9-10; ips close and parallel to margin but ih variable: positioned midway between ips and leg IV insertion or slightly closer to either, oriented parallel and close to edge of notogaster (Fig. 22H) or slightly removed and obliquely oriented (Fig. 21A).

Lateral podosoma and coxisternum (Figs 19C, 20B) — With vertical ridge posterior to leg I insertion (Fig. 22F; r1); ridge-like border (r2) of rudimentary acetabulum distinct, anterior to leg II insertion. Each ridge ending dorsally in small, rounded lobe, just below podocephalic canal. Canal horizontal for most length, deflecting slightly dorsad between r2 and edge of cervical collar (at * in Fig. 22F). Seta eI (8–10) not conspicuously bifurcated: usually with minute distal fork or one short barb (Fig. 22C). Scissure ncx narrow, beginning just posterior to insertion of leg IV, running anteriorly just above leg III insertion, then apparently effacing before reaching sejugal articulation, such that notogaster and epimere III seem fused anteriorly (Fig. 21A). Gland g4 opening apparently absent from region of leg IV insertion. Setal formula for epimeres I-III: 3–1–4; IV neotrichous. Setae of various lengths: relatively short (1c, 2a, 3b: 20–24; 3c: 36; others: 12–16), thin, without barbs.

Anogenital region (Figs 19C, 20B, 22H, I) — Parenthetic'arms' of sclerotized aggenital region tapered posteriorly; consistently with small, round or oval'island-like' sclerite distinctly separate from end of each arm (Figs 20B, 24A). Setae simple, attenuate, without barbs but sometimes vaguely roughened. Exclusive of four epimere III setae, with 17-18 pairs of neotrichous setae on fused epimeral-aggenital plate; most ~10 long but up to 20 near leg IV insertion. Genital aperture oval, plates 50-54 long; both sexes usually with nine pairs of attenuate, nearly smooth genital setae, about third of examined specimens with either eight pairs or asymmetrical (8/9); if only eight present, most anterolateral seta seems missing. Anal aperture ovate in ventral view, with four pairs of equally spaced, attenuate and nearly smooth adanal setae (ad1: 24–28; ad2: 18–20; ad3: 16–18; ad4: 12–14). Adanal lyrifissure ~10 long, inconspicuous, close to and parallel with lateral margin of plate, lateral to ad1. Structure and setation of genitalia as in E. ribagai (see above); ovipositor setae ψ1 ~20, ψ2 ~10, four τ setae 7-13; setae of spermatopositor 5-7 (Fig. 24A). Gravid females with single large egg, 230-260 × 140-150.
Gnathosoma — Subcapitulum (Fig. 21C) longer than wide (~110 × 77–82). Genae with distinct punctation but otherwise smooth (Fig. 19F); mentum basally with distinct, transversely elongated pattern of grooves and mounds. Subcapitular setae (a: 28–32; m, h: 20–24) similar in form: attenuate, without barbs but may be slightly roughened. Adoral seta or1 (12) straight, distinctly barbed, thickest; others (or2: 16; or3: 12) attenuate, slightly barbed, or3 thinnest. Labrum with ciliate distal margin (remainder not studied). Palp (length 69–73) with setation 0–[2+0]–2–7(+ ω); eupathidia and solenidion as in E. ribagai. Vestigial surface crease separating palp femur and genu segments (* in Figs 19B, 21E) very faint. Postpalpal seta (8) similar to eI: bacilliform but usually with minute, inconspicuous distal fork. Chelicera (length 118–123) with smooth seta chb (12–14), several vertical rows of small denticles near mid-length on adaxial face and usually two larger tooth-like spicules of similar size (Fig. 21D); oblique row of denticles on movable digit minute.

Legs (Fig. 23) — General form and properties as in E. ribagai. Average relative lengths of legs I-IV 1: 0.79: 0.82: 0.88; leg I ~ 0.4 times body length. Femur IV ~1.6 length of trochanter IV. Pretarsi heterotridactylous: lateral claws weakly barbed dorsally, empodial claw minute, toothlike, blunt (Fig. 24F, insert). Leg setal and solenidial formulas and their homologies as in E. ribagai (Tables 3, 4) with two exceptions: (1) tarsus I with 24 setae (cn2ʺ present); (2) femur II consistently with five setae (vʹ present). Famulus of tarsus I (Fig. 24G) as in E. ribagai. With 10 tarsus I eupathidia: (it), (p), (u), aʺ, cn2ʺ, mʹ, s.
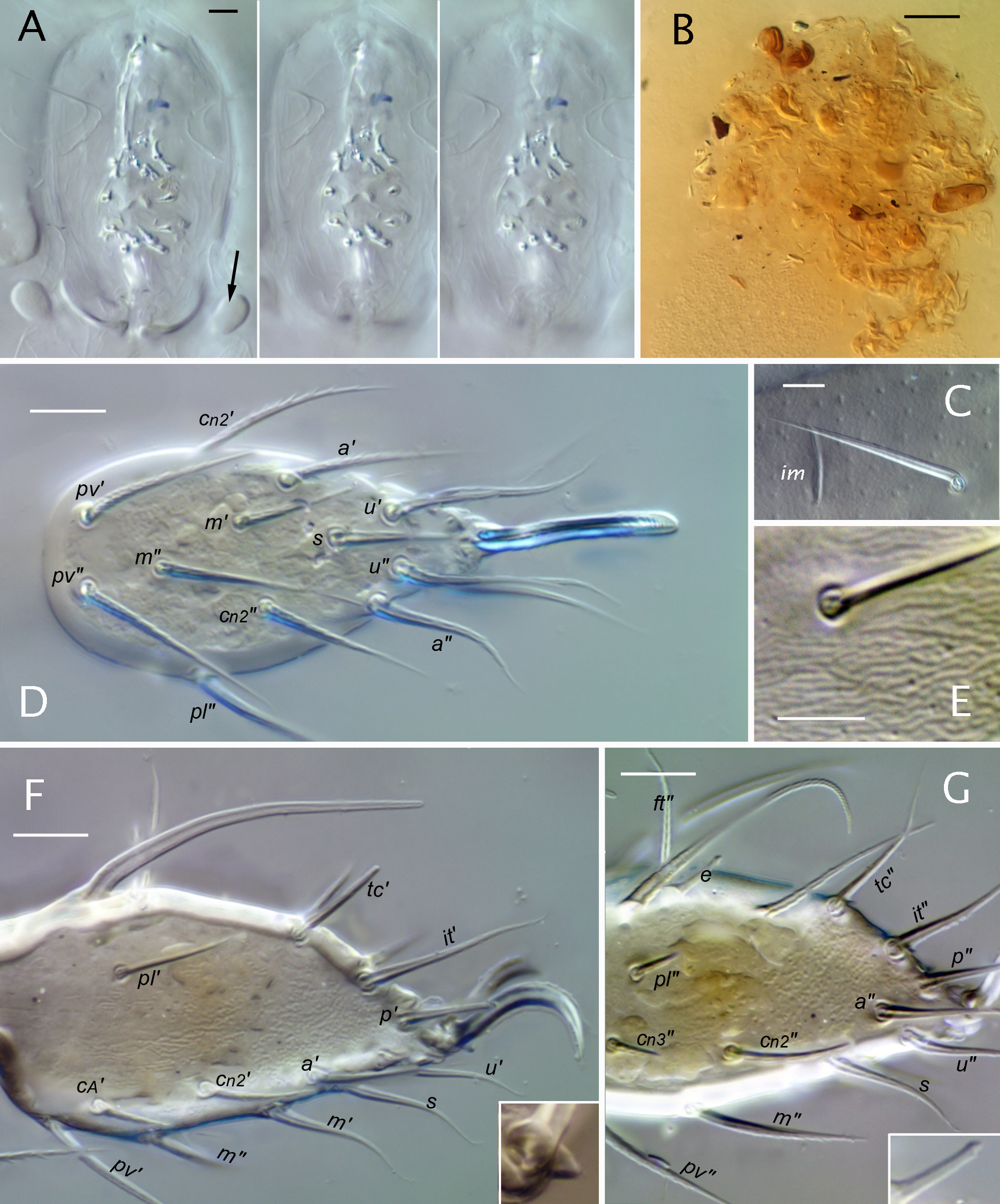
Juveniles
Dimensions — Total length of La (n = 3) 315–332, Pn (n = 10) 398–446, Dn (n = 4) 456–495, Tn (n = 6) 534–611. Total width of La 108–136, Pn 132–156, Dn 146–171, Tn 165–174. Gastronotum length: width ratio, La 1.5-1.8, Pn 1.8, Dn 2.0, Tn 2.0. Length ratio of gastronotum: prodorsum ~ 1.4 in La, increasing to ~ 1.5 in Tn.
Integument — Body cuticle colorless. Surface sparsely punctate (pit diameter up to 1).
Prodorsum (Figs 25A, B, 26F) — Medial emargination of rostrum trapezoid or quadrangular. Setae ro, le, in, exa, exp attenuate, without barbs; in about twice length of le, with exa intermediate (Table 6). Bothridial seta with 6-7 long cilia unilaterally and some inconspicuous small barbs on opposite side; bothridial saccules homomorphic, spherical in larva and nymphs. Seta eI with minute distal fork, rarely with one short barb or acuminate.
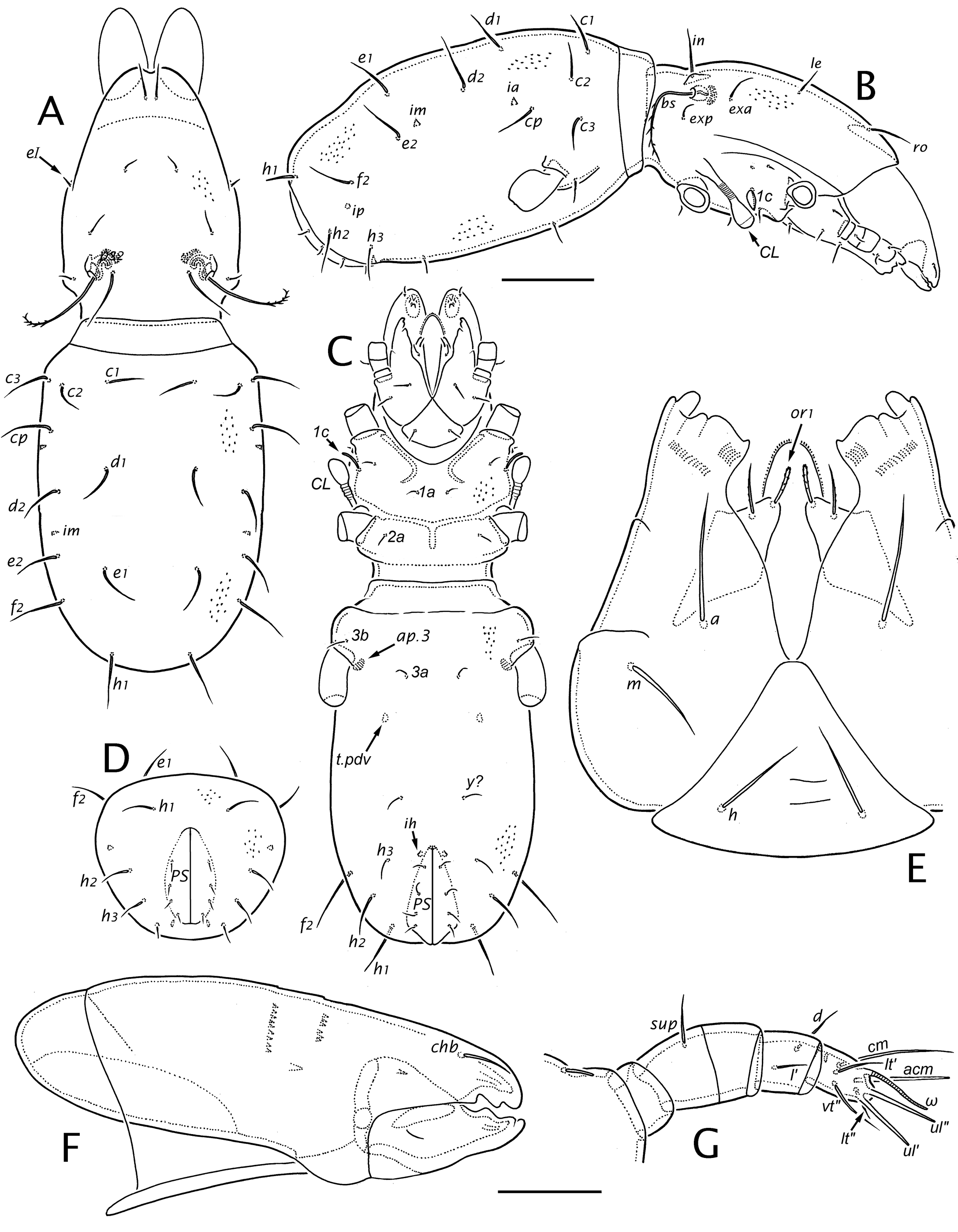
Gastronotic region (Figs 25A-D, 26F) — General structure and setal complements as in E. ribagai. Setae attenuate, without barbs; lengths given in Table 6. In larva, lyrifissures ia, im and ip positioned as in E. ribagai: in nymphs ia shifts dorsally as in E. ribagai, but ip remains low, ventrolateral to seta f2.
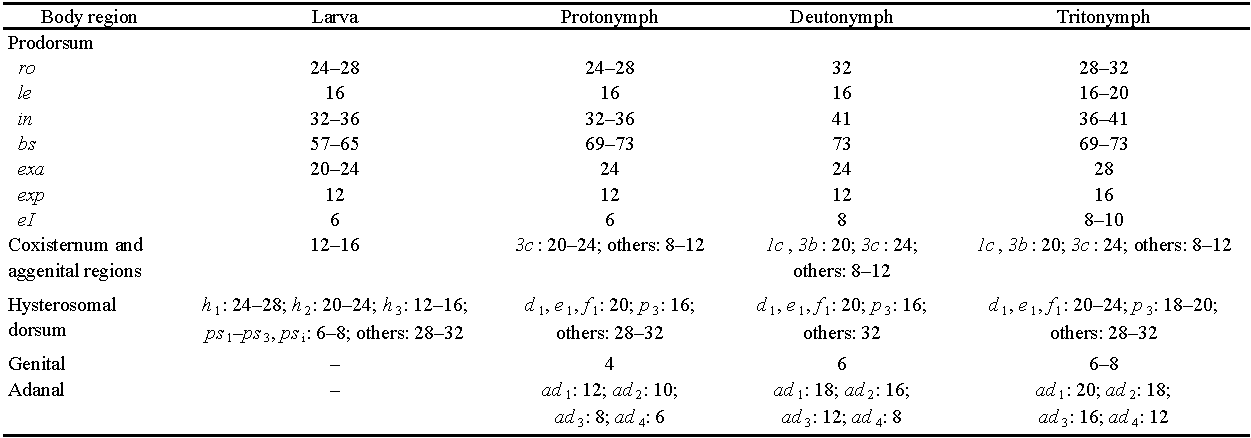
Epimeral region (Figs 25C, 26D, E, G) — Sejugal apodeme absent. Larva with epimeral seta 1c and Claparède's organ as in E. ribagai except stalk or organ with more numerous annuli (~ 10; Fig. 19G), Setae (except 1c in La) thin, attenuate, without barbs; setal complements of epimeres I-III as in E. ribagai, that of epimere IV uncertain, merged with neotrichous setae of aggenital region (see below). Setal lengths given in Table 6.
Anogenital region (Figs 25C, 26D, E, G) — Tendon insertions for dorsoventral muscles as in E. ribagai. Setal lengths during ontogeny given in Table 6; all setae attenuate, without barbs, including ps1 of La and ad1 of Pn. Genital setation (Pn to Tn): 1-4-7. Adanal segment appearing in Dn with full complement of four pairs of setae. Cupules ih, ips and iad appearing in normal ontogenetic pattern. In place of setae x, y in E. ribagai, La with single pair of setae anterior to paraprocts (Fig. 25C; identity equivocal, but perhaps homologous with y considering broader spacing of pair); second seta (presumably x) appearing in Pn (Fig. 26D); combined neotrichous setal complement from epimere IV and aggenital region: La to Tn: 1-5-10-14.

Gnathosoma (Fig. 25E-G) — Subcapitulum longer than wide; setae attenuate, weakly barbed, similar in thickness. Ontogeny of adoral setae as in E. ribagai; or1 thick, distinctly barbed; others attenuate, weakly barbed. General structure of labrum as in E. ribagai. Palp femur and genu fused, but with crease-like vestige of articulation, as in adult; palp setation as in E. ribagai, with seta inf formed in Pn; postpalpal seta slightly bacilliform, smooth. Chelicera similar to that of adult; seta chb smooth. Subcapitular dimensions and setal lengths during ontogeny presented in Table 7.

Legs (Fig. 26A-C) — Structure of legs (including pretarsus) as in E. ribagai. Ontogenetic formulas and homologies of setae and solenidia as in E. ribagai (Tables 3, 4) with following exceptions: trochanter IV seta lʹ forms in Tn (2 setae total) not Ad; seta vʹ consistently appears in Tn (6 setae total) on femur I and in Ad on femur II (5 setae total). Transformations of tarsus I setae to eupathidia as in E. ribagai, except: aʺ transforms in Pn, with aʹ remaining normal through adult; uʺ transforms in Dn; cn2ʺ transforms in Tn (see Table 5, R21). Famulus of tarsus I as in adult.
Material examined
Type locality — USA, Oregon, Lincoln Co., Cascade Head Experimental Forest, 45°3′24″N, 124°0′17″W. Holotype (unsexed) and 5 paratype adults from A1 soil horizon under red alder (Alnus rubra Bong.) forest litter; 12 paratype adults from A1 layer under Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) forest litter; all collected by L. J. Metz (1969). Holotype and 4 paratypes in alcohol, deposited in USNM collection; 5 paratypes (in alcohol) deposited in TSUMZ; 2 paratypes (in alcohol) deposited in CNC. Remaining paratypes (1 on slide, 5 in alcohol) in RNC. Non-type juveniles from same collections (3 La, 10 Pn, 3 Dn, 2 Tn) divided between TSUMZ and RNC.
Other material — California: Marin Co., near Point Reyes, 18-xii-1960, J.S. Buckett, col. (1 Ad; no habitat given; CNC). Oregon: Benton Co., Mary's Peak, North Trailhead #53, 1800′ elev., 22-ii-1976, L. Russell, (1 Ad on slide, 4 Ad, 2 nymphs in alcohol); same, from alder litter around large old stump (4 Ad, 1 nymph in alcohol); Coos Co., Cape Arago, Seven Devils Road, 2-i-1977, L. Russell, from alder litter (2 Ad on slides; CNC). Washington: Wind River Experiment Forest, Pinchot National Forest, 1969, L.J. Metz, from A1 soil horizon under Douglas-fir litter (6 Ad, 2Tn).
Reassessment of Eulohmanniidae
Previous diagnoses of the redundant family-group taxa Eulohmanniidae and Eulohmannioidea have been based on only Eulohmannia ribagai (Grandjean 1954a; Lee 1985; Weigmann 2006; Norton and Behan-Pelletier 2009). Below, we propose a new diagnosis based on all currently available information, including unnamed species discussed above. Corrections and comments relating to previous diagnoses follow the diagnosis.
Expanded diagnosis
Adult
Narrow, elongated (~570–850 um), nearly cylindrical species with dichoid body form; hysterosoma ~1.4–1.7 times longer than proterosoma; pale yellow or orange cuticle, without noticeable melanization. Proterosoma strongly constricted proximally forming waist-like cervical collar, enveloped by hysterosoma during contraction and telescopic folding of broad sejugal articulation. Rostral tectum moderately developed dorsally, narrowing laterally to expose much of retracted gnathosoma. Podocephalic canal conspicuous, extending to cervical collar; supracoxal setae eI strongly or weakly forked. Prodorsum with two pairs of exobothridial setae; exa displaced dorsally, inserted anterior to bothridium. Bothridial seta pectinate; each bothridium with two secretory porose saccules of different size. Notogaster without suprapleural scissure; separated from venter by broad plicature band (na) posterior to metapodosoma but only by narrow scissure (ncx) above legs; na and ncx not connected, notogaster and venter fused in small region immediately behind legs IV; notogastral margin projected on either side as acute angle at junction of na and adanal plicature band bpv, directed toward ventral midline between genital and anal apertures. With 15 pairs of notogastral setae; f1 absent. Opisthonotal gland absent. Epimere II with rudimentary, partial acetabulum at leg II insertion. Epimeres III, IV and aggenital region fully fused, unbroken by epimeral borders or noticeable apodemes; apodeme ap.3 present but small, inconspicuous; small sejugal apodeme present or absent. Epimere IV and aggenital region neotrichous, together with ~15 or more pairs of setae. Genital aperture small, oval, displaced far posteriorly; with 7-10 (usually 9) pairs of genital setae; genital papillae elongated, slightly curved. Aggenital sclerotization incomplete posterior to genital plates (illustrations showing complete envelopment need verification). Anal aperture nearly terminal; adanal and anal (if present) segments each with four pairs of setae. With small preanal apodeme. Ovipositor with six pairs of setae, without coronal (k) setae; spermatopositor with seven pairs (probably including one pair of k-setae). Subcapitulum stenarthric, with small, triangular mentum and large gena; rutellum weakly atelobasic, short, broad, with thumb-like lateral lobe and rutellar comb comprising two rows of cilia on dorsal face; with three pairs of adoral setae, or1 not bifurcated. Palp four-segmented, femur and genu fused, with or without vestige of articulation as inconspicuous crease; tarsal solenidion long, nearly prone, parallel with segment; setae (ul), and usually acm, eupathidial. Chelicera chelate-dentate; proximally inserted in body wall and adaxially emarginate; lamellated organ and oncophysis opʹ present; Trägårdh's organ and seta cha absent. Cheliceral body with vertical rows of small denticles on adaxial face; fixed digit with oblique row of minute denticles on abaxial face (these are present in E. bifurcata, though not shown in Fujikawa's (2014) Fig. 4E. Leg pretarsi usually heterotridactylous with strong lateral claws and minute empodium; rarely monodactylous. Solenidia baculiform or piliform, none flagellate; none coupled to seta d on any segment; genu I with three solenidia, σʹ coupled to small seta lʹ. Tarsus I without proximal accessory setae in l- or v-rows, but with 2-4 c-setae; monotropic setae mʺ and mʹ present; with 9-11 eupathidia, including (p), (u), (it), s and mʹ, plus a- and c-setae according to species; famulus bacilliform, with small conical head. Tibiae I and II with verticil of five setae (cʺ absent).
Juveniles
Body pale, general cuticle somewhat elastic, resistant, strongly layered, birefringent (R8); without distinct sclerites (but see above, Suzuki 1979). Proterosoma without lateral prodorsal rib (`nervure'). Larval Claparède's organ clavate, stalk with uniform, well-defined annulations; seta 1c scaliform in larva, normal in nymphs. With inguinal seta in ps row, but not h row; no paraproctal atrichosy. Ventral neotrichy incremental, beginning in larva. Seta inf of palp femur forms in protonymph. Leg pretarsi monodactylous; empodial claw with small basal spine. Genu pore absent, but genua I-III of larva with seta d vestigial; protonymphal leg IV chaetome: 0-0-0-1-7 (tibia with vʺ present).
Notes on previous diagnoses
Stegasimy. This trait relates to the development of the rostral tectum. In Grandjean's (1954a) definition, a mite is stegasime if the rostral tectum is large enough to cover and protect the gnathosoma when it is retracted, e.g., in a defensive posture. At its extreme development, chelicerae and palps are fully contained within the camerostome, with the tectum forming the'roof' and the subcapitulum forming the'floor' of this secondary chamber. Norton and Behan-Pelletier (2009) followed Grandjean (1969) in describing the prodorsum of Eulohmanniidae as stegasime, but Haumann (1991) and Weigmann (2006) considered it astegasime. The problem lies in applying a rigid terminology (covered or uncovered) to a continuous, gradational trait. In Eulohmanniidae the retracted chelicerae and palps are mostly covered by the modest tectum in dorsal view but are significantly exposed laterally, because the tectum narrows as it approaches the subcapitulum. Haumann (1991) and Weigmann (2006) considered the Eulohmanniidae condition as'secondary' astegasimy; this implies an evolutionary regression, a reversal in the development of the rostral tectum, but no explanation was given and we disagree with this characterization. There is a wide spectrum of tectal development in adult mixonomatans, from virtually none (Nehypochthoniidae) to moderate (Eulohmanniidae, Epilohmanniidae) to strong (Perlohmanniidae, Collohmanniidae, Ptyctima) but we see no evidence of a regressive trend.
Hysterosomal sclerites. Norton and Behan-Pelletier (2009) indicated that epimeres III-IV are seamlessly fused to the notogaster. This is not true: scissure ncx intervenes, except for a very small distance posterior to leg IV.
Notogastral setation. Previous diagnoses all have considered seta f1 present, following Grandjean (1954a). We now believe f1 is absent (R6).
Palp segmentation. Norton and Behan-Pelletier (2009) wrongly listed the palp as having five segments.
Thelytoky. Based only on E. ribagai, the family has been characterized as parthenogenetic, but this is an overextension: some species are almost certainly sexual (R29).
Relationships of Eulohmanniidae
Historical overview of classification
The earliest family-group classification of Eulohmannia was that of Berlese (1913), who considered it one of six genera of the tribe Michaeliini in a very broad concept of the family Nothridae. This was soon adjusted (Berlese 1916b) by including it as one of five genera in Lohmannini [sic], the latter being one of two tribes of his newly proposed Malaconothridae. Sellnick (1928) maintained this classification, but under the elevated family taxon Lohmanniidae. Grandjean (1931) recognized the heterogeneity of Berlese's taxon and proposed the family Eulohmanniidae, which has remained monogeneric except in the classification of Baker and Wharton (1952). These latter authors tentatively included Collohmannia and Perlohmannia, transfers that have been ignored by other authors.
Grandjean (1931) considered Eulohmanniidae to have closest affinity with Nanhermanniidae, which have a ventral plate organization that in some ways is similar (see R4). Willmann (1931), who separately proposed Eulohmanniidae several months later (Hammen 1959), gave a name to this pair of families: the subcohort Diagastres, within the large cohort Aptyctima that included the bulk of oribatid mites. Willmann clearly stated this was a pragmatic grouping, and he doubted this pairing comprised a'systematic unit' but his classification was maintained in synoptic literature into the 1950s (e.g., Vitzthum 1943; Radford 1950; Baker and Wharton 1952).
In his iconic essay on oribatid mite classification, Grandjean (1954a) considered Eulohmanniidae to be one of 11'natural groups', thereby abandoning Willmann's system; he proposed no enveloping higher classification, but two views soon arose. The previously supposed relationship with Nanhermanniidae was recognized formally by Baker et al. (1958) and Woolley and Baker (1958), who included Eulohmanniidae in the newly recognized Nanhermannioidea, along with Nanhermanniidae and Epilohmanniidae. We know of no other authors who used this classification. The same year, Grandjean (1958a) proposed'Perlohmannoidea' (properly Perlohmannioidea Grandjean, 1954a) to include Eulohmanniidae and three other single-family natural groups that he considered relictual: Perlohmanniidae, Epilohmanniidae and Lohmanniidae. This latter superfamily classification was used—with the addition of Collohmanniidae, about which Grandjean (1958a) was equivocal—in an influential series of papers by Balogh (e.g., 1961, 1965) and by Bulanova-Zachvatkina (1967). Subsequently, Grandjean (1969) demoted Perlohmannioidea by essentially reverting to his 1954a opinion about the isolated, relictual nature of the four families. He elevated each to a monofamilial superfamily and did the same with Collohmanniidae. To these he added the speciose Ptyctima (his Euptyctima: Phthiracaroidea and Euphthiracaroidea) and proposed the unranked taxon Mixonomata for the seven superfamilies.
Grandjean's classification of Eulohmannia in the monobasic Eulohmanniidae and redundant Eulohmannioidea has remained in general use, though the acceptance and composition of Mixonomata has varied. Mixonomata was ignored in some important works (e.g., Balogh 1972, Ghilarov and Krivolutsky 1975) but retained mostly in its original sense—adding Nehypochthoniioidea as suggested by Norton and Metz (1980)—in others (e.g., Fujikawa 1991; Balogh and Balogh 1992). Under the name Mesomixosomatae, Lee (1984, 1985; ranked respectively as a subcohort or section) recognized Grandjean's (1969) original concept of Mixonomata except for the removal of Lohmannioidea (see also Norton 2010). A concept of Mixonomata that included Nehypochthonioidea and excluded Lohmannioidea was ranked as an infraorder by Schatz et al. ((2011) and earlier as a supercohort by Norton and Behan-Pelletier (2009); the latter authors used the variant name Mixonomatides to be consistent with the overall mite classification (Krantz and Walter 2009). None of these works altered the redundant classification of Eulohmanniidae in Eulohmannioidea or proposed groupings that linked Eulohmanniidae to other mixonomatans.
Phylogenetic relationships
Inferences from morphology
In a detailed analysis of Perlohmanniidae, Grandjean (1958a) noted that discovering relationships is much hampered in such a relictual, isolated group with fundamental differences in morphology yet little diversity, and this certainly also applies to Eulohmanniidae. The only formal cladistic analysis that included Mixonomata was that of Haumann (1991), who identified it as one of several paraphyletic groupings that have been, and continue to be, used in oribatid mite classifications (see also Norton 1998).
Haumann (1991) could not identify a sister-group of Eulohmannioidea for lack of appropriate synapomorphies, but he included E. ribagai in his cladogram (p. 189) as part of a pectinate series of taxa, positioned between Nothrolohmannia and Lohmanniidae. The latter two have since been considered members of Enarthronota (Hypochthonioidea; Norton 2010). With these enarthronotes removed, the lack of synapomorphies (indicated by broken connecting lines) in the relevant part of Haumann's cladogram would condense it to a polychotomy, though he did not describe it in this way. This polychotomy would comprise five branches: (1) Eulohmannioidea; (2) Perlohmannioidea; (3) Epilohmannioidea; (4) a clade containing Nehypochthonioidea, Collohmannioidea and Ptyctima; and (5) a clade containing all the more derived oribatid mites. Thus, from the standpoint of Eulohmannioidea, there was no resolution of relationships.
Weigmann (2006) seems to have ignored these problems (i.e., the broken lines and unsupported branches) and adopted a somewhat simplified version of Haumann's branching topology. As a result, Weigmann's (2006, his Fig. 1) tree infers that Eulohmannioidea is the basal taxon in his sense of Holonota; its sister-group would be the vast clade that includes: other mixonomatans, plus all Nothrina (= Desmonomata sensu stricto), plus all'higher' oribatid mites (Brachypylina or Circumdehiscentiae), and even Astigmata, though this latter group was not considered (see Norton 1998; Norton and Behan-Pelletier 2009; Schatz et al. 2011). Again, the data in Haumann (1991) neither support nor reject this relationship.
Inferences from DNA
Phylogenies inferred from molecular studies that included Eulohmannia ribagai have been more numerous and diverse. In the first, Maraun et al. (2004) applied three analytical methods to sequences in the D3 region of 28S rDNA. The results differed greatly, and the branch representing Eulohmannioidea was either unresolved (neighbor-joining and maximum parsimony) or (maximum-likelihood) was at the base of an unrealistic clade containing Parhyposomata, Ptyctima and some portions of both Nothrina and Brachypylina—the latter two having major sections elsewhere in the tree. By contrast, in the cladogram of Schaefer et al. (2010), based on a Bayesian analysis of 18S rDNA sequences, E. ribagai is positioned as Weigmann (2006) depicted, and their dating method suggested that the branch diverged in the mid- to late-Ordovician Period.
Relationships of E. ribagai became less clear in several subsequent studies based on 18S that added representatives of a taxon not sampled by Schaefer et al. (2010): the enarthronote superfamily Protoplophoroidea. In a study of the evolution of defense mechanisms, Pachl et al. (2012; their Fig. 1) presented a favored Bayesian inference tree that shows E. ribagai as the sister-taxon of Protoplophoroidea with strong support (posterior probability = 100); this sister-relationship did not exist in their other two analyses (a different Bayesian model and a maximum-likelihood tree) that were more equivocal regarding the position of E. ribagai. A later study (Pachl et al. 2017), in which partial 28S sequences were added to those of 18S, inferred this same odd grouping (E. ribagai + Protoplophoroidea) with all analyses, though in this instance the cluster also included Gehypochthonius (Gehypochthoniidae). This rather astonishing small clade therefore included members of three traditional oribatid mite higher taxa: Mixonomata, Enarthronota and Parhyposomata. Similar clusters appear in other 18S trees: the Bayesian inference tree of Xue et al. (2017) and both maximum-likelihood and neighbor-joining trees of Gong et al. (2018). Most recently, the maximum-likelihood 18S tree of Pachl et al. (2021) disperses this incongruous grouping, nesting Protoplophoroidea within Enarthronota and essentially reverting to the topology of Schaefer et al. (2010) regarding Eulohmanniidae. I.e., if the oddly-placed endeostigmatid family Oehserchestidae were ignored, the labeling in the tree of Pachl et al. (2021) could have been modified to show E. ribagai as the most basal included member of Mixonomata.
The number of specific topologies seen in just these few published 18S trees is striking. Unfortunately, Eulohmanniidae has not been represented in any study using other genes (except the ineffective 28S, see above) or promising approaches, such as mitochondrial gene organization (Arribas et al. 2019; Li and Xue 2019) or genomic-scale analyses (Lozano-Fernandez et al. 2019).
Klimov et al. (2018) examined the phylogeny of acariform mites using partition analyses of six rDNA and coding genes. Although Eulohmanniidae was not included, the study hints that the topological instability and illogical positions of various enarthronote groups and Gehypochthonius, as seen in various trees based only on 18S, is artifactual. Most relevant is that they found Protoplophoroidea and Hypochthonioidea to consistently cluster, in accordance with morphological studies on Enarthronota (Norton 1984, 2001; Haumann 1991), i.e., not interrupted with mixonomatans or other extraneous taxa. Their maximum-likelihood, full partition tree additionally has Gehypochthonius— the only included member of Parhyposomata and an unstable taxon in 18S trees—at the base of the glandulate oribatid mites (Novoribatida of Haumann 1991), where traditional concepts would place it (Norton 1998, Weigmann 2006).
Conclusions
Grandjean's (1969) inclusion of Eulohmanniidae in Mixonomata seemed rather equivocal, since the single species he knew, E. ribagai, lacks two traits that typify Mixonomata and most of the more-derived oribatid mite taxa (Nothrina and Brachypylina): the opisthonotal gland and a notogastral setation in which seta f1 is reduced or lost. Grandjean (1954a) himself linked these two traits, putting great importance on the fact that f1 is susceptible to regression only in groups that either possess the gland or have secondarily lost it. The probable regressive loss of seta f1 (R6) shows that Eulohmanniidae conforms to other mixonomatans in notogastral setation and evokes at least a suspicion that the gland also may have been lost from Eulohmanniidae.
Grandjean (1958a, 1969) and Haumann (1991) both faced problems of mosaic trait distribution within the mixonomatans. While the family is now better known, the problems still exist, so we retain the current redundant classification of Eulohmanniidae in Eulohmannioidea. Eulohmanniidae have clear autapomorphies, some of which are striking: cervical collar; fused and neotrichous hysterosomal venter; narrowly cylindrical body with near-terminal paraprocts. Other autapomorphies are inconspicuous: eupathidial setae (u) on tarsus I (R21); and partly unpaired, perhaps amphistasic setae in the c-row of tarsus I (R20). Among mixonomatans, they have uniquely lost proximal accessory setae in l- and v-rows from tarsus I.
However, the position of Eulohmanniidae as the earliest derivative mixonomatan clade is equivocal. Based on outgroup comparisons with Palaeosomata and Parhyposomata, some plesiomorphic traits are at least consistent with this idea: a pectinate bothridial seta; the absence of paraproctal atrichosy; leg pretarsi with strong lateral claws and a regressed empodial claw (R24); a third genu I solenidion (R22); the lack of flagellate solenidia; the absence of solenidial coupling with seta d on genua and tibiae (R23); the retention of seta vʺ on tibia IV of the protonymph (R16); and the presence of setal pair (m) on tarsus I (R17). The annulated stalk of Claparède's organ is shared with Perlohmanniidae, but also with two of the three families of Parhyposomata (R10), so conceivably it is plesiomorphic within mixonomatans.
Despite retaining these ancestral traits, there are significant synapomorphies with other groups. A preliminary list would include regressive synapomorphies such as the loss of inguinal seta hi in the larva (R13), shared with Collohmanniidae and Ptyctima; the loss of seta cʺ from tibiae I and II, shared with Perlohmanniidae and Ptyctima; and larval suppression of genual seta d, shared with Perlohmanniidae and Nehypochthoniidae (R15). Progressive synapomorphies would include: the coupling of lʹσ, shared with Nehypochthoniidae and Ptyctima (R23); a rutellar comb with the most derived form found among mixonomatans (R25); a palp with fused femur-genu, found in Epilohmanniidae and some Ptyctima (R26); a proximally internalized and adaxially emarginated chelicera, found in Collohmanniidae (R27); cuticular birefringence in juveniles, shared with Perlohmanniidae and Collohmanniidae (R8). Several of these synapomorphies also are shared with Nothrina and Brachypyina, as discussed in the respective Remark. Considering the rather incongruous distribution of these traits, relationships of Eulohmanniidae, as well as the other distinctive mixonomatan families, probably will remain unknown pending focused and diverse molecular analyses.
Remarks
R1 — Body size. According to the catalogue of Castagnoli and Pegazzano (1985) Berlese's collection includes three type specimens of Eulohmannia ribagai, all from Tiarno (Trentino) in the Italian Alps. These are in slide boxes number 66, 70 and 86. Given the date approximations of Norton and Kethley (1989; pp. 433-434), these boxes were filled between 1908 and 1910, so it seems likely that all three were available at the time of the description, but only one length measurement was given:'680 µ'. This would be at the lower end of the size range for our New York population (679–752). However, his Fig. 51 shows a specimen that was maximally contracted. Since a fully extended individual is about 10% longer than if fully contracted, if Fig. 51 represents the measured specimen it would have been at least 730 extended, which is the mean length of New York specimens.
Other literature suggests that size ranges for adults in Palaearctic populations have smaller lower limits. Published length ranges include: Schweizer (1956) 612–675; Bulanova-Zachvatkina (1960) 640–720; Aoki (1975) 670–700; Balogh and Mahunka (1983) 600–700; Weigmann (2006) 600–730. Single measurements include those of Trägårdh (1910) 630, Sellnick (1928) 671 and Willmann (1931) 670. Sergienko (1994) reported 270, but this is an obvious lapsus. Overall, these published lengths range from 600–730, but some caution seems warranted. In some cases, numbers may have been strongly rounded, and often it is not clear if data are new or repeated cumulatively from previous literature, as, for example, the low number of 600 by Balogh and Mahunka (1983) and its possible repetition by Weigmann (2006). Other concerns are the degree of contraction and whether the gnathosoma, which extends beyond the rostrum, is included in the length measurement; this information rarely is stated but there can be a significant effect on measured length (see Material and Methods), and not all reports convincingly state that measured mites were adults. Of the material we studied directly, all juveniles and most adults from Palaearctic populations of E. ribagai fell within the ranges seen in New York; exceptions were several females from Germany (650–708, n = 11) and Khabarovsk (Russian Far East; 650–756, n = 9).
Hammer (1952) reported that the length of her Canadian (Northwest Territories, Reindeer Station) specimens identified as E. ribagai (760–780) was about 15% larger than those of Europe. However, we examined 13 adults from that same location, and these have a broader range: 640–786 (mean 728). Specimens from Alaska (Fairbanks) have a narrower but entirely overlapping range: 679–747 (mean 713; n = 8). While all studied Palaearctic juveniles and all adults from Sweden and China were within the New York range, our small samples (all females) from Germany and Khabarovsk (see above) each included several females below 679.
R2 — Bothridial saccules. Grandjean (1939b) suggested that in E. ribagai these saccules might function as resonating chambers if the trichobothrium were a sound receptor. Later (1969) he thought the saccules were respiratory (tracheal) structures, a role he ascribed to most porose cuticular invaginations. But ultrastructure studies (Alberti et al. 1997) revealed them to be unique secretory organs in E. ribagai. They are distinct from the respiratory saccules of certain other mixonomatans, which may enhance gas exchange in extrinsic cheliceral muscles (Norton and Alberti 1997), and we have no reason to consider them homologous.
R3 — Artifactual gastronotic crease. Upon treatment with lactic acid or other clearing agent, juveniles of E. ribagai and P. metzi may develop a transverse crease between setal rows d and e (Fig. 4E), which is a typical location of scissures in Enarthronota and Parhyposomata (e.g., Strenzke 1963; Norton 1975, 2001). However, when formed the crease varies in distinctness and the reticulate cuticular pattern of the gastronotum is never interrupted, showing that it is an artifact, not a scissure. We believe this variable crease may relate to the dorsoventral muscles that insert in this region (m.pdv in larva, m.mdv in nymphs, Fig. 6A). Hardening of the muscles during preservation, combined with the increased internal pressure and inflation often associated with the clearing process, might buckle the flexible dorsal cuticle of the juveniles. Adults never show this crease, presumably because the cuticle is stronger and there is no dorsoventral muscle in this region.
R4 — Hysterosomal structure, diagastry, pseudodiagastry. Eulohmanniidae and Nanhermanniidae often have been linked in the literature as having adults with a uniquely shared sclerotization pattern, termed diagastry. Grandjean (1954a) coined the term, referring to an adult as diagastric if the gastronotic (notogastral) sclerotization is prolonged without interruption between the genital and anal apertures across the plane of symmetry. While he cited Willmann (1931) for the concept, it was a different aspect of structure that the latter author stressed when proposing the taxon Diagastres for these two families. Willmann viewed the anal aperture as lying at the end of the hysterosoma in Diagastres, rather than being with the genital aperture on a common ventral plate; i.e., the boundary-articulation between dorsum and venter ran behind the genital aperture, not behind the anal aperture.
Early illustrations of Eulohmannia either have shown a continuous ring of sclerotization passing between the anal and genital apertures or lacked sufficient detail in that region to make a judgement (e.g., Sellnick 1928; Willmann 1931, Hammer 1952; Schweizer 1956; Bulanova-Zachvatkina 1960). Correcting his earlier view, Grandjean (1956b, his Fig. 1A) explained that in E. ribagai sclerotization was not in fact continuous and coined the term'pseudodiagastry' to denote the false impression. Difficulties in identifying the limits of the plates (`chitinization' in Grandjean's terminology) and articulations had caused the earlier misinterpretations. Subsequently, the correct general structure of the venter was illustrated by Kunst (1971), Aoki (1975), Weigmann (2006) and Fujikawa (2014). However, unlike Grandjean these latter authors depicted a complete envelopment of the genital aperture by posterior extensions of the sclerotized aggenital region in E. ribagai. We never observed this—regardless of provenance, the interval between genital and anal plates is entirely unsclerotized in the adult, as Grandjean illustrated.
Other than the seamless fusion between epimere IV and the aggenital region, the venter of Eulohmanniidae species differs from the typical macropyline venter, such as that of Perlohmannia, in two conspicuous ways. (1) The nearly terminal adanal plates of Eulohmanniidae adults maintain the curved, valve-like outline from when they were paraproctal, so that collectively with the aggenital region there is an hourglass configuration. (2) The notogastral margin, in the vicinity of seta ps3, projects medially at a strong angle to accommodate the'waist' of the hourglass. As in Perlohmannia, the lack of sclerotization in the small space between the genito-aggenital and ano-adanal regions is plesiomorphic. In identification keys, the anterior part of the hourglass is usually emphasized, resulting in the description of the Eulohmanniidae venter as having a'parabolic line' (or'suture','furrow') running between the genital and anal plates (Balogh 1961, 1972; Balogh and Mahunka 1983; Balogh and Balogh 1992). Setting aside the hourglass shape, nothing is unusual about the anogenital region of Eulohmanniidae, and it seems unnecessary to maintain the esoteric term pseudodiagastry, which seems to have been coined solely to identify a previous error.
Diagastry in Nanhermanniidae is indeed unique and has functional consequences: there is no sejugal articulation (they have a holoid body form) and only the anterior half of what might be considered a circumgastric scissure exists, leaving the hysterosomal cuticle as essentially a broad sclerotized ring between the genital and anal regions. Therefore, the usual source of hemolymph pressure control—dorsoventral musculature pulling top against bottom— seems impossible in nanhermanniid adults. By contrast, in adult Eulohmanniidae the notogaster is separated from a ventral plate by a significant articulating cuticle posterior to leg IV (plicature band na), a bendable narrow scissure (ncx) anterior to IV, and a broad telescoping sejugal articulation. These, and the various associated muscles identified above, provide multiple ways to control body pressure.
R5 — Number of gastronotic setae. The number of setae on the notogaster of adult E. ribagai has been reported differently in the literature. The first definitive statement seems to have been that in Grandjean's (1933) early treatment of oribatid mite development, where he included Eulohmannia (E. ribagai) in a list of genera with 30 notogastral setae (15 pairs) but at that time he lacked full ontogenetic data. For his second general treatment (1949b) he had studied all instars but did not include E. ribagai in his overall analysis due to an unusual and confusing setation in the larva (see R6, R13). He did not mention a specific count in his later summaries (e.g., 1958a, 1969). The first explicit contradictory statement seems to have been that of Bulanova-Zachvatkina (1960) who attributed 14 pairs to E. ribagai, only 12 of which were shown on her illustrations; without comment she later (1975) cited 16 pairs, the number also given by Balogh and Mahunka (1983), Haumann (1991) and Sergienko (1994). Lee (1985) referred to the chaetome as holotrichous, which implies 16 pairs, although he gave no count. None of these authors mentioned the discrepancy with Grandjean (1933). More recently, Weigmann (2006) cited 15 pairs, but without comment. This inconsistency may have influenced Fujikawa's (2014) assertion that the notogastral complement of E. ribagai is variable (15 or 16 pairs).
The discrepancies might relate to illustrations, rather than to direct study. Numerous authors have illustrated E. ribagai (see references in Marshall et al. 1987; Mahunka and Mahunka-Papp 1995) but most figures have been equivocal regarding setal number for one of three reasons. (1) Setae obviously had been overlooked (e.g., Schweizer 1956; Bulanova-Zachvatkina 1960). (2) Possible double-counting: long ventral setae such as epimeral seta 3d, or seta ad1 may also be seen in dorsal views (e.g., Willmann 1931; Hammer 1952 and the unattributed repetition of her figure by Balogh 1961, 1963, 1965, 1972). (3) It may be unclear if marginal setae were repeated in dorsal and ventral views (e.g., Kunst 1971; Aoki 1975 and the repetition of his figure by Balogh and Mahunka 1983; Balogh and Balogh 1992). The most reliable illustrations invariably show 15 pairs: these include figures by Lebrun and Wauthy (1981), Weigmann (2006) and Fujikawa (2014). All our specimens of E. ribagai have 15 pairs, distributed as shown by these later authors, and this chaetome is identical in the other species of Eulohmanniidae we studied herein.
R6 — Homologies of gastronotic setae. In the terminology developed by F. Grandjean, a notogastral complement of 15 pairs of setae is referred to as'unideficient', i.e., lacking one pair from the holotrichous condition of having 4-2-2-2-3-3 pairs in the c, d, e, f, h and ps rows, respectively. In most instances, a unideficient chaetome results from the loss of pair f1; this can be certain, due to the persistence of its alveolar vestige, or implied from other considerations (Grandjean 1954a). In Grandjean's conceptual model of regressive setal evolution, f1 has been considered the weakest of notogastral setae (Travé 1975). However, for the unideficient E. ribagai, Grandjean (1954a; see also 1958a, 1969) explicitly considered f1 to be present. To our knowledge, he never explained his view or indicated which seta might be absent, perhaps due to lingering uncertainties relating to larval setation (see R13).
Lebrun and Wauthy (1981)—the only authors we know to have applied notations to the entire notogastral chaetome of E. ribagai—considered the missing seta to be h3, and they drew a small cuticular mark suspected of being its vestige (their Fig. 2). Grandjean never mentioned having seen such a vestige, and it seems unlikely he would have ignored one if he had seen it; he described various setal vestiges on the cuticle of species in other oribatid mite taxa on numerous occasions and clearly appreciated their significance. Lebrun and Wauthy (1981) did not discuss this cuticular mark, nor did they indicate how many times they observed it, but we looked for it on approximately 40 adults from New York, Canada, Alaska, Germany, Sweden and Khabarovsk and failed to find it. Fujikawa (2014) described E. bifurcata as having 16 pairs of setae (including the purported h3) but our study of the type series failed to find more than 15 pairs on any specimen (see above).
Seta h3 indeed can be regressive in oribatid mites—absent without trace or forming a small alveolar vestige—but in most instances the regression affects only the larva: h3 appears as a distinct seta in the protonymph and remains in subsequent instars. Examples are common in Nothrina and Brachypylina (Grandjean 1949b), with a scattered distribution that indicates multiple evolution of this common regression (see focused reviews by Behan-Pelletier 1997; Norton 1998; Norton and Franklin 2018). A few mixonomatan examples are in Epilohmanniidae (Grandjean 1949b) and Phthiracaridae (Grandjean 1950a) but again, h3 is only delayed to the protonymph rather than being lost. Different ontogenetic patterns in seta h3 occur in Neoliodoidea and Plateremaeoidea. In Neoliodidae h3 is present in the larva then lost at some point in development (e.g., Ermilov et al. 2011; Seniczak et al. 2018). In Gymnodamaeidae h3 is minute or absent in the larva, developed normally in nymphs, and then lost in the adult (e.g., Bayartogtokh and Ermilov 2013, 2014). The only clear examples of h3 being lost from all instars are in Pheroliodidae and Plateremaeidae (e.g., Grandjean 1964b; Seniczak et al. 2020).
We contend that E. ribagai is not another example of h3 loss and that Grandjean (1954a) and Lebrun and Wauthy (1981) were incorrect in their assessment of notogastral setation. Instead, this and other eulohmanniids follow the usual model of unideficiency: pair f1 has been lost from all instars but without leaving a vestige. The hypothesis is not easily accepted if one examines only late nymphs or adult mites, since pairs e1 and e2 are followed immediately behind by two pairs having the same spacing and orientation (Figs 4C, 20A), just as one would expect of f1 and f2 in a holotrichous chaetome. But this uniformity is secondary, as can be understood in the context of developmental changes.
In the larva of both E. ribagai and P. metzi there are 12 pairs of gastronotic setae, contrasting with the 13 pairs expected in a holotrichous larva, so the unideficiency clearly applies to all instars. Also clear is that there are three pairs of setae on larval segment H, arranged parenthetically around the paraproctal segment PS (Figs 3B-D, 25B-D): pair h1 inserts just above the posterodorsal end of the paraprocts and is directed posteriorly or posterodorsally, h3 is a conspicuously smaller seta that inserts posterolateral to cupule ih, and h2 has an intermediate position. This pattern expresses the hypothetical distribution of h-row setae first described by Grandjean (1939d) and exactly mirrors that seen in most oribatid mites, including the diminished size of h3. This same larval complement has been illustrated for Parhyposomata (Strenzke 1963), and mixonomatans such as Nehypochthoniidae (Norton and Metz 1980), Perlohmanniidae (Suzuki 1977) and Collohmanniidae (Grandjean 1966; Norton and Sidorchuk 2014). Therefore, the missing seta is not h3, and must belong to a more anterior segment.
The number and arrangement of c- and d-row setae seems perfectly normal in the larva of E. ribagai and P. metzi (Figs 3A, D, 25A, B). However, in the space between them and the three h-row setae there are only three other pairs: the anterior two of these have the usual disposition for e1-e2, repeating that of d1-d2. Between the e-row and h-row is a single seta; its lateral position identifies it as f2, and the relatively large space between e1 and h1 strongly suggests that f1 is the missing gastronotic seta, despite the absence of a vestige. In later instars we believe seta h1 is displaced anteriorly to fill the sensory gap in the elongating body that would have been caused by the loss of f1.
Does the absence of a vestige negate our argument? To paraphrase Grandjean (1949b), vestiges show us not only that a seta can be regressed, but also that the seta in question might be lost without leaving a trace. There are ample examples of f1 vestiges in mixonomatans and Nothrina, and even Astigmata, but f1 has left no vestige in Brachypylina or the great majority of Astigmata (Grandjean 1954a; Norton 1998). Perhaps the best analogy to the eulohmanniid setation is in species of the nothrine genus Nanhermannia, which also have a cylindrical (though shorter) body form. These mites differ from eulohmanniids in having a larva that clearly lacks h3 and possesses f1, but all subsequent instars add the delayed h3 and lose f1 (Seniczak 1991). Nymphs may possess an f1 vestige or not, but f1 is absent without trace in the adult. With f1 being absent, seta h1 shifts to fill the sensory gap, and in the adult it has a spatial relationship with f2 that mirrors the relationship of e1-e2, just as it does in Eulohmanniidae.
R7 — Opisthonotal gland. Much has been written about the taxonomic distribution and chemistry of this paired exocrine defensive gland, which characterizes the Novoribatida, or'glandulate oribatid mites' (Norton 1998; Raspotnig 2006, 2010). Grandjean (1954a) viewed its distribution as problematic, but most issues disappeared when Astigmata were recognized as derived oribatid mites (OConnor 1984; Norton 1998; Dabert et al. 2010). The absence of these glands in Palaeosomata and Enarthronota is plesiomorphic, but as Grandjean (1954a) first noted, in at least some instances the absence of glands in more derived groups is secondary. They have been lost, for example, in the Ptyctima families Phthiracaridae and Synichotritiidae, as well as in various Astigmata taxa (Norton and Behan-Pelletier 2009, OConnor 2009).
Surprisingly, Grandjean (1969) did not clearly express an opinion about the absence of the gland in two of the families—Lohmanniidae and Eulohmanniidae—in his proposed Mixonomata, a taxon in which most families possess it. Regarding Lohmanniidae, there is now strong morphological (Norton 2010) and molecular (Dabert et al. 2010; Klimov et al. 2018) evidence they are part of the diverse enarthronotan superfamily Hypochthonioidea, so their lack of the gland is plesiomorphic. For Eulohmanniidae the issue seemed solved when the gland was purportedly found in E. bifurcata (Fujikawa 2014): this would suggest it was lost in other species of the family. Our study of the type series failed to find gla on any specimen of E. bifurcata (see above), but its presence in all members of the more basal taxon, Parhyposomata, still is consistent with the hypothesis that ancestors of Eulohmanniidae possessed it.
R8 — Cuticular birefringence. Grandjean (1956b) considered E. ribagai a'singular' oribatid mite in having juveniles with a hysterosomal cuticle that appeared thick and resistant to deformation. He was uncertain if there was general sclerotization in either juveniles or adults, but it seems clear that the adult cuticle differs by having a recognizable exocuticle (Fig. lF, G). Since he utilized polarized light in many observations, it is surprising that Grandjean did not mention the distinct glow from the procuticular layers of juveniles. This trait is absent from Palaeosomata and Parhyposomata, as well as Nehypochthoniidae, Epilohmanniidae and Ptyctima. But the glowing procuticle is shared by Perlohmanniidae and Collohmanniidae—taxa that Grandjean (1958a, 1966) studied in detail—and also various Nothrina and the brachypyline family Hermanniellidae (Norton and Sidorchuk 2014). In these groups the juvenile cuticle seems relatively tougher than that of a typical brachypyline juvenile, which we assume is somehow related to properties that create the glow. The three mixonomatan families also share an artifactual trait that might be related to unusual structure: the procuticle is overlain by an epicuticle that easily separates during clearing treatments prior to observation.
R9 — Supracoxal seta. Fujikawa (2014) differentiated E. ribagai from E. bifurcata partly on the shape of the supracoxal seta (spine) eI: while appearing forked in both species, the two distal tines (cilia) were described as'minute' in E. ribagai (see Grandjean 1939b, his Fig. 2, elcl) but long in E. bifurcata (at least as long as the setal stem in her Fig. 4A). None of our Nearctic specimens of E. ribagai had tines that could be described as minute, though they can appear so if broken (Fig. 8H, bottom), but some from Sweden (Fig. 8J, top) did have very short tines. It is unclear how much of this variation in the shape of eI is within populations, between populations (geographic variation), or even related to the existence of cryptic species.
R10 — Claparède's organ. This organ is considered a metameric homologue of the genital papillae in acariform mites and, like them, has an osmoregulatory function (Alberti 1979; Alberti and Coons 1999). In the large majority of oribatid mites, the structure resembles genital papillae in having a distinct, slightly enlarged distal cap supported by a simple stalk. The stalk tends to be shorter in Nothrina and Brachypylina (e.g., Grandjean 1955) than in early-derivative groups such as Palaeosomata (Grandjean 1954a) and Parhyposomata. In the latter group, which is the basal taxon of Novoribatida, the stalk may be simple (Gehypochthoniidae; Strenzke 1963), or distinctly annulated in part, as in Parhypochthoniidae (Grandjean 1934b) and Elliptochthoniidae (Ermilov and Bayartogtokh 2022). Among mixonomatans, in Perlohmanniidae the stalk is also annulated, giving the impression of having a stack of 8-15 adjacent discs (Grandjean 1958a; Suzuki 1977), while that of Eulohmanniidae is similar (Figs 3C, 8A, 19C; Grandjean 1939c) but with 4-10 annuli. The stalk is simple in Collohmanniidae (Grandjean 1966) and most examined Ptyctima (Grandjean 1934a; Lions 1966; Travé 1975). In Nehypochthoniidae the long stalk has a series of slight but distinct constrictions (Norton and Metz 1980). In Epilohmanniidae the organ is unusually long and uniquely segmented (Grandjean 1946a).
Grandjean (1946a) believed that the segmented organ of Epilohmanniidae represented a primitive structure, and that simpler forms represented regressions, but outgroup comparison suggests otherwise. Epilohmanniidae is not a basal taxon in acariform mites, and no known member of Palaeosomata, Enarthronota, or endeostigmatid mites (e.g., Terpnacaridae; Grandjean 1939a) has an organ with annulation or segmentation. In our view, since the organ is a metameric homolog of genital papillae, it makes sense that their ancestral form is rather similar, i.e., simple. Complex forms with constrictions, annuli or segmentation are derivations that allow long stalks to have controlled flexibility.
R11 — Setation of epimeres III, IV. Beginning in the deutonymph there are four pairs of setae on epimere III of both E. ribagai and P. metzi, and this does not vary in any of our material. Fujikawa (2014; her Fig. 1D) illustrated an adult Hungarian specimen of E. ribagai with four setae on one side and apparently five on the other, yet her written formula noted only three pairs, the same number cited by Balogh and Mahunka (1983) and Balogh and Balogh (1992, their Table 3.3); we believe these are documentation errors, rather than real variations. Each of these publications also reported four pairs of setae on epimere IV, but as noted above the setation of this epimere is complicated by a neotrichy that starts when the epimere first forms in the protonymph. Usually in oribatid mites there is a single pair of epimere IV setae in that instar (Grandjean 1934c), with no other seta present more posteriorly on the venter until reaching the paraproctal region. Instead, the protonymph of both E. ribagai and P. metzi has three pairs in the vicinity of legs IV, plus setae x and y behind them. In later instars, in the absence of clear epimeral borders, it seems impossible to ascribe any specific portion of the increasing neotrichy to epimere IV, but certainly there are more than four pairs in the adult. Travé (1978) thought the ventral neotrichy also involved epimere III, which we do not. However, he did recognize the problem of distinguishing epimeral from aggenital setae and abandoned an earlier (1977) specific statement that there are five pairs on epimere IV.
R12 — Genital setation of E. ribagai. The ontogeny of genital setation, 1-4-7-9 (Pn to Ad), was first documented by Grandjean (1949b, see also 1961). As he showed, it is a common development found in representatives of a diversity of groups, including some Palaeosomata, Parhyposomata, Ptyctima and Nothrina. Most literature has cited nine pairs in the adult (e.g., Bulanova-Zachvatkina 1975; Balogh and Mahunka 1983; Weigmann 2006), but Balogh and Balogh (1992) reported eight pairs (despite showing nine in the illustration), and Fujikawa (2014) gave a range of 8-10, illustrating a European specimen with 10 pairs (her Fig. 1E). We encountered no variation among female E. ribagai, regardless of provenance: all had nine pairs. But males in the population from Reindeer Station (Canada) did vary from 7-9, which would explain the symmetrical setation of seven pairs in the illustration of Hammer (1952 her Fig. 1b) that was based on a mite from this location.
R13 — Setation of aggenital region. As noted by Grandjean (1949b), the unique presence of two pairs of setae between legs IV and the paraprocts of the larva—a region that is glabrous in the larva of other oribatid mite families—creates problems in interpreting homologies. He believed the posterior seta (our y) could be the transcupular inguinal seta of row h, but this seems unlikely as inguinal setae (by definition: Grandjean 1949b) disappear, usually in the subsequent instar. By contrast, y clearly persists in the protonymph of both E. ribagai and P. metzi (Figs 4A, 26D). He thought the more anterior (our x) was likely a secondary (i.e., neotrichous) seta. He further suggested (p. 224) that aggenital setae might not exist in E. ribagai. They are indeed absent throughout ontogeny in numerous genera of Enarthronota and Nothrina, among others, though two pairs are typical of the tritonymph and adult of mixonomatans (e.g., Grandjean op. cit., p. 222). In no oribatid mite is an aggenital seta known to appear prior to the deutonymph.
Grandjean (1949b) seems to have considered the large expanse of cuticle between legs IV and the genital aperture in nymphs and adult to be a hypertrophication of epimere IV, calling its rich setation a'néotrichie coxisternale'. By contrast, Fujikawa (2014) identified the neotrichy as aggenital. Instead, we favor a hypothesis that both epimere IV and the aggenital region contribute to the expansive neotrichous ventral plate in Eulohmanniidae. Neotrichy of epimere IV clearly starts in the protonymph (R11) and there is evidence that aggenital setae or their derivatives are part of the more posterior setation. In adult eulohmanniids, at least two or three setae on each side of the genital plate occupy the same space as aggenital setae in mixonomatans such as Perlohmannia and Collohmannia (Grandjean 1958a; Norton and Sidorchuk 2014). An even more significant comparison is with the mixonomatan family Epilohmanniidae, in which the posterior border of epimere IV is clearly defined: aggenital plates are strongly hypertrophied and medially fused anterior to the genital aperture (e.g., Schuster 1960; Bayartogtokh 2000). Just as some neotrichous epimere IV setae form in the protonymph, we envision setae x and y as precocious aggenital setae or their neotrichous derivatives.
Regardless of the source of the neotrichous setae, we cannot confidently identify any particular number as aggenital seta, and the specific attribution of six pairs to Eulohmannia by Balogh and Balogh (1992; p. 35) or 14 pairs for E. bifurcata by Fujikawa (2014) seems arbitrary.
R14 — Paraproctal setation. The ontogeny of setation on paraproctal valves is rather simple and consistent in our material of E. ribagai (regardless of provenance) and P. metzi. Segment PS has four setae in the larva; following Grandjean's (1949b) concept, the most anterior is an inguinal seta, psi (p4), which is lost when the segment joins the gastronotum in the protonymph. Such inguinal setae are common in early derivative oribatid mites and endeostigmatids (Grandjean 1942b, 1949b; Norton and Behan-Pelletier 2007 [their note 8]; Norton and Fuangarworn 2015 [their remark 11]). Segment AD appears in the protonymph with its full complement of four pairs of setae, and in E. ribagai (but not P. metzi) segment AN appears in the deutonymph, also with four pairs. The setal complements of AD and AN do not change in subsequent instars.
But according to Grandjean's (op. cit. p. 206) analysis, the ontogeny of both segments PS and AD is highly unusual in E. ribagai. Using a shorthand formula, he presented the setal ontogeny as 3–3444–444, which can be interpreted as follows. Segment PS (the first number) has three pairs of setae in the larva, but he was uncertain of the number in subsequent instars, where segment PS moves to the gastronotum; segment AD has three pairs when it appears in the protonymph, but four in subsequent instars; segment AN has four pairs when it appears in the deutonymph and retains this number. Except in taxa with neotrichy or paraproctal atrichosy, in which segments are glabrous when they first appear (Grandjean op. cit. p. 207), it is highly unusual for a segment to add setae after it forms. He cited only two examples of such delay in segment AD setae: E. ribagai and Torpacarus omittens (Lohmanniidae). As discussed above (justification of Paedolohmannia n. gen.) we feel he was wrong about T. omittens: a more parsimonious explanation is that the seta added in the deutonymph is an anal seta.
We also believe Grandjean proposed an incorrect ontogenetic formula for E. ribagai because he overlooked seta ps1 in the larva of E. ribagai, as well as seta ad1 in the protonymph. This might seem unlikely, considering that he was a renowned observer, and that in our material these are the largest setae on their respective segments (Figs 3C, D, 4A). But both these enlarged setae are highly prone to breakage during handling and clearing treatments; they are missing from many of our prepared specimens while the three smaller, more anterior setae of the respective segment are always intact. The large alveolus of a broken ps1 or ad1 can be found if searched for, but it is surprisingly inconspicuous: being close to the lateral edge and near the posterior end of the segment, it becomes almost vertically oriented in usual dorsoventral orientations and is covered by the edge of the gastronotum in lateral orientation. Both ps1 and ad1 (or the alveolus if broken) always are present in our abundant Nearctic material.
Grandjean did not mention the number of larvae or protonymphs he observed, but we studied one protonymph from Austria (Fig. 11C) and it is consistent with our Nearctic material: on one side the enlarged ad1 is present, but on the other only the large alveolus remains. Also consistent are three protonymphs from Germany (Saxony and Berlin states) in the Senckenberg Museum für Naturkunde Görlitz studied by Dr. Ricarda Lehmitz (pers. comm. 2021): one had the large pair ad1 present, the other two showed only their large alveoli. She also studied two larvae from the Czech Republic (Krkonoše Mountains, Šumava Mountains) and while setae ps1 were broken, their large alveoli were present. Similarly, a larva from Norway (near Kumra, Nordland province) studied by Dr. Anna Seniczak (pers. comm. 2022) had the alveoli of ps1 though the setae were lost.
R15 — Vestigial seta d on genua I-III. As part of a benchmark series of papers on the comparative chaetotaxy of oribatid mite legs, Grandjean (1942a) identified eight different'behaviors' of genu seta d during ontogeny. Seven of these relate to patterns in which the seta is somehow diminished in size or suppressed in one or more instars. A pattern he called'larval suppression' was considered unique to E. ribagai at that time, but it is shared by Eulohmannia sp. A and Paedolohmannia metzi. In each instance seta d is represented in the larva by a minute setal vestige inserted in a small pore-like alveolus (Figs 12B, 15G, 26A-C), but a normal seta forms in subsequent instars. A similar development occurs in Perlohmannia (Grandjean 1958A; Suzuki 1977) and Nehypochthonius (new observation; overlooked in original description, cf. Norton and Metz 1980).
This vestige easily might be confused with the so-called'genual pore' (Grandjean 1940) that appears on these same segments in some Nothrina and Mixonomata (but not Eulohmanniidae). By contrast, this pore lacks an internal vestige, exists in all instars, and is always clearly independent of seta d (e.g., Grandjean 1954b; Norton and Sidorchuk 2014).
R16 — Leg IV setation in protonymph. Leg IV of acariform mites forms in the protonymph with a diminished chaetome. Grandjean (1946b) noted many variations, but among oribatid mites the typical protonymphal setation includes seven setae on tarsus I, with all other segments being glabrous. The protonymph of E. ribagai and P. metzi have the usual tarsal setation, but tibia IV consistently forms seta vʺ. It is common, and perhaps plesiomorphic for vʺ to be retained on protonymphal tibia IV in Enarthronota and it forms in large species of Gehypochthoniidae (Parhyposomata) (Norton and Fuangarworn 2015), but these are the only taxa that share this trait, to our knowledge.
R17 — Setal pair (m) on tarsus I. The'poil monotrope', mʺ, is a eustasic larval seta of tarsus I located slightly distal to pvʺ in some early-derivative groups of oribatid mites (Grandjean 1962). It is common in Parhyposomata and Enarthronota (see also Norton and Fuangarworn 2015, their Remark #17) but we are not aware of any mixonomatan other than Eulohmanniidae that has this plesiomorphic seta, which was first reported in E. ribagai by Grandjean (1941b).
Seta mʹ, the pseudosymmetrical complement of mʺ, is less common and always developmental—deutonymphal in Eulohmanniidae (Figs 15I, 24D). We refer to pair (m) as monotropic setae even though the application is grammatically incongruous: derived from the Greek mono (one) and tropos (turning, direction), the name made sense originally because of the asymmetry of mʺ. Seta mʹ was not confidently identified in any oribatid mite until Grandjean (1962) became convinced that a developmental seta could appear distal to fundamental seta pvʹ, instead of proximal to it like a typical proximal accessory setae. The presence of mʹ in Eulohmanniidae is noted here for the first time. The pair (m) has a strong anterior (ʹ) disjunction, which also is true of Gehypochthonius xarifae Strenzke, 1963 (Parhyposomata); although publication delay made the chronology confusing, Grandjean (1962) considered the seta labeled pvʹ by Strenzke (1963, his Fig. 11) to be instead mʹ. This disjunction conforms to the usual anterior offset of the primiventral pair (pv) on tarsus I of oribatid mites (Grandjean 1958b, 1960), although in Eulohmanniidae (pv) have little or no disjunction. By contrast, in most of those Enarthronota that develop both mʹ and mʺ the pair has a posterior disjunction (Norton and Fuangarworn 2015).
The fate of pair (m) during development in Eulohmanniidae—mʺ remaining throughout ontogeny but mʹ becoming eupathidial—appears to be unique. The opposite occurs in several Enarthronota (Norton and Fuangarworn 2015), where the more distal mʺ becomes eupathidial and mʹ remains normal. In Grandjean's (1962) interpretation, both mʺ and mʹ become eupathidial in G. xarifae, in the tritonymph and adult, respectively.
R18 — Iteral setae. Among oribatid mites the ontogeny of iteral setae (it), which are always developmental, varies greatly according to taxon, leg, and even between members of the pseudosymmetrical pair. As summarized by Grandjean (1964a) their ontogeny in E. ribagai is unique, with each leg being different: the development shown in Table 4 is summarized by his formula (n1—[n3, n2]—n3—o). In the context of his conceptual model of regressive evolution in these setae, the early appearance of (it) on tarsus I places it among the more plesiomorphic ontogenies known. The transformation of (it) on tarsus I to eupathidia is shared by a wide range of early-derivative oribatid mites, including members of Palaeosomata, Enarthronota, Parhyposomata, and some other families of Mixonomata.
R19 — Primilateral setae. In the conceptual model of F. Grandjean, primilateral setae are fundamental (present when the leg in question is first formed) and eustasic (formed at that time, or not at all). In a review of their distribution, Grandjean (1959) first reported the complement of primilaterals in E. ribagai—pair (pl) on tarsus I, but only plʹ on tarsus II, with none on III or IV—and noted that it was shared only by the enarthronote genus Eniochthonius (see also Norton and Behan-Pelletier 2007). Unlike Eniochthonius, the tritonymph of E. ribagai adds a seta on the posterior face of tarsus II, in the position typically occupied by accessory seta lʺ if that seta exists. But, as explained below, we believe this seta is not lʺ but instead is the homologue of plʺ that has become delayed (Table 4, Fig. 14B). As Grandjean (op. cit.) indicated, one can distinguish primilateral from accessory l-setae in the adult, even without knowing ontogeny, because both follow rules.
Three rules relevant to proximal (`posterior') accessory setae were developed by Grandjean (1958b) in one of his most insightful papers. (1) They are eustasic, with each potential verticil forming in a specific instar: this concept was developed mostly from studies of Nothrina, but the rule seems to apply also to mites of the mixonomatan families Perlohmanniidae (Grandjean 1958a) and Collohmanniidae (Norton and Sidorchuk 2014), which have many accessory setae. (2) They are subject to regression during evolution, but the regression has segmental directionality, such that in evolutionary time setal losses occur first from more posterior legs; i.e., the'strength' (resistance to regression) of a given seta decreases from tarsus I to IV. (3) Regression also has a reverse ontogenetic directionality: the later a particular eustasic seta forms, the more resistant it is to evolutionary loss.
Unlike Perlohmanniidae and Collohmanniidae, species of Eulohmanniidae have few accessory setae; in fact, there are no more than four in E. ribagai—three in the c-row on tarsus I (see below) and possibly the seta in the lʺ position on tarsus II. But the latter seta fails the tests for a lateral accessory seta: it is present on tarsus II yet is absent from tarsus I (which has only the primilateral pair in the lateral position), and it forms in the tritonymph rather than the adult. According to Grandjean's rules, in a regressive trend over evolutionary time the very last, non-regressed accessory l-seta should be formed on tarsus I of the adult.
While highly explanatory, there is increasing evidence that these rules are not inviolate. Norton and Fuangarworn (2015, see their Remark 18; also Norton and Sidorchuk 2014, their Remark 7) explained that primilateral setae may have become delayed (developmental, or'amphistasic') on the posterior two legs of some Enarthronota. A major factor in this interpretation is that accessory lateral (and also ventral) setae are almost unknown in Enarthronota. Since the posterior lateral seta on tarsus II of Eulohmanniidae disobeys rules for accessory seta lʺ, and has a position exactly appropriate for plʺ, the most parsimonious explanation is that plʺ has become ontogenetically delayed over evolutionary time, i.e., it has become amphistasic. Grandjean (1962) himself used a similar argument when identifying mʹ, the amphistasic anterior seta of the monotropic pair.
To have a single accessory seta on the posterior face of tarsus II would also contradict another widespread pattern, which relates to the priority of setae in a pseudosymmetrical pair. In general, on non-tarsal segments of oribatid mites anterior seta lʹ has priority over lʺ (it is more resistant to regression): i.e., lʺ should not be present unless lʹ also is present. Among mixonomatans this priority also is true of proximal accessory setae on tarsi I and II, e.g., in Perlohmanniidae (Grandjean 1958a) and Nehypochthoniidae (Norton and Metz 1980). In primilateral setae the anterior seta, plʹ, also has priority (Grandjean 1959) so if one member becomes amphistasic it should be plʺ.
R20 — Setae of the c-row. These setae of tarsus I appear to be rare outside Palaeosomata (e.g., Grandjean 1952a, 1954c); to our knowledge they occur only in three mixonomatan families—Perlohmanniidae, Collohmanniidae, and Eulohmanniidae. They are proximal accessory (post-larval) setae that lie at a level between v- and l-row setae when all three rows are present. Accessory setae of the l- and v-rows respectively form proximal to the fundamental primilateral (pl) and primiventral (pv) setae, whereas those of the c-row seem to align with the fundamental antelateral pair (a). In Perlohmanniidae (Grandjean 1958a) and Collohmanniidae (Norton and Sidorchuk 2014) setae of the c-row appear to follow Grandjean's rules for accessory setae (see R19) precisely, though they are not present in the nothrine taxa he used to initially formulate the rules (Grandjean 1958b). In these two families a pair of c-setae are added in each of the last three instars, with the respective notations (cn2), (cn3) and (cA).
In Eulohmanniidae only one series of proximal accessory setae forms. We identify them as c-setae because in their normal (non-eupathidial) form they align with (a) and because no accessory setae appear proximal to either (pl) or (pv)—i.e., they are not l- or v-setae. If either a c- or a-seta transforms to a eupathidium, alignment can be affected since setae tend to be slightly displaced distally and/or ventrally after transformation. We know of no other oribatid mites in which c-setae appear in the absence of other accessory setae. But the c-setae of Eulohmanniidae are unique, to our knowledge, in not precisely following Grandjean's rules (Table 5). Both Paedolohmannia metzi and Eulohmannia sp. A (Aborigen) have the same ontogeny in the c-row: a pair (cn2) forms in the deutonymph, as expected, and these have a null or slight posterior (ʺ) disjunction (Fig. 24D), but then a single posterior seta is added in the tritonymph and a single anterior seta added in the adult. If the latter two setae are eustasic, they would be cn3ʺ and cAʹ, respectively, which are the notations we apply herein. By contrast, E. ribagai forms a single c-seta in each of the last three instars, differing from the other two species in lacking seta cn2ʺ. We have seen no juveniles of Eulohmannia sp. B (Kashmir), but the adult has a single c-seta on each face, which in the other three species occurs only in nymphs; we interpret this as another manifestation of its paedomorphic tarsus.
While Grandjean's second rule of accessory setae applies to c-setae in Eulohmanniidae (they are present only on tarsus I), his third does not: setae remain in the presumed deutonymphal and tritonymphal verticils even though the adult verticil is partially lost. Also, the priority of setae seems to vary among the verticils, with the posterior (ʺ) seta being lost in the deutonymphal and adult verticils in E. ribagai, as expected from general priorities (see R19), whereas the tritonymphal verticil has lost the anterior seta (ʹ). We have no explanation of these observed differences if the setae are eustasic.
An alternative hypothesis is that some c-setae instead have become amphistasic. In such a case, c-seta homologies could not be established based on the instar of appearance. For example, if the tritonymphal seta denoted cn3ʺ in E. ribagai (Fig. 15J) were truly from the ancestral tritonymphal verticil (eustasy hypothesis), one would not expect it to have a more distal position than cn2ʹ from the deutonymphal verticil. However, if the purported cn3ʺ is actually a delayed cn2ʺ (amphistasy hypothesis), then the positions of the two c-setae in the tritonymph would be exactly those of pair (cn2) in the deutonymph of P. metzi and Eulohmannia sp. A (cf. Figs 15J, 24D). At the moment, neither hypothesis seems stronger than the other.
R21 — Tarsus I eupathidia. Eulohmanniidae have a moderate number—9, 10 or 11—of these probable gustatory chemosensilla (Alberti 1998) on tarsus I. This is far fewer than Perlohmanniidae (Grandjean 1958a) or Collohmanniidae (Grandjean 1966; Norton and Sidorchuk 2014), both of which have eupathidia among the abundant proximal accessory setae. Each of the three species for which we have developmental data has a slightly different adult complement and ontogeny of eupathidia (Table 5). We have no juveniles of the paedomorphic Eulohmannia sp. B (Kashmir) but based on a single studied adult there are nine, including (p), (u), (it), s, mʹ and one abaxial c-seta (not yet identified). Eulohmanniidae are almost unique among oribatid mites in having pair (u) eupathidial—according to Grandjean (1958a) this is shared only by some (unnamed) Damaeidae (`Belbidae').
Most ontogenetic transformations are constant: setae (p) are eupathidial from the larva and each species transforms s and uʹ in the protonymph, (it) in the deutonymph, and mʹ in the tritonymph. If seta cn2ʺ is present (Eulohmannia sp. A, P. metzi), it transforms in the tritonymph. We identified only one variation: the transformation of uʺ is either protonymphal, like uʹ, or delayed to the deutonymph (P. metzi).
Transformation of the antelateral setae is most diverse and lacks any recognizable pattern. Both Eulohmannia sp. A and E. ribagai transform aʹ and aʺ, but with reversed ontogenetic priority; P. metzi transforms aʺ surprisingly early (Pn) but aʹ remains normal throughout ontogeny; in Eulohmannia sp. B neither a-seta transforms. We observed few intraspecific variations in eupathidial transformation, and all related to antelateral setae. In Eulohmannia sp. A, aʺ remained normal on one of four deutonymphal legs examined. In E. ribagai seta aʹ transforms in the deutonymph, except on one of 10 tarsi examined from the New York population it remained normal. The transformation of aʺ in E. ribagai is more variable, perhaps differing among populations (but there are few observations). In three tritonymphs from New York and one from Germany, only aʹ was eupathidial, suggesting that aʺ would have transformed in the adult. In another tritonymph from New York, two from Austria (near the type locality), and two from Canada (Cape Breton and Reindeer Station) aʺ had transformed, so both (a) were eupathidial. While variable among specimens, in each instance the state of aʺ was symmetrical in a given mite.
Eulohmanniidae may be unique regarding eupathidial transformation in the monotropic pair (m). In some in Enarthronota (see Norton and Fuangarworn 2015) mʺ becomes eupathidial while mʹ remains normal, and in Gehypochthonius xarifae (Parhyposomata) both mʹ and mʺ become eupathidial, with mʺ transforming first (Grandjean 1962). But we are unaware of any species outside Eulohmanniidae in which mʹ transforms to a eupathidium while mʺ remains normal.
R22 — Complement of solenidia. Eulohmanniidae have a relatively rich complement of solenidia (15 per side), with a development pattern widely shared among early-derivative oribatid mites (Grandjean 1964c). Like most Parhyposomata, Mixonomata and Enarthronota, they have a single (larval) solenidion on tibia I, having lost the developmental ϕ2 that typifies most taxa in the more highly derived Nothrina and Brachypylina. An opposite pattern is seen on genu I, where a developmental solenidion is added to the two larval solenidia. This is known elsewhere only in Palaeacaridae (Palaeosomata; several other palaeosomatans with three solenidia have unknown development) and Nanohystricidae (Enarthronota); in these instances, the third solenidion is tritonymphal or (in Nanohystrix) possibly deutonymphal (Grandjean 1964c; Norton and Fuangarworn 2015), whereas that of Eulohmanniidae appears in the protonymph (Grandjean 1942a). If regression has shaped the solenidial complements of oribatid mites, then that of Eulohmanniidae is the least regressive known for genu I.
R23 — Coupling of solenidia. In the large majority of oribatid mites, seta d is coupled to a solenidion on at least some genua and tibiae or shows evidence that this was true in their ancestors (Grandjean 1942a). In Eulohmanniidae seta d is distant from any solenidion on these segments and is one of the few families outside Palaeosomata and Parhyposomata in which this plesiomorphic separation persists. Among mixonomatans Perlohmanniidae is the only other family lacking such coupling entirely.
However, in all instars seta lʹ of genu I is coupled to solenidion σʹ. This same association is found in the early-derivative mixonomatan family Nehypochthoniidae, in which there are no d-solenidion couplings on leg I, though they exist on other legs (Norton and Metz 1980). An lʹσ coupling also occurs on genu I of Ptyctima, both Phthiracaroidea and Euphthiracaroidea (e.g., Grandjean op. cit.; Hammen 1963; Lions and Norton 1998), in which d-solenidion couplings do exist. We are not aware of lʹ-solenidion couplings on genu I outside Mixonomata, but on genu II of Mucronothrus (Nothrina) a weak coupling develops during ontogeny (Norton et al. 1996).
R24 — Pretarsal ambulacrum. Grandjean (1969) considered the ancestral adult oribatid mite ambulacrum to be tridactylous, with strong empodial and paired lateral claws. He viewed the combination of strong lateral claws and a regressed central (empodial) claw—the'Acaronychus-type' of tridactyly (herein type-A)—as an'exceptional secondary character', shared among members of three of his major groups: Palaeosomata (Archeonothroidea), Parhyposomata (Parhypochthonius) and Mixonomata (Eulohmannia). To these we can now add Elliptochthoniidae (Norton 1975), Nehypochthoniidae (Norton and Metz 1980) and some Gehypochthoniidae (Strenzke 1963, Martinez and Bernava Laborde 2000). Grandjean considered a bidactylous ambulacrum, with the absence of the empodial claw (herein type-B), as a different'exceptional character', found in some primitive Enarthronota—Atopochthoniidae and Phyllochthoniidae (and subsequently in Arborichthoniidae; Norton 1982)—plus some unnamed Gehypochthonius species (probably G. rhadamanthus Jacot, which he had studied previously; Grandjean 1943). At least in Gehypochthoniidae, it seems clear that the type-B ambulacrum represents the end of a regressive trend, since the reduced empodium is either present or absent, according to species. With one exception, all Eulohmanniidae discussed herein have a type-A ambulacrum: the purported absence of an empodial claw in E. bifurcata is an error, as discussed above. Only in Eulohmannia sp. B (Kashmir) is the empodial claw large, and this seems a clear example of neoteny (see above).
In our view, type-A tridactyly was ancestral in Novoribatida, since it is present in all known genera and most known species of Parhyposomata; therefore, we consider the type-A ambulacrum in the mixonomatan families Nehypochthoniidae and Eulohmanniidae as plesiomorphic, rather than secondary. While there are no known extant Enarthronota with a type-A ambulacrum, it existed in the extinct family Protochthoniidae (Norton et al. 1988), so a similar regressive loss may have resulted in the type-B ambulacrum of the enarthronote families noted above. Since the type-A ambulacrum also characterizes Archeonothroidea, which we consider the most basal group of Palaeosomata (Norton and Fuangarworn 2015), it may be ancestral in Sarcoptiformes.
Among mixonomatans, Perlohmanniidae and Epilohmanniidae adults are monodactylous, suggesting paedomorphosis at some point in their evolution, like that of Eulohmannia sp. B. Collohmanniidae are homotridactylous (Norton and Sidorchuk 2014) and Ptyctima are rather plastic in ambulacral structure (e.g., Lions 1964; Lions and Norton 1998), but in no instance is the empodium reduced. Unlike Eulohmanniidae, these other mixonomatans have ambulacra that collectively mirror those of Nothrina and Brachypylina. If the type-A ambulacrum is ancestral in oribatid mites, these more widespread adult forms could have evolved from it by the fixation of a large, paedomorphic empodial claw, while lateral claws either remained large or became susceptible to regression.
R25 — Rutellar comb (= brush, cilary comb). In Nothrina and Brachypylina, the dorsal face of the rutellum usually bears a comb comprising one (usually) or two rows of thin'cilia' that presumably act to clean small particles from the lateral (abaxial) face of the chelicera (Grandjean 1957, Hammen 1968). The comb is secondarily absent—a loss—from many Brachypylina with specialized mouthparts, and probably so in most Astigmata (Norton 1998). Palaeosomata, Enarthronota, and Parhyposomata also lack the comb, but this is an ancestral state.
In mixonomatans—none of which have significantly specialized mouthparts—the comb can be well developed, as in Eulohmanniidae, or have structures that may be homologs of the comb. Such a distribution suggests that the comb evolved within Mixonomata. Grandjean (1958a) considered irregular dorsal ridges on the rutellum of Perlohmannia to represent a reduction (i.e., a loss) of the comb's cilia, but this seems unlikely. If the comb functions by removing debris, what advantage would be gained by their loss from an unspecialized rutellum? The opposite trend seems more likely: a rutellum dorsally roughened by ridges (seen also in Nehypochthoniidae and Epilohmanniidae) represents the primitive form of a cleaning surface, with comb-like projections from the ridges having evolved subsequently.
In Collohmanniidae the rutellum has dorsal ridges that lack cilia, but there are 1-3 (usually two) small spines (Norton and Sidorchuk 2014). Grandjean (1966) thought these spines represented a special form of the comb; if so, it seems an intermediate form, rather than representing the culmination of a trend. Eulohmanniidae have a comb (Fig. 16K; seen by transparency in Figs 10A, C, 21C) as fully developed as those in Nothrina and Brachypylina, so it seems the most highly-derived form among mixonomatans.
R26 — Palp segmentation. The plesiomorphic state in oribatid mites is to retain full independence of the five ancestral articulating segments: trochanter (Tr), femur (Fe), genu (Ge), tibia (Ti) and (Ta) tarsus. All Palaeosomata have this state, but fusions between segments are known in some families of all other major groups. Among mixonomatans, Nehypochthoniidae, Perlohmanniidae and Collohmanniidae retain the plesiomorphic state. Eulohmanniidae have a complete Fe-Ge fusion (misidentified as Tr-Fe by Haumann 1991 and Weigmann 2006) in all instars, with (Paedolohmannia) or without (Eulohmannia) a crease-like vestige of the articulation. In Epilohmanniidae there usually are two apparent segments (Tr-Fe-Ge-Ti fusion) but in some there are seemingly three, as the tibia is only partly fused. Most diverse are the Ptyctima, with three, four or five functional segments (Grandjean 1954a; Hammen 1963; Märkel 1964; Lions and Norton 1998).
The Fe-Ge fusion found in Eulohmanniidae seems the most common type: it exists in all the mixonomatan examples mentioned above in which the palp has either four or three segments. It is also the fusion present in those few Enarthronota with fewer than five palpal segments (Norton et al. 1983). Elliptochthoniidae (Parhyposomata) is a rare exception, in which the single fusion is clearly Tr-Fe. [Haumann 1991 considered Phthiracaridae to have four segments with a Tr-Fe fusion, but this was an error; see Grandjean 1954a; Hammen 1963]. Since the Fe-Ge fusion clearly has evolved independently multiple times, it probably is the optimal location to limit movement, if a less articulated appendage is somehow beneficial.
R27 — Cheliceral internalization and emargination. Chelicerae of acariform mites ancestrally attach to the soft cheliceral frame in a simple fashion, such that the entire appendage extends from the body wall. This is true of Palaeosomata and Parhyposomata, but in Nothrina and Brachypylina (also Astigmata; Norton 1998) the attachment has encroached onto the cheliceral body, such that about the proximal third of the chelicera is internalized (inserted), while the regressed trochanter remains external (Grandjean 1947, his Fig. 2A; Norton and Sidorchuk 2014, their Fig. 11A, B). In most mixonomatans there is no, or very little insertion, but in Collohmanniidae and Eulohmanniidae it is conspicuous.
Another trait seems to correlate well with such insertion: the proximal half of an inserted chelicera is broadly emarginated ventrolaterally on the adaxial face. No functional studies have addressed this, but the combination of insertion and emargination may allow more varied directionality of retractor muscles and therefore cheliceral movements. Another correlation is with Trägårdh's organ, which seems to be well-developed only in groups where the chelicera is internalized and emarginated (R28).
R28 — Trägårdh's organ. This is an elongated, tapering, finger-like oncophysis that evaginates and extends distad from the soft cuticle on the emarginated adaxial face of the chelicera, at the juncture of the cheliceral frame with the trochanter and principal cheliceral segment (Grandjean 1959; Hammen 1968; Alberti et al. 2011). A fully developed Trägårdh's organ is present in all known Nothrina (except Malaconothridae) and Brachypylina. Grandjean (1959) thought the absence of the organ in Malaconothridae represented a loss, presumably because the family is otherwise clearly a member of'Nothroidea' (in his sense) and Norton (1998) considered their similar absence in Astigmata to be a regressive synapomorphy shared with that family.
Fujikawa (2014) purportedly found Trägårdh's organ in E. bifurcata, but we failed to find it in the holotype (see above) and doubt its existence in any Eulohmanniidae. Among Mixonomata, a fully developed Trägårdh's organ is known only in Collohmanniidae (Norton and Sidorchuk 2014), a family that some molecular studies inferred is closely related to, or even within, Nothrina (e.g. Xue et al. 2017; Pachl et al. 2020). Smaller, less conspicuous, and delicate structures having a rather similar position are found in some Ptyctima (reviewed by Lions and Norton 1998); these structures remain poorly known and whether they represent early, plesiomorphic forms of the organ or independently derived structures remains equivocal. In all questionable cases, the chelicera is neither inserted nor adaxially emarginated (R27), conditions that otherwise correlate well with Trägårdh's organ. The fact that the chelicera of Eulohmanniidae is both inserted and emarginated leaves open the possibility that Trägårdh's organ was lost, as in Malaconothridae.
Lohmanniidae appear to be the only family of Enarthronota to possess a structure that has been referred to as Trägårdh's organ (Ermilov 2017; Ren et al. 2017). Grandjean (1950b) first noticed an elongated ridge on the adaxial face of the chelicera in this family, and this ridge ends in a slightly elevated'boss'. He dismissed it as a possible Trägårdh's organ, though he reserved judgement on whether there was some degree of homology. Since it is not a hollow, flexible oncophysis and does not evaginate from the juncture of the chelicera with the cheliceral frame, we believe this ridge on the wall of the chelicera is an independently evolved structure and should not be identified as Trägårdh's organ.
R29 — Thelytoky. Usually referred to by the misapplied term parthenogenesis, thelytoky has been the reproductive mode associated with Eulohmanniidae in the literature, which is understandable since our limited knowledge has been associated exclusively with E. ribagai. But given newly discovered sexual populations of E. ribagai, the probably sexual Eulohmannia sp. A (Aborigen; see above), and the sexual type species of Paedolohmannia, the ancestral reproductive mode of Eulohmanniidae should be reconsidered. Other extant mixonomatans are mostly sexual (Norton et al. 1993): it is the only known mode in Perlohmanniidae and Collohmanniidae and it dominates in Ptyctima (Euphthiracaroidea, Phthiracaroidea). Although Epilohmanniidae include some common thelytokes, probably most species are sexual. Nehypochthoniidae, a monogeneric and phylogenetically relictual taxon, is now the only mixonomatan family that seems to be wholly thelytokous. Outside Mixonomata, a reversal from thelytoky to sexuality was suggested for a group in the nothrine family Crotoniidae (Domes et al. 2007), and two others in Enarthronota (Pachl et al. 2021) but these are diverse groups and there is both morphological and molecular evidence for reversal. Since Eulohmanniidae is morphologically isolated and species-poor, we have no convincing way to test for reversal within it, but the most parsimonious explanation is that Eulohmanniidae was ancestrally sexual and this mode remains in species and populations from northwestern North America and northeastern Asia, where further diversity should be sought.
Acknowledgements
We are grateful for the kind help of many individuals who contributed to various parts of this work. Dr. Satoshi Shimano (Hosei University, Tokyo, Japan) translated relevant parts of papers written in Japanese and facilitated our study of type specimens of Eulohmannia bifurcata. Alexey A. Gubin (Tyumen State University, Tyumen, Russia) produced most of the scanning electron micrographs. Important loans or donations of materials were provided by Drs. Valerie Behan-Pelletier and Wayne Knee (Agriculture Canada, Ottawa); Dr. Derek S. Sikes (University of Alaska Museum, Fairbanks); Dr. Ken-ichi Okumura (National Museum of Nature and Science, Tokyo); Dr. Heinrich Schatz (Leopold-Franzens University of Innsbruck, Austria); and Dr. Ricarda Lehmitz (Senckenberg Museum, Görlitz, Germany). Drs. Lehmitz and Anna Seniczak (University Museum of Bergen, Norway) also made important observations of juvenile E. ribagai from Germany and Norway, at our request. Robin Andrews (University of Alaska, Fairbanks) collected important material from Alaska and helped us interpret glacial refugia. Dr. Jun Chen (Institute of Zoology, Chinese Academy of Sciences, Beijing) clarified Chinese distribution records. We are particularly grateful for constructive comments by Dr. V. Behan-Pelletier on a draft manuscript and for the helpful review by self-identified referee Dr. H. Schatz. This research was partially supported by a grant from the Russian Foundation for Basic Research, project No. 20-54-56054.
References
- Akimov I.A., Yastrebstov A.V. 1991. Skeletal-muscular system of oribatid mites (Acariformes: Oribatida). Zool. Jb. Anat., 121: 359-379.
- Akrami M.A. 2015. An annotated checklist of oribatid mites (Acari: Oribatida) of Iran. Zootaxa, 3963: 451-501. https://doi.org/10.11646/zootaxa.3963.4.1
- Alberti G. 1979. Fine structure and probable function of genital papillae and Claparede organs of Actinotricha. In: Rodriguez J.G. (Ed.). Recent Advances in Acarology, vol. 2. New York: Acad. Press, Inc. p. 501-507. https://doi.org/10.1016/B978-0-12-592202-9.50072-0
- Alberti G. 1998. Fine structure of receptor organs in oribatid mites (Acari). In: Ebermann E. (Ed.). Arthropod Biology: Contributions to Morphology, Ecology and Systematics. Biosystematics and Ecology, Series 14. Öster. Akad. Wiss. p. 27-77.
- Alberti G., Coons B. 1999. Acari - Mites. In: Harrison F.W., Foelix R.F. (Eds). Microscopic anatomy of invertebrates. Volume 8C: Chelicerate Arthropoda. New York: Wiley-Liss. p. 515-1265.
- Alberti G., Storch V., Renner H. 1981. Über den feinstrukturellen Aufbau der Milbencuticula Acari (Arachnida). Zool. Jahrb. Anat., 105: 183-236.
- Alberti G., Norton R.A., Adis J., Fernandez N.A., Franklin E.N., Kratzmann M., Moreno A.I., Weigmann G., Woas S. 1997. Porose integumental organs of oribatid mites (Acari, Oribatida). 2. Fine structure. Zoologica, Stuttgart, 146: 33-114.
- Alberti G., Heethoff M., Norton R.A., Schmelzle S., Seniczak A., Seniczak S. 2011. Fine structure of the gnathosoma of Archegozetes longisetus Aoki (Acari: Oribatida, Trhypochthoniidae). J. Morph., 272: 1025-1079. https://doi.org/10.1002/jmor.10971
- Aoki J. 1959. Record of oribatid mites from Japan. Jap. J. Sanit. Zool., 10: 127-135. https://doi.org/10.7601/mez.10.127
- Aoki J. 1975. Taxonomic notes on some little known oribatid mites of Japan. Bull. Nat. Sci. Mus., Ser. A (Zool.), Tokyo, 1: 57-65.
- Arribas P., Andujar C., Moraza M.L., Linard B., Emerson B.C., Vogler A.P. 2019. Mitochondrial metagenomics reveals the ancient origin and phylodiversity of soil mites and provides a phylogeny of the Acari. Mol. Biol. Evol., 37: 683-694. https://doi.org/10.1093/molbev/msz255
- Ayyildiz N., Subias L.S., Baran S. 2016. Review of the family Perlohmanniidae (Acari: Oribatida) with description of a new species from Turkey. Biologia, 71: 323-327. https://doi.org/10.1515/biolog-2016-0034
- Baker E.W., Wharton G.W. 1952. An Introduction to Acarology. New York: The MacMillan Co. pp. 465.
- Baker E.W., Cunliffe F., Woolley T.A., Yunker C.E. 1958. Guide to the families of mites, with contributions by E. Crabill jr. and G. Nunes. Maryland: Contrib. Inst. Acarol., Col. Park, Nr. 3. pp. 242.
- Balogh J. 1943. Magyarország Páncélosatkái (Conspectus Oribateorum Hungariae). Mat. Term. Közl., 39: 1-202.
- Balogh J. 1961. Identification keys of world oribatid (Acari) families and genera. Acta Zool. Acad. Sci. Hung., 7: 243-344.
- Balogh J. 1965. A synopsis of the world oribatid (Acari) genera. Acta Zool. Acad. Sci. Hung., 11: 5-99.
- Balogh J. 1972. The oribatid genera of the world. Budapest: Akad. Kiadó. pp. 188.
- Balogh J., Balogh P. 1992. The oribatid mites genera of the world. Budapest: Hung. Nat. Mus. Press. pp. 263 (vol. 1), pp. 375 (vol. 2).
- Balogh J., Mahunka S. 1983. Primitive Oribatids of the Palaearctic Region. Budapest: Akad. Kiado. pp. 372.
- Bar I.D., Clark, C.D. 2012. Late Quaternary glaciations in Far NE Russia; combining moraines, topography and chronology to assess regional and global glaciation synchrony. Quat. Sci. Rev. 53: 72-87. https://doi.org/10.1016/j.quascirev.2012.08.004
- Bashkirova E.J. 1958. Oribatid fauna in the virgin steppe of the South-east of the European part of the USSR. Zool. Zh., 37: 193-209.
- Bayartogtokh B. 2000. Oribatid mites of the genus Epilohmannia (Acari: Oribatida: Epilohmanniidae) from Japan and Mongolia. Syst. Appl. Acarol., 5: 187-206. https://doi.org/10.11158/saa.5.1.23
- Bayartogtokh B., Ermilov S.G. 2013. Ontogenetic stages of Gymnodamaeus irregularis, with remarks on morphology of the juveniles of Gymnodamaeidae (Acari: Oribatida: Plateremaeoidea). Int. J. Acarol., 39: 7-25. https://doi.org/10.1080/01647954.2012.739640
- Bayartogtokh B., Ermilov S.G. 2014. Rediscovery of Arthrodamaeus rossicus (Acari: Oribatida: Gymnodamaeidae) with remarks on its generic status and ontogeny of the genus. Int. J. Acarol., 40: 556-569. https://doi.org/10.1080/01647954.2014.962085
- Beck L., Woas S. 1991. Die Oribatiden-Arten (Acari) eines südwestdeutschen Buchenwaldes I. Carolinea, Karlsruhe, 49: 37-82.
- Behan-Pelletier V.M. 1997. The semiaquatic genus Tegeocranellus (Acari: Oribatida: Ameronothroidea) of North and Central America. Can. Ent., 129: 537-577. https://doi.org/10.4039/Ent129537-3
- Behan-Pelletier V.M., Lindo Z. 2019. Checklist of oribatid mites (Acari: Oribatida) of Canada and Alaska. Zootaxa, 4666: 1-180. https://doi.org/10.11646/zootaxa.4666.1.1
- Berlese A. 1905. Acari nuovi. Manipulus III. Redia, 2: 10-32.Berlese A. 1910. Acari nuovi. Manipulus V-VI. Redia, 6: 199-234.
- Berlese A. 1913. Acarotheca Italica. Fasciculus Ius et IIus. Firenze. pp. 221.
- Berlese A. 1916a. Centuria prima di Acari nuovi. Redia, 12: 19-67.
- Berlese A. 1916b. Centuria seconda di Acari nuovi. Redia, 12: 126-177.
- Bernini F., Castagnoli M., Nannelli R. 1995. Arachnida, Acari. In: Minelli A., Rufo S., La Posta S. (Eds). Checklist delle specie della fauna italiana, 24. Bologna: Calderini. pp. 131.
- Bischoff de Alzuet A. 1971. Estudio comparativo de la region genito-anal en los distintos estados de Torpacarus omittens Grandjean (Acari: Lohmanniidae). Rev. Soc. Ent. Argentina, 33: 101-104.
- Bulanova-Zachvatkina E.M. 1960. New representatives of primitive oribatid mites from USSR. Perlohmannoidea Grandj. 1958. (Acariformes, Oribatei). Zool. Zh., 39: 1835-1848.
- Bulanova-Zachvatkina E.M. 1967. Armored mites - Oribatids. Moscow: Higher School. pp. 254.
- Bulanova-Zachvatkina E.M. 1975. Superfamily Eulohmannoidea. In: Ghilarov M.S., Krivolutsky D.A. (Eds). A Key to the Soil-Inhabiting Mites. Sarcoptiformes. Moscow: Nauka. p. 68.
- Castagnoli M., Pegazzano F. 1985. Catalogue of the Berlese Acaroteca. Firenze: Istituto Sperimentale per la Zoologia Agraria. pp. 490.
- Chen J., Liu D., Wang H.-F. 2010. Oribatid mites of China: a review of progress, with a checklist. Zoosymposia, 4: 186-224. https://doi.org/10.11646/zoosymposia.4.1.14
- Cianciolo J.M., Norton R.A. 2006. The ecological distribution of reproductive mode in oribatid mites, as related to biological complexity. Exp. Appl. Acarol., 40: 1-25. https://doi.org/10.1007/s10493-006-9016-3
- Dabert M., Witalinski W., Kazmierski A., Olszanowski Z., Dabert J. 2010. Molecular phylogeny of acariform mites (Acari, Arachnida): Strong conflict between phylogenetic signal and long-branch attraction artifacts. Mol. Phyl. Evol., 56: 222-241 https://doi.org/10.1016/j.ympev.2009.12.020
- Domes K., Norton R.A., Maraun M., Scheu S. 2007. Reevolution of sexuality breaks Dollo's law. Proc. Nat. Acad. Sci. USA, 104: 7139-7144. https://doi.org/10.1073/pnas.0700034104
- Engelmann H.D. 1972. Die Oribatidenfauna des Neißetales bei Ostritz (Oberlausitz). Abh. Berich. Naturk. Görlitz, 47: 1-44.
- Ermilov S.G. 2011. Morphology of ovipositors in oribatid mites of the superfamily Crotonioidea (Acari, Oribatida). Zool. Zh., 90: 1168-1174. https://doi.org/10.1134/S0013873811080112
- Ermilov S.G. 2017. A new species of Lohmannia (Lohmannia) from Vietnam, with supplementary description of L. (Lohmannia) turcmenica (Bulanova-Zachvatkina, 1960). Syst. Appl. Acarol., 22: 193-207. https://doi.org/10.11158/saa.22.2.4
- Ermilov S.G., Bayartogtokh B. 2022. Ontogenetic instars of Elliptochthonius profundus Norton, 1975 (Acari, Oribatida, Elliptochthoniidae), with remarks on juveniles of the superfamily Parhypochthonioidea. Zootaxa, 5187: 53-68. https://doi.org/10.11646/zootaxa.5187.1.6
- Ermilov S.G., Hugo-Coetzee E.A., Khaustov A.A. 2011. Morphology of juvenile instars of Neoliodes terrestris (Wallwork, 1963) and N. ionicus Sellnick, 1931 (Acari: Oribatida: Neoliodidae). Ann. Zool., 61: 817-830. https://doi.org/10.3161/000345411X622633
- Fernandez N.A. 1984. Contribution a la connaissance de la faune Oribatologique d'Argentine. VI. Eohypochthonius (Neoatrichosus) travei nov. sub-gen., nov. sp. Acarologia, 25: 95-106.
- Fuangarworn M., Norton R.A. 2013. Psammochthoniidae n. fam., a paedomorphic family of oribatid mites (Oribatida: Enarthronota) from sandy soil in Thailand, Brazil and the USA. Zootaxa, 3691: 473-499. https://doi.org/10.11646/zootaxa.3691.4.7
- Fujikawa T. 1991. List of oribatid families and genera of the world. Edaphologia, 46: 1-132.
- Fujikawa T. 2014. The second representative of the family Eulohmanniidae Grandjean, 1931 (Acari, Oribatida) from Japan. Edaphologia, 93: 1-10.
- Fujikawa T., Fujita M., Aoki J. 1993. Checklist of oribatid mites of Japan (Acari: Oribatida). J. Acarol. Soc. Japan, 2, Suppl. 1. pp. 121. https://doi.org/10.2300/acari.2.Supplement_1
- Gong X., Chen T.W., Zieger S.L., Bluhm C., Heidemann K., Schaefer I., Maraun M., Liu M., Scheu S. 2018. Phylogenetic and trophic determinants of gut microbiota in soil oribatid mites. Soil Biol. Biochem., 123: 155-164. https://doi.org/10.1016/j.soilbio.2018.05.011
- Ghilarov M.S., Krivolutsky D.A. 1975. A Key to the Soil-Inhabiting Mites. Sarcoptiformes. Moscow, Nauka. pp. 491.
- Grandjean F. 1931. Observations sur les Oribates (1re série). Bull. Mus. nat. Hist. natur. (2), 3: 131-144.
- Grandjean F. 1933. Etude sur le développement des Oribates. Bull. Soc. zool. France, 58: 30-61.
- Grandjean F. 1934a. Phthiracarus anonymum n. sp. Rev. franc. Ent., 1: 51-58.
- Grandjean F. 1934b. Observations sur les Oribates (7e série). Bull. Mus. nat. Hist. natur. (2), 6: 423-431.
- Grandjean F. 1934c. Les poils des épimères chez les Oribates (Acariens). Bull. Mus. nat. Hist. natur. (2), 6: 504-512.
- Grandjean F. 1939a. Quelques genres d'acariens appartenant au groupe des Endeostigmata. Ann. Sci. Nat. Zool., 2: 1-122.
- Grandjean F. 1939b. Observations sur les Oribates (11e série). Bull. Mus. nat. Hist. natur. (2), 11: 110-117.
- Grandjean F. 1939c. Observations sur les Oribates (12e série). Bull. Mus. nat. Hist. natur. (2), 11: 300-307.
- Grandjean F. 1939d. Les segments post-larvaires de l′hysterosoma chez les Oribates (Acariens). Bull. Soc. zool. France, 64: 273-284.
- Grandjean F. 1940. Observations sur les Oribates (13e série). Bull. Mus. nat. Hist. natur. (2), 12: 62-69.
- Grandjean F. 1941a. Statistique sexuelle et parthénogenèse chez les Oribates (Acariens). C.R. Séanc. Ac. Sci., 212: 463-467.
- Grandjean F. 1941b. La chaetotaxie comparée des pattes chez les Oribates (1re série). Bull. Soc. zool. France, 66: 33-50.
- Grandjean F. 1942a. La chaetotaxie comparée des pattes chez les Oribates (2e série). Bull. Soc. zool. France, 67: 40-53.
- Grandjean, F. 1942b. Quelques genres d'Acariens appartenant au groupe des Endeostigmata (2e série). Ann. Sci. nat., Zool., Série 11e (Zoologie), 4: 85-135.
- Grandjean F. 1943. L'ambulacre des Acariens (2e série). Bull. Mus. nat. Hist. natur. (2), 15: 303-310.
- Grandjean F. 1946a. Au sujet de l′organe de Claparède, des eupathidies multiples et des taenidies mandibulaires chez les Acariens actinochitineux. Arch. Sci. phys. natur. (5), Genève, 28: 63-87.
- Grandjean F. 1946b. La signification evolutive de quelques caractères des Acariens (1re série). Bul. Biol. France Belg., 79: 297-325.
- Grandjean F. 1947. L'origine de la pince mandibulaire chez les Acariens actinochitineux. Arch. Sci. phys. Nat., Genève, 29: 305-355.
- Grandjean F. 1949a. Observation et conservation des très petits arthropods. Bul. Mus. nat. Hist. natur., 2e série, 21: 364-369.
- Grandjean F. 1949b. Formules anales, gastronotiques, génitales et aggénitales du développement numérique des poils chez les Oribates. Bul. Soc. zool. France, 74: 201-225.
- Grandjean F. 1950a. Observations sur les Oribates (20e série). Bull. Mus. nat. Hist. natur. (2), 22: 73-80.
- Grandjean F. 1950b. Étude sur les Lohmanniidae (Oribates, Acariens). Arch. Zool. exp. gen., 87: 95-162.
- Grandjean F. 1952a. Observations sur les Palaeacaroides (Acariens) (2e série). Bull. Mus. nat. Hist. natur. (2), 24: 460-467.
- Grandjean F. 1952b. Au sujet de l′ectosquelette du podosoma chez les Oribates superieurs et de sa terminologie. Bull. Soc. zool. France, 77: 13-36.
- Grandjean F. 1954a. Essai de classification des Oribates (Acariens). Bull. Soc. zool. France, 78 (1953): 421-446.
- Grandjean F. 1954b. Posthermannia nematophora n.g., n.sp. (Acarien, Oribate). Rev. franc. Ent., 21: 298-311.
- Grandjean F. 1954c. Etude sur les Palaeacaroides (Acariens, Oribates). Mém. Mus. nat. Hist. natur., 7: 179-274.
- Grandjean F. 1955. L'organe de Claparède et son ecaille chez Damaeus onustus Koch. Bull. Mus. nat. Hist. natur. (2), 27: 285-292.
- Grandjean F. 1956a. Caracteres chitineux de l′ovipositor, en structure normale, chez les Oribates (Acariens). Arch Zool. exp. gen., Notes et Revue, 93: 96-106.
- Grandjean F. 1956b. Observations sur les Oribates (35e série). Bull. Mus. nat. Hist. natur. (2), 28: 282-289.
- Grandjean F. 1957. L'infracapitulum et la manducation chez les Oribates et d'autres Acariens. Ann. Sci. Nat. Série 2. Zool., 19: 234-281.
- Grandjean F. 1958a. Perlohmannia dissimilis (Hewitt) (Acarien, Oribate). Mém. Mus. nat. Hist. natur., 16: 57-119.
- Grandjean F. 1958b. Sur le comportement et la notation des poils accessoires posterieurs aux tarses des Nothroides et d'autres Acariens. Arch. zool. exp. gén., 96: 277-308.
- Grandjean F. 1959. Observations sur les Oribates (40e série). Bul. Mus. nat. Hist. natur., 31: 359-366.
- Grandjean F. 1960. Damaeus arvernensis n. sp. (Oribate). Acarologia, 2: 250-277.
- Grandjean F. 1961. Considérations numériques sur les poils génitaux des Oribates. Acarologia, 3: 620-636.
- Grandjean F. 1962. Nouvelles observations sur les Oribates (2e série). Acarologia, 4: 396-422.
- Grandjean F. 1964a. Nouvelles observations sur les Oribates (3e série). Acarologia, 6: 170-198.
- Grandjean F. 1964b. Pheroliodes wehnkei (Willmann) (Oribate). Acarologia, 6: 353-386.
- Grandjean F. 1964c. La solénidiotaxie des Oribates. Acarologia, 6: 529-556.
- Grandjean F. 1966. Collohmannia gigantea Selln. (Oribate). Première partie. Acarologia, 8: 328-357.
- Grandjean F. 1968. Nouvelles observations sur les Oribates (6e série). Acarologia, 10: 357-391.
- Grandjean F. 1969. Considérations sur le classement des Oribates. Leur division en 6 groupes majeurs. Acarologia, 11: 127-153.
- Grandjean F. 1971. Nouvelles observations sur les Oribates (8e série). Acarologia, 12: 849-876.
- Hammen L. van der 1959. Berlese's primitive oribatid mites. Zool. Verh., Leiden, 40: 1-93.
- Hammen L. van der 1963. The oribatid family Phthiracaridae I. Introduction and redescription of Hoplophthiracarus pavidus (Berlese). Acarologia, 5: 306-317.
- Hammen L. van der 1968. The gnathosoma of Hermannia convexa (C.L. Koch) (Acarida: Oribatina) and comparative remarks on its morphology in other mites. Zool. Verh., Leiden, 94: 1-45.
- Hammen L. van der 1980. Glossary of Acarological Terminology. Volume I. General Terminology. The Hague: Dr. W. Junk Publ. pp. 244.
- Hammen L. van der 1982. Comparative studies in Chelicerata II. Epimerata (Palpigradi and Actinotrichida). Zool. Verh., 196: 1-70.
- Hammer M. 1952. Investigations on the microfauna of Northern Canada, Part I. Oribatidae. Acta Arct., 4: 1-108.
- Hammer M., Wallwork J.A. 1979. A review of the world distribution of oribatid mites (Acari: Cryptostigmata) in relation to continental drift. Biol. Skr. Kong. Dansk. Vidensk. Selsk., 22: 1-31.
- Haumann G. 1991. Zur Phylogenie primitiver Oribatiden, Acari: Oribatida. Graz: Verl. Tech. Univ. Graz. pp. 237.
- Heethoff M., Domes K., Laumann M., Maraun M., Norton R.A., Scheu S. 2007. High genetic divergences indicate ancient separation of parthenogenetic lineages of the oribatid mite Platynothrus peltifer (Acari, Oribatida). Europ. Soc. Evol. Biol., 20: 392-402. https://doi.org/10.1111/j.1420-9101.2006.01183.x
- Hewitt C.G. 1908. On a new phytophagous mite, Lohmannia insignis Berl. var. dissimilis n. var., with notes on other species of economic importance. Mem. Proc. Manch. Lit. Phil. Soc. (Manchester Mem.), Manchester, 52: 1-10.
- Ito M. 1986. An ecological survey on arboreal oribatid mites (Acari: Oribatida) in a subalpine coniferous forest of Shiga-kogen, Central Japan. Edaphologia, 35: 19-26.
- Kaneko N. 1988. Feeding habits and cheliceral size of oribatid mites in cool temperate forest soils in Japan. Rev. Écol. Biol. Sol, 25: 353-363.
- Kethley J. 1990. Acarina: Prostigmata (Actinedida). In: Dindal D.L. (Ed.). Soil Biology Guide. New York: Wiley and Sons. pp. 667-750.
- Kethley J. 1991. A procedure for extraction of microarthropods from bulk soil samples with emphasis on inactive stages. Agriculture, Ecosystems and Environment, 34: 193-200. https://doi.org/10.1016/0167-8809(91)90105-7
- Klimov P.A., OConnor B.M., Chetverikov P.E., Boltond S.J., Pepato A.R., Mortazavi A.L., Tolstikov A.V., Bauchan G.R., Ochoa R. 2018. Comprehensive phylogeny of acariform mites (Acariformes) provides insights on the origin of the four-legged mites (Eriophyoidea), a long branch. Mol. Phyl. Evol., 119: 105-117. https://doi.org/10.1016/j.ympev.2017.10.017
- Krantz G.W., Walter D.E. 2009. A Manual of Acarology. Texas Tech University Press. Lubbock, Texas. pp. 807.
- Kunst M. 1971. Key to the Fauna of Czechoslovakia - Supercohort Oribatei. In: Daniel M., Cerny V. (Eds). Praha: Klic zvireny CSSR IV. Academia, Praha. pp. 531-580.
- Lebrun P. 1971. Ecologie et biocenotique de quelques peuplements d'arthropodes edaphiques. Mem. Inst. Roy. Sci. Nat. Belg., 165: 1-203.
- Lebrun P., Wauthy G. 1981. Quelques observations et réflexions sur les peuplements d'Oribates hypogés (Acariens). Ann. Soc. Roy. Zool. Malacol. Belg., 111: 131-142.
- Lee D.C. 1984. A modified classification for oribate mites (Acari: Cryptostigmata). In: Griffiths D.A., Bowman C.E. (Eds). Acarology VI. Horwood, Chichester. Vol. 1. pp. 241-248.
- Lee D.C. 1985. Sarcoptiformes (Acari) of South Australian soils. 4. Primitive oribate mites (Cryptostigmata) with an extensive, unfissured hysteronotal shield and aptychoid. Rec. South Australian Mus., 19: 39-67.
- Li W.N., Xue X.F. 2019. Mitochondrial genome reorganization provides insights into the relationship between oribatid mites and astigmatid mites (Acari: Sarcoptiformes: Oribatida). Zool. J. Lin. Soc., 187: 585-598. https://doi.org/10.1093/zoolinnean/zlz044
- Lienhard A., Krisper G. 2021. Hidden biodiversity in microarthropods (Acari, Oribatida, Eremaeoidea, Caleremaeus). Nature Portfolio, Sci. Rep., 11: 23123, online, pp. 14. https://doi.org/10.1038/s41598-021-02602-7
- Lions J.C. 1964. La variation du nombre des ongles des pattes de Rhysotritia ardua (C.L. Koch 1836) (Acariens, Oribatei). Rev. Écol. Biol. Sol, 1: 41-65.
- Lions J.C. 1966. Contribution a l′étude du genre Rhysotritia (Acariens, Oribate): Rhysotritia clavata sextiana n. subsp. Vie et Milieu Ser. C, 17: 779-794.
- Lions J.C. 1978. Éléments sur la distribution verticale des Oribates (Acariens) dans les biotopes édaphiques d'un écosystème forestier. Rev. Écol. Biol. Sol, 15: 345-362.
- Lions J.C., Norton R.A. 1998. North American Synichotritiidae (Acari: Oribatida) 2. Synichotritia spinulosa and S. caroli. Acarologia, 39: 265-284.
- Lombardini G. 1962. Acari nuovi del Comelico (Alpi orientali). Ann. Centr. Econ. Montana delle Venezie, Padova, 2 (1960-61): 189-216.
- Lozano-Fernandez J., Tanner A.R., Giacomelli M., Carton R., Vinther J., Edgecombe G.D., Pisani D. 2019. Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida. Nat. Com., 10: 2295, online, pp. 8. https://doi.org/10.1038/s41467-019-10244-7
- Luxton M. 1972. Studies on the oribatid mites of a Danish beech wood soil. I. Nutritional biology. Pedobiologia, 12: 434-463.
- Luxton M. 1981. Studies on the oribatid mites of a Danish beech wood soil. IV. Developmental biology. Pedobiologia, 21: 312-340.
- MacKay J.R. 1974. The Mackenzie Delta area, N.W.T. Geological Survey of Canada, Miscellaneous Report 23. Ottawa: Geol. Surv. Canada. pp. 222.
- Mahunka S. 1987. New and interesting mites from the Geneva Museum. LX. Oribatids from Sabah (East Malaysia). II. Acari Oribatida. Rev. suisse Zool., 94: 765-817. https://doi.org/10.5962/bhl.part.79551
- Mahunka S., Mahunka-Papp L. 1995. The oribatid species described by Berlese (Acari). Budapest: Hung. Nat. Hist. Mus. pp. 325.
- Maraun M., Heethoff M., Schneider K., Scheu S., Weigmann G., Cianciolo J., Thomas R.H., Norton R.A. 2004. Molecular phylogeny of oribatid mites (Oribatida, Acari): evidence for multiple radiations of parthenogenetic lineages. Exp. Appl. Acarol., 33: 183-201. https://doi.org/10.1023/B:APPA.0000032956.60108.6d
- Märkel K. 1964. Die Euphthiracaridae Jacot 1930 und ihre Gattungen (Acari, Oribatei). Zool. Verh., Leiden, 67, 3-78.
- Marshall V.G., Reeves R.M., Norton R.A. 1987. Catalogue of the Oribatida (Acari) of continental United States and Canada. Mem. Ent. Soc. Canada, 139. pp. 418. https://doi.org/10.4039/entm119139fv
- Martinez P.A., Bernava Laborde V. 2000. Gehypochthonius marianoi n. sp. (Acari: Oribatida), from sand dunes in coastal Argentina. Acarologia, 41: 381-389.
- Matthews J.V. Jr. 1974. Quaternary environments at Cape Deceit (Seward Peninsula, Alaska): evolution of a tundra ecosystem. Geol. Soc. Amer. Bull. 85: 1353-1384. https://doi.org/10.1130/0016-7606(1974)85<1353:QEACDS>2.0.CO;2
- Norton R.A. 1975. Elliptochthoniidae, a new mite family (Acarina: Oribatei) from mineral soil in California. J. NY Ent. Soc., 83: 209-216.
- Norton R.A. 1977. A review of F. Grandjean's system of leg-chaetotaxy in the Oribatei and its application to the Damaeidae. In: Dindal D.L. (Ed.). Biology of Oribatid Mites. Syracuse: State Univ. NY, Col. Env. Sci. Forestry. p. 33-62.
- Norton R.A. 1982. Arborichthonius n. gen., an unusual enarthronote soil mite (Acarina: Oribatei) from Ontario. Proc. Entomol. Soc., Washington, 84: 85-96.
- Norton R.A. 1984. Monophyletic groups in the Enarthronota (Sarcoptiformes). In: Griffiths D.A., Bowman C.E. (Eds). Acarology VI, vol. I. Chichester: Horwood. p. 233-240.
- Norton R.A. 1994. Evolutionary aspects of oribatid mite life histories and consequences for the origin of the Astigmata. In: Houck M. (Ed.). Mites. Ecological and evolutionary analyses of life-history patterns. New York: Chapman and Hall. p. 99-135. https://doi.org/10.1007/978-1-4615-2389-5_5
- Norton R.A. 1998. Morphological evidence for the evolutionary origin of Astigmata (Acari: Acariformes). Exp. Appl. Acarol., 22: 559-594.
- Norton R.A. 2001. Systematic relationships of Nothrolohmanniidae, and the evolutionary plasticity of body form in Enarthronota (Acari: Oribatida). In: Halliday R.B., Walter D.E., Proctor H.C., Norton R.A., Colloff M.J. (Eds). Acarology: Proceedings of the 10th International Congress. Melbourne: CSIRO Publishing. p. 58-75.
- Norton R.A. 2007. Holistic acarology and ultimate causes: examples from the oribatid mites. In: Morales-Malacara J.B., Behan-Pelletier V., Ueckermann E., Pérez T.M., Estrada-Venegas E.G., Badil M. (Eds). Acarology XI: Proceedings of the International Congress. México: Inst. Biol. Facult. Ciencias, Univ. Nac. Autón. México; Soc. Latinoamer. Acarología. p. 3-20.
- Norton R.A. 2010. Systematic relationships of Lohmanniidae (Acari: Oribatida). In: Sabelis M.W., Bruin J. (Eds). Trends in Acarology. Proceedings of the 12th International Congress. Dordrecht: Springer. p. 9-16. https://doi.org/10.1007/978-90-481-9837-5_2
- Norton R.A., Alberti G. 1997. Porose integumental organs of oribatid mites (Acari, Oribatida). 3. Evolutionary and ecological aspects. Zoologica, Stuttgart, 146: 115-143.
- Norton R.A., Behan-Pelletier V.M. 2007. Eniochthonius mahunkai sp. n. (Acari: Oribatida: Eniochthoniidae), from North American peatlands, with a redescription of Eniochthonius and a key to North American species. Acta Zool. Acad. Sci. Hung., 53: 295-333.
- Norton R.A., Behan-Pelletier V.M. 2009. Chapter 15, Oribatida. In: Krantz G.W., Walter D.E. (Eds). A Manual of Acarology. Lubbock, Texas: Texas Tech University Press. p. 430-564.
- Norton R.A., Ermilov S.G. 2014. Catalogue and historical overview of juvenile instars of oribatid mites (Acari: Oribatida). Zootaxa, 3833: 1-132. https://doi.org/10.11646/zootaxa.3833.1.1
- Norton R.A., Franklin E. 2018. Paraquanothrus n. gen. from freshwater rock pools in the USA, with new diagnoses of Aquanothrus, Aquanothrinae, and Ameronothridae (Acari, Oribatida). Acarologia, 58: 557-627. https://doi.org/10.24349/acarologia/20184258
- Norton R.A., Fuangarworn M. 2015. Nanohystricidae n. fam., an unusual, plesiomorphic enarthronote mite family endemic to New Zealand (Acari, Oribatida). Zootaxa, 4027: 151-204. https://doi.org/10.11646/zootaxa.4027.2.1
- Norton R.A., Kethley J.B. 1989. Berlese's North American oribatid mites: historical notes, recombinations, synonymies and type designations. Redia, 62: 421-499.
- Norton R.A., Metz L.J. 1980. Nehypochthoniidae (Acari: Oribatei), a new mite family from the Southeastern United States. Ann. Ent. Soc. America, 73: 54-62. https://doi.org/10.1093/aesa/73.1.54
- Norton R.A., Palmer S.C. 1991. The distribution, mechanisms and evolutionary significance of parthenogenesis in oribatid mites. In: Schuster R., Murphy P.W. (Eds). The Acari - Reproduction, Development and Life-history Strategies. London, New York: Chapman and Hall. p. 107-136. https://doi.org/10.1007/978-94-011-3102-5_7
- Norton R.A., Sidorchuk E.A. 2014. Collohmannia johnstoni n. sp. (Acari, Oribatida) from West Virginia (U.S.A.), including description of ontogeny, setal variation, notes on biology and systematics of Collohmanniidae. Acarologia, 54: 271-334. https://doi.org/10.1051/acarologia/20142134
- Norton R.A., OConnor, B.M., Johnston D.E. 1983. Systematic relationships of the Pediculochelidae (Acari: Acariformes). Proc. ent. soc. Washington, 85: 493-512.
- Norton R.A., Bonamo P.M., Grierson J.D., Shear W.A. 1988. Oribatid mite fossils from a terrestrial Devonian deposit near Gilboa, New York. J. Paleont., 62: 259-269. https://doi.org/10.1017/S0022336000029905
- Norton R.A., Kethley J., Johnston D.E., OConnor B.M. 1993. Phylogenetic perspectives on genetic systems and reproductive modes of mites. In: Wrensch D.L., Ebbert M.A. (Eds). Evolution and Diversity of Sex Ratio in Insects and Mites. New York, London: Chapman and Hall. p. 8-99. https://doi.org/10.1007/978-1-4684-1402-8_2
- Norton R.A., Behan-Pelletier V.M., Wang H.F. 1996. The aquatic oribatid mite genus Mucronothrus in Canada and the Western USA (Acari, Trhypochthoniidae). Can. J. Zool., 74: 926-949. https://doi.org/10.1139/z96-106
- OConnor B.M. 1984. Phylogenetic relationships among higher taxa in the Acariformes, with particular reference to the Astigmata. In: Griffiths D.A., Bowman C.E. (Eds). Acarology VI. Horwood, Chichester. Vol. 1. pp. 19-62.
- OConnor, B.M. 2009. Chapter 16, Cohort Astigmatina. In: Krantz G.W., Walter D.E. (Eds). A Manual of Acarology. Lubbock, Texas: Texas Tech University Press. p. 565-657.
- Ohkubo N. 2002. Two species of Epilohmannoides (Acari: Oribatida) of Japan. Edaphologia, 70: 1-5.
- Pachl P., Domes K., Schulz G., Norton R.A., Scheu S., Schaefer I., Maraun M. 2012. Convergent evolution of defense mechanisms in oribatid mites (Acari, Oribatida) shows no ′ghosts of predation past′. Mol. Phyl. Evol., 65: 412-420. https://doi.org/10.1016/j.ympev.2012.06.030
- Pachl P., Lindl A.C., Krause A., Scheu S., Schaefer I., Maraun M. 2017. The tropics as an ancient cradle of oribatid mite diversity. Acarologia, 57: 309-322. https://doi.org/10.1051/acarologia/20164148
- Pachl P., Uusitalo M., Scheu S., Schaefer I., Maraun M. 2021. Repeated convergent evolution of parthenogenesis in Acariformes (Acari). Ecology and Evolution, 11: 321-337. https://doi.org/10.1002/ece3.7047
- Palmer S.C., Norton R.A. 1992. Genetic diversity in thelytokous oribatid mites (Acari, Acariformes, Desmonomata). Bioch. Syst. Ecol., 20: 219-231. https://doi.org/10.1016/0305-1978(92)90056-J
- Pan′kov A.N., Ryabinin N.A., Golosova L.D. 1997. Catalogue of oribatid mites of the Far East of Russia. Part I. Catalogue of oribatid mites of Kamchatka, Sakhalin and the Kuril Islands. Vladivostok, Khabarovsk: Dalnauka Press. pp. 87.
- Pfingstl T., Lienhard A., Baumann J., Koblmüller S. 2021. A taxonomist's nightmare - Cryptic diversity in Caribbean intertidal arthropods (Arachnida, Acari, Oribatida). Mol. Phyl. Evol., 163: 107240, online. pp. 19. https://doi.org/10.1016/j.ympev.2021.107240
- Pollierer M.M., Scheu S. 2021. Stable isotopes of amino acids indicate that soil decomposer microarthropods predominantly feed on saprotrophic fungi. Ecosphere, 12: e03425, online. pp. 15. https://doi.org/10.1002/ecs2.3425
- Purrini K., Bukva V. 1984. Pathenogenetic agents of oribatid mites (Oribatei, Acarina): a gap in the research on population dynamics. In: Griffiths D.A., Bowman C.E. (Eds.). Acarology VI. Horwood, Chichester. Vol. 2. pp. 826-837.
- Purrini K., Weiser J. 1981. Eight new microsporidian parasites of moss mites (Oribatei, Acarina) in forest soils. Z. angew. Ent., 91: 217-224. https://doi.org/10.1111/j.1439-0418.1981.tb04471.x
- Radford C.D. 1950. Systematic check list of mite genera and type species. Int. Union Biol. Sci., 1: 1-232.
- Rafalski J. 1966. Contributions to the knowledge of the Oribatei (Acari) of Poland. I. Fragm. Faun., 12: 347-372. https://doi.org/10.3161/00159301FF1966.12.21.347
- Rajski A. 1961. Ecological-faunistic studies on oribatid mites in several vegetative associations I. Ecology. Pozn. Towarz. Przyj. Nauk, 25: 123-287.
- Rajski A. 1967. Autecological-zoogeographical analysis of moss mites (Acari, Oribatei) on the basis of fauna in the Poznan Environs. Part I. Bull. Entomol. Pol., 37: 69-166. https://doi.org/10.3161/00159301FF1968.14.12.277
- Raspotnig G. 2006. Characterisation of monophyletic oribatid groups by oil gland chemistry - a novel systematic approach in Oribatida (Acari). Abh. Berich. Natur. Görlitz, 78: 31-46.
- Raspotnig G. 2010. Oil gland secretions in Oribatida (Acari). In: Sabelis M.W., Bruin J. (Eds). Trends in Acarology. Proceedings of the 12th International Congress. Dordrecht: Springer. p. 235-239. https://doi.org/10.1007/978-90-481-9837-5_38
- Ren G., Yang M., Liang W., Xie L. 2017. A new species of Meristacarus from China, with supplementary description of Mixacarus (Phyllolohmannia) foliifer (Golosova, 1984) (Acari, Oribatida, Lohmanniidae). Syst. Appl. Acarol., 22: 1989-2007. https://doi.org/10.11158/saa.22.11.15
- Riha G. 1951. Zur Ökologie der Oribatiden in Kalksteinböden. Zool. Jahrb. Syst., 80: 408-450.
- Ryabinin N.A., Pan′kov A.N. 2002. Catalogue of oribatid mites of the Far East of Russia. Part II. Continental part of the Far East. Vladivostok, Khabarovsk, Russia: Dalnauka Press. pp. 92.
- Schäffer S., Kerschbaumer M., Koblmüller S. 2019. Multiple new species: Cryptic diversity in the widespread mite species Cymbaeremaeus cymba (Oribatida, Cymbaeremaeidae). Mol. Phyl. Evol., 135: 185-192. https://doi.org/10.1016/j.ympev.2019.03.008
- Schaefer I., Norton R.A., Scheu S., Maraun M. 2010. Arthropod colonization of land - Linking molecules and fossils in oribatid mites (Acari, Oribatida). Mol. Phyl. Evol., 57: 113-121. https://doi.org/10.1016/j.ympev.2010.04.015
- Schatz H., Behan-Pelletier V.M., OConnor B.M., Norton R.A. 2011. Suborder Oribatida van der Hammen, 1968. In: Zhang Z.-Q. (Ed.). Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148: 141-148. https://doi.org/10.11646/zootaxa.3148.1.26
- Schuster R. 1956. Der Anteil der Oribatiden an den Zersetzungsvorgängen im Boden. Z. Morph. Ökol. Tiere, 45: 1-33. https://doi.org/10.1007/BF00699814
- Schuster R. 1960. Über die Morphologie und Artengliederung der Gattung Epilohmannia Berlese 1917 (Acari, Oribatei). Zool. Anz., 165: 197-213.
- Schweizer J. 1956. Die Landmilben des Schweizerischen Nationalparkes. 3. Teil, Sarcoptiformes Reuter 1909. Ergebnisse der wissenschaftlichen Untersuchungen des schweizerischen Nationalparks, Neue Folge, Liestal, 5: 213-377.
- Schweizer J. 1957. Die Landmilben des Schweizerischen Nationalparkes. 4. Teil: Ihr Lebensraum, ihre Vergesellschaftung unter sich und ihre Lebensweise. Ergebnisse der wissenschaftlichen Untersuchungen des schweizerischen Nationalparks, Neue Folge, Liestal, 6: 11-107.
- Sellnick M. 1928. Formenkreis: Hornmilben, Oribatei. In: Brohmer P., Ehrmann P., Ulmer G. (Eds.). Die Tierwelt Mitteleuropas. Leipzig: Quelle und Meyer, (3) 9. p. 1-42.
- Seniczak S. 1991. The morphology of juvenile stages of moss mites of the family Nanhermanniidae (Acari, Oribatida) 1. Zool. Anz., 227: 319-330.
- Seniczak S., Seniczak A., Kaczmarek S. 2018. Morphological ontogeny of Platyliodes scaliger (Acari, Oribatida, Neoliodidae), with comments on Platyliodes Berlese. Syst. Appl. Acarol., 23: 25-41. https://doi.org/10.11158/saa.23.1.3
- Seniczak S., Ivan O., Kaczmarek S., Seniczak A. 2020. Morphological ontogeny of Lopheremaeus mirabilis (Acari: Oribatida: Plateremaeidae), and comments on Lopheremaeus Paschoal. Syst. Appl. Acarol., 25: 2147-2164. https://doi.org/10.11158/saa.25.12.1
- Sergienko G.D. 1994. Archoribatida - Lower oribatids. In: Topachevsky (Ed.). Fauna of the Ukraine 25. Kiev: Naukova Dumka. p. 5-204.
- Shafer A.B.A., Cullingham C.I., Côté S.D., Coltman D.W. 2010. Of glaciers and refugia: a decade of study sheds new light on the phylogeography of northwestern North America. Mol. Ecol. 19: 4589-4621. https://doi.org/10.1111/j.1365-294X.2010.04828.x
- Siepel H., de Ruiter-Dijkman E.M. 1993. Feeding guilds of oribatid mites based on their carbohydrase activities. Soil Biol. Biochem., 25: 1491-1497. https://doi.org/10.1016/0038-0717(93)90004-U
- Strenzke K. 1963. Entwicklung und Verwandtschaftsbeziehungen der Oribatidengattung Gehypochthonius (Arach., Acari). Senck. Biol., 44: 231-255.
- Subías L.S. 2004. Listado sistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes, Oribatida) del mundo (1758-2002). Graellsia, 60: 3-305.
- Subías L.S. 2022. Listado sistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes, Oribatida) del mundo (1758-2002). Monografías electrónicas S.E.A. (Sociedad Entomológica Aragonesa), n° 12. pp. 538. Available from: http://www.sea-entomologia.org
 https://doi.org/10.3989/graellsia.2004.v60.iExtra.218
https://doi.org/10.3989/graellsia.2004.v60.iExtra.218 - Suomalainen E. 1950. Parthenogenesis in animals. Advances in Genetics, 3: 193-253. https://doi.org/10.1016/S0065-2660(08)60086-3
- Suzuki K. 1977. Ontogenetic development of Perlohmannia gigantea (Aoki, 1960). I. Morphology of larval stase (Acari: Cryptostigmata). Acta Arach., 27: 95-107. https://doi.org/10.2476/asjaa.27.Specialnumber_95
- Suzuki K. 1979. Oribatid fauna of Hachioji City (2). Mem. Educ. Inst. Private Schools in Japan, 64: 117-123.
- Taberly G. 1988. Recherches sur la parthénogenèse thélytoque de deux espèces d'Acariens Oribates: Trhypochthonius tectorum (Berlese) et Platynothrus peltifer (Koch). IV. Observations sur les mâles ataviques. Acarologia, 29: 95-107.
- Tadros M.S. 1975. The correlation between occurrence of both soil mites (Oribatei) and soil fungi in Qualubia Governorate. Vest. Ceskoslovenske Spol. Zool., 29: 294-296.
- Trägårdh I. 1910. Acariden aus dem Sarekgebirge. Naturw. Unters. Sarek., Zool., 4: 375-586.
- Travé J. 1967. Phyllochthonius aoutii nov. gen., nov. spec., un Enarthronota (Acarien, Oribate) nouveau de Cote d'Ivoire, avec la creation d'une superfamilie nouvelle, Phyllochthonoidea. Zool. Med., 42: 83-105.
- Travé J. 1975. Sur les stases immatures de deux genres d'Oribotritiidae (Oribates). Acarologia, 17: 536-545.
- Travé J. 1977. La néotrichie epimerique d'Hermannia jesti sp. n. (Oribate). Acarologia, 19: 123-131.
- Travé J. 1978. La néotrichie chez les Oribates (Acariens). Acarologia, 20: 590-602.
- Travé J. Vachon M. 1975. François Grandjean 1882-1975 (Notice biographique et bibliographique). Acarologia, 17: 1-19.
- Travé J., André H.M., Taberly G., Bernini F. 1996. Les Acariens Oribates. Wavre, Belgium: AGAR and Société internationale des Acarologues de Langue française. pp. 110.
- Vitzthum H. 1943. Acarina. In: Bronn H.G. (Ed.). Klassen und Ordnungen des Tierreiches, vol. 5: Arthropoda, part IV: Arachnoidea, Book 5: Acarina. Leipzig: Becker and Erler Kom.-Ges. pp. 1011.
- Wafa A.K., El-Kifl A.H., Tadros M.S. 1964. Ecological studies on oribatids in Giza Region (Egypt). Proc. 1st Int. Congr. Acarol. 1963, Acarologia, fasc. hors. sér., 6: 183-186.
- Wauthy G., Fain A. 1991. Observations on the legs of Trimalaconothrus maniculatus Fain & Lambrechts, 1987 (Acari, Oribatida). Part 1. Larva, leg IV of nymphs and fundamental phanerotaxy. Acarologia, 32: 415-434.
- Weigmann G. 2006. Hornmilben (Oribatida). Die Tierwelt Deutschlands. Teil 76. Keltern: Goecke and Evers. pp. 520.
- Willmann C. 1931. Moosmilben oder Oribatiden (Cryptostigmata). In: Dahl F. (Ed.): Die Tierwelt Deutschlands, Bd. 22, vol. 5. Jena: Gustav Fischer. pp. 79-200.
- Woodring J.P., Cook E.F. 1962. The internal anatomy, reproductive physiology, and molting process of Ceratozetes cisalpinus (Acarina, Oribatei). Ann. Ent. Soc. America, 55: 164-181. https://doi.org/10.1093/aesa/55.2.164
- Woas S. 2002. Acari: Oribatida. In: Adis J. (Ed.). Amazonian Arachnida and Myriapoda. Sofia: Pensoft Publishers. p. 21-291.
- Woolley T.A., Baker E.W. 1958. A key to the superfamilies and principal families of the Oribatei (Sarcoptiformes: Acarina). Ent. News, 69: 85-92.
- Xue X.F., Dong Y., Deng W., Hong X.Y., Shao R. 2017. The phylogenetic position of eriophyoid mites (superfamily Eriophyoidea) in Acariformes inferred from the sequences of mitochondrial genomes and nuclear small subunit (18S) rRNA gene. Mol. Phyl. Evol., 109: 271-282. https://doi.org/10.1016/j.ympev.2017.01.009



2022-05-04
Date accepted:
2022-09-22
Date published:
2022-09-28
Edited by:
Baumann, Julia

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Norton, Roy A. and Ermilov, Sergey G.
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)