



The oribatid mite (Acari: Oribatida) community of a high-Andean cushion peatland in southern Peru
Hense, Jonathan  1
; Schittek, Karsten
2
; Forbriger, Markus3
; Mächtle, Bertil
4
; Schäbitz, Frank
5
and Bonkowski, Michael
6
1
; Schittek, Karsten
2
; Forbriger, Markus3
; Mächtle, Bertil
4
; Schäbitz, Frank
5
and Bonkowski, Michael
6
1✉ Biology Education, Nees-Institute, University Bonn, Meckenheimer Allee 170, 53315 Bonn, Germany.
2Institute of Geography Education, University of Cologne, Gronewaldstr. 2, 50931 Cologne, Germany.
3Institute of Geography, Heidelberg University, Im Neuenheimer Feld 348, 69120 Heidelberg, Germany.
4Institute of Geography, Heidelberg University, Im Neuenheimer Feld 348, 69120 Heidelberg, Germany.
5Institute of Geography Education, University of Cologne, Gronewaldstr. 2, 50931 Cologne, Germany.
6Terrestrial Ecology, Institute of Zoology, University of Cologne, Zülpicher Str. 47b, 50674 Cologne, Germany.
2022 - Volume: 62 Issue: 4 pages: 879-891
https://doi.org/10.24349/idxe-mfsxOriginal research
Keywords
Abstract
Introduction
The oribatid mite's species of high-Andean cushion peatlands were initially explored by the groundbreaking work of Hammer (1958, 1961), but recent studies are only available for cushion peatlands of northern and central Chile (Covarrubias and Mellado 2003; Covarrubias 2004a, b, 2009; Figueroa and Covarrubias 2005). In contrast, the oribatid fauna of the Peruvian peatlands is largely unknown.
The cushion peatlands of the high Andes above ~3000 m a.s.l., locally called bofedales, are a unique ecosystem, characterised by extreme environmental conditions with seasonal drought, diurnal frost-thaw cycles, hypoxia, strong winds and intense solar radiation (Ruthsatz 1993; Squeo et al. 2006; Schittek et al. 2012). As a direct consequence of these harsh environmental conditions, parts of the cushion peatland regularly die off, or are infrequently covered by debris from erosion and landslides. This results in a dynamic mosaic of ecotopes. More recently, anthropogenic impacts, especially land-use as pasture dramatically increased the disturbance intensity of cushion peatlands (Ruthsatz 1995; Ramsar-Convention 2005, Poblete 2007), leading to an extremely heterogenous and dynamic mosaic of different successional stages (Schittek et al. 2012, 2015). In general, habitat heterogeneity is a major driving force of species richness (Adams 2009). This connection is well studied in above-ground settings (Mittelbach et al. 2001), but it has been only rarely addressed for below-ground communities (Nielsen et al. 2010). Moreover, habitat heterogeneity is changing in time and its implications, amplitude, and temporal characteristics are the main research focus of the paleoenvironmental discipline. In order to advance paleoenvironmental analyses, it is claimed to intensify multiproxy approaches where different indicators of habitat change are combined (Birks and Birks 2006). In the last two decades, Oribatid mites gained more interest as paleoenvironmental indicators (Erickson and Platt 2013) but were used only once in the Neotropics (Chepstow-Lusty et al. 2007). High-Andean cushion peatlands showed high potential as archives for paleoenvironmental analyses and subfossil oribatid mites are deposited and well preserved over time (Schittek et al. 2015).
Considering the lack of knowledge on oribatid mites of Peruvian peatlands and a need of new paleoenvironmental indicators, the aim of this study was: (i) to characterise the taxonomic richness of oribatid mites in a cushion peatland; (ii) to describe changes in community composition within the soil heterogeneity of the peatland; and (iii) to test if identification to species level can be attained using deposited and damaged dead specimens.
Material and Methods
Study site
The Cerro Llamoca peatland (referred later as CLP) is situated in the western Cordillera of the Andes of southern Peru (14°10′S, 74°44′W) at an altitude between 4150 and 4250 m a.s.l., southward of the name giving mountain peak'Cerro Llamoca', and covers an area of 11.6 hectares (Figure 1). It comprises a part of the unique cushion peatland (''bofedal'') ecosystems situated on slopes, in wet brooks or valley bottoms at 4000-4900 m a.s.l. in the Andes. The cushion peatlands are of significant importance for the local human population: they represent pastureland for wild and domestic animals and are important water reservoirs during drought periods (Ruthsatz 1995).
The study area is characterised by a tropical climate (Lauer 1993), in the sense where extreme diurnal temperature fluctuations, due to the high altitude, are overlaid by distinct annual fluctuations of temperature and precipitation. The ERA-Interim model, renders a mean annual precipitation of 349 mm for the CLP with a marked seasonal pattern (Dee at al. 2011), of which about 90% falls during the austral summer (December to March). Within this wet season, rainy days are distinctly clustered with short periods of thunderstorms and heavy rain, followed by longer dry periods (Garreaud 2000). This precipitation pattern could be termed as ''monsoon-like'' (Zhou and Lau 1998). The peatlands act as an important buffer by absorbing all precipitation and releasing it throughout the year (Verzijl and Quispe 2013). The main cushion-forming plants of the CLP are the Juncaceae Distichia muscoides Nees & Meyen 1834 and Oxychloe andina Phil. 1860 (Schittek et al. 2012, 2015). Overall, the uppermost part of the CLP shows clearly visible transitions between the following ecotopes: (i) Springs in the upper area are bordered by large Distichia muscoides cushions, although most of them are degraded by trampling of hoofed herbivores, such as cattle. Trampling destroys the vital uppermost plants and hence exposes the underlying peat, which fosters stronger evaporation or direct drainage of stored water from the peat cushions and facilitates further vegetation damage and vulnerability to erosion (Schittek et al. 2012): (ii) Due to erosion on the surrounding slopes, transportation of eroded sediments and further accumulation processes in the valley bottoms, the middle section is characterised by a large debris fan covering the peatland. Solely Oxychloe andina inhabits some of the wet debris layers of this area. (iii) The lowest section is less affected by degradation and shows a typical mosaic-like pattern of well-established large Distichia muscoides cushions, interspersed by several accompanying species e.g. Zameioscirpus muticus, Plantago tubulosa, Gentiana prostrata and Werneria pygmaea. The cushions are often encircled by a frame of Deyeuxia chrysantha reeds and are usually separated by small and shallow pools (Schittek et al. 2012).
Sampling strategy and preparation
The sampling aimed to get a full overview of the oribatid mite community of the Cerro Llamoca peatland. Accordingly, five representative ecotopes characterizing distinct microhabitats within the peatland were sampled (Figure 1d). From each ecotope, one core was taken on 28/10/2011 with a soil-corer (5cm diameter x 15cm length). Depending on the depth of the bedrock and other soil characteristics, varying core length from 8 - 24 cm could be retrieved. The cores were immediately divided into two equal halves of the length and stored in half-shell plastic tubes. One half was injected immediately with ca. 0.5 ml ethanol (96%) cm-1 to preserve and fix the soil fauna and to avoid spatial displacements of the invertebrates. The other half was used for the characterization of the soil substrates and categorized as: (i) dense Distichia muscoides-cushion (Figure 2a); (ii) mineral debris (Figure 2b); (iii) silty peat of Oxychloe andina plant material (Figure 2c); (iv) humous silty mud (Figure 2d); and (v) dry peat of dead (Distichia muscoides) material (Figure 2e).

In the laboratory, each half soil core was subsampled into layers of 1 cm thickness. From all five subsampled cores, the samples of each second centimetre (n=41) were chosen for the examination of the mesofauna (see Figure 2). For the analysis, the volume of every sample was quantified by placing the soil in a graduated measuring cylinder and filling it up with distilled H2O to a fixed volume of 10 ml with accuracy 0.1 ml. The samples were then macerated in 10% KOH for at least four hours at 4 °C before being separated by wet sieving into two fractions of \textless112-250µm and \textgreater250µm, respectively. Subsequently, the individual fractions were diluted in water, spread out on a white picking tray (10 x 6cm) and completely scanned in multiple steps under a dissecting microscope at 10-12x magnification. All mesofaunistic individuals or fragments of these were picked out and stored in ethanol (96%) until mounting. In this study, only the data on oribatid mites is presented. Juvenile oribatid mites were not included in the data.

Mounting and identification of oribatid mites
The oribatid mites were cleared in gently heated lactic acid (96%), following the procedure of Weigmann (2006) and mounted in Hoyer's medium (Baker and Wharton 1952), which was stained with potassium iodide and iodine to increase the contrast (Faraji and Bakker 2008). For identification, different light microscopes with magnifications from 50-400x, and literature by Hammer (1958, 1961, 1962a, b), Balogh and Balogh (1988, 1990), Balogh (1972), Norton and Behan-Pelletier (2009) and an interactive glossary by Hunt et al. (1998) were used. The nomenclature is based on Subías (2004, update 2021).
Photos were taken with a Canon EOS 600D mounted on a Zeiss Axio Scope A1 light microscope with differential interference contrast (DIC) illumination, using the remote-control software SmartShooter v.1.1.14. Multi-layer stacking with 10-30 pictures per specimen was performed by the Zerene Stacker v.1.04 software with image processing and measuring width and length by Adobe Photoshop CS4.
According to their species rank-abundance distribution, the species were subdivided according to the abundance classification criteria of Engelmann (1978) into dominant (\textgreater10%), subdominant (\textless10%), recendent (< 2%), subrecendent (\textless1%) and sporadic (\textless0.3%) taxa. These ratios refer to all individuals of a single oribatid mite species in relation to all the total abundance.
Statistical Analyses
The species richness was calculated by rarefaction analysis using the software EstimateS ver. 9.10 (Colwell 2013) and BioDiversity Pro (McAleece et al. 1997), following the procedures by Gotelli and Colwell (2011) and Colwell et al. (2012). For all other statistical analyses, the software R (R Core Team 2018) was used.
Comparisons of multiple, independent and non-parametric datasets were achieved using the Kruskal-Wallis-test with a subsequent post-hoc procedure corrected by the number of tests (Giraudoux 2015). The influences of different parameters on the oribatid mite community were tested by multiple regressions.
To characterise the ranked abundance distribution, different models were fitted to the curve using maximum likelihood estimation with the vegan package (Oksanen et al. 2019). In this package, five models (brokenstick, pre-emption, log-normal, Zipf and Zipf-Mandelbrot) are implemented. The best model fit was determined by the lowest Akaike information criterion (AIC) value.
Results
In total, 17 oribatid mite species were identified, by investigating of 689 adult specimens or remains of these from all 41 samples. The community comprised eight families, of which the most diverse families were the Ceratozetidae with six taxa and the Scheloribatidae with three taxa. Only 85 (12.4%) of the 689 oribatid mite specimens could not be identified because of damage to taxonomically important structures. A mean richness of 2.5 species per sample (SD= 2.7) was discovered, ranging from only one species (Zetomimus furcatus) in the Distichia muscoides cushion substrate to up to 11 species in the dry peat substrate (see Figure 4b). A full overview of the identified oribatid mite community is given in Table 1.
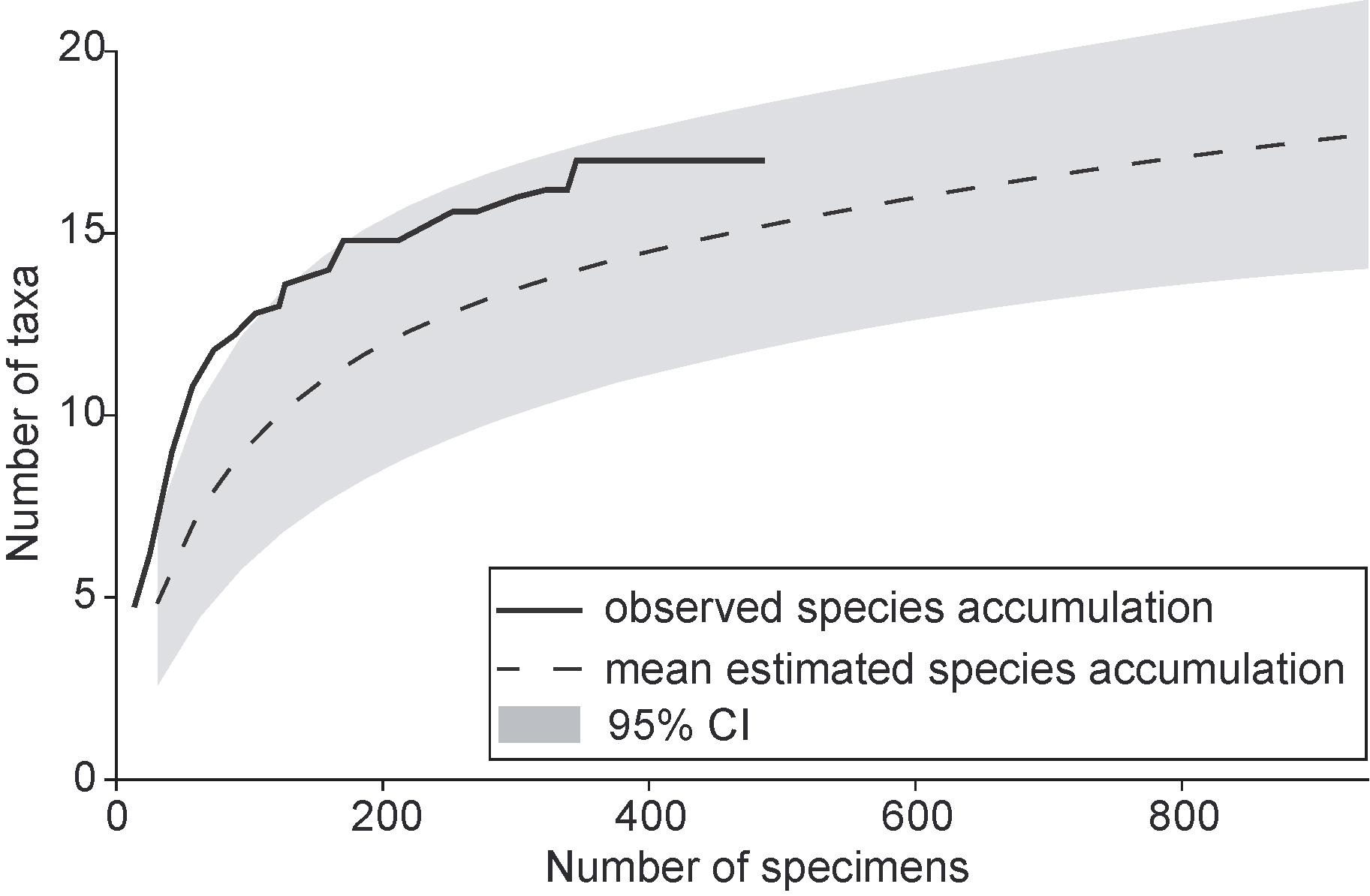


For all samples, the rarefaction analysis shows a non-saturating estimation curve reaching 18 taxa at 1000 counted specimens (Figure 3). The real species accumulation runs in accordance with the upper limit of the 95% confidence interval (CI), which ranges from 13 to 21 species at 1000 counted specimens. The community structure of the peatland inhabiting oribatid mites showed a best fit to a niche preemption model with an AIC support of 59.75 (Figure 5). For the other fitted models, higher AICs were estimated (Zipf-Mandelbrot: 62.3; Lognormal: 63.6; Brokenstick: 64.3; Zipf: 69.4). Tectocepheus sp. (23.8%), Edwardzetes armatus (17.0%) and Trichoribates hammerae (13.7%) were the dominant species of the oribatid mite community. These most abundant species were accompanied by a group of subdominant species: Neoamerioppia notata (9.5%), Malaconothrus monodactylus (5.6%), Nanhermannia elegantissima (5.3%), Scheloribates confundatus (3.5%) and S. pallidulus (3.5%). In addition, there are some recedent species like Zetomimus furcatus (1.4%), Tyrphonothrus maior (1.1%) and Scheloribates elegantulus (1.0%). Two subrecendet species Sphaerozetes chavinensis and Oxyoppia suramericana with \textless1% abundance and four sporadically occurring taxa (Zetomimus nigrisetosus, Aquanothrus sp., Ceratozetes (Magellozetes) sp. and Oripoda sp.) with \textless0.32% complete the species inventory.
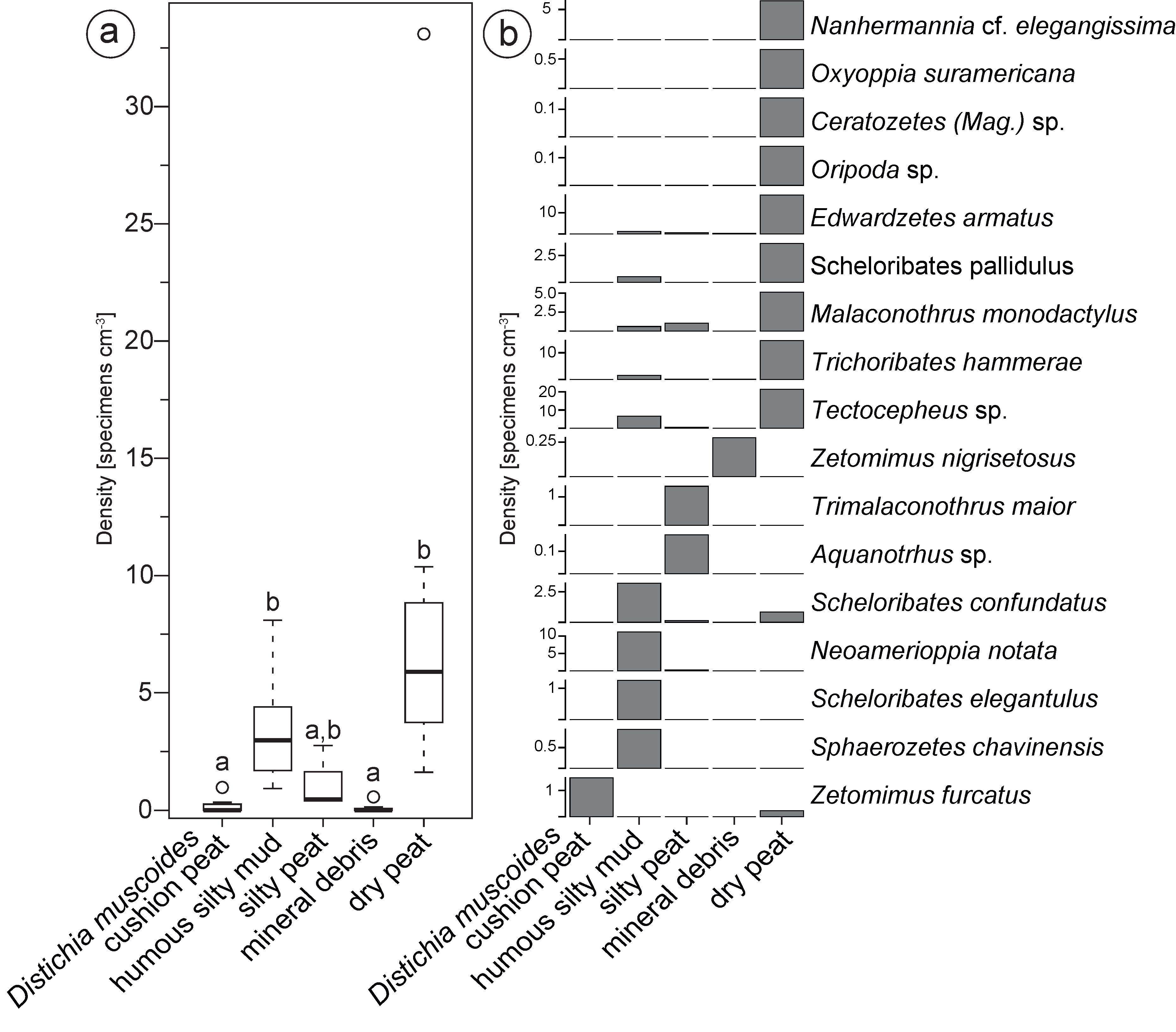
Distribution within the ecotopes
In mean, the oribatid mite community showed an abundance of 3.0 specimens per cm3 (SD= 5.9) (Figure 4a). However, the abundance of oribatid mites differed significantly within different microhabitats of the peatland (H(4)= 27.3, p\textless0.001). Post hoc comparisons showed higher densities specimens cm-3 in the degraded dry Distichia muscoides peat substrate (M= 8.96, SD=10.11) and in the humous silty mud (M= 3.40, SD= 2.30) (see Figure 4a). The different microhabitats had a significant influence (β=0.62, p\textless0.001) on the density of oribatids (F(2,32)= 7.4, p\textless0.001, R2=.27) and explained 27.33% of the overall variation. In the investigated soil cores, the abundance of oribatid mites varied with depth (Figure 2), however, the sample depth had no significant influence on the inhabiting oribatid mite density (β=0.18, p=0.27).

Discussion
The rarefaction analysis of the oribatid mite fauna indicates sufficient sampling density in the current study. The confirmed (17 species) and by rarefaction estimated species richness (18 species) lies within the range of 18 and 13 species per 1000 specimens reported in Chilean cushion peatlands (Covarrubias 2004a; Figueroa and Covarrubias 2005). However, the oribatid mite communities of other cushion peatlands, such as a Patosia clandestina-dominated type in northern Chile, with only seven species are distinctly less diverse (Covarrubias 2009). Nevertheless, cushion peatlands are species rich in comparison to other ecosystems of the high Andes (forest, cactus zone, dwarf shrub heath, tussock grass steppe, and cushion plant zone), which achieve only a mean of 5.4 oribatid species (Covarrubias 2004a).
According to the classification of Engelmann (1978), Tectocepheus sp., Edwardzetes armatus and Trichoribates hammerae were the dominant species of the oribatid mite community. Species of the genus Tectocepheus are extremely ubiquitous, favored by parthenogenetic reproduction and belong to the most common oribatid mites throughout the world (Nübel-Reidelbach 1994; Hajmová and Smrž 2001; Subías 2004, update 2021; Weigmann 2002, 2006). Tectocepheus sp. is reported from cushion peatlands in Chile (Covarrubias 2004a; Figueroa and Covarrubias 2005). In contrast, Edwardzetes armatus and Trichoribates hammerae were exclusively found at altitudes \textgreater4200 m a.s.l. inhabiting wet habitats (Hammer 1958, 1961; Balogh and Balogh 1990). Trichoribates hammerae is endemic to Peru, but Edwardzetes armatus is occurring thoughout the Neotropical region (Subías 2004, update 2021).
The dominant species were accompanied by a group of subdominant species: Neoamerioppia notata, Malaconothrus monodactylus, Nanhermannia elegantissima, Scheloribates confundatus and S. pallidulus. Among these, Neoamerioppia notata is known from moist to wet habitats in the high Andes of Bolivia (Hammer 1958). Malaconothrus monodactylus was reported at high densities from a cushion peatland in Chile (Covarrubias 2004a, 2009). However, it is frequently found in bogs and peatlands throughout the Holarctic and Neotropic ecozones (Subías 2004, update 2021; Weigmann 2006). Nanhermannia elegantissima is a well-known species from the entire Neotropic and other regions (Subías 2004, update 2021) and also from high-Andean cushion peatlands (Hammer 1958; Covarrubias 2004a, b; Figueroa and Covarrubias 2005). The genus Nanhermannia is composed of secondary decomposers feeding mainly on fungi (Schneider et al. 2004; Ermilov and Łochyńska 2008) and show a slow development despite being parthenogenetic. Scheloribates confundatus was found in very wet habitats in Peru and Argentina (Hammer 1961), whereas S. pallidulus is a cosmopolitan species and typical for peat bogs and forest soils in Europe, Eurasia and North America (Ivan et al. 1997; Subías 2004, update 2021; Weigmann 2006; Murvanidze and Kvavadze 2010; Barreto and Lindo 2018; Behan-Pelletier and Lindo 2019).
In addition, some recedent species like Zetomimus furcatus, Tyrphonothrus maior and Scheloribates elegantulus could be determined.
Zetomimus furcatus is one of the few fully aquatic oribatids (Fischer and Schatz 2010), characterised as a specialist of oligotrophic Holarctic peat bogs and other swampy habitats (Schelvis and van Geel 1989; Murvanidze and Kvavadze 2010; Weigmann 2006). Similarly, Tyrphonothrus maior is a semi-aquatic, semi-cosmopolitan and parthenogenetically reproducing species, occurring in very wet habitats (Balogh and Balogh 1988; Palmer and Norton 1991; Subías 2004, update 2021; Schatz and Behan-Pelletier 2008; Lehmitz and Maraun 2016; Minor et al. 2019; Barreto and Lindo 2021). In contrast, Scheloribates elegantulus was found in moss between heathery shrubs (Hammer 1961) and in semi-deciduous forest fragments surrounded by savanna (Ferreira et al. 2012). This species shows a Neotropical distribution (Subías 2004, update 2021).
The community structure showed a best fit to a niche preemption model (Wilson 1991). This fit characterises a community in which each species arrives randomly and preempts a constant fraction of the remaining resources. This is typical for small, stressed or pioneer communities (May 1975). Together with the frequent occurrence of parthenogenetic species, the composition and structure of the oribatid community likely reflects the selective forces acting in the extremely harsh environmental conditions of the high Andean cushion peatlands such as extreme diurnal temperature fluctuations, intense solar radiation and seasonal droughts (Ruthsatz 1993; Squeo et al. 2006; Schittek et al. 2012).
The oribatid density in our study ranged from 0.08 specimens cm-3 in the debris ecotope to 8.96 specimens cm-3 in the degraded ecotope, and it was much higher than reported in the soils of other cushion peatlands with densities ranging between 0.2-1.4 specimens cm-3 (Figueroa and Covarrubias 2005; Covarrubias 2009). These latter values, however, were obtained by Berlese-Tullgren funnel extraction of the living oribatid mites, while we handpicked all dead and damaged specimens from the sample. In particular the relatively high oribatid densities in dry peat likely indicate higher availability of food resources (e.g. fungi) due to higher decomposition rates in the degraded parts of the peatlands. Unpublished data on high fungi spore concentrations in the dry peat samples supports this hypothesis (Dzakovic, unpublished data). If this connection can be confirmed in future studies, the density of oribatid mites may thus be used as an indicator of peatland degradation.
In view of the extremely low abundances of mites in cushion peatland soils we consider the hand-picking procedure described in this study as superior. In addition, by including the remains of dead specimens, we could obtain a much more complete species inventory of the oribatid mite community. Most importantly, we were able to identify even heavily damaged oribatid mites with the existing literature. This gives us a good precondition to use dead and preserved specimens from older soil deposits as indicators of the past changes in ecotone composition in future studies.
Conclusion
This investigation represents the first comprehensive study of the oribatid mite fauna of a high-Andean cushion peatland from Peru. The oribatid mites inhabit only distinct substrates within the full range of the peatland's heterogeneous ecotopes. In the dense Distichia muscoides cushions, almost no oribatids could be observed, but relatively high densities in dry peatland parts may indicate increased degradation of the peatlands due to increased decomposer activity caused by trampling of cows.
It is demonstrated that the identification of oribatid mites is also possible using in situ dead specimens obtained from the soil samples by hand-picking. Identification of dead species was particularly important to complement the species inventory by rare taxa. For taxonomic determination, the absence of the legs, which is often the case in dead oribatid mite specimens, is only a problem for specific taxa. However, the absence of legs requires an adapted use of the available identification keys for South America. The comments on morphological features, given in this study, provide a means to ease the use of fossil remains as indicators for environmental change in these habitats. With the synthesis of the taxonomic, distributional and autecological data, a useful tool for (paleo-)environmental research could be developed. More research on oribatid mites of the high Andes and especially for Peru is needed to obtain a complete species inventory of this region. On account of the age, and due to the remoteness and isolation of the cushion peatland ecosystems, its oribatid mite fauna may serve as an ideal model system for evolutionary studies.
Acknowledgements
The authors give their sincere thanks to Steffen Woas and Ludwig Beck (both State Museum of Natural History Karlsruhe, Germany), René Covarrubias (Chile), Mark Maraun (University of Göttingen, Germany) and Roy A. Norton (Syracuse, USA) for invaluable taxonomic help. Special thanks go to Alexander-Chepstow-Lusty whose comments significantly improved the manuscript.
acarologia_4548_supplementary.pdf
References
- Adams J. 2009. Species richness: patterns in the diversity of life. Chichester: Springer. 386 pp.
- Baker E., Wharton G. 1952. An introduction to acarology. New York: The Macmillan Company. 482 pp.
- Balogh J. 1972. The oribatid genera of the world. Budapest: Akademiai Kiado. 188 pp.
- Balogh J., Balogh P. 1988. Oribatid mites of the neotropical region I. Amsterdam: Elsevier. 336 pp.
- Balogh J., Balogh P. 1990. Oribatid mites of the neotropical region II. Amsterdam: Elsevier. 334 pp.
- Barreto C., Lindo Z. 2018. Drivers of decomposition and the detrital invertebrate community differ across a hummock-hollow microtopology in boreal peatlands. Ecoscience, 25: 39-48. https://doi.org/10.1080/11956860.2017.1412282
- Barreto C., Lindo Z. 2021. Checklist of oribatid mites (Acari: Oribatida) from two contrasting boreal fens: an update on oribatid mites of Canadian peatlands. Syst. Appl. Acarol., 26: 866-884. https://doi.org/10.11158/saa.26.5.4
- Behan-Pelletier V. M., Lindo. Z. 2019. Checklist of oribatid mites (Acari: Oribatida) of Canada and Alaska. Zootaxa, 4666: 1-180. https://doi.org/10.11646/zootaxa.4666.1.1
- Birks H.H., Birks. H.J. 2006. Multi-proxy studies in palaeolimnology. Veget. Hist. Archaeobot., 15: 235-251. https://doi.org/10.1007/s00334-006-0066-6
- Chepstow-Lusty A., Frogley M., Bauer B., Leng M., Cundy A., Boessenkool K., Gioda K. A. 2007. Evaluating socio-economic change in the Andes using oribatid mite abundances as indicators of domestic animal densities. J. Archaeol. Sci., 34: 1178-1186. https://doi.org/10.1016/j.jas.2006.12.023
- Colwell R. 2013. EstimateS: Statistical estimation of species richness and shared species from samples. Version 9. Available from: http://purl.oclc.org/estimates

- Colwell R., Chao A., Gotelli N., Lin S., Mao C., Chazdon R., Longino J. 2012. Models and estimators linking individual-based and sample based rarefraction, extrapolation and comparison of assemblages. J. Plant Ecol., 5: 3-21. https://doi.org/10.1093/jpe/rtr044
- Covarrubias R. 2004a. Ácaros oribátidos (Acari: Oribatida) de la región Altiplánica de Chile. Acta Ent. Chilena, 28: 33-39.
- Covarrubias R. 2004b. La pequeña fauna de artrópodos que vive dentro del suelo: buscando en el Altiplano. Informes de Investigación, 2: 1-8.
- Covarrubias R. 2009. Microartrópodos de la estepa altoandina altiplánica, con detalle de especies de oribátidos (Oribatida: Acarina). Neotrop. Entomol., 38: 482-490. https://doi.org/10.1590/S1519-566X2009000400007
- Covarrubias R., Mellado I. 2003. Microartrópodos de suelos asociados a vegetación altiplanica. I. parque nacional Volván Isluga, Chile. Acta Ent. Chilena, 27: 25-35.
- Dee D. P., Uppala S. M., Simmons A. J., Berrisford P., Poli P., Kobayashi S., Andrae U., Balmaseda M. A., Balsamo G., Bauer P., Bechtold P., Beljaars A. C. M., van de Berg L., Bidlot J., Bormann N., Delsol C., Dragani R., Fuentes M., Geer A. J., Haimberger L., Healy S. B., Hersbach H., Hólm E. V., Isaksen L., Kållberg P., Köhler M., Matricardi M., McNally A. P., Monge-Sanz B. M., Morcrette J.-J., Park B.-K., Peubey C., de Rosnay P., Tavolato C., Thépaut J.-N.,Vitart F. 2011. The ERA-Interim reanalysis: configuration and performance of the data assimilation system. Q. J. Roy. Meteor. Soc., 137: 553-597. https://doi.org/10.1002/qj.828
- Engelmann H.-D. 1978. Zur Dominanzklassifizierung von Bodenarthropoden. Pedobiologia, 18: 378-380. https://doi.org/10.1016/0011-2275(78)90090-5
- Erickson J., Platt. R. 2013. Oribatid Mites. In: Elias, S. (Ed.). The encyclopedia of quaternary science. Amsterdam: Elsevier. p. 697-698. https://doi.org/10.1016/B978-0-444-53643-3.00276-4
- Ermilov S., Łochyńska M. 2008. The influence of temperature on the development time of three oribatid mite species (Acari, Oribatida). North West J. Zool., 4: 274-281.
- Faraji F., Bakker F. 2008. A modified method for clearing, staining and mounting plant-inhabiting mites. Eur. J. Entomol, 105: 793-795. https://doi.org/10.14411/eje.2008.105
- Ferreira R., Franklin E., Pereira de Souza J., de Moraes J. 2012. Soil oribatid mite (Acari: Oribatida) diversity and composition in semi-deciduous forest fragments in eastern Amazonia and comparison with the surrounding savanna matrix. J. Nat. Hist., 12: 1-14. https://doi.org/10.1080/00222933.2012.707245
- Figueroa M., Covarrubias R. 2005. Oribátidos (Acarina: Oribatida) en vegas de altura de la región Metropolitana, Chile. Acta Ent. Chilena, 29: 37-44.
- Fischer B., Schatz H. 2010. Hornmilbenarten (Acari: Oribatida) in Feuchtgebieten Südtirols (Italien). Gredleriana, 10: 209-226.
- Garreaud R. 2000. Intraseasonal variability of moisture and rainfall over the South American Altiplano. American Meterological Society - Monthly Weather Reviews, 128: 3337-3346. https://doi.org/10.1175/1520-0493(2000)128<3337:IVOMAR>2.0.CO;2
- Giraudoux P. 2015. pgrimess: Data analysis in ecology. R package version 1.6.0. Available from: https://cran.r-project.org/web/packages/pgrimess/index.html
- Gotelli N.,Colwell R. 2011. Estimating species richness. Biological diversity: frontiers in measurement and assessment, 12: 39-54.
- Hajmová M., Smrž J. 2001. Reproductive and nutritional biology of Tectocepheus velatus (Acari: Tectocepheidae) in different biotopes. In: Halliday R., Walter D., Proctor H., Norton R., Colloff M. (Eds). Acarology: Proceedings of the 10th International Congress. Melbourne: Csiro Publishing. pp. 255-261.
- Hammer M. 1958. Investigations on the Oribatid fauna of the Andes Mountains. I. The Argentine and Bolivia. Biol. Skr. Dan. Vid. Selsk., 10(1): 1-129.
- Hammer M. 1961. Investigations on the Oribatid fauna of the Andes Mountains. II. Peru. Biol. Skr. Dan. Vid. Selsk., 13(1): 1-157.
- Hammer M. 1962. Investigations on the Oribatid fauna of the Andes Mountains. III. Chile. Biol. Skr. Dan. Vid. Selsk., 13(2): 1- 96.
- Hammer M. 1962. Investigations on the Oribatid fauna of the Andes Mountains. VI. Patagonia. Biol. Skr. Dan. Vid. Selsk., 13(3): 1-37.
- Hunt G., Norton R. A., Kelly J., Colloff, M., Lindsay, S. 1998. Oribatid mites - an interactive glossary.
- Ivan Q., Vasiliu N., Vasiliu M. 1997. The fauna and structure of the Oribatid mites communities from height peat bogs in the Eastern Carpathians (Romania). The Poiana Stampei and Luciana nature reserves. Anurual Muzeului Natinal al Bucovinci, 14: 35-42.
- Lauer W. 1993. Human development and environment in the Andes: a geoecological overview. Mt. Res. Dev., 13: 157-166. https://doi.org/10.2307/3673633
- Lehmitz R., Maraun M. 2016. Small-scale spatial heterogeneity of stable isotopes signatures ($δ ^1^5$N, $δ^1^3$C) in Sphagnum sp. transfers to all trophic levels in oribatid mites. Soil Biol. Biochem., 100: 242-251. https://doi.org/10.1016/j.soilbio.2016.06.005
- May R. M. 1975. Patterns of species abundance and diversity. In: Cody M. L., Diamond J. M. (Eds). Ecology and Evolution of Communities. Cambridge: The Belknap Press of Harvard University Press. pp. 81-120.
- McAleece N., Gage J., Lambshead J., Patterson G. 1997. Biodiversity professional. Version 2.0. Available from: http://www.sams.ac.uk/peter-lamont/biodiversity-pro
- Minor M. A., Ermilov S.G., Philippov D.A. 2019. Hydrology-driven environmental variability determines abiotic characteristics and Oribatida diversity patterns in a Sphagnum peatland system. Exp. Appl. Acarol., 77: 43-58. https://doi.org/10.1007/s10493-018-0332-1
- Mittelbach G., Steiner C. F., Scheiner S. M., Gross K., Reynolds H., Waide R., Willig M., Dodson S. I., Gough L. 2001. What is the observed relationship between species richness and productivity? Ecology, 82: 2381-2396. https://doi.org/10.1890/0012-9658(2001)082[2381:WITORB]2.0.CO;2
- Murvanidze M., Kvavadze E. 2010. An inventory of Oribatid mites, the main decomposers in bogs of Colchic Lowland (Caucasus, Georgia). In: Sabelus M., Bruin J. (Eds). Trends in Acarology. Berlin: Springer. pp. 175-178. https://doi.org/10.1007/978-90-481-9837-5_28
- Nielsen U. N., Osler G. H., Campbell C. D., Neilson R., Burslem D., van der Wal R. 2010. The enigma of soil animal species diversity revisited: the role of small-scale heterogeneity. PloS one, 5: e11567. https://doi.org/10.1371/journal.pone.0011567
- Norton R. A., Behan-Pelletier V. M. 2009. Suborder Oribatida. In: Krantz G., Walter D. (Eds). A manual of acarology. Lubbock: Texas Tech University Press. pp. 430-564.
- Nübel-Reidelbach E. 1994. Taxonomy and systematics of the genus Tectocepheus Berlese, 1895 (Acari, Oribatei). Andrias, 12: 3-94.
- Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., Minchin P. R., O'Hara R. B., Simpson G. L., Solymos P., Stevens M. H. H., Eduard S., Wagner H. 2019. vegan: community ecology package. R package version 2.5-6. Available from http://CRAN.R-project.org/package=vegan
- Palmer S., Norton R. A. 1991. Taxonomic, geographic and seasonal distribution of thelytokous parthenogenesis in the Desmonomata (Acari: Oribatida). Exp. Appl. Acarol., 12: 67-81. https://doi.org/10.1007/BF01204401
- Poblete R. 2007. Uso de vegas y bofedales de la zona cordillerana y precordillerana de la región de Atacama. Chile, Universidad de Chile, Santiago de Chile.
- R Core Team. 2019. R: A language and environment for statistical computing. Available from http://www.R-project.org
- Ramsar-Convention 2005. Regional strategy for the conservation and sustainable use of High Andean wetlands. Proceedings from Wetlands and water: supporting life, sustaining livelihood, Kampala.
- Ruthsatz B. 1993. Flora und ökologische Bedingungen hochandiner Moore Chiles zwischen 18°00′ (Arica) und 40°30′ (Osorno) südl. Breite. Phytocoenologia, 23: 157-199. https://doi.org/10.1127/phyto/23/1993/157
- Ruthsatz B. 1995. Vegetation und Ökologie tropischer Hochgebirgsmoore in den Anden Nord-Chiles. Phytocoenologia, 25: 185-234. https://doi.org/10.1127/phyto/25/1995/185
- Schatz H., Behan-Pelletier V. 2008. Global diversity of oribatids (Oribatida: Acari: Arachnida). Hydrobiologia, 595: 323-328. https://doi.org/10.1007/978-1-4020-8259-7_35
- Schelvis J., van Geel B. 1989. A palaeoecological study of the mites (Acari) from a lateglacial deposit at Usselo (The Netherlands). Boreas, 18: 237-243. https://doi.org/10.1111/j.1502-3885.1989.tb00398.x
- Schittek K., Forbriger M., Mächtle B., Schäbitz F., Wennrich V., Reindel M., Eitel B. 2015. Holocene environmental changes in the highlands of the southern Peruvian Andes (14°S) and their impact on pre-Columbian cultures. Clim. Past, 11: 27-44. https://doi.org/10.5194/cp-11-27-2015
- Schittek K., Forbriger M., Schäbitz F., Eitel, B. 2012. Cushion Peatlands - Fragile water resources in the High Andes of Southern Peru. Landscape and Sustainable Development, 4: 62-85.
- Schneider K., Migge S., Norton R. A., Scheu S., Langel R., Reineking A., Maraun M. 2004. Trophic niche differentiation in soil microarthropods (Oribatida, Acari): evidence from stable isotope ratios (15N/14N). Soil Biol Biochem, 36: 1769-1774. https://doi.org/10.1016/j.soilbio.2004.04.033
- Squeo F. A., Warner B. G., Aravena R., Espinoza D. 2006. Bofedales: high altitude peatlands of the central Andes. Rev. Chil. de Hist. Nat., 79: 245-255. https://doi.org/10.4067/S0716-078X2006000200010
- Subías L.S. 2004, update 2021. Listado sistemático, sinonímico y biogeográfico de los àcaros Oribátidos (Acariformes: Oribatida) del mundo (except fósiles), 16ª actualización. pp. 532. Available from: http://bba.bioucm.es/cont/docs/RO_1.pdf (accessed 07 Jul. 2021).
- Verzijl A., Quispe S.G. 2013. The system nobody sees: irrigated wetland management and Alpaca herding in the Peruvian Andes. Mt. Res. Dev., 33: 280-293 https://doi.org/10.1659/MRD-JOURNAL-D-12-00123.1
- Weigmann G. 2002. Morphological variability between and within populations of Tectocepheus (Acari, Oribatida, Tectocepheidae) from the velatus-complex in Central Europe. In: Bernini, F., Nannelli, R., Nuzzaci, G., de Lillo, E. (Eds). Acarid phylogeny and evolution: adaptation in mites and ticks. Dordrecht: Springer. pp. 141-152. https://doi.org/10.1007/978-94-017-0611-7_15
- Weigmann G. 2006. Hornmilben (Oribatida). In: Dahl F. Die Tierwelt Deutschlands, Teil 76. Keltern: Goecke & Evers. pp. 520.
- Wilson J. B. 1991. Methods for fitting dominance/diversity curves. J. Veg. Sci., 2: 35-46. https://doi.org/10.2307/3235896
- Zhou J.,Lau K.-M. 1998. Does a monsoon climate exist over South America? JCLI, 11: 1020-1040. https://doi.org/10.1175/1520-0442(1998)011<1020:DAMCEO>2.0.CO;2



2022-04-15
Date accepted:
2022-08-16
Date published:
2022-09-06
Edited by:
Pfingstl, Tobias

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Hense, Jonathan; Schittek, Karsten; Forbriger, Markus; Mächtle, Bertil; Schäbitz, Frank and Bonkowski, Michael
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)