



Two new species of Euscheloribates (Acari, Oribatida, Scheloribatidae) from the Philippines
Ermilov, Sergey G.  1
and Corpuz- Raros, Leonila
2
1
and Corpuz- Raros, Leonila
2
1✉ Institute of Environmental and Agricultural Biology (X-BIO), Tyumen State University, Tyumen, Russia.
2Institute of Weed Science, Entomology and Plant Pathology, College of Agriculture and Food Science, and Museum of Natural History, University of the Philippines Los Banos, College, Laguna, Philippines.
2022 - Volume: 62 Issue: 3 pages: 811-820
https://doi.org/10.24349/dipc-7xfqZooBank LSID: 81D97E2F-8889-4AD7-8545-892BBDE17101
Original research
Keywords
Abstract
Introduction
The oribatid mite genus Euscheloribates (Acari, Oribatida, Scheloribatidae) was proposed by Kunst, 1958 with Euscheloribates samsinaki Kunst, 1958 as type species. According to the catalogue of Subías (2004, updated 2022) the genus comprises two subgenera and 14 species, which are distributed in the Palaearctic, Tropical and Subtropical regions. The main generic traits of Euscheloribates are: rostrum rounded, pointed or incised; lamella simple; prodorsal setae comparatively long; bothridial seta with developed head; pteromorph broad, immovable; 10 pairs of short notogastral setae or their alveoli; three pairs of genital, 0–1 pair of aggenital, 1–2 pairs of anal, and 3 pairs of adanal setae; all legs monodactylous (see Kunst 1958; Balogh and Balogh 1992; Subías 2020).
Among the oribatid mite materials collected from the Philippines, we found two new species of Euscheloribates. The main goal of the paper is to describe these two new species.
At present, one species of Euscheloribates, E. payatosensillus (Corpuz-Raros, 1979) was registered in the Philippine fauna (Corpuz-Raros and Ermilov 2009).
Methods
Observation and documentation — Specimens were mounted in lactic acid on temporary cavity slides for measurement and illustration. Body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the notogaster. Notogastral width refers to the maximum width of the notogaster in dorsal view. Lengths of body setae were measured in lateral aspect. All body measurements are presented in micrometers. Formulas for leg setation are given in parentheses according to the sequence trochanter-femur-genu-tibia-tarsus (famulus included). Formulas for leg solenidia are given in square brackets according to the sequence genu-tibia-tarsus. Drawings were made with a camera lucida using a Leica transmission light microscope ''Leica DM 2500''.
Terminology — Morphological terminology used in this paper follows that of Grandjean: see Travé & Vachon (1975) for references; Norton (1977) for leg setal nomenclature; and Norton & Behan-Pelletier (2009) for overview.
Abbreviations — Prodorsum: lam = lamella; plam = prolamella; slam = sublamella; Al = sublamellar porose area; tlam = translamella; tu = tutorium; plc = prodorsal lateral carina; ro, le, in, bs = rostral, lamellar, interlamellar, and bothridial seta, respectively; ex = exobothridial setal alveolus; Ad = dorsosejugal porose area; D = dorsophragma; P = pleurophragma. Notogaster: c, la, lm, lp, h, p = notogastral setae; Sa, S1, S2, S3 = sacculi; ia, im, ip, ih, ips = lyrifissures; gla = opisthonotal gland opening. Gnathosoma: a, m, h = subcapitular setae; or = adoral seta; d, l, cm, acm, ul, su, lt, vt, inf, sup = palp setae; ω = palp solenidion; cha, chb = cheliceral setae; Tg = Trägårdh's organ. Epimeral and lateral podosomal regions: 1a, 1b, 1c, 2a, 3a, 3b, 4a, 4b = epimeral setae; Ah = humeral porose area; PdI, PdII = pedotectum I and II, respectively; dis = discidium; cir = circumpedal carina. Anogenital region: g, ag, an, ad = genital, aggenital, anal, and adanal setae, respectively; iad = adanal lyrifissure; p.o. = preanal organ. Legs: Tr, Fe, Ge, Ti, Ta = trochanter, femur, genu, tibia, and tarsus, respectively; p.a. = porose area; ɛ = famulus; d, l, v, ev, bv, ft, tc, it, p, u, a, s, pv, pl = setae; ω, σ, φ = solenidia.
Taxonomy
Euscheloribates guitingensis n. sp.
ZOOBANK: 6CD7359C-86AF-471D-8E7F-6C920A485421 ![]()
(Figures 1a-c; 3a, c, e-g; 4a-d)


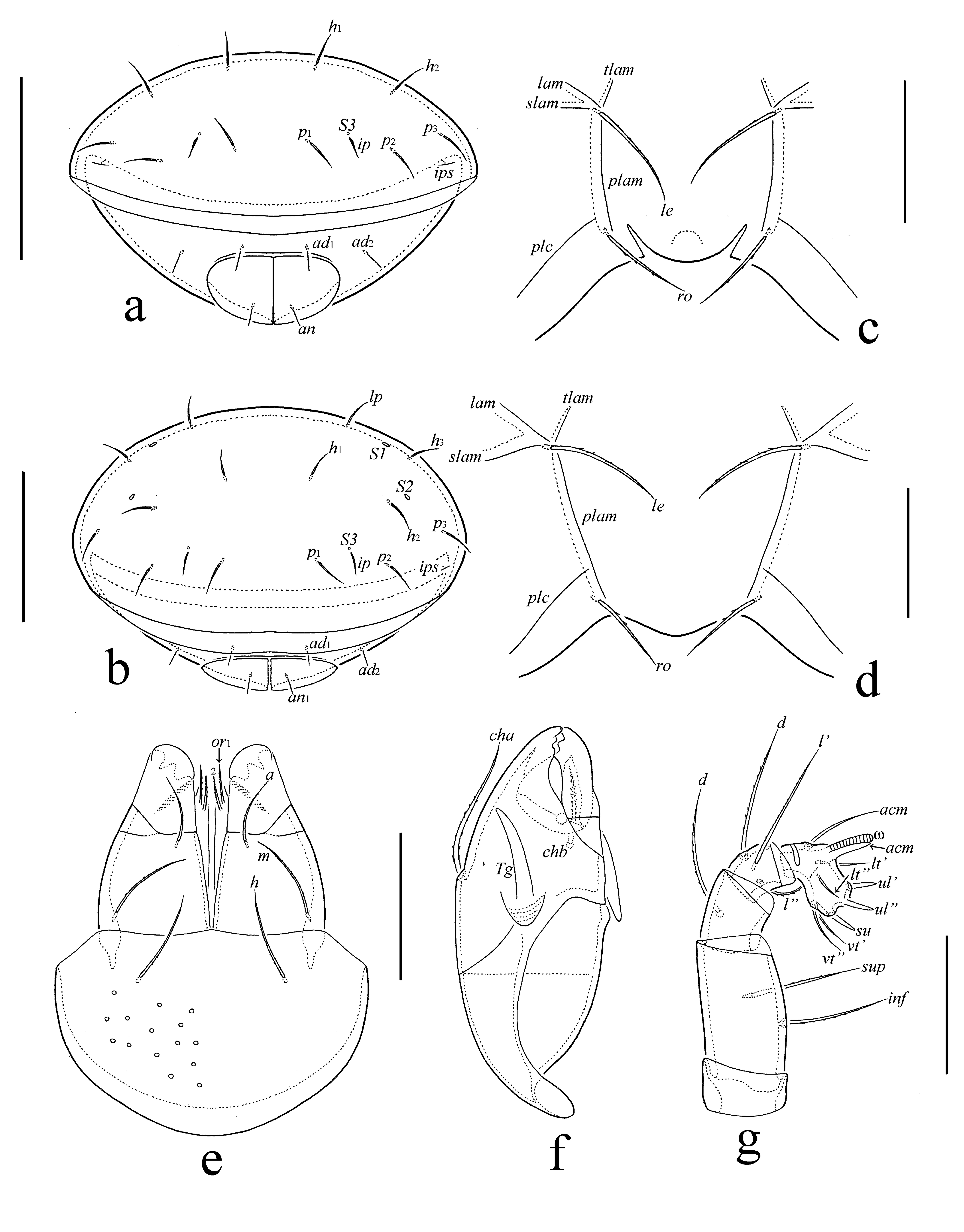
Diagnosis — Body length: 249–282. Notogaster, epimeral and anogenital regions partially with sparse foveolae. Rostrum with two incisions. Translamella represented by two short ridges directed backwards. Tutorium present. Rostral and lamellar setae long, setiform, slightly barbed; interlamellar seta long, setiform, erect, barbed; ro shortest, le longest. Bothridial seta with short stalk and elongate clavate, slightly barbed head. Anterior notogastral margin distinctly convex medially. All notogastral setae short, setiform, slightly flexible, smooth. Epimeral and anogenital setae short, setiform, smooth; both aggenital setae present; each anal plate with one seta. Circumpedal carina long. Leg tarsus I with 16 setae, tarsus III with 13 setae, trochanter III with one seta, genua IV with one seta.
Description of adult — Measurements – Body length: 282 (holotype, female), 265–282 (four paratype females), 249 (three paratype males); body width: 149 (holotype), 140–157 (four paratype females), 123–132 (three paratype males).
Integument – Body color brown. Cuticle microporose (visible in dissected specimens under high magnification, ×1500). Notogaster (except median and lateral part), epimeral region and partially anogenital region sparsely foveolate (diameter of foveola up to 2).
Prodorsum – Rostrum with two incisions. Lamella about 1/2 length of prodorsum. Prolamella lineate. Sublamella about 2/3 length of prodorsum. Sublamellar porose area rounded (4–6) or oval (6–8 × 4). Translamella absent, only two short ridges developed near lamellae, directed backwards. Tutorium medium-sized, simple. Rostral (22–24) and lamellar (34–36) setae setiform, slightly barbed. Interlamellar seta (28–30) setiform, erect, barbed. Exobothridial seta represented by alveolus. Bothridial seta (24–30) with short (4–6), smooth stalk and longer (18–24), elongate clavate, slightly barbed head. Bothridium with lateral scale. Dorsosejugal porose area oval (6 × 2–4). Dorsophragma distinctly elongate.
Notogaster – Anterior notogastral margin distinctly convex medially. Ten pairs of notogastral setae (14–16) setiform, thin, slightly flexible, smooth. Four pairs of sacculi drop-like; S3 smallest, sometimes poorly visible. Opisthonotal gland opening and all lyrifissures distinct.
Gnathosoma – Subcapitulum size: 65–69 × 45–49; subcapitular (a: 12; m, h: 16) and adoral (6) setae setiform, roughened. Palp (length: 41–45) setation: 0–2–1–3–9 (+ω); solenidion swollen distally. Postpalpal seta (4) spiniform, smooth. Chelicera (length: 65–69) with two setiform, barbed setae (cha: 24; chb: 14–16).
Epimeral and lateral podosomal regions – Epimeral formula: 3–1–2–2; setae (c: 4; others: 10–12) setiform, thin, smooth. Humeral porose area Ah distinct; Am not observed. Pedotecta I, II represented by small laminae; pedotectum II rounded distally in ventral aspect. Discidium triangular, rounded distally. Circumpedal carina comparatively long, anteriorly reaching pedotectum II.
Anogenital region – Three pairs of genital (10–12), one pair of aggenital (10–12), one pair of anal (10–12), and three pairs of adanal (12–14) setae setiform, thin, smooth. Adanal lyrifissure distinct. Marginal porose area not observed. Ovipositor elongated (85 × 24), blade (32) shorter than length of distal section (beyond middle fold; 53); each of the three blades with four smooth setae, ψ1 ≈ τ1 (20) setiform, ψ2 ≈ τa ≈ τb ≈ τc (8) slightly thorn-like; six coronal setae (4) spiniform.
Legs – Claw slightly barbed on dorsal side. Porose area on all femora and on trochanters III, IV well visible; porose area on all tarsi and tibiae absent. Formulas of leg setation and solenidia: I (1–5–2–4–16) [1–2–2], II (1–5–2–4–15) [1–1–2], III (1–2–1–3–13) [1–1–0], IV (1–2–1–3–12) [0–1–0]; homology of setae and solenidia indicated in Table 1. Famulus short, slightly swollen distally, inserted posterior to solenidion ω2; seta d on femora III, IV inserted dorsobasally on segments. Solenidion ω1 on tarsus I, ω1 and ω2 on tarsus II and σ on genu III slightly bacilliform; other solenidia setiform.
Material examined — Holotype (female) and seven paratypes (three males and four females): Philippines, Sibuyan Is., Romblon Province, Mt. Magdiwang, Mount Guiting-guiting Natural Park, leaf litter in forest (Museum number: JCBN 17 0330-2), 30.III.2017 (leg. O.L. Eusebio).
Type deposition — The holotype is deposited in the collection of the Senckenberg Museum of Natural History, Görlitz, Germany; seven paratypes are deposited in the collection of the Tyumen State University Museum of Zoology, Tyumen, Russia. All specimens are preserved in 70% solution of ethanol with a drop of glycerol.
Etymology — The species name guitingensis refers ro the place of origin, Mount Guiting-guiting Natural Park.
Remarks — Euscheloribates guitingensis n. sp. is morphologically most similar to Euscheloribates olszanowskii Bayartogtokh & Shimano, 2020 from Palau in having aggenital setae, clavate bothridial seta with short stalk and one pair of anal setae. However, the new species differs from the latter by larger body size (length: 249–282 versus 203–208), in the presence (versus absence) of tutorium, two incisions on the rostrum and partial foveolate notogaster and ventral plate, and by reduced leg chaetome (e.g. tarsus I with 16 setae, tarsus III with 13 setae, trochanter III and genua IV with one seta versus tarsus I with 19 setae, tarsus III with 15 setae, trochanter III and genua IV with two setae).
Euscheloribates magdiwangensis n. sp.
ZOOBANK: EA8D40AF-4219-4543-95AE-99B0A7379957 ![]()
(Figures 2b, d, 4a-c;)

Diagnosis — Body length: 265–298. Rostrum rounded. Translamella represented by two short ridges directed backwards. Tutorium present. Rostral and lamellar setae long, setiform, slightly barbed; interlamellar seta long, setiform, erect, barbed; ro shortest, le longest. Bothridial seta with short stalk and elongate clavate, slightly barbed head. Anterior notogastral margin distinctly convex medially. All notogastral setae short, setiform, slightly flexible, smooth. Epimeral and anogenital setae short, setiform, smooth; both aggenital setae present; each anal plate with two setae. Circumpedal carina long. Leg tarsus I with 16 setae, tarsus III with 13 setae, trochanter III with one seta, genu IV with one seta.
Description of adult — Measurements – Body length: 265 (holotype, male), 298 (two paratype females), 265 (one paratype male); body width: 132 (holotype), 149, 166 (two paratype females), 132 (one paratype male).
Integument – Body color brown. Cuticle microporose (visible in dissected specimens under high magnification, ×1500).
Prodorsum – Rostrum slightly protruding, rounded. Lamella about 1/2 length of prodorsum. Prolamella lineate. Sublamella about 2/3 length of prodorsum. Sublamellar porose area rounded (4–6) or oval (8 × 4). Translamella absent, only two short ridges developed near lamellae, directed backwards. Tutorium medium-sized, simple. Rostral (28–32) and lamellar (49–53) setae setiform, slightly barbed. Interlamellar seta (36–41) setiform, erect, barbed. Exobothridial seta represented by alveolus. Bothridial seta (24–30) with short (4–6), smooth stalk and longer (20–24), elongate clavate, slightly barbed head. Bothridium with lateral scale. Dorsosejugal porose area oval (6 × 2–4). Dorsophragma distinctly elongate.
Notogaster – Anterior notogastral margin distinctly convex medially. Ten pairs of notogastral setae (14–16) setiform, thin, slightly flexible, smooth. Four pairs of sacculi drop-like; S3 smallest, sometimes poorly visible. Opisthonotal gland opening and all lyrifissures distinct.
Gnathosoma – Subcapitulum size: 65–69 × 45–53; subcapitular (a: 12; m, h: 16) and adoral (6) setae setiform, roughened. Palp (length: 41–45) setation: 0–2–1–3–9 (+ω); solenidion swollen distally. Postpalpal seta (4) spiniform, smooth. Chelicera (length: 65–69) with two setiform, barbed setae (cha: 24; chb: 16).
Epimeral and lateral podosomal regions – Epimeral formula: 3–1–2–2; setae (c: 4; others: 10–12) setiform, thin, smooth. Humeral porose area Ah distinct; Am not observed. Pedotecta I, II represented by small laminae; pedotectum II rounded distally in ventral aspect. Discidium triangular, rounded distally. Circumpedal carina comparatively long, anteriorly reaching pedotectum II.
Anogenital region – Three pairs of genital (10–12), one pair of aggenital (10–12), two pairs of anal (10–12), and three pairs of adanal (12–14) setae setiform, thin, smooth. Adanal lyrifissure distinct. Marginal porose area not observed.
Legs – Claw slightly barbed on dorsal side. Porose area on all femora and on trochanters III, IV well visible; porose area on all tarsi and tibiae absent. Formulas of leg setation and solenidia: I (1–5–2–4–16) [1–2–2], II (1–5–2–4–15) [1–1–2], III (1–2–1–3–13) [1–1–0], IV (1–2–1–3–12) [0–1–0]; homology of setae and solenidia indicated in Table 1. Famulus short, slightly swollen distally, inserted posterior to solenidion ω2; seta d on femora III, IV inserted dorsobasally on segments. Solenidion ω1 on tarsus I, ω1 and ω2 on tarsus II and σ on genu III slightly bacilliform; other solenidia setiform.


Material examined — Holotype (male) and three paratypes (one male and two females): Philippines, Sibuyan Is., Romblon Province, Mt. Magdiwang, Mount Guiting-guiting Natural Park, leaf litter in forest (Museum number: JCBN 17 0330-2), 30.III.2017 (leg. O.L. Eusebio).
Type deposition — The holotype is deposited in the collection of the Senckenberg Museum of Natural History, Görlitz, Germany; three paratypes are deposited in the collection of the Tyumen State University Museum of Zoology, Tyumen, Russia. All specimens are preserved in 70% solution of ethanol with a drop of glycerol.
Etymology — The species name magdiwangensis refers ro the place of origin, Mt. Magdiwang.
Differential diagnosis — Euscheloribates magdiwangensis n. sp. is morphologically most similar to Euscheloribates mixtus (Mahunka, 1978) from Mauritius in having aggenital setae, clavate bothridial seta with short stalk and rounded rostrum, and in the absence of complete translamella and foveolate body ornamentation. However, the new species differs from the latter by smaller body size (length: 265–298 versus 320–325), in the presence (versus absence) of two short parts of translamellar line directed backwards, longest lamellar seta on the prodorsum (versus interlamellar seta longest), 10 pairs (versus one pair p1 developed, others represented by alveoli) of well visible notogastral setae and two pairs (versus three pairs) of setae on epimeres III and IV, and by localization of only one (versus two) genital seta in medioanterior part of genital plate.
Remarks
- Subías (2004, updated 2022) divided the genus Euscheloribates into nominate subgenus and the subgenus Euscheloribates (Trischeloribates) Hammer, 1971. His opinion (Subías 2020) is based on the presence (in Euscheloribates) or absence (in Trischeloribates) of aggenital setae. However, some other genera in Scheloribatidae (e.g. Muliercula Coetzer, 1968; Scheloribates Berlese, 1908; Perscheloribates Hammer, 1973) include species with or without aggenital setae; therefore, this morphological trait has no value to support the subgenera within Euscheloribates.
- Kunst (1958) described Euscheloribates and included it in Scheloribatidae. Later, several other related genera were proposed and synonymized with Euscheloribates (see Subías 2004, updated 2022): Mahunka (1978) described Tribates as a representative of Oribatulidae; Balogh and Balogh (1984) described Nanobates as a representative of Scheloribatidae; Balogh and Balogh (1992) described Corynobates as a representative of Scheloribatidae; Mahunka (2000) described Berndibula as a representative of Oribatulidae; and Mahunka (2009) described Ambrobates as a representative of Haplozetidae. To support establishment of these genera, some morphological traits were used by these authors, which are often used only at the species level (e.g. presence or absence of aggenital setae, number of setae on anal plate, morphology of rostrum). Due to this, a confusion arose, associated with the creation of invalid genera and incorrect placement in different families.
- Balogh (1970) described Birobates and included it in Oribatulidae. Aoki and Ohkubo (1974) placed Birobates in Oripodidae, and Balogh and Balogh (1984) included this genus into their newly proposed family Birobatidae. At present, this genus is considered as member of the family Oripodidae (Subías 2004; Colloff and Cairns 2011).
- Euscheloribates and Birobates are similar and do not have obvious differences and seem to be closely related; therefore, their independent generic status and placement in different families looks improbable. For example, based on their morphology, we have no doubt that E. guitingensis n. sp. and E. magdiwangensis n. sp. clearly belong to the same genus, but, according to the concept by the above-mentioned authors, they could have been included in either in Euscheloribates or in Birobates. The main problem is that most of the known species in these two genera (including the type species) are not well described, and there is no information on some important traits such as presence/absence of sublamella, sublamellar porose area, tutorium, keel-shaped ridge, humeral and marginal porose areas, and setation of legs. Thus, a detailed morphological revision is required, especially the type species to understand the generic status of Euscheloribates and Birobates, and to determine their respective family placement, and their species composition.
Acknowledgements
We cordially thank O.L. Eusebio for collecting materials used in this study; J.C.B. Naredo for facilitating transmittal of the specimens, together with other oribatids, to Russia; and two anonymous reviewers for valuable comments. This research was partially supported by the cooperative agreement No. FEWZ-2021-0004 from the Russian Ministry of Science and Higher Education.
References
- Aoki J., Ohkubo N. 1974. A proposal of new classification of the family Oripodidae (s. lat.), with description of new species. Bul. Nat. Sci. Mus., Tokyo, 17(2): 117-147.
- Balogh J. 1970. New oribatids (Acari) from New Guinea. II. Acta Zool. Acad. Sci. Hung., 16(3-4): 291-344.
- Balogh J., Balogh P. 1984. A review of the Oribatuloidea Thor, 1929 (Acari: Oribatei). Acta Zool. Hung., 30(3-4): 257-313. https://doi.org/10.1016/0305-4179(84)90004-4
- Balogh J., Balogh P. 1992. The oribatid mite genera of the World. Vol. 1. Budapest: Hungarian National Museum Press. 263 p.
- Bayartogtokh B., Shimano S. 2020. Oribatid mites of the families Scheloribatidae and Oripodidae (Acari, Oribatida) from Babeldaob Island of Palau. Ann. Zool, 70(3): 345-360. https://doi.org/10.3161/00034541ANZ2020.70.3.007
- Berlese A. 1908. Elenco di generi e specie nuove di Acari. Redia, 5: 1-15.
- Coetzer A. 1968. New Oribatulidae Thor, 1929 (Oribatei, Acari) from South Africa, new combinations and a key to the genera of the family. Mem. Inst. Invest. Cien. Moç., 9(A): 15-126.
- Colloff M.J., Cairns A. 2011. A novel association between oribatid mites and leafy liverworts (Marchantiophyta: Jungermanniidae), with a description of a new species of Birobates Balogh, 1970 (Acari: Oribatida: Oripodidae). Austral. J. Ent., 50: 72-77. https://doi.org/10.1111/j.1440-6055.2010.00782.x
- Corpuz-Raros L.A. 1979. Philippine Oribatei (Acarina). I. Preliminary list of species and descriptions of forty new species. Philip. Agr., 62(1): 1-82.
- Corpuz-Raros L., Ermilov S.G. 2019. Catalogue of oribatid mites (Acari: Oribatida) from the Malay Archipelago. Zootaxa, 4716(1): 1-240. https://doi.org/10.11646/zootaxa.4716.1.1
- Hammer M. 1971. On some oribatids from Viti Levu, the Fiji Islands. Det Kong. Dansk. Vidensk. Selsk. Biol. Skr., 16(6): 1-60.
- Hammer, M. 1973. Oribatids from Tongatapu and Eua, the Tonga Islands, and from Upolu, Western Samoa. Det Kong. Dansk. Vidensk. Selsk. Biol. Skr., 20(3): 1-70.
- Kunst M. 1958. Euscheloribates samsinaki n. g., n. sp., eine neue Moosmilbe aus der Tschechoslowakei (Acarina-Oribatei). Cas. opic. Ceskoslov. spol. ent., 55(1): 67-70.
- Mahunka S. 1978. Neue und interessante milben aus dem Genfer museum XXVII. A first survey of the oribatid (Acari) fauna of Mauritius, Reunion and the Seychelles I. Rev. suisse Zool., 85(1): 177-236. https://doi.org/10.5962/bhl.part.82228
- Mahunka S. 2000. Oribatids from Hong Kong III (Acari: Oribatida: Microzetidae and Oribatulidae) (Acarologia Genavensia XCV). Arch. Sci., Genève, 53(3): 177-184.
- Mahunka S. 2009. Oribatids from Madagascar IV (Acari: Oribatida). Rev. suisse Zool., 116(3-4): 337-352. https://doi.org/10.5962/bhl.part.81323
- Norton R.A. 1977. A review of F. Grandjean's system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal D.L. (Ed.). Biology of oribatid mites. Syracuse: SUNY College of Environmental Science and Forestry. pp. 33-61.
- Norton R.A., Behan-Pelletier V.M. 2009. Oribatida. In: Krantz G.W., Walter D.E. (Eds). A Manual of Acarology (TX). Lubbock: Texas University Press. Chapter 15. pp. 430-564.
- Subías L.S. 2004. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes, Oribatida) del mundo (1758-2002). Graellsia, 60 (número extraordinario): 3-305. https://doi.org/10.3989/graellsia.2004.v60.iExtra.218
- Subias L.S. 2020. Adiciones al listado mundial de ácaros oribátidos (Acari, Oribatida) (15ª actualización). Rev. Ibér. Aracnol., 36: 3-12.
- Subías L.S. 2022. Listado sistemático, sinonímico y biogeográfico de los Ácaros Oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles), 17ª actualización. 537 pp. Available from: http://bba.bioucm.es/cont/docs/RO_1.pdf
 (accessed March 2022).
(accessed March 2022). - Travé J., Vachon M. 1975. François Grandjean. 1882-1975 (Notice biographique et bibliographique). Acarologia, 17(1): 1-19.



2022-04-27
Date accepted:
2022-07-22
Date published:
2022-08-29
Edited by:
Pfingstl, Tobias

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Ermilov, Sergey G. and Corpuz- Raros, Leonila
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)