



A new species of Augeriflechtmannia (Prostigmata: Tetranychidae) from Haloxylon ammodendron (Amaranthaceae) in Iran and a key to the world species
Mahdavi, Sayed Mosayeb  1
; Latifi, Malihe
2
; Asadi, Mahdieh
3
and Auger, Philippe
4
1
; Latifi, Malihe
2
; Asadi, Mahdieh
3
and Auger, Philippe
4
1Department of Plant Protection, College of Agriculture, Vali-e-Asr University of Rafsanjan, Iran.
2✉ Department of Plant Protection, College of Agriculture, Vali-e-Asr University of Rafsanjan, Iran.
3Department of Plant Protection, College of Agriculture, Shahid Bahonar University of Kerman, Kerman, Iran.
4CBGP, INRAE, CIRAD, Institut Agro, IRD, Univ Montpellier, Montpellier, France.
2022 - Volume: 62 Issue: 4 pages: 898-907
https://doi.org/10.24349/4ry0-lfqdZooBank LSID: 62B46D0D-4836-44A6-8C4C-4B2D58B8FDC5
Original research
Keywords
Abstract
Introduction
Augeriflechtmannia was first described as Diplonychus Auger & Flechtmann in 2003. After that, Mahdavi et al. (2021) refined the genus diagnosis. It is a quite small genus of the tribe Tetranychini Reck, 1950 with only five species: A. armeniaca (Bagdasarian, 1951) collected from Amaranthus hybridus and Salsola dendroides (Amaranthaceae) in Armenia, A. astragalus Mahdavi, Asadi, Latifi & Seeman, 2021 collected from Astragalus sp. (Fabaceae) and A. iraniensis (Mahdavi & Ueckermann, 2013) collected from Salsola sp. (Amaranthaceae) (Mahdavi et al, 2013) both in Iran, A. jeridi (Auger & Flechtmann, 2003) collected from Arthrophytum schmittianum (Amaranthaceae) and A. penisinuosus (Auger & Flechtmann, 2003) collected from Cornulaca monacantha and Salsola sp. (Amaranthaceae) both in Tunisia.
In this paper, one more species of this genus, Augeriflechtmannia haloxyloni sp. nov. is described and illustrated based on females and males collected from Haloxylon ammodendron (Amaranthaceae) in Kerman Province, Iran. A key to the world species of Augeriflechtmannia is also provided.
Material and methods
Leaves and twigs of H. ammodendron (Fig. 1) were collected in bags and brought to the laboratory. The twigs were cut layer by layer and washed in a solution of water and commercial detergent. The dipping-washing-filtering method (Boller 1984) was used to remove the mites from plant material. The resulting solution was filtered through a sieve (400 Mesh), and the mites retained on the sieve were washed with 70% ethanol into a Petri dish, then collected individually under a stereomicroscope and mounted in Hoyer's medium. Mites were described with an Olympus® BX51 phase-contrast compound microscope. Measurements were done using an imaging system including DinoCapture® 2.0 software are given in micrometers (μm). The measurements of the holotype are given in parentheses after the range of measurements for the paratypes. The drawings were made using a drawing tube attached to a microscope with 1000x and 1600x magnifications. Final illustrations were drawn with Corel® draw x8 2016 and edited using Photoshop® CC 2017. The terminology and setal notations used for the description follow that of Lindquist (1985). Depositories are cited using the following abbreviations:

- SBUK —Collection of the Acarology Laboratory, Shahid Bahonar University of Kerman, Kerman, Iran.
- CBGP — Centre de Biologie pour la Gestion des Populations - Continental Arthropod Collection. https://doi.org/10.15454/D6XAKL
 .
. - QM —Queensland Museum, South Brisbane, Australia.
Family Tetranychidae Donnadieu, 1875
Subfamily Tetranychinae Berlese, 1913
Tribe Tetranychini Reck, 1950
Genus Augeriflechtmannia Koçak & Kemal, 2008
Diplonychus Auger & Flechtmann, 2003: 75. (Preoccupied)
Augeriflechtmannia Koçak & Kemal, 2008: 4
Augeriflechtmannia, Mahdavi et al., 2021: 703
Type-species: Augeriflechtmannia jeridi (Auger & Flechtmann, 2003).
Augeriflechtmannia haloxyloni sp. nov.
ZOOBANK: DF1218EE-9273-42AA-AE46-56AFB8A6C01F ![]()
(Figs. 2–8)
Type material
Male (holotype), 9 males, 11 females, two deutonymphs and two protonymphs paratypes ex. Haloxylon ammodendron (Amaranthaceae), Kerman-Kerman Province, Iran, 30°14′N 57°13′E, altitude 1835m a.s.l. and Rafsanjan-Kerman Province, Iran, 30°20′N 56°09′E, altitude 1616m a.s.l, on 15 October 2017 and 03 June 2021, respectively, coll. S. M. Mahdavi.
Type deposition
Holotype and all paratypes at SBUK, except one female and one male paratype at CBGP and one female paratype at QM.
Diagnosis
Male – Empodia I-II with two clawlike structures and a strong dorsomedian spur; empodia III-IV deeply split into two clawlike structures, each with two pairs of dorsal appendant hairs without dorsal spurs. Aedeagus turned upwards, without knob, ending in a short sigmoid distal part; axis of the proximal part of sigmoid end forming an acute to a slightly acute angle with the shaft axis. Female. Dorsal striae without lobes; setation of tibiae and tarsi I–IV tibiae 8+(1ω)-7-5-5, tarsi 13+(1ω)+2 dupl.-11+(1ω)+1 dupl.-9+(1ω)-9+(1ω).
Description
Male (Holotype) — (n=9; Figs. 2–4).

Dorsum – Color in living specimens light-yellow to orange with red eyes. Dorsal striae without lobes. Dorsal body setae slender and longer than distances between consecutive setae. Lengths of setae v2 41–55 (41), sc1 80–84 (84), sc2 46–49 (46), c1 59–68 (59), c2 64–72 (64), c3 55–64 (64), d1 62–64 (64), d2 66–68 (66), e1 60–63 (63), e2 62–65 (65), f1 53–57 (52), f2 41–44 (41), h2 23–30 (30).
Venter – Ventral striae without lobes. Lengths of setae: ps1 9 (9), ps2 9 (9), g1 11 (11), g2 11 (11), ag 20–25 (20), h3 18–20 (20).
Gnathosoma – Peritreme straight distally ending in a small bulb (Fig. 2e). Palp setation typical, spinneret (suζ) about 2.5 times as long as broad, 4 long, 1.5 wide, solenidion (ω) length 4 (Fig. 2f).
Legs – (Fig. 2a–b). Setation of legs I–IV: coxae 2-2-1-1, trochanters 1-1-1-1, femora 10-6-4-4, genua 5-5-4-4, tibiae 8+(4ω)-7-5-5, tarsi 13+(3ω)+2 dupl.-11+(1ω)+1 dupl.-9+(1ω)-9+(1ω); Empodia I–II with two clawlike structures and a strong dorsomedian spur (Fig. 2c); empodia III-IV deeply split into two clawlike structures, each with two dorsal appendant hairs without dorsal spurs (Fig. 2d). Tarsus I with four tactile setae (l′1, l″1, v′2, v″1) and two solenidia (ω′1, ω″2) well proximal to a proximal set of duplex setae, one solenidion (ω″1) and one tactile seta (v′1) at the level of proximal duplex setae (Fig. 2a). Tarsus II with one tactile seta (v′2) and one solenidion (ω″1) well proximal to duplex setae, two tactile setae (v′1, pv″) near to proximal duplex setae (Fig. 2b). Lengths of legs (from trochanter): I 168–170 (168), II 135–137 (135), III 137–140 (137), IV 153–160 (153).
Aedeagus – Holotype, (Figs. 3a–4). Aedeagus turned upwards, without knob, ending in a short sigmoid distal part, the axis of proximal part of sigmoid end forming an acute to slightly acute angle with the shaft axis. Aedeagus with some obvious variations in the length of sigmoid distal part (Fig. 3a–f; Fig. 4 and Table 1).




Female — (n=11; Figs. 5–8).


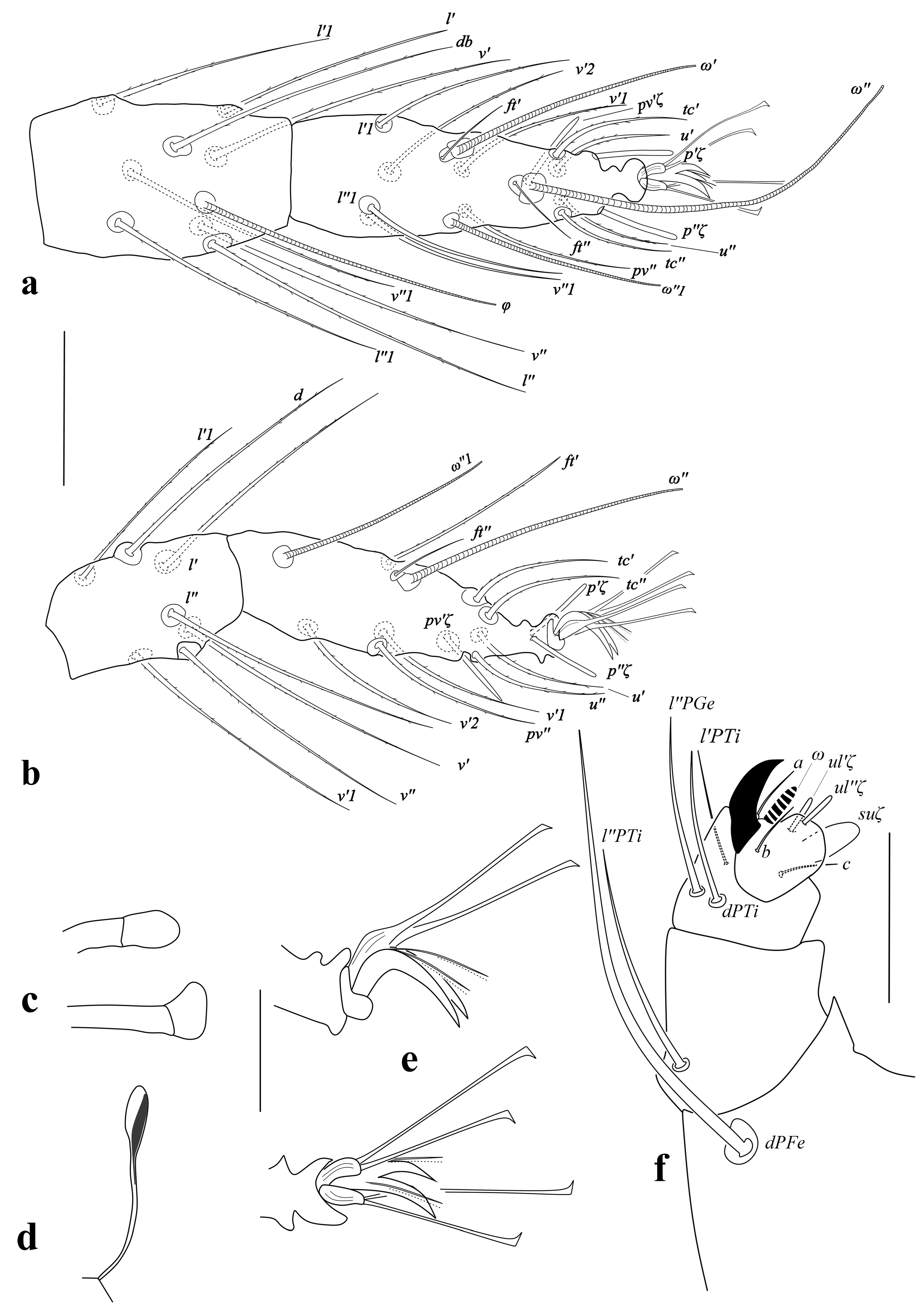
Length – (v2–h2) 270–281; width (c3–c3) 205–216.
Dorsum – Color in living specimens light-yellow to orange with red eyes. Dorsohysterosomal striae transverse between all dorsocentral setae (Fig. 5), without lobes (Fig. 6b). Dorsal body setae slender and longer than distances between consecutive setae and reach second seta behind. Lengths of setae: v2 56–60, sc1 101–117, sc2 54–74, c1 87–89, c2 87–89, c3 74–86, d1 84–89, d2 88–94, e1 81–83, e2 77–88, f1 77–82, f2 69–73, h2 33–51. Distances between dorsal setae: v2–v2 53–63, sc1–sc1 65–77, sc2–sc2 159–170, c1–c1 63–64, c2–c2 125–132, c3–c3 205–216, d1–d1 61–63, d2–d2 140–142, e1–e1 42–49, e2–e2 139–145, f1–f1 32–33, f2–f2 81, h2–h2 20–22.
Venter – Ventral striae without lobes; pregenital striae with irregular hourglass pattern (Fig. 8). Lengths of setae: 1a 35–40, 3a 32–39, 4a 38–45, ag 42–47, g1 26–28, g2 28–29, ps1 14–15, ps2 13–15, h3 25–30; distances between setae 1a–1a 31–33, 3a–3a 54–63, 4a–4a 39–42, ag 64–72. Spermatheca (Fig. 7d) ending in a small bulb.
Gnathosoma – Peritreme straight, distally ending in the small bulb (Figs. 5a, 7c); palp setation typical, spinneret (suζ) about 2 times as long as broad, 3–4 long, 1.5–2 wide, solenidion (ω) length 3 (Fig. 7f). Stylophore rounded, longer than wide.
Legs – (Fig. 7a, b). Setation of legs I–IV: coxae 2-2-1-1, trochanters 1-1-1-1, femora 10-6-4-4, genua 5-5-4-4, tibiae 8+(1ω)-7-5-5, tarsi 13+(1ω)+2 dupl.-11+(1ω)+1 dupl.-9+(1ω)-9+(1ω); empodia I–IV deeply split into two clawlike structures each with two pairs of dorsal appendant hairs without dorsal spurs (Fig. 7e). Tarsus I with four tactile setae (l′1, l″1, v′2, v″1) well proximal to proximal set of duplex setae, and one solenidion (ω″1) two tactile setae (v′1, pv″) at the level of proximal duplex setae (Fig. 7a). Tarsus II with one tactile seta (v′2) and one solenidion (ω″1) well proximal to duplex setae and two tactile setae (v′1, pv″) at the level of proximal duplex setae (Fig. 7b). Lengths of legs: I 180–199, II 155–161, III 177–181, IV 180–185.
Etymology
The specific epithet ''haloxyloni'' is derived from the specific name of the host plant Haloxylon ammodendron (Amaranthaceae).
Remarks
Augeriflechtmannia haloxyloni sp. nov. is similar to A. penisinuosus (Auger & Flechtmann, 2003) in global shape of the aedeagus and leg chaetotaxy, but it can be distinguished from the latter by three features: In males, 1) the aedeagus bear a long sigmoid distal part in A. penisinuosus (Fig. 3g) vs. short in A. haloxyloni sp. nov. (Fig. 3a–f); 2) empodia I-II are different in A. penisinuosus vs. empodia I–II are similar among all the specimens of A. haloxyloni we observed. In females: 3) the dorsal striae bear lobes in A. penisinuosus (Fig. 6a) vs. dorsal striae without lobes in A. haloxyloni.
As we found adults and juveniles together during sampling, indicating that the mite is active as reproduction takes place, the absence of lobes on the integument of female dorsal striations is a morphological feature specific to the new species that do not result from diapause-inducing environmental conditions experienced by the specimens. Indeed, despite the mite were collected in two sampling events, in June and October, all the specimens examined are are devoid of lobes on the dorsal integument.
Despite some obvious variations in the aedeagus length, we assumed that all the specimens belong to the same species. In our opinion, there are three lines of evidence to consider these variations as intraspecific: First, similar variations are found among specimens collected in the same host in both localities: Kerman and Rafsanjan (Figs. 3a–f and Table 1). Second, there are some intermediate forms between aedeagi with a longer and aedeagi with a shorter sigmoid distal end. Third, it is not possible to link variations in the morphology of the aedeagus to variations in other morphological characters or biological characteristics.
Key to the world species of Augeriflechtmannia
1. Male: tibiae I–IV with 8+(4ω)-7-5-5 setae
...... 2
— Male: tibiae setal formula not as above
...... 3
2. Female: dorsal striae without lobes. Male: empodia I-II similar (Fig. 2c)
...... A. haloxyloni sp. nov.
— Female: dorsal striae with lobes. Male: empodia I-II not similar
...... A. penisinuosus
3. Male: tibiae I–IV with 9+(3ω)-7-6-6 setae; knob of aedeagus with a slight angle to that of the shaft, the dorsal margin of knob curved, posterior projection about 1.6 times as long as anterior projection; knob twice as long as neck (Fig. 3i)
...... A. astragalus
— Male: tibiae setal formula not as above
...... 4
4. Male: tibiae I–IV with 10+(3ω)-7-6-7 setae; end of aedeagus almost rounded without indentation, bend dorsally at an angle of 90 degrees (Fig. 3j)
...... A. iraniensis
— Male: tibiae I–IV with 9+(5ω)-7-6-7 setae; aedeagus bent dorsally at a right angle, with a small acute anterior projection pointing upwards and a small posterior angulate projection (Fig. 3h)
...... A. jeridi
Note
As the male of A. armeniaca is unknown, this species is indistinguishable from other species of Augeriflechtmannia (Mahdavi et al. 2021). For this reason, it was not included in the key.
References
- Auger, P., Flechtmann, C. (2003). Two species of a new genus of Tetranychidae (Acari: Prostigmata) from south Tunisia. International Journal of Acarology, 29, 75-80. https://doi.org/10.1080/01647950308684324
- Bagdasarian, A. T. (1951). Contributions to the fauna of spider mites (fam. Tetranychidae) of Yerevan and its environs. Akad. Nauk Arm. S.S.R. Izv. Biol. S.Kh. Nauk, 4, 368-374.
- Berlese, A. (1913). Acarotheca Italica. Tipografia di M. Ricci, Firenze, Italy.
- Boller, E. E. (1984). Eine einfache Ausschwemm-methode zur schnellen Erfassung von Raubmilben, Trips und anderen Kleinarthropoden im Weinbau. Schweizerisch Zeitschrift fur Biologie, Obst- und Weinbau. 120, 77-111.
- Donnadieu, A. L. (1875). Recherches pour servir à l′histoire des Tétranyques. Thèse. Faculté des Sciences de Lyon. 134 pp. Also published in. Annales de la Société Linnéenne de Lyon. (n. ser.), 22(1876), 29-136.
- Koçak, A., Kemal, M. (2008). Nomenclatural notes on the genus-group names of the order Acarina. Miscellaneous Papers, 145, 1-6.
- Lindquist, E. E., Helle, W., & Sabelis, M. W. (1985). Spider mites: their biology, natural enemies, and control (Vol. 1A). Amsterdam, Elsevier Science Publishers, B.V., pp.3-28.
- Mahdavi, S. M., Asadi, M., Latifi, M., Seeman, O. D. (2021). New species of Augeriflechtmannia and Tetranychus (Acari: Trombidiformes: Tetranychidae) and a review of Augeriflechtmannia. Systematic Applied Acarology, 26(4), 698-712. https://doi.org/10.11158/saa.26.4.4
- Mahdavi, S. M., Asadi, M., Ueckermann, E. A., Farzan, S. (2013). A new species of Tetranychus Dufour, 1832 (Acari, Trombidiformes: Tetranychidae) from Iran. Systematic Applied Acarology, 18(3), 245-251. https://doi.org/10.11158/saa.18.3.7
- Reck, G. F. (1950). Spider mite fauna from Georgia (Tetranychidae: Acarina) Trudy Zoologicheskogo Instituta Akademia Nauk Gruz.S.S.R., 9, 117-134.



2022-04-16
Date accepted:
2022-08-08
Date published:
2022-09-09
Edited by:
Migeon, Alain

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Mahdavi, Sayed Mosayeb; Latifi, Malihe; Asadi, Mahdieh and Auger, Philippe
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)