



Ontogeny of South African intertidal oribatid mite species (Acari, Oribatida, Ameronothroidea) and supplements to adult morphology.
Hugo-Coetzee, Elizabeth A.  1
; Baumann, Julia
2
; Neethling, Jan-Andries
3
; Bardel-Kahr, Iris4
and Pfingstl, Tobias
5
1
; Baumann, Julia
2
; Neethling, Jan-Andries
3
; Bardel-Kahr, Iris4
and Pfingstl, Tobias
5
1✉ Terrestrial Invertebrate Department, National Museum, PO Box 266, Bloemfontein 9300, South Africa & Department of Zoology and Entomology, University of the Free State, Bloemfontein, South Africa.
2Institute of Biology, Karl-Franzens University of Graz, Universitaetsplatz 2, 8010 Graz, Austria.
3Terrestrial Invertebrate Department, National Museum, PO Box 266, Bloemfontein 9300, South Africa.
4Institute of Biology, Karl-Franzens University of Graz, Universitaetsplatz 2, 8010 Graz, Austria.
5Institute of Biology, Karl-Franzens University of Graz, Universitaetsplatz 2, 8010 Graz, Austria.
2022 - Volume: 62 Issue: 3 pages: 721-753
https://doi.org/10.24349/q6ks-5cs8Original research
Keywords
Abstract
Introduction
The marine intertidal oribatid mites of the superfamily Ameronothroidea are air-breathing animals living on the edge of the constant fluctuating cycle of low tide and high tide, of submersion and emersion and variable temperatures. Consequently, they have adapted to this environment in their morphology, behavior, reproduction, respiration and physiology (Bücking et al. 1998; Pfingstl 2013a, 2017). Ameronothroid mites live in the littoral zone in algae mats, rock crevices, on boulders, in barnacles and tubeworms, and man-made structures (Marshall and Pugh 2002; Pfingstl 2013a, 2017) or in estuaries and mangrove forests (Pfingstl et al. 2014). They feed mainly on algae, but also on lichens and cyanobacteria (Luxton 1966; Pugh and King 1985; Bücking et al. 1998; Pfingstl 2013a).
Ameronothroidea consists of four families namely, Ameronothridae, which occurs in the northern polar and cold temperate regions, the Podacaridae of the southern polar and cold temperate regions and the Fortuyniidae and Selenoribatidae of the subtropics and tropics (Procheş and Marshall 2001; Pfingstl and Schuster 2014; Pfingstl 2017; Norton and Franklin 2018). Four species from three families of Ameronothroidea have been recorded along the South African coast (Procheş and Marshall 2002; Pfingstl et al. 2021a). There was a record of a fifth species, namely Ameronothrus bilineatus, which was found in a salt marsh near Port Alfred (Weigmann 1975), but this occurrence was suggested to be introduced (Procheş and Marshall 2001) and since then there have been no further records of this species. However, the distribution of the four native species follows the general trend where Fortuyniidae and Selenoribatidae are limited to the subtropical regions of the east coast and the Podacaridae to the warm- to cold-temperate regions of the south coast (Procheş and Marshall 2002; Pfingstl 2016; Pfingstl et al. 2021a). Fortuynia elamellata micromorpha Marshall & Pugh, 2002 (Fortuyniidae) and Schusteria ugraseni Marshall & Pugh, 2000 (Selenorbatidae), were described from samples of 1998 and 1999 from barnacles, oyster shells and shale deposits on the mid- to upper rocky shores from Park Rynie, KwaZulu-Natal on the east coast of South Africa; Selenoribates divergens Pfingstl, 2015c (Selenoribatidae) was originally described from Egypt, and therefore the occurrence of this species along the sub-tropical east coast of South Africa (Pfingstl et al. 2021a) needs further investigation. Procheş and Marshall (2002) reported a Schusteria species (Schusteria sp. 2), from St. Lucia, which may refer to Sel. divergens, however since that material is lost, and the exact sample location unknown, the identity is assumed, but not confirmed. Halozetes capensis Coetzee & Marshall, 2003 (Podacaridae) was described from samples taken in November 2000 from finely-branched algae in the mid- and upper eulittoral zone from Kommetjie on the south-western coast and Nature's Valley on the southern coast. The Halozetes sp. reported in Procheş and Marshall (2002) supposedly refers to H. capensis. The descriptions of the adults of these species were sufficient, only omitting information or drawings of legs and the gnathosoma in some species. With the exception of the tritonymph of H. capensis, the juveniles of these species are unknown.
The description of oribatid juvenile morphology is often neglected. This can be attributed to several reasons such as the difficulty in associating juveniles with the adult in samples or museum collections (this association is even more difficult in high diversity groups such as the Oppioidea or Oripodoidea), or the small size and cryptic or endophagous behavior of juveniles (Norton and Ermilov 2014). Despite the scarcity of information of juveniles, it is an important aspect contributing to knowledge of ecology, systematic and phylogeny and even to homology and applications of nomenclature with regard to body and leg setation (Norton 1977; Norton and Ermilov 2014; Zhang 2018). Interestingly, juveniles of mites associated with aquatic and semi-aquatic or intertidal environments are the best known among the brachypyline oribatids, with juveniles of more than half of the Ameronothroidea known or partly known. Possible reasons include the medium to large size of species, the distinctive setae or sclerites on the juveniles (Norton and Ermilov 2014), excellent historic studies so that the general habitus of juveniles of these mites are well known (e.g., Grandjean 1955, 1966, 1968; Hammen 1963; Schuster 1963; Luxton 1967; Behan-Pelletier 1997) and the recent interest in littoral mites. Adults and juveniles of littoral mites are also often collected together because of their aggregation behavior facilitating the adult-juvenile association (Bücking et al. 1998; Krisper and Schuster 2008; Pfingstl 2013a). Currently, information of juvenile morphology is known for 69% of Fortyniidae, 37% of Selenoribatidae, 59% of Podacaridae and 63% of Ameronothridae species (see Appendix 1).
During a comprehensive project examining the biodiversity and distribution of intertidal mites along the South African coast, juvenile stages of all four species were collected. The main aim of the paper is to give supplementary descriptions of the adults and descriptions of all juvenile stages of F. elamellata micromorpha, Sel. divergens, Sch. ugraseni and H. capensis and to compare the juveniles with other known juveniles of the respective genus.
Material and methods
Sample collection
This study forms part of a larger study in which samples were taken at different locations along the coastline of South Africa (Pfingstl et al. 2021a). Fresh specimens for this study originated from Sheffield Beach, Umkomaas, Umdloti, St. Lucia (all KwaZulu-Natal), East London (Eastern Cape) and Nature's Valley (Western Cape) sampled in 2019 and 2020. In addition, previous collected specimens of F. e. micromorpha (collected in 1998-1999) and H. capensis (2000), housed at the National Museum, Bloemfontein, were used (these included holotypes, paratypes and juveniles). Localities and sample details are given in the respective species descriptions under: Material examined. Samples (app. 10 cm3) of littoral algae, barnacle and mussel shells were scraped off rocks with a knife or a small shovel and then put in Berlese-Tullgren funnels for 12 to 24 hours to extract mites. Most mites extract within 24 hours from these rather small samples with rare occurrences after this extraction time. Specimens were collected alive in small plastic containers with plaster of Paris, sorted with a fine brush and preserved in ethanol (100%) for morphological investigation. Juveniles were assigned to a specific species only if adults of a single species were contained in the respective sample. Specimens are stored in the Acarology collection of the National Museum, Bloemfontein, South Africa (abbreviated as NMB) and in the collection of the Senckenberg Museum für Naturkunde Görlitz.
Preparation and visual documentation
Specimens were mounted in lactic acid on temporary cavity slides for measurement and illustration. Body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the notogaster. Notogastral width refers to the maximum width in dorsal view. Lengths of body setae were measured in the aspect they could be best observed. All body measurements are presented in micrometers. In most instances the range of sizes are given, but for body measurements the range is given followed by the mean in brackets. Drawings were made with a camera lucida using a Nikon transmission light microscope ''Nikon Eclipse 50i″. Drawings were made with software CorelDraw 2021. For photographic documentation, specimens were air-dried and photographed with a Keyence VHX-5000 digital microscope using automated image stacking.
Morphological terminology used in this paper follows Grandjean (1966, 1968) and Norton and Behan-Pelletier (2009); Norton (1977) for setal nomenclature; Norton and Franklin (2018) for notation of epimeral setae (e.g. Remark 15, setae are named according to their relative position and not to their ontogenetic appearance).
Abbreviations
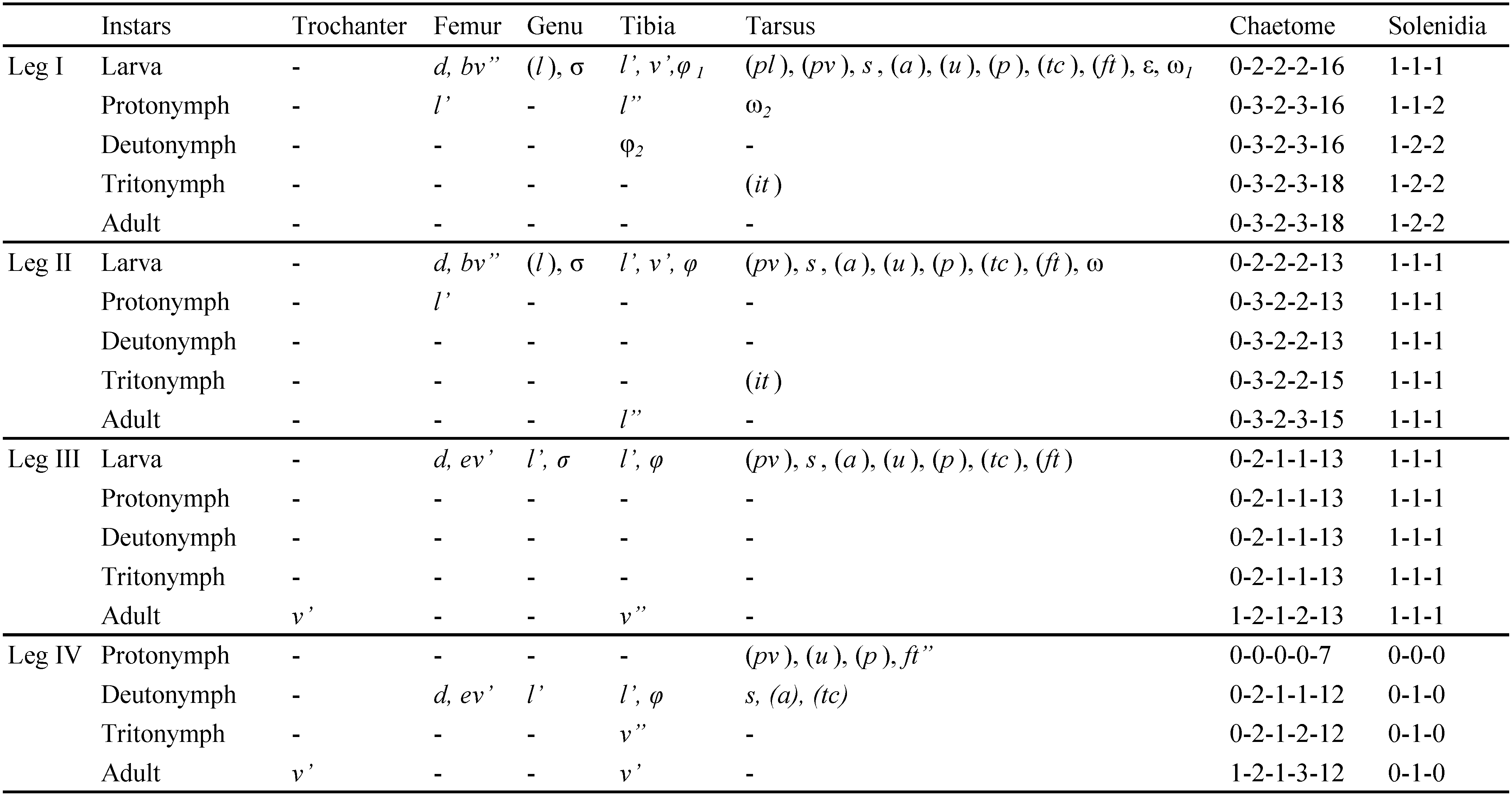
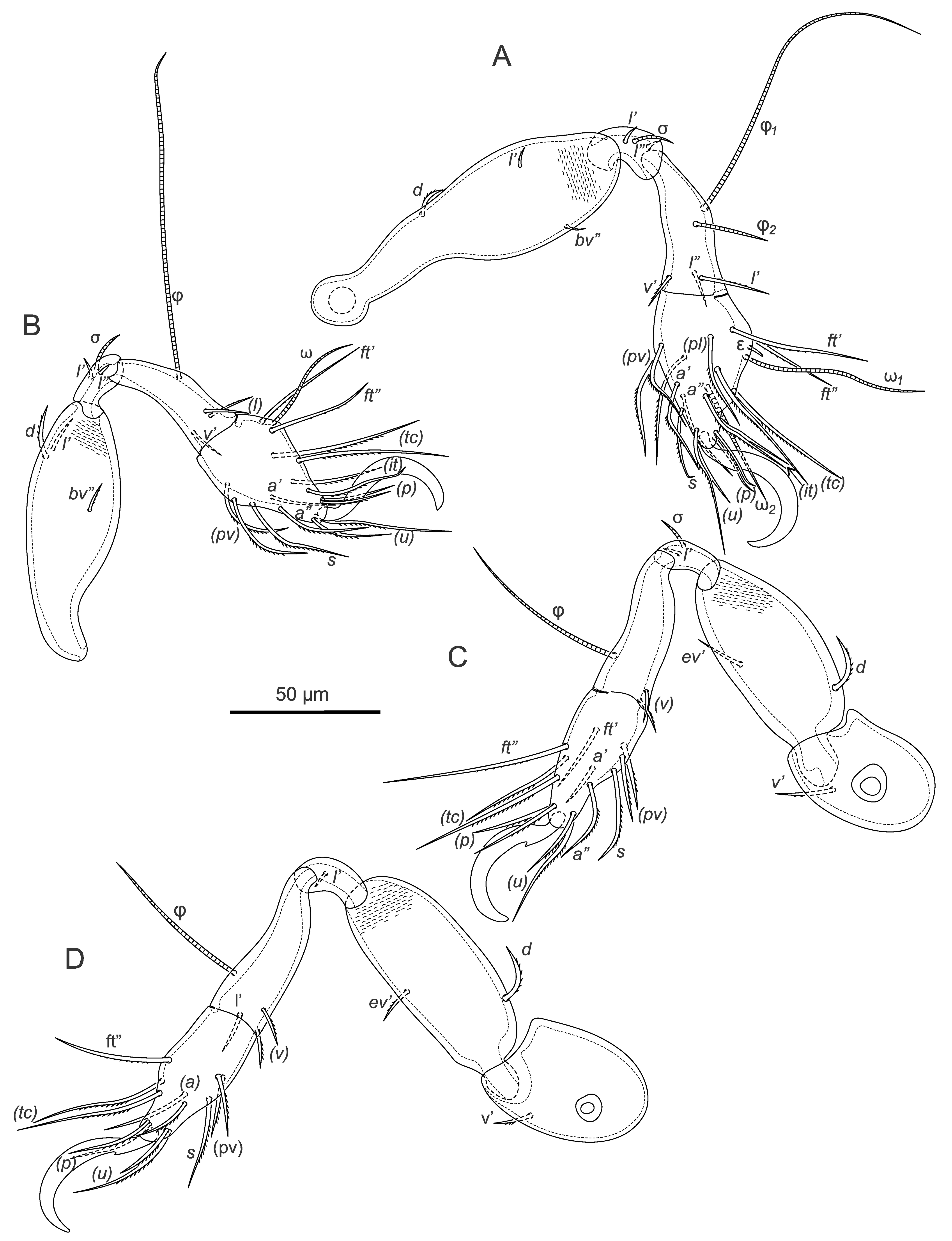
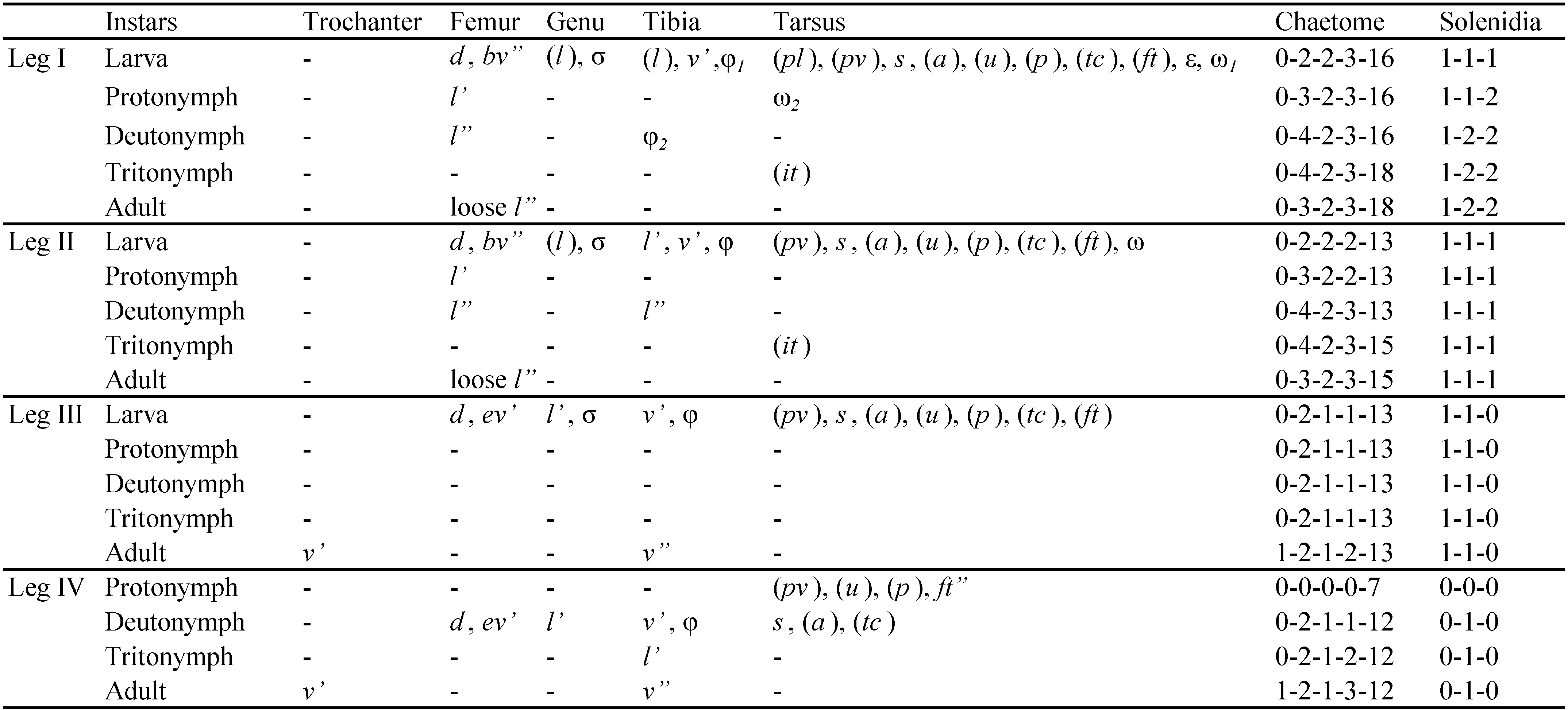
The following abbreviations are used in the text, figures and tables: prodorsum: ro, le, in, bs, ex - rostral, lamellar, interlamellar, bothridial and exobothridial setae, respectively; notogaster: c, da, dm, dp, la, lm, lp, h, p - notogastral setae; ia, im, ip, ih, ips - lyrifissures; gla - opisthonotal gland opening; gnathosoma: a, m, h - subcapitular setae; cha, chb - cheliceral setae, Tg - Trägårdh's organ; epimeral and lateral podosomal regions: 1a, 1b, 1c, 2a, 3a, 3b, 3c, 4a, 4b - epimeral setae; PdI, PdII - pedotecta I and II respectively; ce, ci - prodorsal canals of van der Hammen's organ; dis - discidium; Cl - Claparède's organ; s1 , s2 - enanthiophyses; anogenital region: g, ag, an, ad - genital, aggenital, anal, and adanal setae, respectively; iad - adanal lyrifissure; β - tendon β; legs: Tr, Fe, Ge, Ti, Ta - leg trochanter, femur, genu, tibia, and tarsus, respectively; pa - leg porose area; ω, σ, φ - leg solenidia; ɛ - leg famulus; d, l, v, ev, bv, ft, tc, it, p, u, a, s, pv, pl - leg setae.
Results
Family Fortuyniidae Hammen, 1963
Genus Fortuynia Hammen, 1960
Fortuynia elamellata Luxton, 1967
Fortuynia elamellata micromorpha Marshall & Pugh, 2002
Adults


Measurements: females (n=2) length 399-414 (407), width 202-218 (210), males (n=5) length 392-408 (399), width 201-220 (211); holotype (female) length 397, width 190, paratypes (females) (n=3) length 398-408 (403), width 202-209 (206); body sizes in range of the specimens of the original description, females (n=7) length 380-404, width 206-215.
Integument (Figures 1 and 2): brown to dark brown, genital plate slightly darker, legs with slight darkening of proximal part of tarsi, distal part of tibiae, distal two thirds of femora and entire genua and trochanters; body surface finely punctate, lighter areas around insertions of notogastral setae; granulate in acetabular regions; chelicerae punctate.
Prodorsum (Figures 1A, C): rostrum broadly rounded, slightly flattened apex in dorsal view; no lamellar ridges; ro (35-41), le (26-35), smooth, in, ex represented by alveoli; bs (30-36) smooth, medially incurving, with clavate head; bothridium cup-like; transverse line developed posterior to insertion of alveoli of ro.
Gnathosoma (Figure 2E): Setae a, m, h thin, smooth; pedipalp setation 0-2-1-3-9(+ω), ω on tarsus erect, not associated with eupathidium acm; femur with large porose area; chelicera with teeth, smooth seta chb shorter than barbed seta cha.
Notogaster (Figures 1A, C): elongate oval in dorsal view; lenticulus anteriorly with irregular border; 14 pairs of smooth, setiform notogastral setae, relatively long, h2 longest, c2 , p3 shortest (c1 60-73, c2 32-40, da, dm, dp 47-67, la, lm, lp 43-70, h1 35-50, h2 81-104, h3 50-67, p1 58-85, p2 34-53, p3 30-37), c3 absent; five pairs of distinct lyrifissures present, ia postero-lateral to c2 , im between lm and lp, ih far lateral to h3 , ip and ips lateral to p2 ; gla lateral to im.
Lateral aspect (Figure 1C): PdI round, small; cuticular canals of van der Hammen's organ present, canal ce very short, reaching posterior border of bothridium, canal ci absent.
Ventral (Figures 1B, C): all epimeral setae setiform, smooth, setation 3-1-3-2, seta 1b (22-32) slightly longer than others (13-23); discidium rounded; five pairs of genital (g1 19-23, g2-5 16-19), one pair of aggenital (ag 15-19), two pairs of anal (an1 26-35, an2 20-31) and three pairs of adanal (ad1 40-54, ad2 27-36, ad3 22-27) setae thin, smooth; lyrifissure iad medially to ad2 , slightly obliquely orientated; anal plates slightly triangular (length 81-86, width 29-33), preanal organ triangular; genital plates trapezoid, large (length 65-80, width 38-49, no distinct difference in size between males and females but plates in females by trend slightly larger).
Legs (Figures 2A-D): monodactylous, long hook-like claws with small teeth dorsally; porose areas pa on Fe I-IV and Tr III, IV distinct; tarsi with proximal lyrifissure; famulus short, straight, weakly blunt-ended; all solenidia long, setiform; for leg setation and solenidia see Table 1.


Juvenile stages
Common features – apheredermous; dark brown in colour, integument plicate and soft except for the more sclerotised centrodorsal plate, median posterior of centrodorsal plate with a less sclerotised area forming a narrow inverted ′v′ (except in larva where the plate is uniform in sclerotisation); body finely granulate, with more distinct granulation around insertions of dorsal gastronotal setae and in furrows and acetabular regions, granulation on prodorsum more distinct in the larva and deutonymph; prodorsum triangular, hysterosoma oval in dorsal view; rostrum broadly rounded; le setiform, short, smooth, in represented by alveoli; bothridia small, cup-like, bs with short stalk and smooth head clavate in lateral view, globose in dorsal view; ventral side with specific pattern of folds i.e., folds between epimeres 2 and 3, anterior and posterior of genital area and lateral to anal orifice, with pores within these folds leading into tracheal tubes; legs monodactylous with large hook like claws, with small teeth dorsally; porose areas on legs as in adults; for leg setation and solenidia see Table 1.
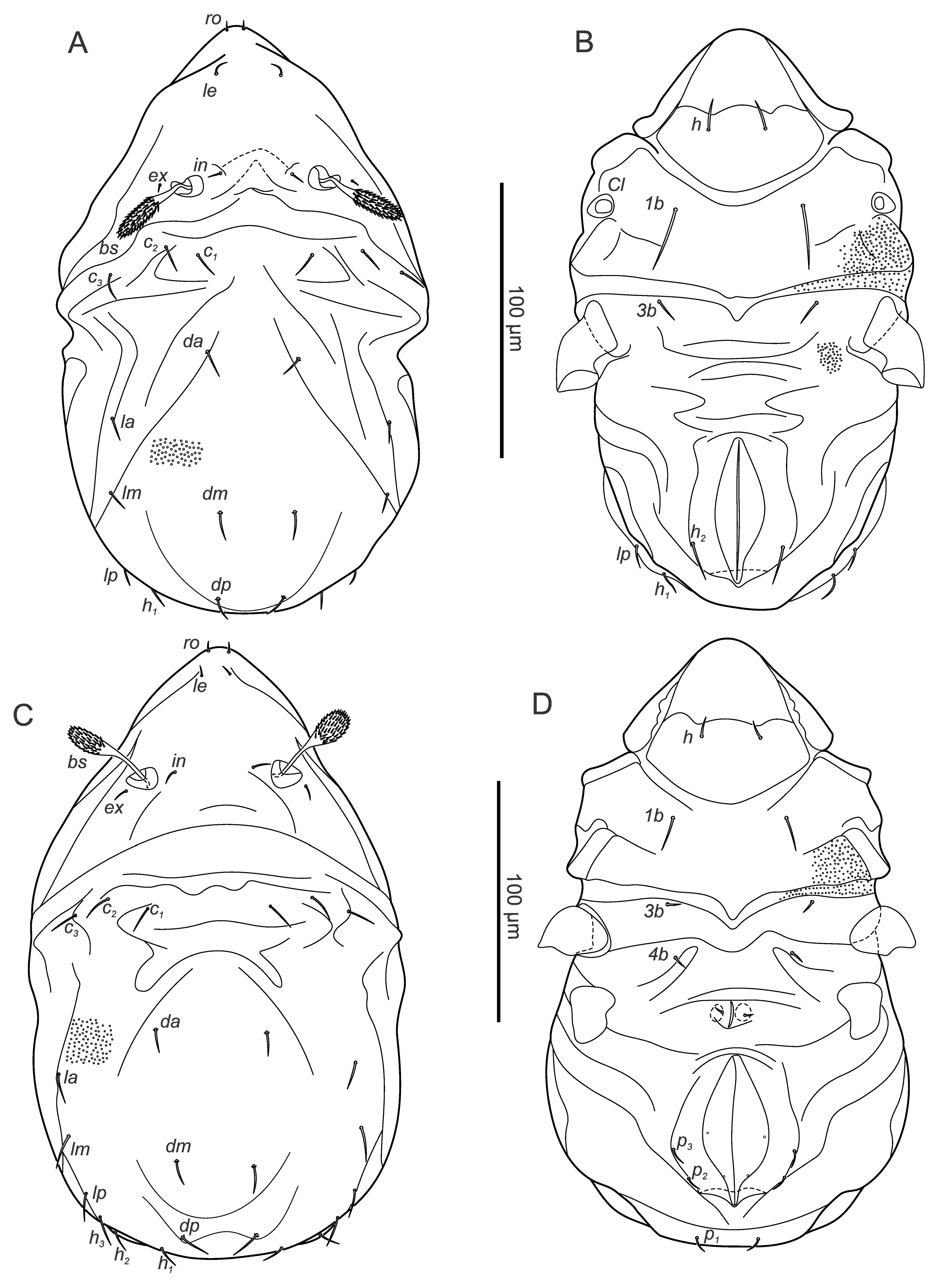
Larva – (Figures 3A, B) (n=7) length: 175-224 (201), width: 103-139 (118).
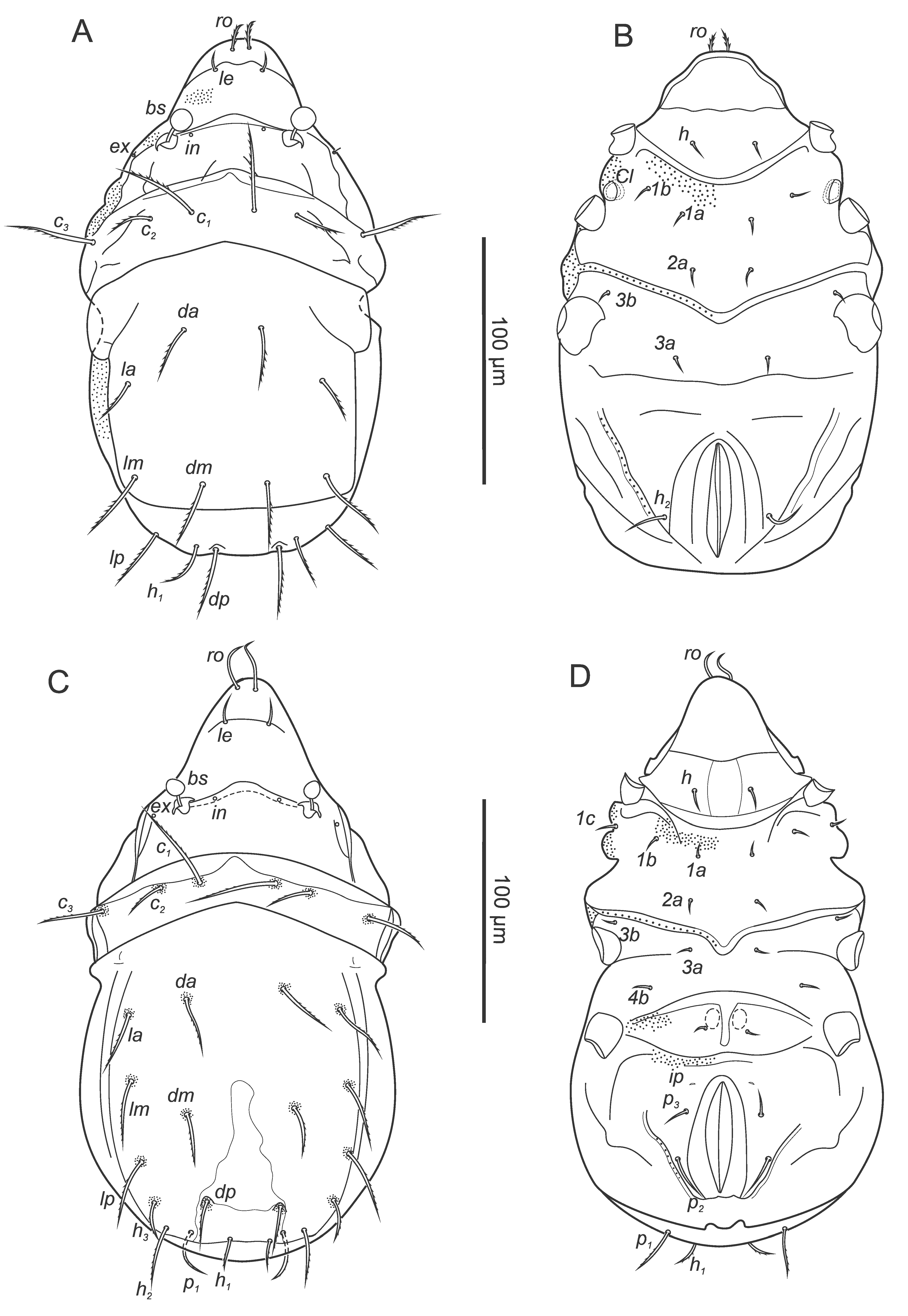
Gastronotic region: ro setiform, short, barbed, ex minute; 11 pairs of gastronotal setae (c1-3 , da, dm, dp, la, lm, lp, h1-2 ) setiform, barbed, except smooth seta h2 ; h2 lateral to anal folds on ventral side.
Ventral region of idiosoma: epimeral setation 2-1-2 (1a, 1b, 2a, 3a, 3b), setiform, smooth, small; Claparède's organ present, no protective seta observed.
Protonymph – (Figures 3C, D) (n=9) length 234-273 (255), width 112-147 (131).
Gastronotic region: ro setiform, smooth, curved, ex represented by alveolus; 15 pairs of gastronotal setae (h3 , p1-3 added), setiform, finely barbed (except p2-3 smooth), p2-3 lateral to anal opening; seta c1 longer than other setae.
Ventral region of idiosoma: epimeral setation: 3-1-2-1 (1c, 4b added), setae setiform, short, one pair of short genital setae; cupule ip anterior to anal valves distinct.
Deutonymph – (Figures 4A, B) (n=13) length 291-327 (313), width 134-176 (157).

Gastronotic region: ro setiform, smooth, ex represented by alveolus; 15 pairs of setiform, roughened gastronotal setae, except p2-3 smooth, p2-3 lateral to posterior part of anal folds, h2 longest.
Ventral region of idiosoma: epimeral setation: 3-1-2-2 (4a added); two pairs of short genital setae, one pair of aggenital setae, three pairs adanal setae lateral to anal opening, ad1 longest, two vestigial pairs of anal setae; cupule iad anterior to anal opening.
Tritonymph – (Figures 4C, D, supplementary figure 1) (n=4) length 381-400 (391) width 177-215 (198).
Gastronotic region: ro setiform, smooth, ex represented by alveolus; 15 pairs of setiform, smooth gastronotal setae, c1 , h2 longest, p2-3 ventrally, distanced from anal orifice.
Ventral region of idiosoma: Epimeral setation: 3-1-3-2 (3c added), four pairs of setiform genital, one pair of aggenital, three pairs of adanal, two pairs of anal setae; cupule iad anterior to anal valves.
Material examined
Holotype (NMB 4734.1.1) and three paratypes (NMB 4734.1.2) from Park Rynie, KwaZulu-Natal, South Africa, 30°21′S, 30°43′E, from barnacles, oyster shells and shale deposits, mid to upper rocky shore, 1998-1999, collected by D.J. Marshall and K. Ugrasen (in the original description the holotype should have been deposited elsewhere, but was instead recently sent to NMB); six adults, four larvae, seven protonymphs, six deutonymphs, one tritonymph from Sheffield Beach, KwaZulu-Natal, South Africa, 29°29.737′S, 32°15.127′E, from Bostrychia sp. algae from rock, upper eulittoral zone, 27 Feb. 2020, collected by T. Pfingstl (NMB 4717.1); one adult, one larva, four deutonymphs, two tritonymphs from Umkomaas, KwaZulu-Natal, 30°12.672′S, 30°48.00′E, from Gelidum sp. algae from tubeworms and rock in the lower eulittoral zone, 02 Mar. 2020, collected by T. Pfingstl (NMB 4720.1, NMB 4721.1); two larvae, two protonymphs, three deutonymphs and one tritonymph from Umdloti, KwaZulu-Natal, 29°40.560′S, 31°6.957′E, from Gelidum sp. algae on rock, barnacles, mussels, lower to median eulittoral zone, 29 Feb. 2020, collected by T. Pfingstl (NMB 4719.2).
Remarks
The original description of the adult by Marshall and Pugh (2002) provides good morphological information and the drawings are detailed, but there are no drawings of the lateral side of the body or the legs. Also, canal ce is present, but ci is absent, whereas the authors stated both are absent. The adults of the other locations match the adults of Park Rynie of the original description.
Some or all juvenile stages of several species of Fortuynia are known, namely F. arabica Bayartogtokh et al., 2009, F. atlantica Krisper & Schuster, 2008, F. churaumi Pfingstl et al., 2019, F. dimorpha Pfingstl, 2015, F. elamellata elamellata Luxton, 1967, F. maledivensis Pfingstl, 2015, F. longiseta Pfingstl, 2015, F. rotunda Marshall & Pugh, 2002, F. shibai Aoki, 1974, F. smiti Ermilov et al., 2013, F. taiwanica Bayartogtokh et al., 2009 and F. yunkeri Hammen, 1963 (see Appendix 1).
The morphology of most Fortuynia juveniles is similar in habitus and development of setation, except for the juveniles of F. arabica and F. taiwanica (Bayartogtokh et al. 2009), which may be attributed to several factors (see Pfingstl et al. 2021b). For the other 10 species, the main differences lie in the length and morphology of setae (Pfingstl and Schuster 2012a) and other minor characteristics such as ontogeny of certain leg setae, especially on the femora. One characteristic of the larva of F. elamellata micromorpha that is distinctly different is the strongly barbed rostral setae. The juveniles of F. elamellata elamellata are not very well described (Luxton 1967), but in general terms, besides the larger sizes they seem to be similar to F. elamellata micromorpha. The only difference is in the placement of the lamellar setae in the larva which is very close together in the nominate species and further from each other in the subspecies. However, only one badly damaged larva specimen was used for the description of F. elamellata elamellata.
Family: Selenoribatidae
Genus: Selenoribates Strenzke, 1961
Selenoribates divergens Pfingstl, 2015
Adult

Measurements: females (n=2): length 285-293 (289), width 157-175 (166), males (n=8): length 269-284 (279), width 162-174 (166); measurements in the original description of specimens from Egypt within range of South African specimens, mean length 273, mean width 164.
Integument (Figure 5): brown in colour, antero-medial notogaster with lighter area, ventrally area posterior to genital plates darker than anterior to this area, genital and anal plates same colour as this posterior area, trochanters III and IV of legs darker than rest of legs; body densely granulate, larger granules irregularly sparsely distributed, denser bigger granules in acetabular region, legs with small dense granules.
Prodorsum (Figures 5A, C): rostrum with small lateral humps and rounded median apex; lamellar ridge absent; short line extending from bothridium anteriorly; ro (8-9), in, ex (3-4) short, simple, le branched in two sub-equal parts (10-16); bothridium large cups opening laterally, with postbothridial tubercle; bs with medium long stem and clavate, spinose head (30-41).
Gnathosoma: same as in Pfingstl (2015c).
Notogaster (Figures 5A, C): oval in dorsal view; dorsosejugal furrow incomplete; tubercle present antero-laterally, posterior to bothridial tubercle; two x-shaped ridges antero-laterally, slightly anterior of seta c1 ; 14 pairs of notogastral setae (c1-2 , da, dm, dp, la, lm, lp, h1-3 , p1-3 ), branched basally, branches sub-equal in length (10-20), c3 absent; five pairs of lyrifissures distinct: ia in humeral region, im anterior to la, ih lateral to lp and h3 , ip lateral to p3 close to lateroventral border of notogastral plate, ips laterally to p2 and p3 ; gla lateral to la.
Lateral aspect (Figure 5C): PdI directed anteriorly, rounded; discidium pointed; laterosejugal furrow developed as a groove, extending from s1 to bothridium; enantiophyses (s1 , s2 ) present as strong opposite triangular projections.
Ventral region (Figure 5B): median sternal cavity on epimeron I demarcated by a ridge; all setae smooth, simple; epimeral setation 1-0-1-1, seta 1b very long (33-38)< 3b (12-14)< 4b (7-9); three pairs of genital (5-7), two pairs of anal and three pairs of adanal setae (6-9); genital plate (length 32-40, width 19-23) and anal plate (length 56-63, width 20-24) large; pre-anal organ triangularly rounded in ventral view; lyrifissure iad parallel and adjacent to anterior lateral border of anal plate.
Legs: see drawings in Pfingstl (2015c); monodactylous, long hook-like claws with proximo-ventral tooth; femora with ventral carinae; porose areas absent; solenidion φ1 on Ti I long, but not necessarily curving backwards; for setation and solenidia see Table 2.
Juvenile stages
Common features – apheredermous; light brown in colour, integument strongly plicate and soft, except for the more sclerotised centrodorsal plate; body finely granulate (less distinct in larva), with more distinct granulation in the acetabular regions; prodorsum triangular, hysterosoma oval in dorsal view; rostrum with weak lateral humps and rounded apex; le, ro setiform, short, smooth, simple; in and ex very short; bothridia small, cup-like, bs with medium stalk and clavate spinose head; plicate centrodorsal shield bearing central dorsal setae, da, dm, dp on small apophyses; no cupules discernable; folds lateral to anal orifice; epimeral seta 1b very long; legs monodactylous with hook like claws, small tooth proximo-ventrally on each claw; leg porose areas not visible; for leg setation and solenidia see Table 2.
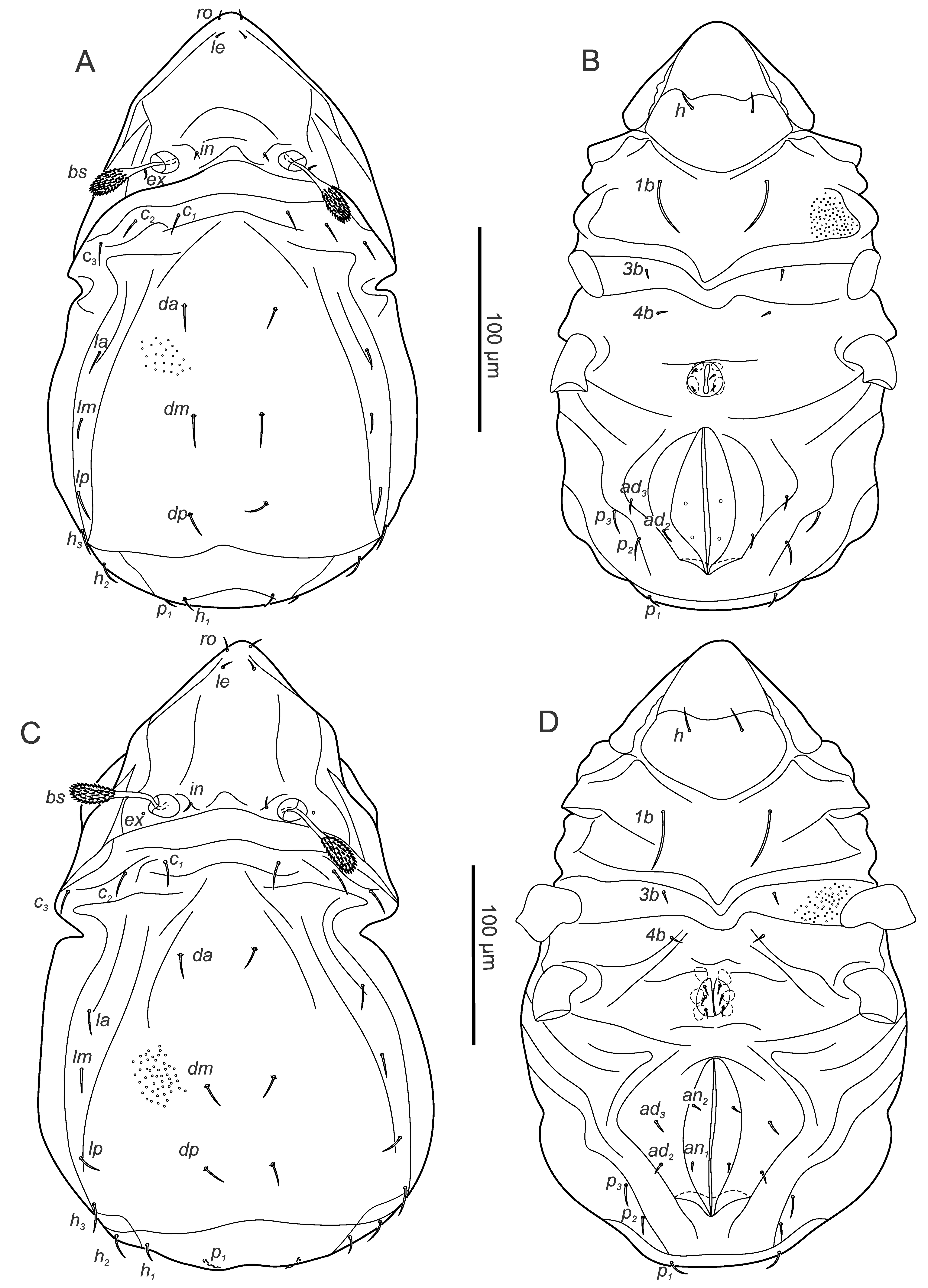
Larva – (Figures 6A, B) (n=1) length: 170, width: 109.

Gastronotic region: 11 pairs of simple gastronotal setae (c1-3 , da, dm, dp, la, lm, lp, h1-2 ), setae da, dm, dp setiform, thickened, barbed, setae h1 , h2 slightly thickened, other setae thin setiform, smooth; h2 lateral to anal folds on ventral side.
Ventral region of idiosoma: epimeral setation 1-0-1-0 (1b, 3b), setiform, smooth, 3b short; Claperède's organ present, not covered by protective seta.
Protonymph – (Figures 6C, D) (n=9) length 163-199 (178), width 87-108 (100).
Gastronotic region: 15 pairs of simple gastronotal setae (h3 , p1-3 added), setiform, smooth, seta h2 of similar length as setae da, dm, dp; p2-3 lateral to anal folds.
Ventral region of idiosoma: Epimeral setation: 1-0-1-0, seta setiform; one pair of short genital setae.
Deutonymph – (Figures 7A, B) (n=1) length 235, width 138.

Gastronotic region: 15 pairs of gastronotal setae, smooth, setae da, dm, dp branched with one small and one larger branch, setae la, lm, lp, h1-3 , p1 branched into sub- equal branches, setae c1-3 , p2-3 simple setiform, p2-3 ventrally, posterior to anal folds.
Ventral region of idiosoma: epimeral setation: 1-0-1-1 (4b added); two pairs of short genital setae, three pairs of short setiform adanal setae lateral to anal valves, two vestigial pairs of anal setae.
Tritonymph – (Figures 7C, D, supplementary figure 2) (n=8) length 245-268 (257), width 139-159 (148).
Gastronotic region: 15 pairs of gastronotal setae (as in deutonymph).
Ventral region of idiosoma: epimeral setation as in deutonymph; three pairs of setiform genital, three pairs of adanal, two pairs of anal setae.
Material examined
Ten adults and 19 juveniles from Mission Rocks, iSimangaliso Wetland Park, St. Lucia, KwaZulu-Natal, South Africa, 28°16.708′S, 32°29.143′E, from Bostrychia sp. algae, median eulittoral zone, 3 Mar. 2020, collected by T. Pfingstl (NMB 2722.1).
Remarks
The original description of this species with material from Egypt provides very good morphological information and drawings of all aspects including the legs (Pfingstl 2015c). The South African specimens are similar in morphology to the Egyptian specimens, except for the presence of small lateral humps on the rostrum, the small tubercle antero-laterally on the notogaster and lyrifissure ih not located on the lateroventral border of the notogaster. Considering the large geographic distance between Egypt and South Africa, there is a possibility that these differences are indicative of a closely related but genetically distinct new species. However, the differences are very subtle and could also be a result of geographic variation. Without genetic data, we cannot conclude that the South African specimens represent a new species.
Some juvenile stages of three other species are known, namely S. satanicus Pfingstl, 2013 (larva), S. quasimodo Pfingstl, 2013 (proto- and tritonymph) and S. mediterraneus Grandjean, 1966 (deuto- and tritonymph) (Grandjean 1966; Pfingstl 2013b; see Appendix 1). The larvae of S. divergens and S. satanicus are similar in general habitus and morphology of setae and integument, but differ in the form of the bothridial seta which is with a medium long stem and clavate spinose head in the former and long, flagelliform in the latter (also different in adults).
The protonymph and tritonymph of S. divergens and S. quasimodo differ most noticeably by the number of gastronotal setae and many more gastronotal folds in the latter. The nymphs of S. divergens have 15 pairs of setae, whereas the protonymph of S. quasimodo has 24 pairs and the tritonymph 44 pairs stemming from a multiplication of certain setae. Furthermore, the setae in the tritonymph of S. divergens are branched, while the setae in S. quasimodo are setiform, with setae da, dm, dp thickened and serrated. Setae ex is minute in S. divergens and vestigial in S. quasimodo. There is only one difference on the leg setae in the number of setae on Fe II (S. divergens proto- and tritonymph: 3 setae, S. quasimodo protonymph: 2, tritonymph: 4). Other characteristics, which also differ in the adults, are the form of the bothridial setae (S. divergens head clavate, spinose, S. quasimodo flagelliform) and the number of adanal setae (S. divergens three pairs, S. quasimodo two pairs).
In general, the deuto- and tritonymph of S. divergens and S. mediterraneus look similar in habitus and gastronotal folds. An obvious difference is that S. divergens has branched gastronotal setae with setae da, dm, dp slightly thickened while S. mediterraneus has setiform setae and all setae are thin. There are differences on the leg setae in the number of setae on Fe I and II (S. divergens: 3 setae, S. mediterraneus: 4). Other characteristics, which also differ in the adults, is the form of the bothridial setae (S. divergens: head clavate, spinose, S. mediterraneus: spatuliform) and the number of adanal setae (S. divergens: three pairs, S. mediterraneus: two pairs).
Family: Selenoribatidae Schuster, 1963
Genus: Schusteria Grandjean, 1968
Schusteria ugraseni Marshall & Pugh, 2000
Adult

Measurements: females (n=9): length 368-389 (378), width 227-244 (227), males (n=9): length 351-394 (369), width 220-245 (229); neotype (female): length 384, width 236; no difference in size between individuals from the two locations (Sheffield Beach and Winterstrand); measurements in the original description are within close range, females: length 331, 334, width 205, 220.
Integument (Figures 8 and 9): brown in colour, antero-medial notogaster with lighter area, ventrally the area posterior to genital plates darker than the anterior area, genital and anal plates dark with small foveolae; legs lighter brown, except trochanter III and IV dark brown; femora distally with faint striae; body distinctly granulate with large granules interspersed with small granules, larger granules on PdI.
Prodorsum (Figures 8A, C): rostrum rounded in dorsal view; ro, le, in short (6-11), fine, smooth, ex as alveolus; bothridium, large, rectangular with small lateral incision and small opening for bs; bs with relative long stem and clavate, spinose head (38-55).
Gnathosoma: similar to S. littorea (see Grandjean 1968).
Notogaster (Figures 8A, C): oval in dorsal view; dorsosejugal furrow incomplete medially; 15 pairs (c1-3 , da, dm, dp, la, lm, lp, h1-3 , p1-3 ) fine, short, smooth setae, all of similar length (10-18), c3 not necessarily closer to c1 than to c2 as in original description, but indeed c3 closer to lyrifissure im than to ia; five pairs of distinct lyrifissures: ia next to anterior lateral border of notogaster, im posterior to c3 , ih laterally to lm and lp, ips laterally to p3 , ip between p2 and p3 ; gla postero-laterally to im.
Lateral aspect (Figure 8C): PdI directed anteriorly, rounded; discidium rounded triangularly.
Ventral (Figures 8B, C): all setae smooth, fine; epimeral setation 1-0-1-1, seta 1b very long (43-56) < 3b (8-11)< 4b (6-8); three pairs of genital (5-7), two pairs of anal and two pairs of adanal (7-8) setae; lyrifissure iad parallel, antero-laterally to anal plates; setae ad3 posterior to iad, ad2 and ad3 close together; genital plates (length 49-60, width 26-30) and anal plates (length 79-86, width 29-33) large; pre-anal organ triangular in ventral view.
Legs (Figures 9A-D): monodactylous, hooked claw with small tooth proximo-ventrally; most setae barbed, except thickened blunt antiaxial lateral seta on genua I-IV; Fe I, II with only three setae (as in S. littorea), on tarsi seta u′ longer than u″; for setation and solenidia see Table 3.
Juvenile stages
Common features – apheredermous; light brown in colour, integument strongly plicate and soft, except for the more sclerotised centrodorsal plate; body strongly granulated; prodorsum triangular, hysterosoma oval in dorsal view; rostrum rounded; ro, le, in setiform, short, smooth, fine; bothridia small, cup-like; bs with medium stalk and clavate spinose head; plicate centrodorsal shield bearing central dorsal setae, da, dm, dp on very small apophyses; all gastronotal setae simple, fine, short; no cupules discernible; folds lateral to anal orifice; epimeral seta setiform, smooth, 1b very long, seta 3b shorter; legs monodactylous with hooked claw and small tooth proximo-ventrally; leg porose areas not visible; for setation and solenidia see Table 3.
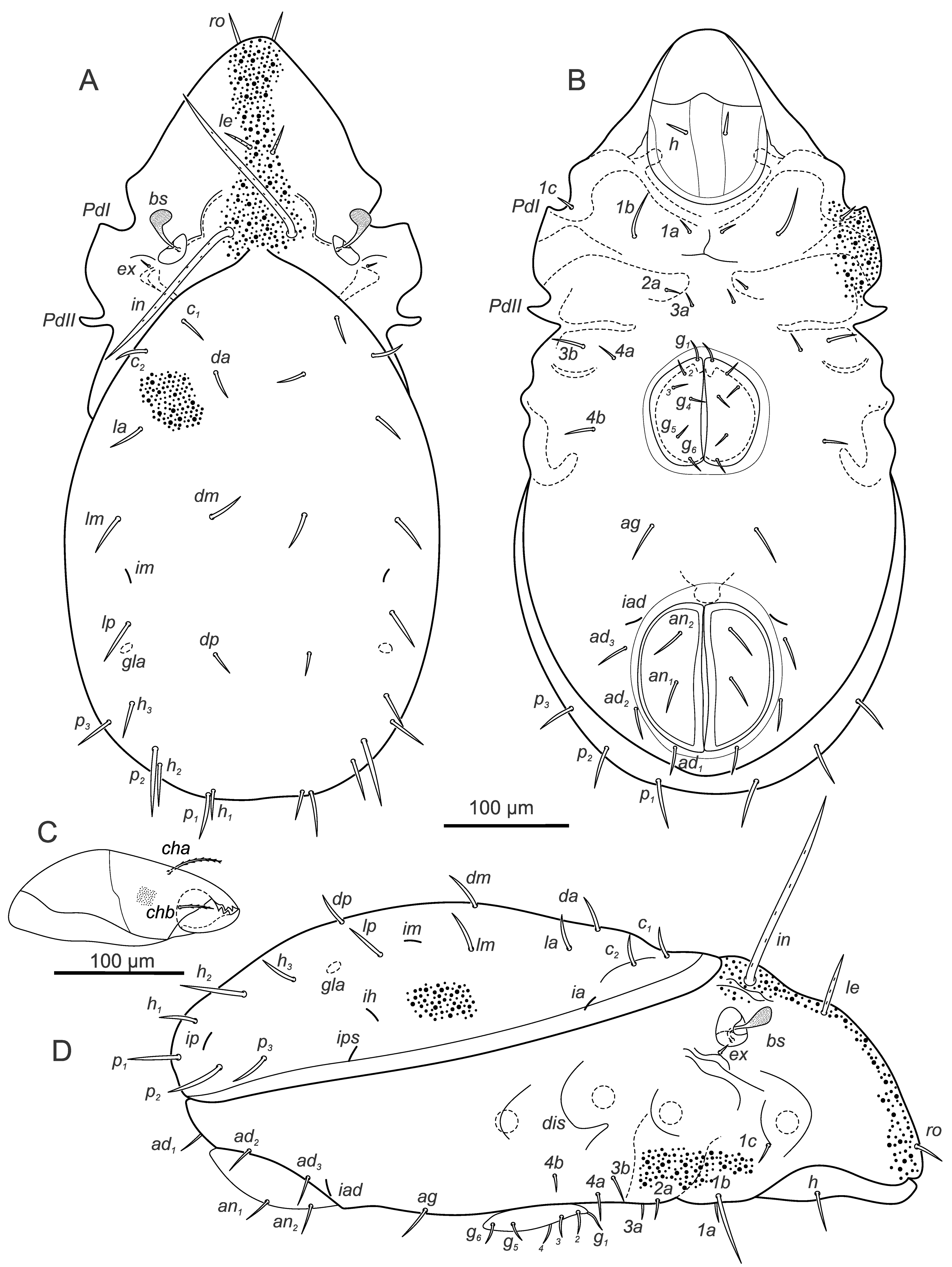
Larva – (Figures 10A, B) (n=5) length 179-207 (194), width 102-124 (115).
Gastronotic region: ex similar to in; 11 pairs of gastronotal setae (c1-3 , da, dm, dp, la, lm, lp, h1-2 ); h2 lateral to anal opening on ventral side, slightly longer than other setae.
Ventral region of idiosoma: epimeral setation 1-0-1-0 (1b, 3b); Claparède's organ present, no protective seta observed.
Protonymph – (Figures 10C, D) (n=4) length 219-246 (234), width 135-146 (140).
Gastronotic region: ex similar to in; 15 pairs of gastronotal setae (h3 , p1-3 added), p2-3 lateral to anal opening.
Ventral region of idiosoma: epimeral setation: 1-0-1-1 (4b added); one pair of short genital setae; two pairs of white spots on anal valves.
Deutonymph – (Figures 11A, B) (n=11) length 252-303 (281), width 153-194 (169).
Gastronotic region: ex similar to in; 15 pairs of gastronotal setae (as in protonymph), p2-3 ventrally, laterally to anal folds.
Ventral region of idiosoma: epimeral setation: 1-0-1-1; two pairs of short genital setae, two pairs of short setiform adanal setae lateral to anal folds, two vestigial pairs of anal setae on anal valves.
Tritonymph – (Figures 11C, D, supplementary figure 3) (n=8) length 320-366 (338), width 191-233 (207).
Gastronotic region: ex vestigial; 15 pairs of gastronotal setae (as in deutonymph), setae p2-3 postero-laterally to anal folds.
Ventral region of idiosoma: epimeral setation 1-0-1-1; three pairs of setiform genital, two pairs of adanal, two pairs of anal setae present.
Material examined
Four adults and 28 juveniles from Winterstrand, East London, Eastern Cape, South Africa, 33°5.67′S, 27°47.759′E, from algae in rock and tubeworms, eulittoral zone, 22 Oct. 2020, collected by J.A. Neethling (NMB 4707.1); six adults from Sheffield Beach, KwaZulu-Natal, South Africa, 29°29.737′S, 2°15.127′E, from Bostrychia sp. algae from rock, upper eulittoral zone, 27 Feb. 2020, collected by T. Pfingstl (NMB 4717.2).
Neotype designation
The holotype and paratype of S. ugraseni have been lost (Marshall, personal communication) and therefore a neotype should be selected. There is no material available from the type locality (Park Rynie). The specimens used here originate from Sheffield Beach (app. 116 km from Park Rynie) and Winterstrand (app. 612 km from Park Rynie). Due to the closer proximity, we selected a neotype from Sheffield Beach (NMB 4717.2.1) and five neo-paratypes from Sheffield Beach (NMB 4717.2.2) and four neo-paratypes from Winterstrand (NMB 4707.1.1) stored in the Acarology collection of NMB. Additional specimens will be stored in the Senckenberg Natural History Museum, Görlitz, Germany.
Remarks
The original description of Marshall and Pugh (2000) provides good morphological information and drawings of all aspects of the body, but not the legs. The adult specimens from Sheffield and Winterstrand match the description of the specimens from Park Rynie of the original description.
In general appearance the juveniles of S. ugraseni are very similar to those of the other two species of which juvenile stages are known: S. littorea Grandjean, 1968 (all stages) and S. melanomerus Marshall & Pugh, 2000 (protonymph) (Grandjean 1968; Pfingstl 2016). They have the same appearance and specific patterns of lateral and ventral folds. The protonymph of S. ugraseni differs from S. melanomerus in the presence of short ex (vestigial in S. melanomerus), two pairs of white spots on the anal valves (vs absent) and setae l′ on Fe I and II (vs absent).
A distinct difference between the adults of S. ugraseni and S. littorea is the number of anal and adanal setae with two pairs of anal and two pairs of adanal setae in the former, but one pair of anal and two pairs of adanal setae in the latter species. This trait is also different in the juveniles of these two species: the deuto- and tritonymph of S. ugraseni with two pairs of adanal setae and S. littorea with one pair, deutonymph and tritonymph of S. ugraseni with two pairs of anal (vestigial in deutonymph) while S. littorea has one pair in deuto- and tritonymph (also vestigial in deutonymph); the anal valves of the protonymph of S. ugraseni has two pairs of white spots, but they are absent in S. littorea. There is also a difference in the leg setae with seta l″ on Ti II present in the deuto- and tritonymphs, while it is only appearing in the adult of S. littorea. Another difference is the presence in S. ugraseni of a short ex seta in larva, proto- and deutonymphs and alveolus in tritonymph and adult, while a short ex seta is present in all stages of S. littorea and in the adult.
Family Podacaridae Grandjean, 1955
Genus: Halozetes Berlese, 1916
Halozetes capensis Coetzee & Marshall, 2003
Adult
Measurements: Adult body size: Kommetjie: holotype (female) length 637, width 351 (the scale bar in the drawing of the original description (Coetzee & Marshall 2003) is correct, but the sizes given in the text are incorrect and are rectified here), paratypes: female (n=1): length 662, width 374, males (n=4) length 565-595 (585), width 292-327 (314); Nature's Valley: females (n=2) length 590-617 (604), width 326-339 (333), males (n=3) length 571-586 (580), width 300-321 (311).
Integument (Figure 12): dark brown in colour; leg tarsi, tibiae lighter brown; dorsal side distinctly granulate with various sized granules, on prodorsum granules medially from anterior to posterior along hour-glass shape of ro, le and in, ventrally small area of granules between leg I and II, remainder of ventral side without granulation, legs with granules of various sizes; chelicera punctate.
Prodorsum (Figures 12A, D): rostrum rounded in dorsal view, slightly projected in lateral view; ro (20-31) smooth, le (35-69) roughened, in very long (150-181), roughened, directed upwards, ex short (8-13), smooth; bothridium cup-like with small opening for bs; bs with short stalk and clavate head (34-39).
Gnathosoma (Figure 12C): palp similar to that of H. belgicae mickii (Coetzee 2000), palp setation 0-2-1-3-9+1ω, palp solenidion ω and eupathidium acm connected, forming double horn; cheliceral setae setiform, barbed, chb slightly shorter than cha.
Notogaster (Figures 12A, D): elongated oval in dorsal view; anterior margin projecting anteriorly, dorsosejugal furrow medially indistinct; 14 pairs of smooth, stout notogastral setae (c1-2 , da, dm, dp, la, lm, lp, h1-3 , p1-3 ), c3 absent, ranges in the lengths of the dorsal setae on the notogaster quite large within populations, in the holotype the lengths towards the lower ranges (see Figure 2 in Coetzee and Marshall 2003, Figure 12 here depicts longer setae), mostly seta h2 longer than the others (c1 14-21, c2 18-25, da, dm, dp 12-28, la 10-27, lm 13-40, lp 16-47, h1 , h3 10-29, h2 27-59, p1 17-34, p2 -3 14-46); five pairs of distinct lyrifissures, ia postero-laterally to c2 , im between lm and lp, ih lateral to lp, ips lateral to ih close to latero-ventral border of notogaster, ip anterior to p1 ; gla posterior to lp.
Lateral aspect (Figure 12D): PdI small lamina, directed forward; discidium rounded.
Ventral (Figures 12B, D): all setae smooth; epimeral setation 3-1-2-2, seta 1b (40-56) < 3b (18-33) < 4a, 4b (13-24) < 1a, 2a, 1c, 3a (9-19); six pairs of genital (g1 15-19, g2-6 12-16), one pair of aggenital (22-39), two pairs of anal (17-29) and three pairs of adanal (ad1 14-18, ad2-3 17-25) setae present; anal plates large (length 114-141, width not measured due to distortion of plates in many specimens), genital plates larger in females than males (female: length 106-115, males 84-92; width not measured due to distortion of plates in many specimens), also genital plates of females with distinct posterior interlocking triangle, males with weaker triangle.
Legs: matching drawings in Coetzee and Marshall (2003); tridactylous, middle claw strong, long hook-like claws, weakly serrated dorsally; porose areas present; for setation and solenidia see Table 4.
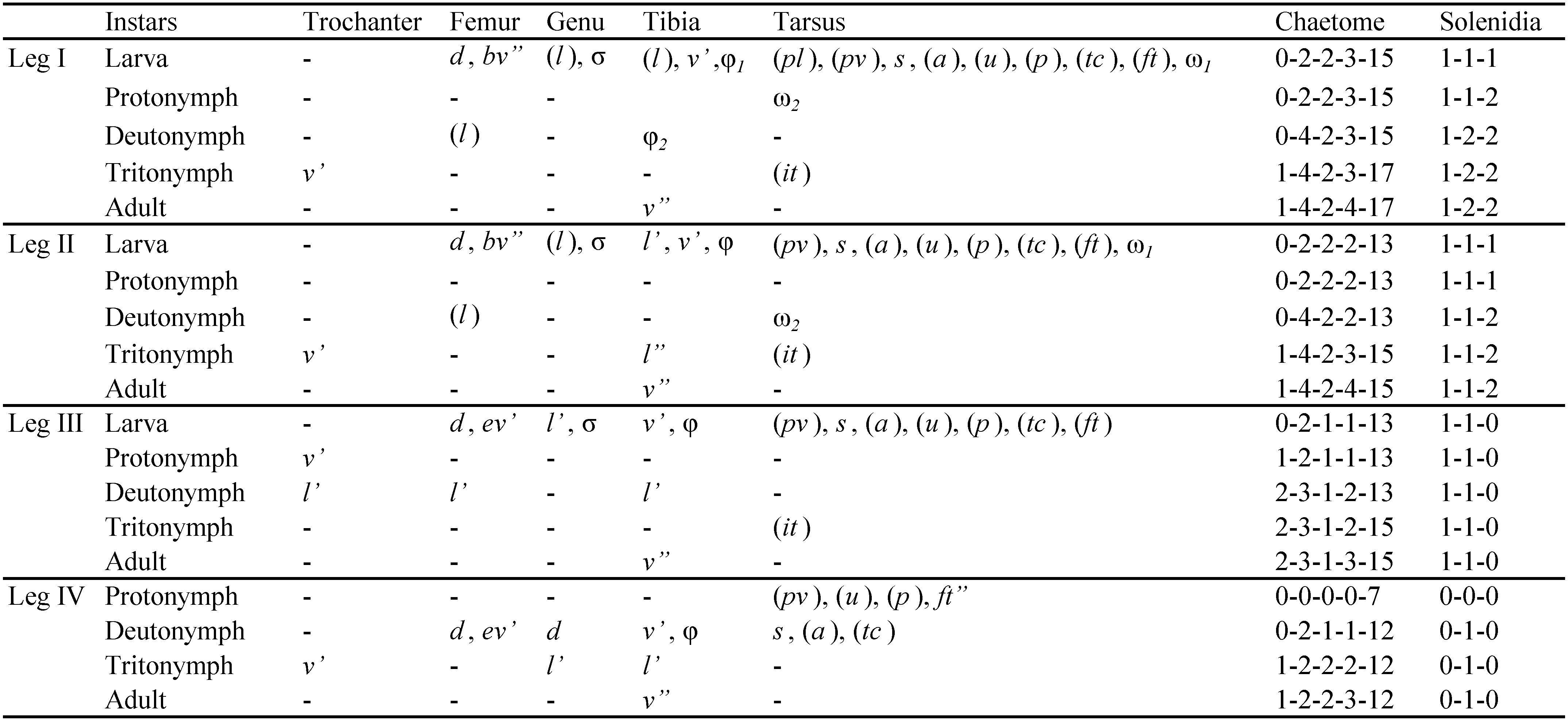
Juvenile stages
Common features – apheredermous; dark brown in colour, integument strongly plicate and soft, body distinctly granulate; three oval sclerites in posterior region of prodorsum, three pairs of sclerites on notogaster laterally between setal rows of l-series and d-series, anterior pair irregular in shape, others rounded or oval; prodorsum triangular, hysterosoma oval in dorsal view; rostrum rounded; ro short, le of medium length, in very long, all three setae roughened, ex very short, smooth; bothridia small; gastronotal setae thickened, setiform, smooth or slightly roughened; no cupules discernible; folds lateral to anal opening; epimeral region with folds; legs monodactylous; leg porose areas visible; for setation and solenidia see Table 4.
Larva – (Figures 13A, B) (n=8) length 268-298 (282), width 143-169 (154).
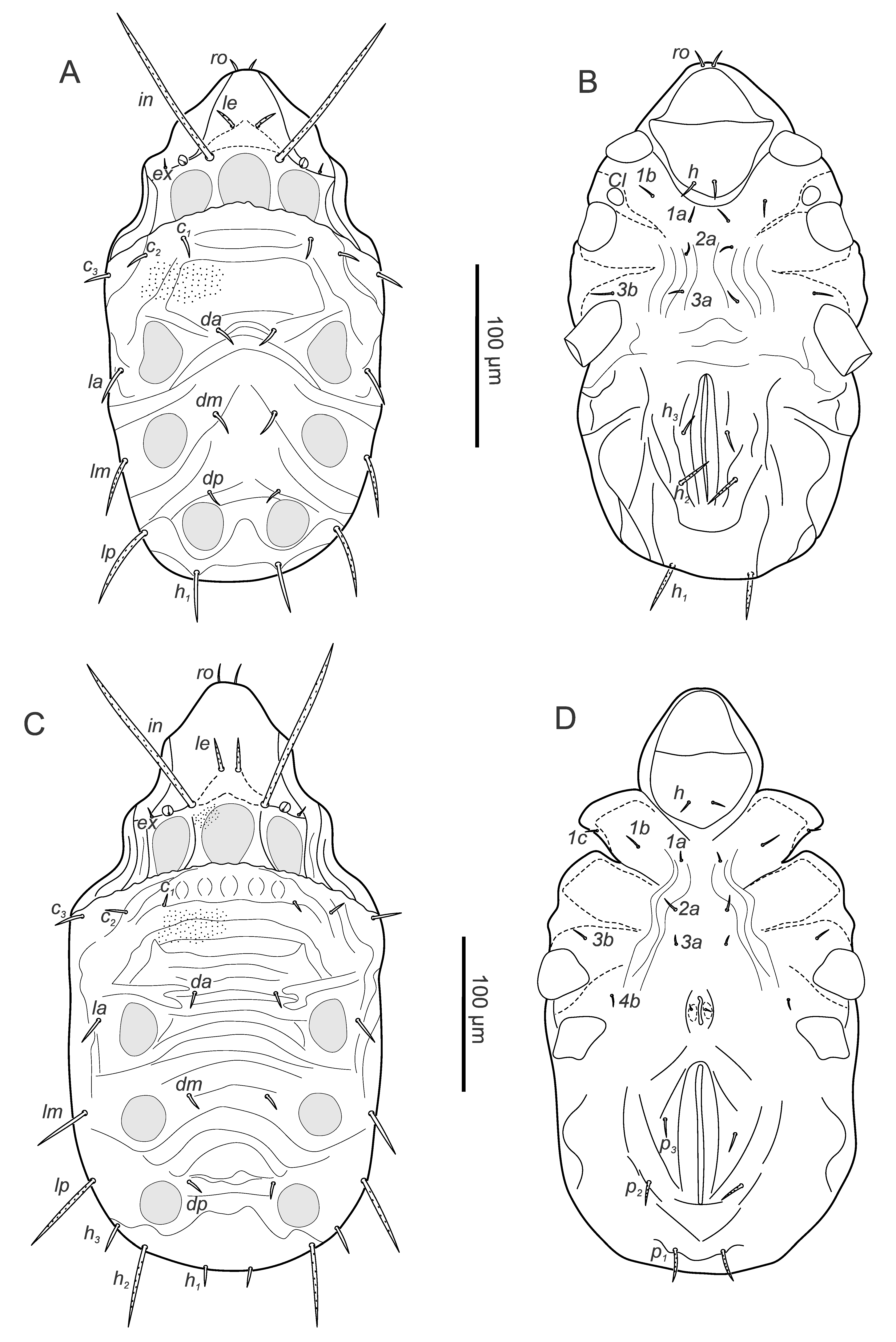
Gastronotic region: bs not visible externally, 12 pairs of gastronotal setae (c1-3 , da, dm, dp, la, lm, lp, h1-3 ), setae lp longest; h2 , h3 lateral to anal opening on ventral side.
Ventral region of idiosoma: epimeral setation 3-1-2-0 (1a, 1b, 1c, 2a, 3a, 3b) short setiform, smooth, third seta of first epimere forms protective scale over respective Claparède's organ.
Protonymph – (Figures 13C, D) (n=8) length 321-397 (361), width 185-221 (204).
Gastronotic region: bs not visible externally; 15 pairs of gastronotal setae (p1-3 added), seta lm slightly longer and lp and h2 much longer than others, p2-3 lateral and p1 posterior to anal folds.
Ventral region of idiosoma: epimeral setation: 3-1-2-1 (4b added), setae smooth, setiform; one pair of short genital setae.
Deutonymph – (Figures 14A, B) (n=9) length 424-479 (439), width 230-254 (241).

Gastronotic region: bs not visible externally; 15 pairs of gastronotal setae, setae lp and h2 longest, p2-3 , laterally to anal folds; some dorsal setae of specimens from Nature's Valley distinctly longer than specimens from Kommetjie (Nature's Valley c1-2 15-21, c3 24-27, da, dm 23-27, dp 11-16, la, 24-28, lm 31-34, h1-3 12-18, Kommetjie c1-2 8-10, c3 21-22, da, dm 8-14, dp 8-9, la, 14-16, lm 15-20, h1-3 8-13), other setae slightly shorter (Nature's Valley lp 40-47, h2 59-66, Kommetjie lp 47-59, h2 72-78).
Ventral region of idiosoma: epimeral setation: 3-1-2-2 (4a added); three pairs of short genital setae, three pairs of short setiform adanal setae lateral to anal opening, aggenital setae added.
Tritonymph – (Figures 14C, D) (n=11) length 492-636 (561) width 284-324 (307).
Gastronotic region: bs with short stalk and very small, rounded head; 15 pairs of gastronotal setae, setae lp and h2 longest, p1-3 ventrally, p1-2 posterior to anal folds; some dorsal setae of specimens from Nature's valley slightly longer or shorter than specimens from Kommetjie but not as distinct as in deutonymph (see difference in setal lengths in Figure 14 here compared to Figure 10 in Coetzee and Marshall (2003)).
Ventral region of idiosoma: epimeral setation as in deutonymph; five pairs of setiform genital, one pair of aggenital, three pairs of adanal, two pairs of anal setae.
Material examined
Five adults, one protonymph, nine deutonymphs, six tritonymphs from Nature's Valley, Western Cape, South Africa, 33°59.333′S, 23°32.796′E, from Bostrychia intricata algae on rocks, upper eulittoral zone, 23 Feb. 2019, collected by T. Pfingstl (NMB 4671.1); holotype (NMB 4150.1), five adult paratypes (the paratypes of the original description could not be identified, as they were within a vial of 59 individuals), eight larvae, seven protonymphs, four deutonymphs, five tritonymphs from Kommetjie, Western Cape, South Africa, 34°13′S, 18°32′E, from finely branched algae on rocks, middle and upper eulittoral zone, Oct. 2000, collected by D.J. Marshall (NMB 4150).
Remarks
The original description by Coetzee and Marshall (2003) of H. capensis provides very good morphological information and drawings of all aspects including legs. The adults from Nature's Valley of our samples do not differ from those of Kommetjie. The variation in the length of dorsal setae, even within a population, is noteworthy.
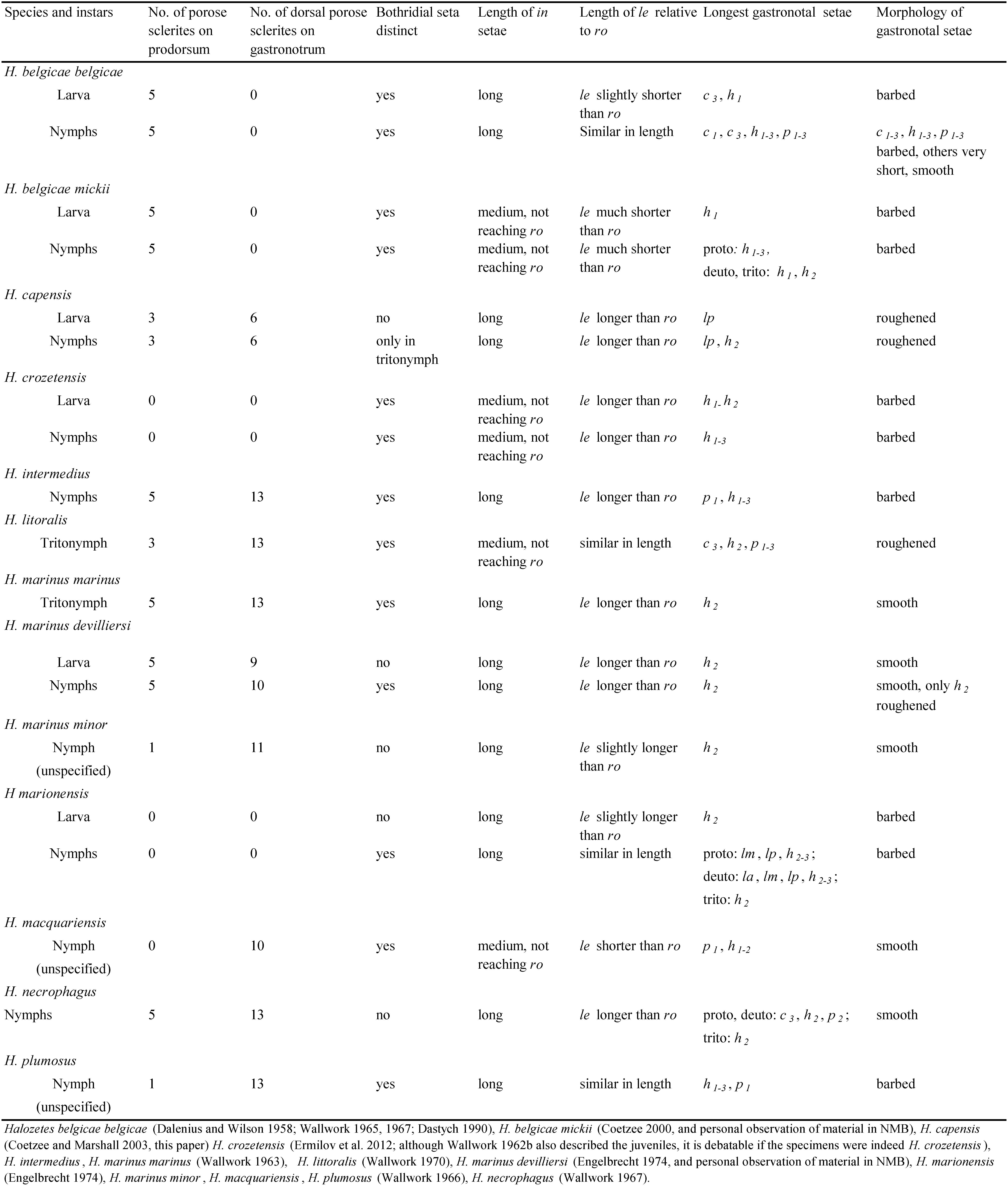
Some or all juvenile stages of several Halozetes species and subspecies are known, namely H. belgicae belgicae (Michael, 1903), H. belgicae mickii Coetzee, 2000, H. capensis, H. crozetensis (Richters, 1907), H. intermedius Wallwork, 1963, H. litoralis Wallwork, 1963, H. marinus marinus (Lohmann, 1907), H. marinus devilliersi Engelbrecht, 1974, H. marinus minor Wallwork, 1966, H. marionensis Engelbrecht, 1974, H. macquariensis (Delenius, 1958), H. necrophagus Wallwork, 1967 and H. plumosus Wallwork, 1966 (Norton & Ermilov 2014, also Appendix 1). Ermilov et al. (2012) provided very useful information and a key to the juveniles of the various species. The juveniles of H. capensis differ from other known juveniles mostly in the number of porose sclerites on the prodorsum and gastronotum, the distinctness of bs, length of setae in and le and in the morphology of gastronotal setae (see Table 5 for a summary of the differences).
Acknowledgments
Many thanks to the South African authorities for issuing permits: CapeNature (permit no. CN44-31-7224), Eastern Cape Parks & Tourism Agency (permit no. RA_0295), South African National Parks (SANPARKS) (permit no. CRC/2019-2020-002-2019/V1, reference no. COEH/AGR/002-2019/2019-2022/V1), iSimangaliso Wetland Park and Department Environment, Forestry and Fisheries (DEFF) (permit no. RES2020/93).
This study was funded by the OeAD – Austrian Agency for International Cooperation in Education and Research (project no. ZA 13/2019) together with the National Research Foundation; South-Africa/Austria Joint Scientific and Technological Cooperation (grant no. 116060).
Appendix
Species of the superfamily Ameronothroidea for which information on juveniles exist in the literature

acarologia_4538_supplementary.pdf
References
- Bayartogtokh B., Chatterjee T. 2010. Oribatid mites from marine littoral and freshwater habitats in India with remarks on world species of Thalassozetes (Acari: Oribatida). Zool. Stud., 49(6): 839-854.
- Bayartogtokh B., Chatterjee T., Chan B.K.K., Ingole B. 2009. New species of marine littoral mites (Acari: Oribatida) from Taiwan and India, with a key to the world's species of Fortuynia and notes on their distributions. Zool. Stud., 48: 243-261.
- Behan-Pelletier V.M. 1997. The semiaquatic genus Tegeocranellus (Acari: Oribatida: Ameronothroidea) of north and central America. Can. Entomol., 129: 537-577. https://doi.org/10.4039/Ent129537-3
- Bücking J., Ernst H., Siemer F. 1998. Population dynamics of phytophagous mites inhabiting rocky shores - K-strategists in an extreme environment? In: Ebermann E. (Ed). Arthropod Biology: Contributions to morphology, ecology and systematics. Biosystematics and Ecology Series 14. Österreichische Akademie der Wissenschaften, Vienna. p. 93-143.
- Coetzee L. 2000. A new subspecies of Halozetes belgicae (Acari, Oribatida, Ameronothridae) from the marine supra-littoral of the sub-Antarctic Marion Island. Acarologia, 40: 439-447.
- Coetzee L., Marshall D.J. 2003. A new Halozetes species (Acari : Oribatida: Ameronothridae) from the marine littoral of southern Africa. Afr. Zool., 38(2): 327-331.
- Dalenius P., Wilson O. 1958. On the soil fauna of the Antarctic and of the sub-Antarctic Islands. The Oribatidae (Acari). Ark. Zool., 11(23): 393-425.
- Dastych H. 1990. Some notes on Antarctic mites (Acari). Entomol. Mitt. Zool. Mus. Hamburg, 10 (139/140): 43-56.
- Engelbrecht C.M. 1974. The genus Halozetes (Oribatei, Acari) on Marion Island. Navors. Nas. Mus. 3(1): 1-25.
- Engelbrecht C.M. 1975. New ameronothroid (Oribatei: Acari) taxa from Republic of South Africa and the islands Gough and Marion. Navors. Nas. Mus., 3(4): 53-88.
- Ermilov S.G., Stary J., Block W. 2012. Morphology of juvenile instars of Ameronothridae (Acari: Oribatida). Zootaxa, 3224: 1-40. https://doi.org/10.11646/zootaxa.3224.1.1
- Grandjean F. 1949. Formules anales gastronotiques, genitales et aggenitales du developpement numerique des poils chez les Oribates. Bull. Soc. Zool. Fr., 74: 201-225.
- Grandjean F. 1955. Sur un acarien des iles Kerguelen. Podacarus auberti (Oribates). Mem. Mus. Nat. Hist. Natur. Paris, 8 : 109-150.
- Grandjean F. 1966. Selenoribates mediterraneus n. sp. et les Selenoribatidae (Oribates). Acarologia, 8: 129-154.
- Grandjean F. 1968. Schusteria littorea n. g., n. sp. et les Selenoribatidae (Oribates). Acarologia, 10: 116-150.
- Hammen van der L. 1963. Description of Fortuynia yunkeri nov. spec., and notes on the Fortuyniidae nov. fam. Acarologia, 5: 152-167.
- Krisper G., Schuster R. 2008. Fortuynia atlantica sp. nov., a thalassobiontic oribatid mite from the rocky coast of the Bermuda Islands (Acari: Oribatida: Fortuyniidae). Ann. Zool., 56(2): 419-432. https://doi.org/10.3161/000345408X326753
- Luxton M. 1966. Laboratory studies on the feeding habits of saltmarsh Acarina, with notes on their behavior. Acarologia, 8: 163-174.
- Luxton M. 1967. Two new mites from the rocky shore of North Auckland. New Zeal. J. Mar. Fresh. Res., 1: 76-87. https://doi.org/10.1080/00288330.1967.9515194
- Luxton M. 1992. Oribatid mites from the marine littoral of Hong Kong (Acari: Cryptostigmata). In: Morton B (Ed). The marine flora and fauna of Hong Kong and southern China III. Proceedings of the Fourth International marine Biological Workshop: the marine Flora and Fauna of Hong Kong and Southern China. Hong Kong: Hong Kong University Press. p. 211-227.
- Marshall D.J., Pugh P.J.A. 2000. Two new species of Schusteria (Acari: Oribatida: Ameronothroidea) from marine shores in southern Africa. Afr. Zool., 35(2): 201-205. https://doi.org/10.1080/15627020.2000.11657091
- Marshall D.J., Pugh P.J.A. 2002. Fortuynia (Acari: Oribatida: Ameronothroidea) from the marine littoral of southern Africa. J. Nat. Hist, 36: 173-183. https://doi.org/10.1080/00222930010002775
- Michael A.D. 1882. Further notes on British Oribatidae. J. R. Microsc. Soc. (series 2), 2: 1-18. https://doi.org/10.1111/j.1365-2818.1882.tb00175.x
- Michael A.D. 1888. British Oribatidae. Vol. II. Ray Society, London, 337-657.
- Michael A.D. 1903. Acarina (Oribatidae). Expedition Antarctique Belge. Resultats du Voyage du S.Y. Belgica en 1897-1898-1899. Rep. Sci. Zool. Acariens Libres, 10: 1-6.
- Norton R.A. 1977. A review of F. Grandjean's system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal D.L. (Ed). Biology of oribatid mites. Syracuse: SUNY College of Environmental Science and Forestry. p. 33-61.
- Norton R.A., Behan-Pelletier V.M. 2009. Suborder Oribatida. Chapter 15. In: Krantz G., Walter D.E. (Eds.). A Manual of Acarology. Texas Tech University Press, Lubbock, p. 430-564.
- Norton R.A., Ermilov S.G. 2014. Catalogue and historical overview of juvenile instars of oribatid mites (Acari: Oribatida). Zootaxa, 3833 (1): 1-132. https://doi.org/10.11646/zootaxa.3833.1.1
- Norton R.A., Franklin E. 2018. Paraquanothrus n. gen. from freshwater rock pools in the USA, with new diagnoses of Aquanothrus, Aquanothridae, and Ameronothridae (Acari, Oribatida). Acarologia, 58(3): 557-627. https://doi.org/10.24349/acarologia/20184258
- Pfingstl T. 2013a. Habitat use, feeding and reproductive traits of rocky-shore intertidal mites from Bermuda (Oribatida: Fortuyniidae and Selenoribatidae). Acarologia, 53(4): 369-382. https://doi.org/10.1051/acarologia/20132101
- Pfingstl T. 2013b. Revealing the diversity of a once small taxon: the genus Selenoribates (Acari, Oribatida, Selenoribatidae). Zookeys, 312: 39-63. https://doi.org/10.3897/zookeys.312.5478
- Pfingstl T. 2015a. An interesting case of sexual dimorphism in intertidal mites: Fortuynia dimorpha sp. nov. (Acari, Oribatida, Fortuyniidae). Syst. Appl. Acarol., 20: 567-578. https://doi.org/10.11158/saa.20.5.10
- Pfingstl, T. 2015b. The intertidal Fortuyniidae (Acari: Oribatida): new species, morphological diversity, ecology and biogeography. Zootaxa, 3957: 351¬-382. https://doi.org/10.11646/zootaxa.3957.4.1
- Pfingstl T. 2015c Morphological diversity in Selenoribates (Acari, Oribatida): new species from coasts of the Red Sea and the Indo-Pacific. Int. J. Acarol., 41(4): 356-370. https://doi.org/10.1080/01647954.2015.1035321
- Pfingstl T. 2016. Marine associated mites (Acari: Oribatida: Ameronothroidea) of the African continent: biogeography, new species and distribution records. Syst. Appl. Acarol., 21: 320-334. https://doi.org/10.11158/saa.21.3.7
- Pfingstl T. 2017. The marine-associated lifestyle of ameronothroid mites (Acari, Oribatida) and its evolutionary origin: a review. Acarologia, 57(3): 693-721. https://doi.org/10.24349/acarologia/20174197
- Pfingstl T., Krisper G. 2014. Plastron respiration in marine intertidal oribatid mites (Acari, Fortuyniidae and Selenoribatidae). Zoomorphology, 133: 359-378. https://doi.org/10.1007/s00435-014-0228-5
- Pfingstl T., Schatz H. 2017. New littoral mite species (Acari, Oribatida, Fortuyniidae) from the Galápagos archipelago, with ecological and zoogeographical considerations. Zootaxa, 4244: 39-64. https://doi.org/10.11646/zootaxa.4244.1.2
- Pfingstl T., Schuster R. 2012a. First record of the littoral genus Alismobates (Acari: Oribatida) from the Atlantic Ocean, with a redefinition of the family Fortuyniidae based on adult and juvenile morphology. Zootaxa, 3301: 1-33. https://doi.org/10.11646/zootaxa.3301.1.1
- Pfingstl T., Schuster R. 2012b. Carinozetes nov. gen. (Acari: Oribatida) from Bermuda and remarks on the present status of the family Selenoribatidae. Acarologia, 52(4): 377-409. https://doi.org/10.1051/acarologia/20122067
- Pfingstl T., Schuster, R. 2014. Global distribution of the thalassobiontic Fortuyniidae and Selenoribatidae (Acari, Oribatida). Soil Org., 86(2): 125-130.
- Pfingstl T., Baumann J., Lienhard A., Schatz H. 2017. New Fortuyniidae and Selenoribatidae (Acari, Oribatida) from Bonaire (Lesser Antilles) and morphometric comparison between Eastern Pacific and Caribbean populations of Fortuyniidae. Syst. Appl. Acarol., 22(12): 2190-2217. https://doi.org/10.11158/saa.22.12.11
- Pfingstl T., Baumann J., Neethling J.A., Bardel-Kahr I., Hugo-Coetzee E.A. 2021a. Distribution patterns of intertidal oribatid mites (Acari, Oribatida) from South African shores and their relationship to temperature. Afr. J. Mar. Sci., 43(2): 215-225. https://doi.org/10.2989/1814232X.2021.1912825
- Pfingstl T., De La Paz J.C., Hernández-Teixidor D. 2020. First record of intertidal oribatid mites (Acari, Oribatida) from the Canaries - a new species and its complete ontogeny. Syst. Appl. Acarol., 25(10): 1901-1914. https://doi.org/10.11158/saa.25.10.13
- Pfingstl T., Hiruta S.F., Hagino W., Shimano S. 2021b. Juvenile morphology of seven intertidal mite species (Acari, Oribatida, Ameronothroidea) from the East Asian Region. Int. J. Acarol., 47(6): 536-554. https://doi.org/10.1080/01647954.2021.1965656
- Pfingstl T., Hiruta S.F., Wagner M., Hagino W., Shimano S. 2019b. First record of the family Ameronothridae (Acari: Oribatida) from Japan - new species, juvenile morphology, ecology and biogeographic remarks. Int. J. Acarol., 45(5): 315-327. https://doi.org/10.1080/01647954.2019.1629624
- Pfingstl T., Lienhard A., Jagersbacher-Baumann J. 2014. Hidden in the mangrove forest: the cryptic intertidal mite Carinozetes mangrovi sp. nov. (Acari, Oribatida, Selenoribatidae). Exp. Appl. Acarol., 63(4): 481-495. https://doi.org/10.1007/s10493-014-9802-2
- Procheş Ş., Marshall D.J. 2001. Global distribution patterns of non-halacarid marine intertidal mites: implications for their origins in marine habitats. J. Biogeogr., 28: 47-58. https://doi.org/10.1046/j.1365-2699.2001.00513.x
- Procheş Ş., Marshall D.J. 2002. Diversity and biogeography of southern African intertidal Acari. J. Biogeogr., 29: 1201-1215. https://doi.org/10.1046/j.1365-2699.2002.00772.x
- Pugh P.J.A., King P.E. 1985. Feeding in intertidal Acari. J. Exp. Mar. Biol. Ecol., 94: 269-280. https://doi.org/10.1016/0022-0981(85)90064-4
- Resch B., Baumann J., Pfingstl T. 2019. The intertidal genus Indopacifica (Acari, Oribatida, Selenoribatidae): new species, juvenile morphology and distribution. Zootaxa 4647: 322-335. https://doi.org/10.11646/zootaxa.4647.1.19
- Schubart H. 1970. Ameronothrus schusteri n. sp., eine neue Oribatide von der Küste Jugoslawiens (Arachnida: Acari). Senckenb. Biol., 51(5-6): 425-432.
- Schubart H. 1975. Morphologische Grundlagen für die Klärung der Verwandtschaftsbeziehungen innerhalb der Milbenfamilie Ameronothridae (Acari, Oribatei). Zoologica, 123: 24-91.
- Schuster R. 1963. Thalassozetes riparius n. gen., n. sp., eine litoralbewohnende Oribatide von bemerkenswerter morphologischer Variabilität (Acari-Oribatei). Zool. Anz., 171: 391-403.
- Sitnikova L.G. 1968. On the question of evolution in the family Podacaridae Grandj. Second All-Union Symposium on Oribatid mites. Institute of Zoology and Parasitology of the Academy of Sciences of the Lithuanian SSR, Vilnyus. p. 9-10.
- Sitnikova L.G. 1975. The superfamily Ameronothroidea Balogh, 1961. In: Ghilyarov M.S. (Ed). Key to Soil Inhabiting Mites. Sarcoptiformes. Moscow: Nauka Press. p. 232-234.
- Wallwork J.A. 1962a. A redescription of Notaspis antarcticus Michael, 1903 (Acari: Oribatei). Pac. Insects, 4(4): 869-880.
- Wallwork J.A. 1962b. Notes on the genus Pertorgunia Dalenius, 1958 from Antarctica and Macquarie (Acari: Oribatei). Pac. Insects, 4(4): 881-885.
- Wallwork J.A. 1963. The oribatei (Acari) of Macquarie Island. Pac. Insects, 5(4): 721-769.
- Wallwork J.A. 1965. The cryptostigmata (Acari) of Antarctica with species reference to the Antarctic Peninsula and South Shetland Islands. Pac. Insects, 7(3): 453-468.
- Wallwork J.A. 1966. More oribatid mites (Acari: Cryptostigmata) from Campbell I. Pac. Insects, 8(4): 849-877.
- Wallwork J.A. 1967. Cryptostigmata (Oribatid mites). In: Gressit J.L. (Ed). Entomology of Antarctica. Antarctic Research series, 10. Washington, D.C. American Geophysical Union. p. 105-122. https://doi.org/10.1029/AR010p0105
- Wallwork J.A. 1970. Acarina: Cryptostigmata of South Georgia. Pac. Insects Monogr., 23: 161-178.
- Wallwork J.A. 1981. A new aquatic oribatid mite from Western Australia (Acari: Cryptostigmata: Ameronothridae). Acarologia, 22(3): 333-339.
- Weigmann G. 1975. Vorkommen von Ameronothrus (Acari, Oribatei) im Litoral Südafrikas. Veröff. Inst. Meeresforsch. Bremerhav., 15: 65-67.
- Zhang Z.-Q. 2018. Accelerating studies on the ontogeny and morphological diversity in immature mites. Zootaxa, 4540(1): 5-6. https://doi.org/10.11646/zootaxa.4540.1.3



2021-12-06
Date accepted:
2022-04-29
Date published:
2022-08-10
Edited by:
Roy, Lise

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Hugo-Coetzee, Elizabeth A.; Baumann, Julia; Neethling, Jan-Andries; Bardel-Kahr, Iris and Pfingstl, Tobias
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)