



Infestation of laboratory colonies of the soft tick, Ornithodoros moubata Murray 1877 (Acari: Ixodida: Argasidae) by Tyrophagus fanetzhangorum Klimov & OConnor 2009 (Acari: Astigmatina: Acaroidea: Acaridae)
Filatov, Serhii  1
; Erhart, Jan2
and Rego, Ryan O.M.3
1
; Erhart, Jan2
and Rego, Ryan O.M.3
1✉ Department of Pediatrics and the National School of Tropical Medicine, Baylor College of Medicine, Houston, Texas, USA
2Institute of Parasitology, Biology Centre of the Czech Academy of Sciences, České Budějovice, Czechia.
3Institute of Parasitology, Biology Centre of the Czech Academy of Sciences, České Budějovice, Czechia & Faculty of Science, University of South Bohemia, České Budějovice, Czechia.
2022 - Volume: 62 Issue: 1 pages: 223-227
https://doi.org/10.24349/xuqf-20hwShort note
Keywords
Abstract
Laboratory colonies of medically and veterinary important hematophagous arthropods are crucial resources for bettering our understanding of vector-borne diseases and for the discovery of new and efficient ways to keep at bay these threats to animal and public health. Many sub-disciplines within the research field equally depend on the availability of live vectors to conduct lab experiments and how the organism in question was reared and maintained before the study commenced is critical for the reproducibility of results and their interpretation (Wilson and Harrup, 2018). However, most published papers leave out natural history details behind the colony work, at best limiting the information to a few succinct words about the geographical origin and laboratory maintenance conditions. Until recently, little attention has been devoted to the diverse micro- and macro-symbionts and other co-inhabitants occurring in established research colonies of arthropod vectors.
With the increasing recognition of the importance of symbiotic microbiota in vector-pathogen interactions (Rego et al., 2020), research focus is slowly shifting towards the vector as a holobiont, but observations on the macroscopic symbionts of hematophagous arthropods (either in culture or in the field conditions) remain exceedingly rare. Here, we describe a serendipitous finding of an astigmatid mite infesting a long-term laboratory colony (over 20 years old) of the soft tick Ornithodoros moubata (Murray, 1877) and discuss the possible nature of this association.
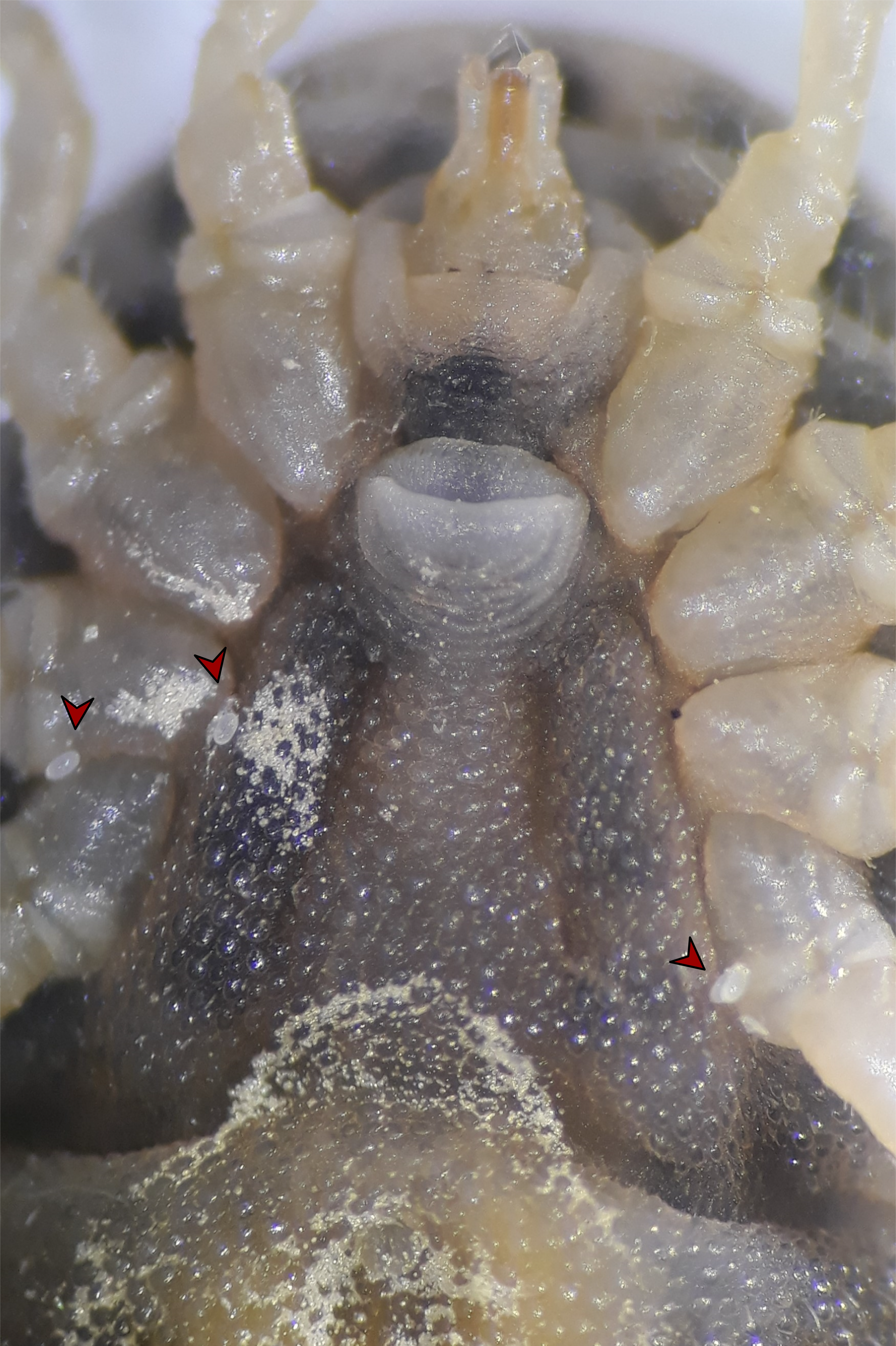
While sexing individual ticks in the laboratory colony at the Institute of Parasitology, our attention was attracted by a few rapidly moving, small, yellowish mites emerging from the integumental folds around the coxae of a late-stage O. moubata nymph. The mites were moving erratically across the ventral surface of the tick, disappearing behind the coxae and coming into sight again, probably disturbed by the bright light of the stereomicroscope. Upon closer inspection, we were able to find more mites on 4 other ticks out of the 50 individuals contained in the same box. No mites were found crawling on the dorsal surface of the ticks, on the filter paper within, or at the bottom and on the walls of the plastic box with the soft ticks. Further checks of other, randomly picked boxes from the colony revealed the same pattern, with a few mites occurring on live, normal looking O. moubata and, sometimes, on dead ticks as well. In one case, a dead tick had multiple mites of different developmental stages on its ventral side (Suppl. video https://figshare.com/s/79bf7d0e975713252c21 ![]() ) and in some instances mite eggs, along with active stages, could be observed attached to the ventral cuticle of a living specimen (Fig. 1).
) and in some instances mite eggs, along with active stages, could be observed attached to the ventral cuticle of a living specimen (Fig. 1).
To identify the mite species, we collected 20-30 specimens with fine-tipped forceps and placed them in 96% ethanol. Total genomic DNA from the specimens was extracted using the DNAeasy Tissue Kit (Qiagen), as previously described (Dabert et al., 2008). Briefly, mites were transferred into 180 µl of ATL buffer, incubated at 56°C overnight, then incubated with added Proteinase K at the same conditions for 72 hours. DNA was then isolated according to the manufacturer's instructions. The central part of the Cytochrome oxidase I (COI) was amplified using the primers described in Yang et al. (2011) and sequenced in both directions. The raw chromatograms were manually checked, quality trimmed and assembled into a consensus sequence using the Unipro UGENE software (Okonechnikov et al., 2012). The obtained consensus sequence was identified using the GenBank Nucleotide database with the BLAST tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi ![]() ). The obtained COI sequence was deposited in the GenBank database under the accession number OK036578.
). The obtained COI sequence was deposited in the GenBank database under the accession number OK036578.
The molecular identification revealed that the mites infesting O. moubata belong to the species Tyrophagus fanetzhangorum Klimov & OConnor, 2009 (99.7% nucleotide identity, 100% coverage to KY9862266), which only recently has been formally described and separated from the closely related Tyrophagus putrescentiae (Schrank, 1781) using both morphological and molecular evidence (Klimov and OConnor, 2009). Very little information about the former species can be found in the literature. Therefore, we provide a brief review of ecological information on T. putrescentiae for the following discussion, anticipating the two species may share many ecological features.
T. putrescentiae is a ubiquitous mite, well known as a pest of stored products and a source of medically important house dust mite allergens (Klimov and OConnor, 2009). It also has been reported infesting various research colonies of insects, such as the Africanized honey bees (Texeira et al., 2014), the invasive Aedes spp. mosquitoes (Serpa et al., 2004), and Drosophila spp. (Brown and Lloyd, 2015), although the effect of these infestations on colony fitness is largely unknown. We did not observe any effects of T. fanetzhangorum either on infested individuals of O. moubata or on their colonies. Concerning the possible source of the infestation, we suspect that the mite could have gotten into tick colonies through contact with laboratory animals used to feed the ticks.
Tyrophagus spp. are generally considered to be fungivores although predation on soil micro-invertebrates, such as nematodes, has been increasingly recognized as a dietary habit occurring in this group of acarines (Krantz, 2009; Walter et al., 1986). Because of the high relative humidity required to maintain laboratory tick colonies, they are often contaminated by a variety of environmental saprophytic fungi, which can sometimes be responsible for an increased mortality in ticks (Bonnet et al., 2021). However, as wild ticks spend most of their lifetime in the soil (Burtis et al., 2019), they must have evolved a tolerance to frequently encountered and potentially dangerous microbes, albeit their pathogenicity for ticks might be species- and/or context-dependent (Loosová et al., 2001; Pekas et al., 2017). In our experience, Ornithodoros ticks in laboratory colonies rarely experience massive die-offs that could be attributed to pathogenic fungi, but dead ticks represent a favorable substrate for colonization by various molds (Figure 2). These, in turn, could represent the required resource for the build-up of fungivorous mite populations. The constant presence of the mites on live and apparently healthy ticks but not on the rearing boxes however, merits further observations on the basic biology of these acarines.

Acaroid mites (family Acaridae) are well-known for their phoretic associations with various invertebrates and it has been hypothesized that the family's ancestors were saprophagous inhabitants of animal nests that adapted to utilize nidicolous arthropods for dispersal between resource patches (Murillo Rojas, 2019). Although the genus Tyrophagus subsequently lost the specialized phoretic deutonymph stage, which is present in most other genera (Murillo Rojas, 2019), our observations of the close association of T. fanetzhangorum with O. moubata suggest a perfect ecological match and potential for mutualistic or at least commensalistic interactions between the species. It is unknown if such interactions can occur in the field, but the occurrence of T. putrescentiae on other nidicolous ectoparasites of animals such as fleas (Karbowiak et al., 2013) hints toward this possibility.
To sum up, we report a curious case of a laboratory interaction between a tick species and an acaroid mite that conceivably may also occur in naturally infested animal burrows, habitats that often represent incredibly species-rich ecosystems (Krivokhatsky, 1994). More work will now be required to understand the importance of this interaction for each species and its occurrence in natural populations.
Acknowledgments
The short research stay of the first author at the Institute of Parasitology, Biology Centre of the Czech Academy of Sciences was funded through a Visegrad Scholarship project (International Visegrad Fund). We thank the two anonymous reviewers and Dr Karen McCoy for providing helpful comments and suggestions on earlier drafts of the manuscript.
References
- Bonnet S.I., Blisnick T., Al Khoury C., Guillot J. 2021. Of fungi and ticks: Morphological and molecular characterization of fungal contaminants of a laboratory-reared Ixodes ricinus colony. Ticks and Tick-borne Dis., 12: 101732. https://doi.org/10.1016/j.ttbdis.2021.101732
- Brown A.N., Lloyd V.K. 2015. Evidence for horizontal transfer of Wolbachia by a Drosophila mite. Exp. Appl. Acarol., 66: 301-311. https://doi.org/10.1007/s10493-015-9918-z
- Burtis J.C., Yavitt J.B., Fahey T.J., Ostfeld R.S. 2019. Ticks as soil-dwelling arthropods: an intersection between disease and soil ecology. J. Med. Entomol., 56: 1555-1564. https://doi.org/10.1093/jme/tjz116
- Dabert J., Ehrnsberger R., Dabert M. 2008. Glaucalges tytonis sp. n.(Analgoidea, Xolalgidae) from the barn owl Tyto alba (Strigiformes, Tytonidae): compiling morphology with DNA barcode data for taxon descriptions in mites (Acari). Zootaxa, 1719: 41-52. https://doi.org/10.11646/zootaxa.1719.1.2
- Karbowiak G., Solarz K., Asman M., Wróblewski Z., Slivinska K., Werszko J. 2013. Phoresy of astigmatic mites on ticks and fleas in Poland. Biol. Lett., 50: 87-94. https://doi.org/10.2478/biolet-2013-0007
- Klimov P.B., OConnor B.M. 2009. Conservation of the name Tyrophagus putrescentiae, a medically and economically important mite species (Acari: Acaridae). Int. J. Acarol., 35: 95-114. https://doi.org/10.1080/01647950902902587
- Krantz G. 2009. Habits and habitats. A manual of Acarology. 3rd edition. Texas Tech Univ. Press, Lubbock, Texas, USA: 64-82.
- Krivokhatsky V.A. 1994. Arthropods inhabiting rodent burrows in the Karakum desert. Biogeography and Ecology of Turkmenistan. Springer. p. 389-402. https://doi.org/10.1007/978-94-011-1116-4_23
- Loosová G., Jindrák L., Kopacek P. 2001. Mortality caused by experimental infection with the yeast Candida haemulonii in the adults of Ornithodoros moubata (Acarina: Argasidae). Folia Parasitol., 48: 149-153. https://doi.org/10.14411/fp.2001.023
- Murillo Rojas P. 2019. Disentangling Species Boundaries and the Evolution of Habitat Specialization for the Ecologically Diverse Mite Family Acaridae. The University of Michigan. pp. 102.
- Okonechnikov K., Golosova O., Fursov M., the U.t. 2012. Unipro UGENE: a unified bioinformatics toolkit. Bioinform., 28: 1166-1167. https://doi.org/10.1093/bioinformatics/bts091
- Pekas A., Palevsky E., Sumner J.C., Perotti M.A., Nesvorna M., Hubert J. 2017. Comparison of bacterial microbiota of the predatory mite Neoseiulus cucumeris (Acari: Phytoseiidae) and its factitious prey Tyrophagus putrescentiae (Acari: Acaridae). Sci. Rep., 7: 1-12. https://doi.org/10.1038/s41598-017-00046-6
- Rego R.O., Lopez J.E., Cabezas-Cruz A. 2020. Biological Drivers of Vector-Pathogen Interactions. Front. Cell. Infect. Microbiol., 10. https://doi.org/10.3389/fcimb.2020.609495
- Serpa L.L.N., Franzolin M.R., Barros-Battesti D.M., Kakitani I. 2004. Tyrophagus putrescentiae predando insetos adultos de Aedes aegypti e Aedes albopictus em laboratório. Rev. de Saude Publ., 38: 735-737. https://doi.org/10.1590/S0034-89102004000500019
- Texeira É.W., dos Santos L.G., Matioli A.L., Alves M.L.T.M.F. 2014. First report in Brazil of Tyrophagus putrescentiae (Schrank)(Acari: Acaridae) in colonies of Africanized honey bees (Apis mellifera L.). Interciencia, 39: 742-744.
- Walter, D.E., R.A. Hudgens and D.W. Freckman 1986. Consumption of nematodes by fungivorous mites Tyrophagus spp. (Acarina: Astigmata: Acaridae). Oecologia 70: 357-361. https://doi.org/10.1007/BF00379497
- Wilson A.J., Harrup L.E. 2018. Reproducibility and relevance in insect-arbovirus infection studies. Curr. Opin. Insect Sci., 28: 105-112. https://doi.org/10.1016/j.cois.2018.05.007
- Yang B., Cai J., Cheng X. 2011. Identification of astigmatid mites using ITS2 and COI regions. Parasitol. Res., 108: 497-503. https://doi.org/10.1007/s00436-010-2153-y



2021-09-06
Date accepted:
2021-12-02
Date published:
2022-02-08
Edited by:
McCoy, Karen

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Filatov, Serhii; Erhart, Jan and Rego, Ryan O.M.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)