



Water mites of the genus Hydrodroma Koch, 1837 (Acari, Hydrachnidia: Hydrodromidae) from Argentina, with description of two new species
Pešić, Vladimir1 and Smit, Harry2
1✉ Department of Biology, University of Montenegro, Cetinjski put b.b., 81000 Podgorica, Montenegro.
2Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, the Netherlands.
2022 - Volume: 62 Issue: 1 pages: 68-83
https://doi.org/10.24349/xmar-43rwZooBank LSID: 35607D3A-544C-4DA1-8639-088C0E8A157C
Original research
Keywords
Abstract
Introduction
Water mites of the genus Hydrodroma Koch, 1837 have been found in all biogeographic regions except Antarctica (see Smit 2020). Species of this genus are often abundant in different types of standing waters like lakes, ponds, ditches and pool areas of streams (Di Sabatino et al. 2010; Gerecke 2017).
To date, 33 species of the monotypic family Hydrodromidae are known worldwide (Zhi-Qiang et al. 2011; Pešić et al. 2021a, b; Pešić and Smit 2022), with 12 species and subspecies of the genus Hydrodroma described from South America (Lundblad 1941, 1953; K. Viets 1954a, b; Cook 1980, 1988). Hydrodroma stalagmophila Lundblad, 1941, and H. ungulata Lundblad, 1941, both described from southern Brazil (Lundblad 1941) are well distinguished by the absence of swimming setae. South American Hydrodroma with swimming setae on the legs have been traditionally assigned to two further species: populations with long, medially directed posteromedial projections of Cx-I were assigned to H. peregrina (Koenike, 1905), which includes the nominal subspecies and H. peregrina robusta K. Viets, 1954, described from northeastern Brazil (K. Viets 1954a). Populations with short, medially directed posteromedial projections of Cx-I were attributed to H. despiciens (Müller, 1776) and its subspecies H. despiciens brevirostris Lundblad, 1941, H. despiciens clavipes Lundblad, 1953, H. despiciens crassiseta K. Viets, 1954, H. despiciens longiseta K. Viets, 1954, H. despiciens macronyx K. Viets, 1954, H. despiciens micronyx K. Viets, 1954 and H. despiciens pauciseta Lundblad, 1954. Of these subspecies, H. clavipes, originally described from Colombia, raised to species rank by Cook (1980), the status of the others is still not clear, and as stated by Cook (1988) requires taxonomic revision.
In this paper we report and describe four Hydrodroma species from Argentina, two of these as new for science.
Material and Methods
Water mites were collected by hand netting, sorted live in the field, and immediately preserved in Koenike-fluid by the junior author. In the lab, selected specimens were dissected, and slide mounted in Faure's medium. Holotypes and some paratypes will be deposited in the Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Buenos Aires (MBR), some paratypes will be deposited in Naturalis Biodiversity Center in Leiden (RMNH).
All measurements are in µm. The photographs of ejaculatory complexes were made using a camera on a Samsung Galaxy smartphone. The following abbreviations are used: Ac = acetabulum; Cx-I = first coxa; dL = dorsal length; H = height; I-L-4-6 = fourth-sixth segments of first leg; L = length; NHRS = Swedish Museum of Natural History, Stockholm; NP = National Park; P-1-P-5 = palp segment 1-5; W = width.
Systematics
Family Hydrodromidae K. Viets, 1936
Genus Hydrodroma Koch, 1837
Diagnosis (based on Di Sabatino et al. 2010) — Idiosoma soft, without sclerotized dorsalia and ventralia, not laterally compressed wider than high. Cx-I+II medially separated by a fine membranous line; a row of long fine setae along the medial margins of Cx-I, and posterior margins of Cx-II, -III, and –IV. Genital flaps with more or less equally rounded lateral and concave medial margins, bearing fine setae and numerous Ac arranged in several rows along medial edge. Excretory pore sclerotized. Leg claws without claw blade, with a dorsal clawlet. P-2 with pectinate mediodistal setae. P-4 with a long, pointed extension reaching tip of the slender and elongate P-5.
Hydrodroma peregrina (Koenike, 1905)
Figures 1-2

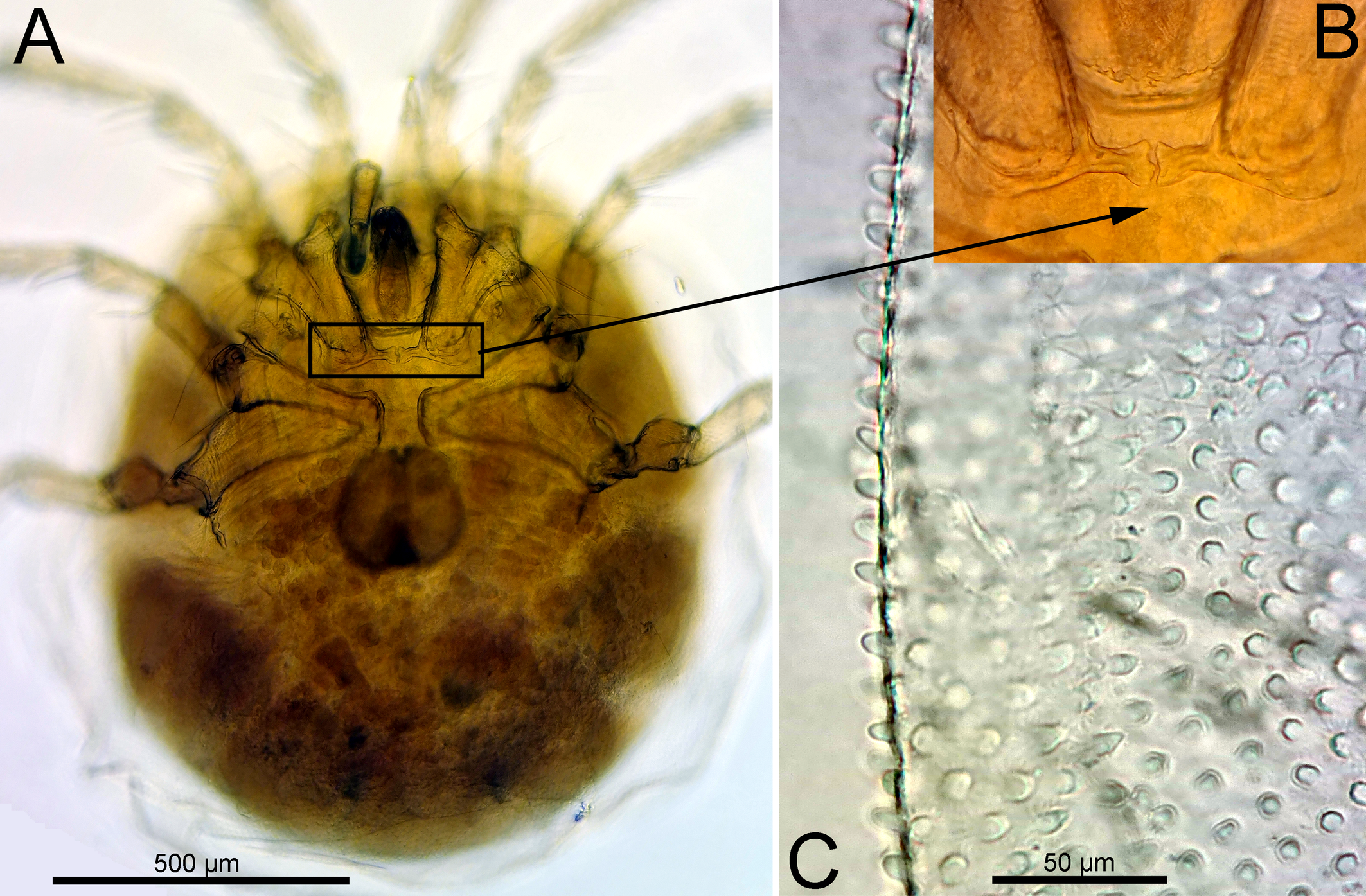
Material examined — Argentina, Prov. Río Negro, Río Negro near General Conesa, 9 Nov. 1999, leg. Smit, 1♀ dissected and slide mounted (RMNH).
Morphology — Female — Integument papillae apically rounded (Figure 1C). Cx-I+II with well-developed, medially directed projections of Cx-I separated by a fine membranous line as indicated in Figures 1A, 2B. Genital flaps each with 75 Ac in at most 6 longitudinal rows and 33 setae, all solid and flat (Figure 1B). Legs with relatively small claws (L 22-30; L ratio claw/segment 5, 6-11 %). Leg setae numbers: II-L-5 posterior 7; III-L-4 posterior 11; III-L-5 posterior 9; IV-L-4 anterior 11, posterior 12; IV-L-5 anterior 0, posterior 8-9. P-2 with three pectinate distal setae on the medial surface, dorsodistal seta long and slender (Figure 1C).
Measurements: Idiosoma L 1269, W 1080. Coxal field L 541; Cx-III W 559; L Cx-I+II, 288, Cx-III+IV, 311. Coxal setae numbers: Cx-I, 12; Cx-II, 20; Cx-III, 17-19; Cx-IV, 14-15. Genital field W 247, genital plate L 209, maximum W 109.
Gnathosoma vL 223; chelicera total L 303, L basal segment 244, claw 63, L ratio basal segment/claw 3.9. Palp total L 500, dL/H, dL/H ratio: P-1, 55/50, 1.1; P-2, 86/65, 1.33; P-3, 59/59, 1.0; P-4, 208/48, 4.3; P-5, 92/23, 3.9; L ratio P-2/P-4, 0.41. dL I-L-1-6: 72, 105, 128, 211, 250, 236; claw L 27, – 10.6 % L I-L-5. dL II-L-2-6: 125, 156, 272, 319, 269; claw L 22, – 6.9 % L I-L-5. dL III-L-2-6: 128, 159, 268, 306, 288; claw L 30, – 9.8 % L I-L-5. dL IV-L-1-6: 136, 184, 253, 372, 378, 317; claw L 30, – 7.9 % L I-L-5.
Remarks — Hydrodroma peregrina is often reported from the Neotropics, from Argentina to Mexico (Lundblad 1941; Cook 1980). It was originally described by Koenike (1905) from the Rio Grande do Sul in Brazil. In the original description, Koenike (1905) illustrated the coxal field but did not give neither an illustration of palp nor information on the leg setation. In describing specimens from Paraguay and Brazil, Lundblad (1941) stated that he did not find any differences with Koenike's type material. We were able to locate the type material of H. peregrina in the Koenike's collection in the Museum für Naturkunde der Humboldt-Universität Berlin. The type material was in very poor condition but it was possible to recognize the presence of the characteristic long, medially directed posteromedial projections of Cx-I. In regard to the shape of coxal field and palps our specimen agrees well with specimens of H. peregrina s. str. as illustrated by Lundblad (1941).
In describing specimens from Mexico, Cook (1980) mentioned that his specimens differ from each other in proportional size of the genital field, separating the large and the small genital field forms, respectively. Our specimen as well as the specimen from Brazil illustrated by Lundblad (1941) have genital field proportions more or less intermediate between those of the two Mexican forms (see Cook 1980).
Hydrodroma peregrina robusta K. Viets, 1954, a subspecies described by K. Viets (1954a) from northeastern Brazil differs from the nominal species in the absence of swimming setae on II-leg and the characteristic slender shape and setation of the palp (P-4 L/H ratio 6.0); P-2 with four long and slender, a hair-like mediodistal pectinate setae, see Figures 16a-b in K. Viets 1954a). In regard to these differences, there is no reason to support the assignment of this taxon as subspecies of H. peregrina and we propose to the taxon to the level of a separate species H. robusta stat. nov.
Distribution — Brazil, Paraguay (Koenike 1905; Lundblad 1941), Mexico (Cook 1980), Argentina (this paper). It is possible that populations formerly attributed to H. peregrina represent several distinct species.
Hydrodroma longiseta K. Viets, 1954 stat. nov.
Figure 3

Material examined — Argentina, Prov. Entre Rios, ditch Brazo Largo, Delta de Paraná, N of Zarate, 33°54.621 S, 58°51.154 W, 6 Nov. 1999, leg. Smit, 1♂, dissected and slide mounted (RMNH).
Morphology — Male — Integument papillae apically rounded (Figure 3B). Posteromedial projections of Cx-I short, separated by a fine membranous line (Figure 3A). Genital flaps each with 62 Ac in at most 5 longitudinal rows, and 25 setae, all hollow (Figure 3A). Legs with relatively small claws (L 17-22 µm, ratio claw L/segment 5 L 10-13 %). Leg setae numbers: II-L-posterior 0; III-L-4 posterior 5-6; III-L-5 posterior 5; IV-L-4 anterior 5-6, posterior 5; IV-L-5 anterior 0, posterior 4. P-2 with three pectinate distal setae on medial surface, dorsodistal seta hair-like, long and slender (Figure 3B).
Measurements: Idiosoma L 806, W 650. Coxal field L 353; Cx-III W 347; L Cx-I+II, 183, Cx-III+IV, 191. Coxal setae numbers: Cx-I, 14-15; Cx-II, 17; Cx-III, 15-16; Cx-IV, 12. Genital plate L 156-162. Ejaculatory complex L 130.
Gnathosoma vL 152; chelicera total L 193, L basal segment 153, claw 41, L ratio basal segment/claw 3.8. Palp total L 325, dL/H, dL/H ratio: P-1, 34/31, 1.13; P-2, 58/45, 1.27; P-3, 28/39, 0.72; P-4, 138/30, 4.6; P-5, 67/26, 2.6; L ratio P-2/P-4, 0.42. dL I-L-2-6: 64, 83, 128, 158, 159; claw L 20, – 12.8 % L I-L-5. dL II-L-2-6: 75, 94, 159, 190, 191; claw L 20, – 10.7 % L I-L-5. dL III-L-1-6: 47, 72, 91, 150, 169, 163; claw L 17, – 10.2 % L I-L-5. dL IV-L-2-6: 100, 128, 197, 217, 200; claw L 22, – 10.1 % L I-L-5.
Remarks — Our specimen is provisionally assigned to Hydrodroma longiseta K. Viets, 1954. This taxon was originally described by K. Viets (1954a) as a subspecies of H. despiciens. Hydrodroma longiseta resembles H. despiciens in shape of the coxal field, with short and medially directed posteromedial projections of Cx-I, but differs in the absence of swimming setae on II-L and in the shape of the palp with a long and slender, hair-like dorsodistal pectinate seta on the medial surface of P-2 (see Figures 14a-c in K. Viets 1954a). In regard to these differences, there is no reason to support the assignment of this taxon as subspecies of H. despiciens. In the shape of the coxal field and the palp our specimen from Delta de Paraná matches the original description. Anyhow, the variability of more specimens including material from the locus typicus needs to be examined to clarify the taxonomy of this species.
Distribution — Brazil („H. despiciens longiseta'' in K. Viets 1954a), Argentina (this study).
Hydrodroma argentinensis Pešić & Smit sp. nov.
ZOOBANK: 87C5C9D7-5081-4A02-87FE-0774D298DBA1 ![]()
Figures 4-6
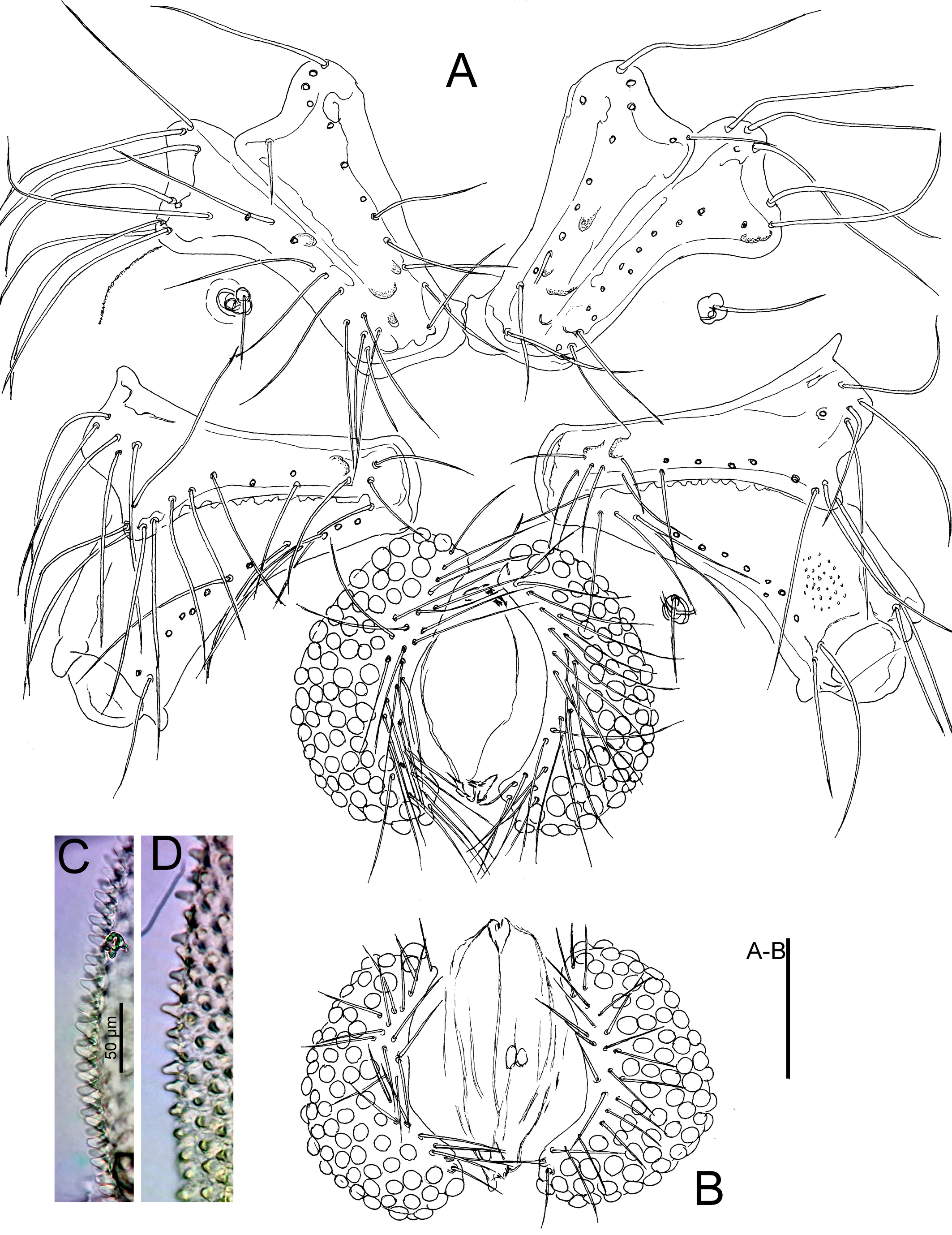


Material examined — Holotype ♂ (MBR), Argentina, Prov. Río Negro, Manso River, Nahuel Huapi NP, 41º20.858 S, 71º36.300 W, alt. 817 m, 17 Feb. 2018, leg. Smit, dissected and slide mounted. Paratypes: 2♂, 2♀, same data as holotype, 1♀ dissected and slide mounted (RMNH); Argentina, Prov. Río Negro, Rio Manso Medio near Cascada Los Alerces, Nahuel Huapi NP, 16 Nov. 1999, leg. Smit, 1♂, 6♀ (MBR).
Other material — Argentina: Prov. Neuquén, Río Collón Curá, 30 km E of Junín de los Andes, 39°59.8333 S, 70°50.1075 W, 12 Nov. 1999, leg. Smit, 1♀ (RMNH); Prov. Neuquén, small lake ± 15 km N of Villa La Angostura, along Rutá 234, Lanín NP, 12 Nov. 1999, leg. Smit, 3♂, 12♀ (1♂, 1♀ dissected and slide mounted, RMNH).
Diagnosis — Integument papillae of two types: main papillae longish, each papilla surrounded by six very flat elevations forming a hexagonal pattern, in tangential view apically rounded (Figures 4C-D, 6C-D). Posteromedial projections of Cx-I short. Genital plates with 58–81 pairs of Ac in at most 5 longitudinal rows. Legs with moderately long claws (L 31-34 µm; L ratio claw/segment 5, 10-15%). Leg setae numbers: II-L-posterior 0; III-L-4 posterior 7-10; III-L-5 posterior 6-8; IV-L-4 anterior 7-10, posterior 6-9; IV-L-5 anterior 0-3, posterior 5-7.
Description — Posteromedial projections of Cx-I short, separated by a fine membranous line. Genital flaps with rounded lateral and strongly concave medial margins, each with 63 (holotype, paratypes and other specimens: 58-81) Ac, and 31 (25-38) setae, with sexual dimorphism (♂: hollow, ♀: more solid and flat). P-2 with three pectinate mediodistal setae, similar in size.
Male (holotype; in parentheses some measurements of paratypes, n = 2; in square brackets some measurements of the specimen from small lake ± 15 km N of Villa La Angostura): Idiosoma L 894 (950) [1190], W 825. Coxal field L 463 (420) [531]; Cx-III W 509 (472) [652]; L Cx-I+II 222 (231) [266], Cx-III+IV 241 (234) [261]. Coxal setae numbers: Cx-I, 14 (11-13) [14]; Cx-II, 21 (18-20) [26]; Cx-III, 21 (16-21) [22]; Cx-IV 15-17 (14-17) [18]. Genital plate L 214-216 (216-219) [220-223]. Ejaculatory complex L [205].
Gnathosoma vL 180 [188]; chelicera total L 236 [261], L basal segment 195 [217], claw 46 [52], L ratio basal segment/claw 4.3 [4.2]. Palp total L 382 [407], dL/H, dL/H ratio: P-1, 48/34, 1.4 [45/36, 1.26]; P-2, 64/52, 1.24 [72/54, 1.33]; P-3, 39/52, 0.76 [41/56, 0.72]; P-4, 158/36, 4.4 [169/38, 4.5]; P-5, 73/19, 3.9 [80/19, 4.3]; L ratio P-2/P-4, 0.41 [0.43].
dL I-L-1-6: 50 [63], 78 [81], 100 [100], 161 [173], 219 [236], 225 [234]; claw L 33 [33] – 14.8 (13.4) [13.9] % L I-L-5. dL II-L-1-6: 55, 94, 125, 222, 272, 272; claw L 34, – 12.5 (11.1) % L I-L-5. dL III-L-1-6: 53, 106, 127, 211, 250, 254; claw L 33, 13.1 (11.1) % L I-L-5. dL IV-L-1-6: 100, 141, 177, 261, 284, 279; claw L 31, – 11 (10.2) % L I-L-5.
Female (paratype from Manso River, n = 1; in square brackets some measurements of the specimen from small lake ± 15 km N of Villa La Angostura): Idiosoma L 1016 [1000], W 818. Coxal field L 484 [477]; Cx-III W 603 [575]; L Cx-I+II, 234 [230], Cx-III+IV, 255 [242]. Coxal setae numbers: Cx-I, 16 [15]; Cx-II, 23 [22]; Cx-III, 20 [16]; Cx-IV, 16 [16]. Genital flaps large, L 183-189 [184].
Gnathosoma vL 191 [203]; chelicera total L 274 [261], L basal segment 213 [211], claw 59 [52], L ratio basal segment/claw 3.6 [4.1]. Palp total L 423 [422], dL/H, dL/H ratio: P-1, 48/41, 1.2 [45/38, 1.2]; P-2, 70/55, 1.27 [70/50, 1.39]; P-3, 42/52, 0.82 [43/52, 0.84]; P-4, 180/39, 4.6 [180/36, 5.0]; P-5, 83/17, 4.8 [84/19, 4.5]; L ratio P-2/P-4, 0.39 [0.39].
dL I-L-1-6: 58 [59], 84 [81], 106 [97], 183 [172], 241 [234], 238 [229]; claw L 33 [34], – 13.5 [14.5] % L I-L-5. dL II-L-1-6: 56, 105, 136, 245, 294, 284; claw L 33, – 11.1 % L I-L-5. dL III-L-1-6: 64, 108, 135, 222, 278, 272; claw L 32, – 11.5 % L I-L-5. dL IV-L-1-6: 98, 150, 194, 287, 325, 299; claw L 34, – 10.5 % L I-L-5.


Etymology — Named after the country where the new species was collected.
Discussion — In regard to the shape of the coxal field with relatively short, medially directed posteromedial projections of Cx-I, the new species resembles H. despiciens, a species considered previously to have a nearly cosmopolitan distribution (see Gerecke 2017 for a discussion). In former times, Hydrodroma populations from Brazil (Lundblad 1941), Argentina (Cook 1980) and Chile (Cook 1988) have been attributed to H. despiciens s. str., while six additional subspecies of the latter were introduced for other South American populations (see below). Recently, H. despiciens was redescribed by Gerecke (2017) allowing a reliable identification and enhancing the taxonomic status of populations previously assigned to that species. Studies on extra-European populations, e.g., from Australia (Pešić and Smit 2007a, b, 2011) have already shown that the concept of a cosmopolitan distribution of H. despiciens must be rejected.
In addition to the relatively smaller claw (mean L in males 27-29, in females 29-30 µm; L ratio claw/segment 5, 8-12%; data taken from Gerecke 2017), H. despiciens, can be separated from the new species from Argentina by the presence of one swimming seta on II-L-5 and an increased number of swimming setae on III/IV-L (more than 10 on both posterior III-L-4 and anterior IV-L-4). Moreover, H. despiciens differs in the absence of swimming setae from anterior IV-L-5. However, swimming setae numbers of single segments may overlap and the numbers of all segments must be considered together.
Based on the original descriptions, we give here a brief overview of the South American subspecies of H. despiciens with their main features (if not mentioned otherwise, data taken from original descriptions or calculated from figures given therein). Cook (1988) stated that it is more likely that some of these subspecies deserve to be ranked as distinct species.
Hydrodroma despiciens brevirostris Lundblad, 1941, was described from a pond near S. Antonia in Villarica, Paraguay. Based on the original description, the female holotype (prep. 23331, NHRS) is characterized by the presence of swimming setae on II-L. Two other females (preps. 2330 and 2332, both NHRS) included by Lundblad (1941) in the type series, however, probably are not conspecific with the holotype. Both paratypes, as mentioned by Lundblad (1941) do not possess swimming setae on II-L and dorsodistal seta on the medial surface P-2 is longer and slender (hair-like) as in H. longiseta (see above for a description). K. Viets (1954a) already suggested that these females should be assigned more correctly to H. longiseta.
Hydrodroma despiciens pauciseta Lundblad, 1944 was described from a stream in Sanibeni (Peru) on the basis of a single female. The characteristic feature mentioned by Lundblad (1944) for this subspecies is the reduced number of swimming setae (III-L-4, 1; III-L-5, 2; IV-L-4, 1-2; IV-L-5, 1).
Hydrodroma despiciens clavipes Lundblad, 1953 was described from Colombia, and later on considered by Cook (1980) a separate species on the basis of material collected from Mexico. In regard to the absence of swimming setae on II-L and a strongly reduced swimming setation on III/IV-L (one single, short seta each III-L-4/5, and IV-L-4/5) this species seems to be closely related to H. despiciens pauciseta Lundblad, 1944.
Hydrodroma despiciens macronyx K. Viets, 1954 was described from the river Beberibe in the Brazilian state Pernambuco. In addition to the absence of swimming setae on II-L and reduced swimming setae numbers on III/IV-L (III-L-4 with 3-5 swimming setae; IV-L-4 with 3 in ♂, 6-7 in ♀; IV-L-5 with one in ♂, 3 swimming setae in ♀) this subspecies is characterized by large leg claws (42 µm), shortened swimming setae on III/IV-L and slender palpal segments, especially P-4 (L/H ratio P-4 6.3).
Hydrodroma despiciens crassiseta K. Viets, 1954, was described from the Brazilian Amazon region (Igarapé do Guaranazal). Based on the original description, this subspecies differs from other subspecies of H. despiciens by an increased number of swimming setae on II-L (II-L-4 with 4; II-L-5 with 10 swimming setae). Hydrodroma despiciens micronyx K. Viets, 1954, was described from a swamp in Recife, northeastern Brazil. It resembles H. despiciens crassiseta from which it differs in reduced numbers of swimming setae on II-L (II-L-4 with one, II-L-5 with two swimming setae).
Variability — We found considerable variation in the number of swimming setae on the anterior IV-L-5 between populations from rivers and lakes (a small lake in Lanín NP). Most specimens (eight out of 15) from the small lake N of Villa La Angostura did not have swimming setae on anterior IV-L-5, while other specimens bear here at least one (rarely two) swimming seta(e) at least on one leg; three specimens possessed swimming setae on anterior IV-L-5 on both legs. In the population from the Manso River most specimens possessed 1-2 swimming setae on the anterior IV-L-5 but several specimens were without swimming setae on anterior IV-L-5 on one or both legs.
Distribution — Argentina.
Hydrodroma ventana Pešić & Smit sp. nov.
ZOOBANK: 2018E6DD-D27A-4202-A2AF-103611065A51 ![]()
Figures 7-9

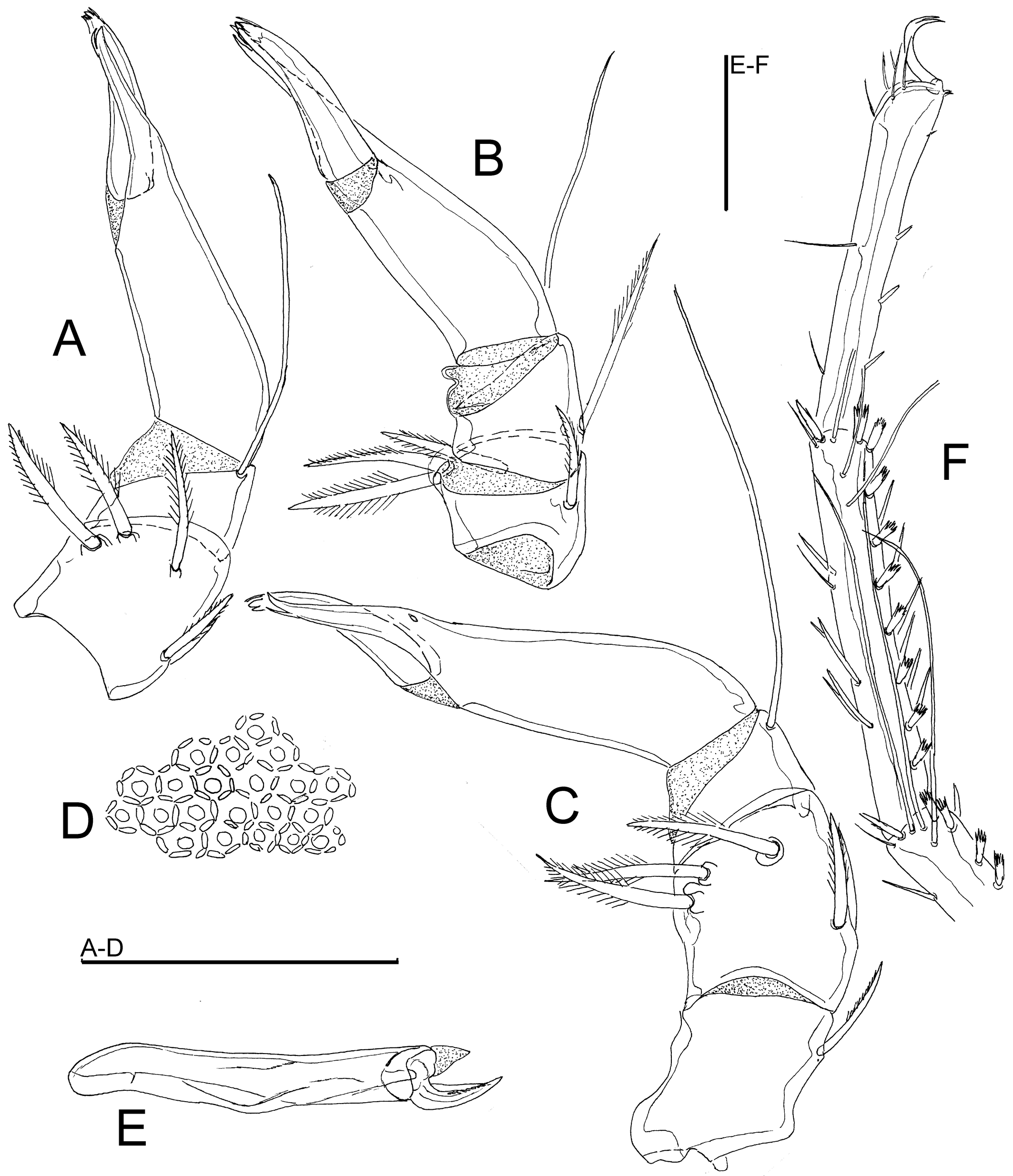
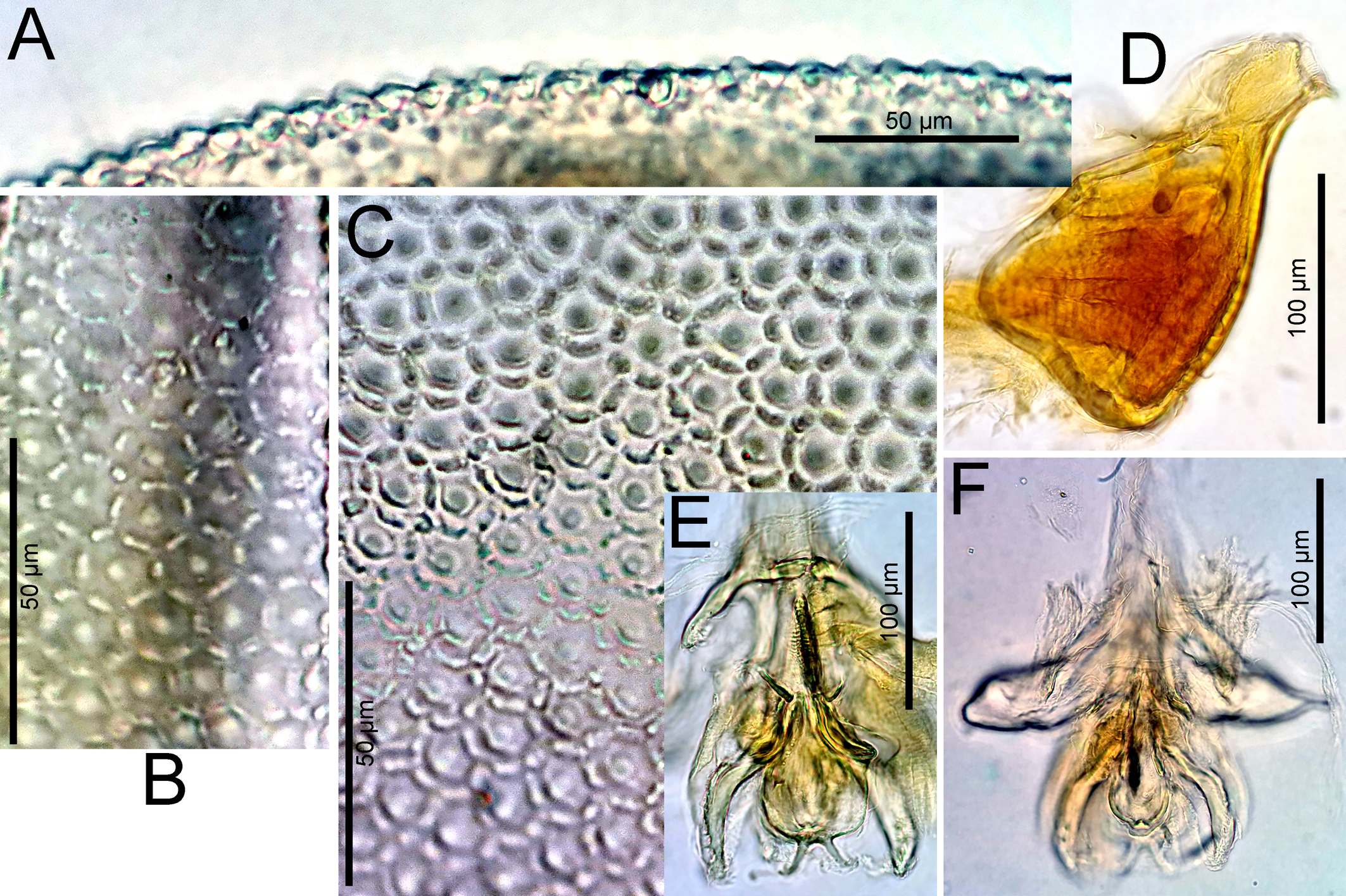
Material examined — Holotype ♂ (NBR), Argentina, Prov. Buenos Aires, Arroyo Sauce Grande, Sierra de la Ventana, 10 Nov. 1999, leg. Smit, dissected and slide mounted. Paratypes: 2♂, 2♀, same place and data as holotype, 1 ♂, 1♀ dissected and slide mounted (RMNH); 1♀, Arroya Naposté, Sierra de la Ventana, Estancia Funke E of Torquist, 38°07.998 S, 62°05.229 W, alt. 334 m, 8 Mar. 2018, leg. Smit (NBR).
Diagnosis — Main integument papillae flattened, surrounded by six small, little prominent elevations (Figures 9B-C), in tangential view short and apically rounded (Figure 9A). Posteromedial projections of Cx-I short. Genital plates with 37–45 pairs of Ac in at most 4 longitudinal rows. Legs with relatively long claws (L 39-42; L ratio claw/segment 5, 13-22 %). Leg setae numbers: II-L-posterior 0; III-L-4 posterior 2-4; III-L-5 posterior 2-3; IV-L-4 anterior 1-3, posterior 3-4; IV-L-5 anterior 0, posterior 1-3.
Description — Posteromedial projections of Cx-I short, medially separated by a fine membranous line. Genital flaps with rounded lateral and strongly concave medial margins, each with 37-45 Ac and 23-28 hollow setae. P-2 with three pectinate mediodistal setae similar in size.
Male (holotype; in parentheses some measurements of paratype, n = 1): Idiosoma L 1112 (875), W 931 (838). Coxal field L 447; Cx-III W 470; L Cx-I+II 230, Cx-III+IV, 219. Coxal setae numbers: Cx-I, 17 (15); Cx-II, 23 (23); Cx-III, 14 (12); Cx-IV, 14-15 (17). Genital plate L 188 (173). Ejaculatory complex (184).
Gnathosoma vL 167; chelicera total L 239, L basal segment 188, claw 52, L ratio basal segment/claw 3.6. Palp total L 328, dL/H, dL/H ratio: P-1, 42/33, 1.27; P-2, 50/47, 1.07 (61/48, 1.26); P-3, 33/42, 0.78 (31/47, 0.67); P-4, 142/33, 4.3 (147/34, 4.3); P-5, 61/17, 3.5 (61/16, 3.9); L ratio P-2/P-4, (0.41). dL I-L-2-6: 66, 88, 144, 177, 191; claw L 39, – 22.0 % L I-L-5. dL II-L-2-6: 91, 119, 188, 223, 223; claw L 39, – 17.5 % L I-L-5. dL III-L-2-6: 94, 125, 188, 215, 209; claw L 40.6, – 18.9 % L I-L-5. dL IV-L-2-6: 125, 172, 253, 266, 231; claw L 42.2, – 15.9 % L I-L-5.
Female: Idiosoma L 1395, W 1156. Coxal field L 528; Cx-III W 506; L Cx-I+II 263, Cx-III+IV, 264. Coxal setae numbers: Cx-I, 14-18; Cx-II, 24; Cx-III, 12 Cx-IV, 16. Genital plate large, L 192.
Gnathosoma vL 213; chelicera total L 275, L basal segment 217, claw 59, L ratio basal segment/claw 3.7. Palp total L 371, dL/H, dL/H ratio: P-1, 42/42, 1.0; P-2, 70/52, 1.36; P-3, 38/48, 0.77; P-4, 155/34, 4.5; P-5, 66/17, 3.8; L ratio P-2/P-4, 0.45. dL I-L-2-6: 77, 105, 170, 216, 200; claw L 41, – 18.8 % L I-L-5. dL II-L-2-6: 100, 131, 226, 263, 228; claw L 42, – 16.0 % L I-L-5. dL III-L-2-6: 116, 141, 212, 250. 228; claw L 42.2, – 16.9 % L I-L-5. dL IV-L-2-6: 147, 200, 293, 313, 265; claw L 41, – 13.0 % L I-L-5.

Etymology — Named after the Sierra de la Ventana, the name is a noun in apposition.
Discussion — The new species resembles H. argentinensis sp. nov. in the absence of swimming setae on II-L and a similar shape of the palp (L/H ratio P-4 ˂ 6.0; three pectinate distal setae on P-2 similar in size). The new species differs from H. argentinensis sp. nov. in a lower number of genital acetabula (˂ 50 Ac), reduced number of swimming setae on III/IV-L (less than 4 on both posterior III-L-4 and anterior IV-L-4), legs with a relatively large-sized claw (L ratio claw/segment 5, 13-22 %) and in the shape of integument papillae (more flattened, in tangential view short, and less densely arranged than in H. argentinensis sp. nov., compare figures 4C-D with 9A). Moreover, the new species differs in the absence of swimming setae on the anterior IV-L-5 (see under H. argentinensis sp. nov. about variation in the swimming setation of the latter species).
In comparison with the six known South American subspecies of H. despiciens, the new species is most similar to H. despiciens macronyx K. Viets, 1954a, described from northeast Brazil (Pernambuco). The latter subspecies is distinguished from the new species from Argentina in more slender palp segments, especially P-4 (L 208 µm; L/H ratio P-4 6.3) and shortened swimming setae on III/IV-L.
Distribution — Argentina.
Acknowledgements
HS is indebted to the Direccion de Conservacion y Manejo (Buenos Aires) for their permission to collect in the national parks of Argentina. Truus van der Pal (Alkmaar) assisted with the field work. We are indebted to Anja Friederichs (Museum für Naturkunde der Humboldt-Universität Berlin) for her help in finding the type material of Hydrodroma peregrina. We thank Joanna Mąkol (Wrocław) and two anonymous reviewers, whose constructive comments greatly improved this work.
References
- Cook D.R. 1980. Studies on neotropical water mites. Mem. Amer. Ent. Inst., 31: 1-645.
- Cook D.R. 1988. Water mites from Chile. Mem. Amer. Ent. Inst., 42: 1-356.
- Di Sabatino A., Gerecke R., Gledhill T., Smit H. 2010. Chelicerata: Acari II. In: Gerecke R, editor. Chelicerata: Araneae, Acari I. Süßwasserfauna von Mitteleuropa, 7, 2-2: 1-134. München: Elsevier Spektrum Akademischer Verlag. https://doi.org/10.1007/978-3-8274-2266-8_1
- Gerecke R. 2017. The water mites of the genus Hydrodroma (Acari, Hydrachnidia, Hydrodromidae) in Europe and Africa. Ecol. Monten., 13: 1-24. https://doi.org/10.37828/em.2017.13.1
- Koenike F. 1905. Zwei neue Wassermilben aus den Gattungen Megapus und Diplodontus. Zool. Anz., 28): 694-698.
- Lundblad O. 1941. Die Hydracarinenfauna Südbrasiliens und Paraguays. Erster Teil. Svensk. Vetenskapsakad. Handling., Stockholm (s.3), 19 (7): 1-183.
- Lundblad O. 1944. Einige neue und wenig bekannte Hydracarinen aus Südamerika. Ent. Tidskr., 65 (3-4): 135-162.
- Lundblad, O. 1953. Die Hydracarinenfauna von Colombia. Ark. Zool. (s.2), 5 (8): 435-585.
- Pešić V., Smit H. 2007a. Water mite species of the genus Hydrodroma Koch (Acari: Hydrachnidia, Hydrodromidae) from Australasia. Part I. Zootaxa, 1389: 31-44. https://doi.org/10.11646/zootaxa.1389.1.2
- Pešić V., Smit H. 2007b. Water mite species of the genus Hydrodroma Koch (Acari: Hydrachnidia, Hydrodromidae) from Australia. Part II. Zootaxa, 1509: 41-50. https://doi.org/10.11646/zootaxa.1509.1.4
- Pešić V., Smit H 2011. A new species of the genus Hydrodroma Koch, 1837 (Acari, Hydrachnidia, Hydrodromidae), with a key to the hitherto known six species of the genus in Australia. ZooKeys, 143: 13-22. https://doi.org/10.3897/zookeys.143.2115
- Pešić V., Smit H. 2022. Hydrodroma angelieri (Acari, Hydrachnidia: Hydrodromidae) a new water mite species from Corsica based on morphological and DNA barcode evidence. Acarologia, 62(1): 3-11. https://doi.org/10.24349/l06c-j0qm
- Pešić V., Smit H., Mary N.J. 2021a. Two new water mite species of the genus Hydrodroma Koch, 1837 from New Caledonia (Acari, Hydrachnidia: Hydrodromidae). Acarologia, 61: 581-590. https://doi.org/10.24349/aGHX-uIU1
- Pešić V., Zawal A., Saboori A., Smit H. 2021b. New records of water mites (Acari, Hydrachnidia) from Iran with the description of one new species based on morphology and DNA barcodes. Zootaxa, 5082: 425-440. https://doi.org/10.11646/zootaxa.5082.5.2
- Viets, K. 1954a. Süßwassermilben (Hydrachnellae, Acari) aus Südamerika. Arch. Hydrobiol., 49 (1/2): 1-224. https://doi.org/10.1007/BF00023587
- Viets K. 1954b. Wassermilben aus dem Amazonasgebiet (Hydrachnellae, Acari).(Systematische und ökologische Untersuchungen). Bearbeitung der Sammlungen Dr. R. Braun, Aarau und Dr. H. Sioli, Belém. Schweiz. Z. Hydrol., Basel, 16(1): 78-151. https://doi.org/10.1007/BF02485767
- Zhang Z.-Q., Fan Q.-H., Pešić V., Smit H., Bochkov A.V., Khaustov A.A., Baker A., Wohltmann A., Wen T.H., Amrine J.W., Beron P., Lin J., Gabrys G., Husband R. 2011. Trombidiformes. In: Z.-Q. Zhang (ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa, 3148: 129-138. https://doi.org/10.11646/zootaxa.3148.1.24



2021-11-02
Date accepted:
2022-01-13
Date published:
2022-01-17
Edited by:
Mąkol, Joanna

This work is licensed under a Creative Commons Attribution 4.0 International License
2022 Pešić, Vladimir and Smit, Harry
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)