



A new genus and new species of macronyssid mite (Mesostigmata: Gamasina: Macronyssidae) from Brazilian caves including molecular data and key for genera occurring in Brazil
Gomes-Almeida, Brenda K.1 and Pepato, Almir R.2
1Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Zoologia, Pós-graduação em Zoologia/Laboratório de Sistemática e Evolução de Ácaros Acariformes. Av. Antônio Carlos, 6627, Pampulha, Belo Horizonte, MG, Brasil, CEP 31270–901.
2✉ Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas, Departamento de Zoologia, Pós-graduação em Zoologia/Laboratório de Sistemática e Evolução de Ácaros Acariformes. Av. Antônio Carlos, 6627, Pampulha, Belo Horizonte, MG, Brasil, CEP 31270–901 & Tyumen State University, 10 Semakova Str., 625003 Tyumen, Russia.
2021 - Volume: 61 Issue: 3 pages: 501-526
https://doi.org/10.24349/acarologia/20214447ZooBank LSID: 9C3C87C0-5371-4A8E-80B1-2F4E89632BDC
Original research
Keywords
Abstract
Introduction
The family Macronyssidae Oudemans, 1936 includes mites that can be associated with a variety of vertebrates, such as birds, reptiles and mammals, including humans (Radovsky 2010; Orlova et al. 2017; Bassini-Silva et al. 2019). The life cycle of these mites comprises five stages of development: egg, larva, protonymph, deutonymph and adults (male or female) (Radovsky 1966; 1967). Of these, only protonymph and adults are hematophagous, other stages are inactive and can be found on the substrate or roost of hosts (Radovsky 2010).
Radovsky (2010) reviewed the Macronyssidae family, recognizing seven new genera for the family, from new species discovered by him or by splitting other genera, mainly Ornithonyssus Sambon, 1928, which included at least a third of the Macronyssid genera (Zumpt and Till 1953; Fonseca 1954; 1960; Radovsky 1966; 1969; Micherdzinski 1980; Shepherd and Narro 1983).
Currently, Macronyssidae comprises 34 valid genera, with occurrences worldwide. Of these, at least 23 genera occur in the Neotropical region associated with different host taxa: Synasponyssus Radovsky and Fuman 1969, Parichoronyssus Radovsky 1966, Radfordiella Fonseca 1948, Chiroecetes Herrin and Radovsky 1974, Mitonyssoides Yunker, Lukoschus and Giesen 1990, Macronyssoides Radovsky 1966, Nycteronyssus Saunders and Yunker 1973, Mitonyssus Yunker and Radovsky 1980, Cryptonyssus Radovsky 1966, Chiroptonyssus Augustson 1945, and Macronyssus Kolenati 1858 are associated exclusively with bats; Steatonyssus Kolenati 1858 with bats and other mammals; Acanthonyssus Yunker and Radovsky 1966, Argitis Yunker and Saunders 1973, Lepidodorsum Saunders and Yunker 1975, Lepronyssoides Fonseca 1941 and Lagidonyssus Radovsky 2010 with rodents, despite Lepronyssoides can be found associated with other mammals; Pellonyssus Clark and Yunker 1956 with birds; Ornithonyssus Sambon 1928 with birds and mammals, including humans; Draconyssus Yunker and Radovsky 1966, Ophionyssus Megnin 1884, Endophionyssuus Radovsky 2010 on lizards and snakes; and, finally, Thigmonyssus Radovsky 2010 found associated with anteaters.
Thirteen macronyssid genera are recorded from Brazil thus far: Chiroptonyssus, Lepronyssoides, Macronyssoides, Macronyssus, Mitonyssoides, Mitonyssus, Ophionyssus, Ornithonyssus, Parichoronyssus, Pellonyssus, Radfordiella, Steatonyssus and Thigmonyssus (Bassini-Silva et al. 2020). They comprise twenty-seven species of mites occurring in localities in the Southeast (Minas Gerais, Rio de Janeiro and São Paulo), Middle-west (Mato Grosso, Mato Grosso do Sul and Goiás), Northeast (Alagoas, Bahia, Ceará, Maranhão, Paraíba, Pernambuco, Piauí and Rio Grande do Norte) and South (Paraná and Rio Grande do Sul) (Fonseca 1948; Fonseca 1954; Radovsky 1967; Whitaker Jr and Mumford 1977; Azevedo et al. 2002; Almeida et al. 2011; Nieri-Bastos et al. 2011; Moras et al. 2013; Silva et al. 2017; Bassini-Silva et al. 2019; 2020; Oliveira et al. 2020).
Herewith, we aim at describing a new genus and new species based on adults and protonymphs found in caves located in the states of Minas Gerais, Rio de Janeiro (Southeastern) and Pará (Eastern Amazon region).
Material and methods
Specimens were collected during surveys for the subterranean fauna, in iron ore and limestone caves in Minas Gerais and Rio de Janeiro states, Southeastern Brazil and from caves in Pará state, North Brazil (Figure 1A). Mites were found freely dwelling on the cave floor, from entrance, penumbra and aphotic zone, during the dry and rainy seasons (Table 1; Figure 1B).


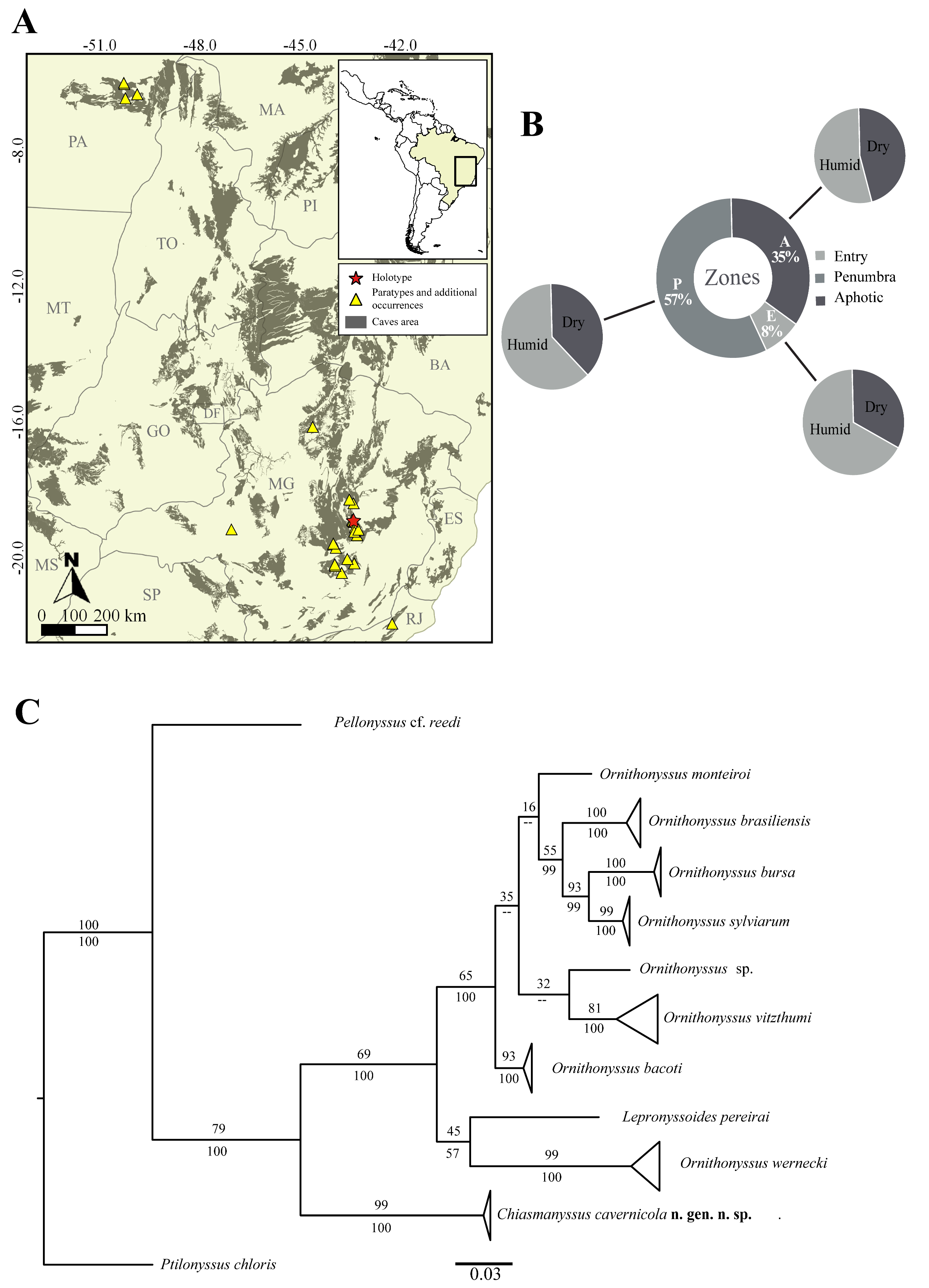
Specimens were mounted as permanent microscope slides using Hoyer's medium. Measurements and illustrations were made using the Leica DM 750 optical microscope with an ICC50 W digital camera attached. All measurements are given in micrometers (μm). Illustrations were prepared using the live image overlay technology adapted from Sidorchuk and Vorontsov (2014). Map was produced with QGIS 3.18.1 program and figures were prepared with Krita v.4.4.2 program. Terminology of dorsal and ventral chaetotaxy and the nomenclature of the dorsum based on Radovsky (2010). The terminology of sternal pores was reported according to Evans and Till (1979).
Holotype, paratypes and additional individuals (Supplementary material 1) are deposited at the Acarological Collection, Centro de Coleções Taxonômicas, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte City. Collection acronym: UFMG AC.
Molecular procedures
DNA extraction — Specimens were preserved in 95–100 % ethanol and stored at −20 °C. Genomic DNA was extracted from single specimens using a QIAamp DNA Micro kit (Qiagen) following the manufacturer's protocol, except by using two steps of the final elution, leading to a final volume of 50 µl.
16S rRNA gene amplification — A fragment of 16S rRNA was chosen due its availability for other Macronyssidae, since was employed in previous studies on the Macronyssid genus Ornithonyssus. The amplification of mitochondrial large ribosomal subunit (16S) fragment of ~ 410 bp was conducted using the primers proposed by Mangold et al. (1998): 16S + 1 (5'-CTG CTC AAT GAT TTT TTA AAT TGC TGT GG-3'); 16S–1 (5' -CCG GTC TGA ACT CAG ATC AAG T-3'). Amplifications were performed in 20 µl of final volume, with Platinum Taq DNA Polymerase (Invitrogen) in a Mastercycler nexus (Eppendorf) thermocycler. Master mix for initial PCR contained 2.0 µl of PCR buffer (1X), 1.4 µl MgCl2 (50 mM), 1.4 µL of dNTP (10 mM each), 0.8 µl of each oligonucleotide primer. Amplifications used 1–3 µl of genomic DNA and included an initial denaturing step of 2 min at 94 °C, followed by 35 cycles, comprising: denaturing at 94 °C for 30s, annealing for 35s, and extension at 72 °C for 45 s. The annealing temperature for the first seven cycles increased in 0.3 °C every cycle, ranging from 47 °C to 48.8 °C, remaining 28 cycles used an annealing temperature of 50 °C. Final extension of 5 min. at 72 °C.
Sequencing and editing chromatogram — PCR products found positive in 1% agarose gel electrophoresis were purified using the Ampure® (Agencourt) kit and sequenced using a 3730 DNA Analyzer. Sequencing primers as for PCRs. Chromatograms were resolved in ChromasPro 1.41 (Technelysium Pty Ltd).
Sampling, alignment and 16S secondary structure — Newly obtained sequences and those downloaded from GenBank are reported in Table 2, along voucher material collection numbers. This fragment was chosen because it led to the largest sample of terminals identified to species and with vouchering data. It is beyond the objectives of this article perform a throughout phylogenetic analyses of Macronyssidae, something that would require more markers and terminals. The aim of including this fragment is just to verify if it supports the inclusion of the new genus in the relatively well sampled Ornithonyssus. Preliminary automatic alignment was performed using MUSCLE (Edgar 2004) and then manually aligned with aid of sequence editor BioEdit 7.2.1 (Hall 1999) considering the 16S secondary structure as reported for other arthropods (Smith and Bond 2003; Gillespie et al. 2006). A full annotated alignment is provided in the supplementary material. Therein, matching parentheses and dots were used to indicate stem and loop regions, respectively; structural helix numbering is given after Gillespie et al. (2006). Length variable regions inferred to be ambiguously aligned were classified into regions of expansion and contraction (REC), non-pairing regions of ambiguous alignment (RAA) and excluded from downstream phylogenetic analyses. Access to the Brazilian genetic heritage was registered in SisGen under register number A68A31E.
Molecular Phylogenetic Analysis — Model for sequence evolution was chosen using the AICc criterion implemented in jModeltest (Darriba et al. 2012). Maximum Likelihood inference was run in PhyML (Guindon et al. 2010) online at http://www.atgc-montpellier.fr/phyml/ (accessed January 18nd 2021) and Bayesian analyses in MrBayes 3.2.7 (Ronquist et al. 2012). Inter and intraspecific distances were calculated in MEGA7 (Kumar et al. 2016).
Result
DNA sequencing and analyses
The final alignment counted with 288 positions after having their 3' and 5' extremities trimmed, and regions of ambiguous alignment excluded. Among them, 186 positions were conserved across all macronyssid species and 78 parsimony informative. Whereas intraspecific mean p-distances among sequences varied from 0.0005 in O. sylviarum (Canestrini and Fanzago 1877) (n= 24) to 0.0094 in O. wernecki (Fonseca 1935a) (n= 3), among Ornithonyssus and Lepronyssoides species, interspecific p-distances varied from 0.0462 (O. bacoti (Hirst 1913) x O monteroi (Fonseca 1941) and 0.1435 (O. bursa (Berlese 1888) x O. wernecki and L. pereirai (Fonseca 1935a). Pairwise mean interspecific p-distances are summarized in Table 3. The results suggest the 16S fragment sequenced here is a useful barcoding marker for molecular identification.

The model HKY+G was chosen using jModeltest and used in Maximum likelihood and Bayesian analyses. They agree to each other and similar to Nieri-Bastos et al. (2011) recovers L. pererai and O. wernecki in a basal position relative to remaining Ornithonyssus species (Figure 1C). Chiasmanyssus cavernicola n. gen. n. sp. branches basal in the topology obtained, being recovered in an isolated position relative to Ornithonyssus and Lepronyssoides species from which 16S sequences are available.
Taxonomy
Family Macronyssidae Oudemans, 1936
Diagnosis according to Radovsky (2010).
Chiasmanyssus n. gen.
ZOOBANK: 1C2CF818-9683-4DB2-BE34-DAD69DC435F5 ![]()
(Figures 3–10)
Type species: Chiasmanyssus cavernicola, new species.
Diagnosis — Female large, its sternal shield square, distinctly sclerotized, with a smoother area on central sternal shield X-shaped; sternal shield bearing three pairs of setae and two pairs of pores; without sternal glands; anterior median protusion in epigynal shield extends under the sternal shield; anterior spur on coxa II lacking. Adults with dorsal shield entire, reticulated and with j and J series complete. Adults and protonymphs with setae J5 minute; all setae on idiosoma, legs and gnathosoma slender and smooth, except palp with disto-ventral seta on genu spatulate. Females and protonymphs with chelae simple and slender, without spines or other processes. Male with four pairs of conspicuous pores near setae j3, z4, J3 and one laterally between J4 and J5 setae; peritrema short, extending from posterior margin of coxae II to the anterior margin of coxae IV and peritrematic shield with nodule fused to the middle of the shield.
Etymology — The generic name comes from Greek word ''chiasma'' referring to the distinct X-shaped area on sternal shield and ''nyssus'' , to prick, referring to the parasitic life.
Chiasmanyssus cavernicola n. gen. n. sp.
ZOOBANK: A34F4774-19C6-41E7-AFCB-023910A345A0 ![]()
Diagnosis — Female: Elongated dorsal shield, holotrichous, bearing 27 pairs of setae, all setae long and subequal, except j2, z2, s1–s2 small (and J5 minute). Unarmed dorsum with approximately 84 pairs of long setae and ventral opisthosoma with approximately 48 pairs of setae; peritrema long, extending from level of anterior margin of coxae II to the middle of coxae IV; palpal trochanter with a small blade-like ventral process. Male: anterior margin of the dorsal shield not discernible from the soft cuticle; with neotrichous condition on opisthonotal region; a pair of additional minute setae, laterally to J4 and J5, on neotrichous area of opisthonotal region. Four pairs of caudal setae on unarmed dorsum and four pairs of setae on unarmed venter. Protonymph: Subrectangular podonotal shield with lateral borders biconcave at level of z2 and z4, posterior region almost square; with j2 very close to j1. Trapezoidal pygidial shield with J4 and Z4 small than others (except J5 minute). Unarmed dorsum with 12 pairs of long setae. Unarmed venter with four pairs of median setae between ventral shields and a pair of caudal setae, without setae flanking the anus.
Description. Female — Measurements summarized in table 4.
(Figures 2–4)
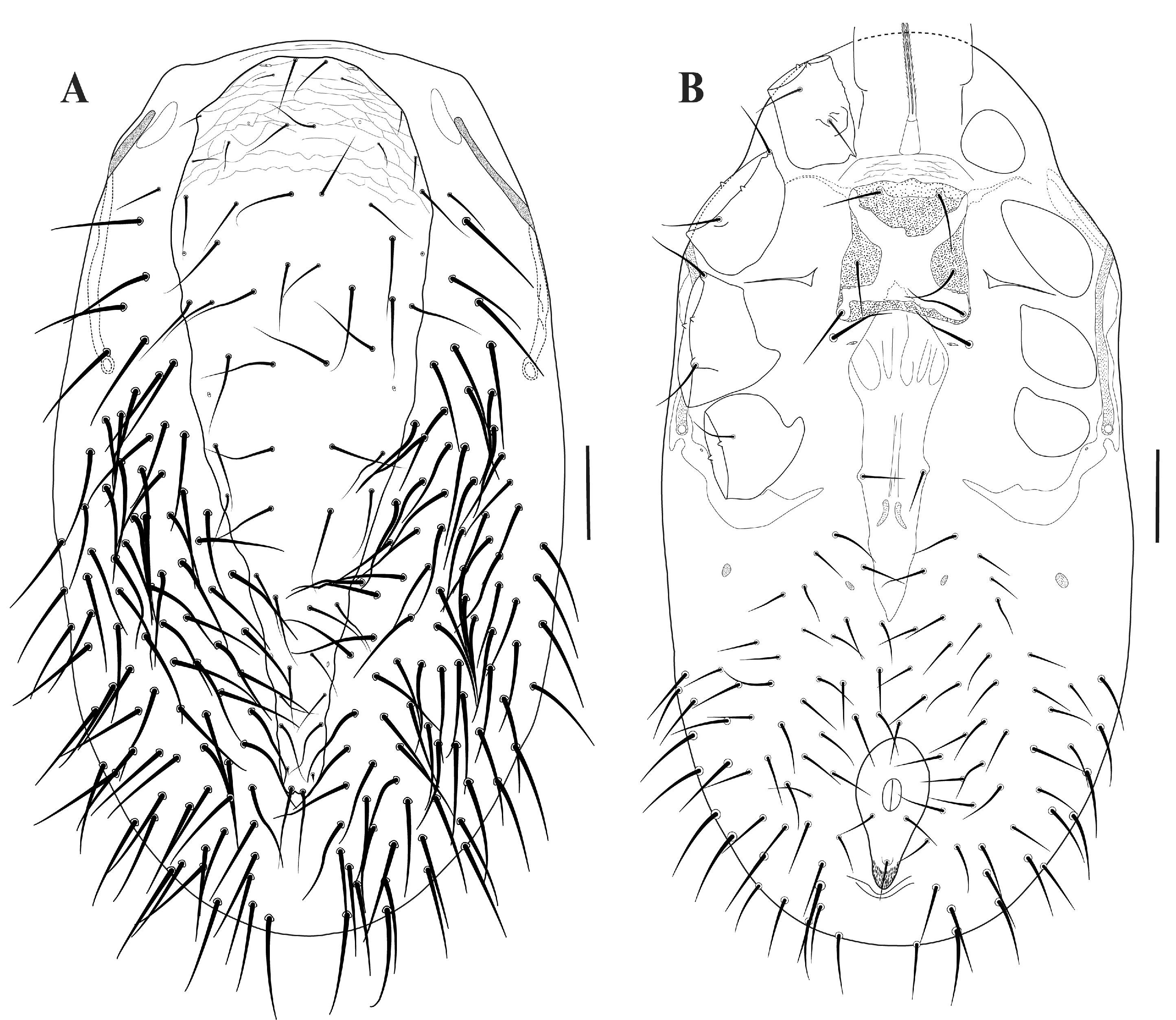


Idiosoma — (Figures 2A–B). Oval, moderately sclerotized and striated integument; all setae smooth. Dorsum (Figure 2A). Dorsal shield entire, elongated, longer than wide, and narrow, even at its widest, distinctly tapered in the middle of opisthonotal region; reticulated sculpture on all its area; holotrichous condition with 27 pairs of setae, 16 on podonotum and 11 on opisthonotum; j and J series complete with j1, j2 and j3 on dorsal shield; all setae long and subequal except j2, z2, s1–s2 small and J5 as microsetae (Figure 3A); one pair of lyrifissures near on margin lateral to setae j1 and four pair of pores on margin lateral to setae j3 and J4 and near s4 and z6. Unarmed dorsum with approximately 84 pairs of long setae. Venter (Figure 2B). Tritosternal base not expanded; Tritosternum bipartite with long and fimbriate laciniae (Figure 3B). Presternal area is weakly sculptured. Sternal shield nearly square, anterior margin with a small medial prominence; strongly sclerotized and punctate, except for a smoother X-shaped portion on central sternal shield (Figure 3C); anterior and posterior corners absent; three pairs of sternal setae relatively long, st1 and st2 on the sclerotized part and st3 inserted on X-shaped region on shield; two pairs of sternal pores. Metasternal setae (st4) on unarmed integument with third pair of pores accompanying metasternal setae. Epigynal shield expanded anteriorly, median projection extends under the sternal shield, no anterior scale-like markings; posterior margin pointed (Figure 3C); one pair of genital setae, approximately at the middle of epigynial plate. Anal plate pyriform; long and narrow cribrum; one pair of adanal setae (pan) and one postanal seta (pon) (Figure 3D). Endopodal plates small and triangular between level of coxae II and III. Opisthosoma with approximately 48 pairs of setae ventrally, slender, except caudal setae, being thicker, similar to the setae on unarmed dorsum. Peritreme long, extending from anterior margin of coxae II to the middle of coxae IV (Figure 2B). Peritrematic shield free from dorsal shield but interrupted by two ''nodules'' on the anterior dorsum between coxae I and II and in the middle ventral between coxae II and III (Figure 2A–B). Peritrematic shield posterior to the stigma fused with the parapodal shield around the posterior margin of coxa IV (Figure 2B).

Legs — Legs long and slender; legs I and IV longer than others; leg setae smooth. Coxae II and III may have posterior seta on a low protuberance; coxae II lacks anterodorsal spines; coxae lack ventral spurs or spur-like spines and ridges. Chaetotaxy of legs: I= 0 0/1 0/1 0 (2), 1 0/2 1/1 1 (6), 2 3/1 2/3 2 (13), 2 3/2 3/1 2 (13), 2 3/2 3/1 2 (13), 4 8/4 2/2 6/3 4 (33) + 6 solenidia + 1 macroseta; II= 0 0/1 0/1 0 (2), 1 0/2 0/1 1 (5), 2 3/1 2/2 1 (11), 2 3/1 2/1 2 (11), 2 2/1 2/1 2 (10), 3 3/2 1/1 3/2 3 (18); III= 0 0/1 0/1 0 (2), 1 1/2 0/1 0 (5), 1 2/1 1/0 1 (6), 2 2/1 2/1 2 (10), 2 1/1 2/1 2 (9), 3 3/2 1/1 3/2 3 (18) and IV= 0 0/1 0/0 0 (1), 1 1/2 0/1 0 (5), 0 2/2 1/0 1 (6), 2 2/1 3/1 1 (10), 2 1/1 3/1 2 (10), 3 3/2 1/1 3/2 3 (18). Sensory field of tarsus I has distal setae (d1–d2) on pedicels; setae s4 (solenidion) fused at base; two setae (solenidion) additional in tarsus (''s?''), a seta near ds and d2, and a very posterior seta; one macroseta, long and slender with distinctly wide socket on dorsal (seta ''m'', probable).
Gnathosoma — (Figure 4A–F). Deutosternal groove with nine denticles arranged 1–1–1–1–1–1–1–1–1. Gnathosoma with three pairs of hypostomal setae (hyp1, hyp2 and hyp3) slender and subequal and one pair of gnathosomal setae (pc). Trochanter–tarsus chaetotaxy formula 0/2 (2); 1 1/0 1/0 1/0 1 (5); 2 1/0 1/0 1/0 1 (6); 2 2/1 2/0 3/1 3 (14) and tarsus with 16 setae + apotele forked. Disto-ventral with circular (spatulate) and membranous seta (Figure 4C) and one pair of dorsal lyrifissure on posterior margin of palpal genu (Figure 4D). Palpal trochanter has a small blade-like ventral process, not extending beyond the segment (Figure 4E). Chelicera with chelae simple and slender, without spines or other process (Figure 4F).
Description. Male — Measurements summarized in table 4
(Figures 5–7)



Idiosoma — (Figure 5A–B). As in female but smaller. Dorsum (Figure 5A). Dorsal shield entire, occupying most of the dorsal idiosoma; anterior margin of the dorsal shield not discernible from the soft cuticle; posterior narrow to setae J5; reticulate sculpture; dorsal shield with j and J series complete, j1, j2 and j3 on dorsal shield and J5 minute, bears approximately 56 pairs of setae; neotrichous area on opistonothal region; a pair of additional minute setae, laterally to J4 and J5, on neotrichous area of opistonothal region; one pair of lyrifissures near on margin lateral to setae j1 and six pairs of pores visible on dorsal shield, four pairs of conspicuous pores near setae j3, z4, J3, one laterally to J4 and J5 setae, and two supplementary pairs positioned close together, in the dorsal shield margin, laterally the setae J2; four pairs of caudal setae on unarmed dorsum. Venter (Figure 5B). Tritosternum as in female. Venter has a holoventral shield; reticulated sculptured; bears two pairs of lyrifissures and three pairs of setae on sternal region; third pair of pores near the metasternal and genital setae on shield; ventral region broadened posteriorly, with 10–11 setae on each side. Anal region with three post-anal setae, cribrum wide and anal ring present. Endopodal plates small and triangular, between coxae III and IV. Unarmed ventre with four pairs of setae. Peritrema much shorter than in female, extending from posterior margin of coxa II to anterior margin of coxa IV. Peritrematic shield anteriorly fused to dorsal shield, with nodule fused to the middle of the shield. Peritrematic shield posterior to stigma as in females.
Legs (Figure 6A–C and Figure 7A–C). Legs as in female. Chaetotaxy of legs as in female, including tarsus I sensory field (Figure 7B–C).
Gnathosoma — As in female. Palpal trochanter lacks a ventral process and chelicera with spermatodactyl short (Figure 7D).
Description. Protonymph — Measurements summarized in table 4.
(Figures 8–9).

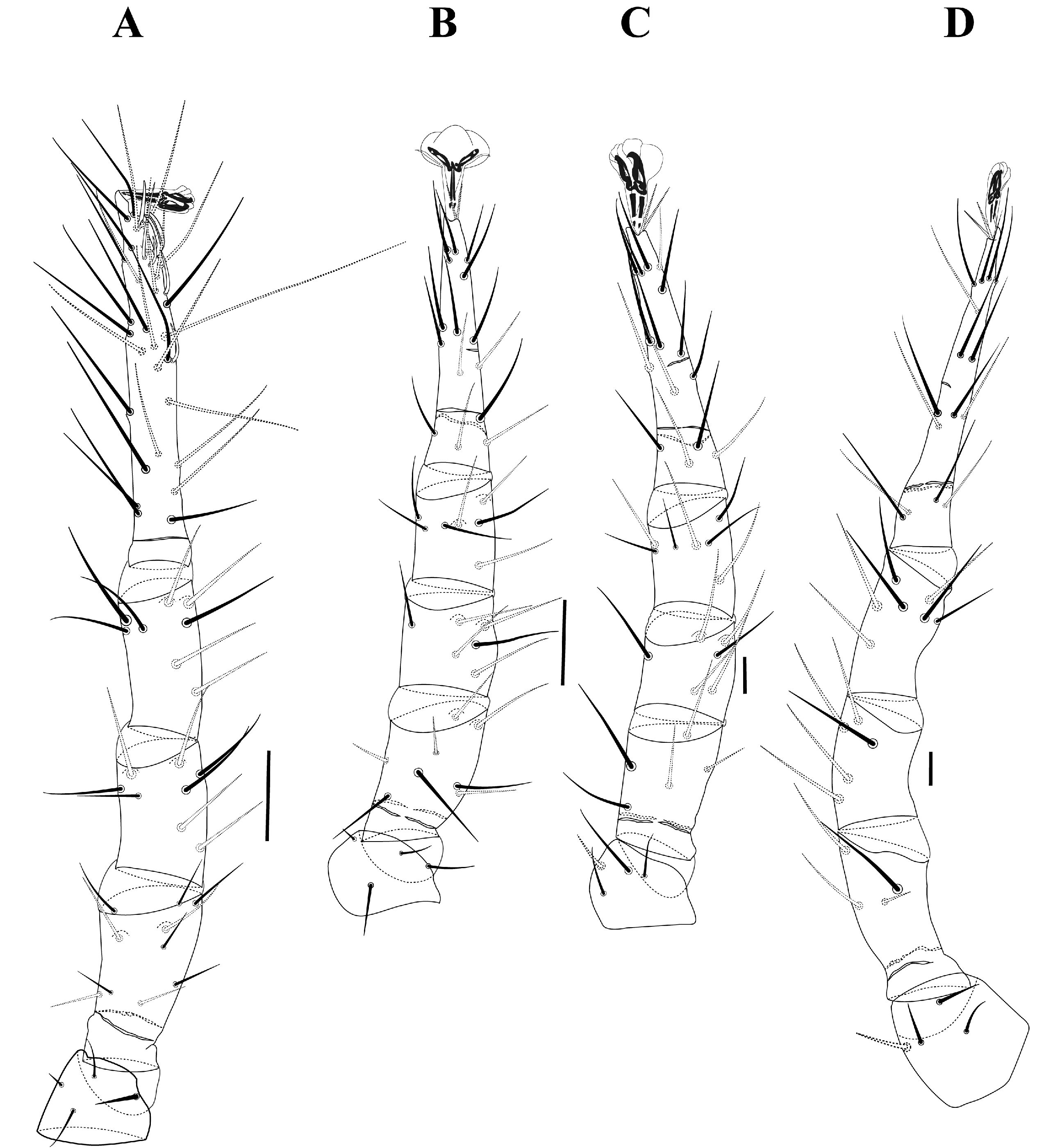
Idiosoma — (Figure 8A–B). Smaller than adults. Dorsum (Figure 8A): Subrectangular podonotal shield with lateral borders biconcave at level of z2 and z4, posterior region almost square; some line sculpture on dorsal shield surface; 11 pairs of setae, with j2 very close to j1; two pairs of pores near setae j3 and s5 on podonotal shield. Dorsum with two pairs of medial setae between the shields (J1–J2) and a lateral pair (J3). Mesonotal shield (or median diastema or platelets) comprising three pairs of platelets. Pygidial shield trapezoidal, as long as wide, with pairs of setae: J4–J5, Z3–Z5 and S5; J4 and Z4 much smaller than other setae (except J5, minute) on the pygidial shield; three pairs of pores near setae Z3, Z4 and J5 on pygidial shield. Unarmed dorsum has three pairs of free ''nodules'', two pairs large near to lateral edges and one pair very small on anterior region. Unarmed dorsum with 12 pairs of long setae, dorsal and ventral caudal setae may be present 1 or 2 minute barbs (clearly observed just under immersion). Venter (Figure 8B). Tritosternum as in adults. Sternal shield resembles a pentagon; three pairs of slender and long sternal setae; two pairs of lyrifissures posterior to st1 and st2. Unarmed venter with five pairs of setae, four pairs of median setae between ventral shields and a pair of caudal setae (without seta flanking the anus); setae pair between coxae IV are slender than other ventral setae. Anal shield subtriangular with postanal setae long and subequal; Peritreme short, extending from middle of coxae III to middle of coxae IV.
Legs — (Figure 9A–D). Chaetotaxy of the legs: I= 0 0/1 0/1 0 (2), 1 0/2 0/0 1 (4), 2 3/1 1/1 2 (10), 1 2/1 2/1 1 (8), 1 2/1 2/1 1 (8), 4 6/3 1/1 6/3 3 (27) + 6 solenidia+ 1 macroseta; II= 0 0/1 0/1 0 (2), 1 0/1 0/1 1 (4), 1 2/1 2/1 1 (8), 1 2/0 2/0 1 (6), 1 1/1 2/1 1 (7), 3 3/2 1/0 3/2 3 (17); III= 0 0/1 0/1 0 (2), 1 0/1 0/1 1 (4), 1 2/1 1/0 0 (5), 1 2/0 2/0 1 (6), 1 1/1 2/1 1 (7), 3 3/2 1/0 3/2 3 (17) and IV= 0 0/1 0/0 0 (1), 1 1/2 0/0 0 (4), 0 2/1 1/0 0 (4), 1 2/0 2/0 0 (5), 1 1/1 2/1 1 (7), 3 3/2 1/0 3/2 3 (17). Tarsus I sensory field has setae as in adults.
Gnathosoma — The gnathosoma, including chelicerae, similar to females, except the trochanter–tarsus palp chaetotaxy formula: 0/1 (1); 1 0/0 1/0 1/0 1 (4); 1 1/0 1/0 1/0 1 (5); 1 4/1 4/1 1 (12) and tarsus with 15 setae + forked apotele. Palpal trochanter lacks a ventral process.
Deutonymph and Larva — Unknown.
Host — Unknown
Etymology — The specific name ''cavernicola'' refers to its habitat.
Material studied — Holotype: Female [UFMG AC 171121], on 12-26/Aug./2013 at 18°56'10.6''S 43°24'45.5''W, in cave CSS-09, Conceição do Mato Dentro municipality. Paratypes: Female [UFMG AC 180094], on 26/Jun- 06/Jul./2018, at 19°20'35.0''S 43°18'28.8''W, in cave MCFC-0012, Itambé do Mato Dentro municipality; Female [UFMG AC AC171375], on 21/Nov. and 02/Dec./2016, in cave HOLC-0090/97, at 19°36'10.5''S 44°00'43.1''W, Pedro Leopoldo municipality; Female [UFMG AC 180044], on 26-28/Mar./2018, at 20°12'07.8''S 43°58'03.7''W, in cave MS-0030, Nova Lima municipality; Female [UFMG AC 170277], on 16-26/Jan./2017 at 19°09'58.7''S 43°15'59.3''W, in cave SPT-0180, Conceição do Mato Dentro municipality; Three female [UFMG AC 160229, 160327, 160214] and one male [UFMG AC 160322], on 10-27/Apr./2015, at 16°08'56.0''S 44°37'39.0''W, in cave Lapa Sem Fim, Luizlândia municipality; Female [UFMG AC 180121], on 26/Jun. and 06/Jul./2018, at 19°19'52.2''S 43°18'41.8''W, in cave MCFC-0009, Itambé do Mato Dentro municipality; Female [UFMG AC 200027], male [UFMG AC 180341] and protonymph [UFMG AC 200032], on 30/Apr. and 05/May/2018, at 18°55'08.0''S 43°25'13.6''W, in cave CSS-0007, Conceição do Mato Dentro municipality; Protonymph [UFMG AC 180049], on 26-28/Mar./2018, 20°12'08.7''S 43°58'03.4''W, in cave MS-0031, Nova Lima municipality. All localities in Minas Gerais state, Brazil. Coordinates given in WGS-84 All mites were collected freely dwelling on cave floor by the staff of the company Carste Ciência e Meio Ambiente. The additional material analyzed can be seen in the supplementary material 1.
Discussion — The new genus differs from Synasponyssus, Bewsiella Domrow 1958, Glauconyssus Uchikawa 1991, Ichoronyssus Kolenati 1858 and Parichoronyssus by bearing less than three pairs of pores on the sternal shield. These genera have the third pair of sternal pores on the shield or as in Synasponyssus on the complex sternal-median shield, included in the so-called group ''A'' by Radovsky (2010).
Chiasmanyssus cavernicola n. gen. n. sp. resembles all other genera of Macronyssidae by sharing the peritrematic shield posterior to the stigma fused to the parapodal shield around the posterior margin of coxa IV in adults, a character regarded as apomorphic in the family, but absent in several genera (Bewsiella, Glauconyssus, Ichoronyssus, Synasponyssus and Parichoronyssus) probably close to the laelapine stem (Radovsky 2010), and by the presence of an extra seta in the tibia III, genu III and IV.
Those setae are most likely lost in some derived genera: i) all extra seta absent from Chiroecetes; ii) only extra setae on genu III in Megistonyssus Radovsky 1966; iii) tibia III and genu IV both with one extra seta in Mitonyssoides and Acanthonyssus; iv) only genu IV with one extra seta in Macronyssus and Radfordiella (one species R. oudemansi Fonseca 1948); v) Pellonyssus, Atrichonyssus Radovsky 2010 and Ophionyssus have two extra setae in genu IV. Radovsky (2010) suggests that absence or presence of one or more extra setae on any or all tibia III, genu III and IV is useful for setting apart closely related genera within the family.
Chiasmanyssus n. gen. differs from all other genus described by the morphology of the sternal shield in females with its distinct ''X'' shaped area. Some genera have the sternal shield with some distinct sclerotization but not comparable to this: Chelanyssus Zumpt and Till 1953 and Steatonyssus have a sclerotized band in posterior margin (Till and Evans 1964; Till and Evans 1966; Radovsky 1966); Chiroptonyssus has a heavily sclerotized posterior band extending anteriad on the sides of the shield and Lagidonyssus has the shield divided into two parts, which anterior part shortest and a posterior part (from first pair of pores) more strongly sclerotized and with setae st2 and st3 (Fonseca 1935b; Radovsky 1967; 2010).
The presence of the seta J5 minute is similar to Macronyssus and Megistonyssus, among the most derived genera. However, it is distinguished from these genera by the absence of anterior spur of coxa II. In Macronyssus, the protonymphs have only 10 setae in shield podonotal, with j1 setae are anterior to the podonotal shield and in Megistonyssus the pygidial shield has only four setae pairs and a setae pair flanking the anal shield in the unarmed venter (Radovsky 1966; 1967; 2010).
Chiasmanyssus n. gen. is closely related to genus Cryptonyssus Radovsky 1966. It can be set apart, however, by only five pairs of setae on venter, four between the shields and one caudal (without setae flanking the anus); shape of pygidial and podonotal shield and presence of only 12 setae pairs on the dorsal integument. Furthermore, females lack marginal setae on dorsal shield, while males lack stout or spinose anterior setae on coxae II and III (Radovsky 1966; 1967).
Despite the scanty molecular data that could be retrieved from GenBank, the 16S fragment sequenced apparently is a good bar-coding marker, since it has a considerable gap between intra- and inter-specific distances. This fragment also supported Chiasmanyssus n. gen. as an independent lineage, highly supported in a clade recovered as sister-group of Ornithonyssus and Lepronyssoides.
Finally, it remains as a mystery the host of the new species. As the large number of individuals dwelling on caves floor, vertebrates that use them as nesting sites or shelter are candidates. However, authors have been surveying bats for several months, the most abundant vertebrates found in those caves and, despite finding most of mesostigmata lineages usually associated to bats in neotropics, could not find Chiasmanyssus n. gen. on them thus far.
Keys for genus of Macronyssidae from Brazil
Females
1. Peritrematic shield posterior to the stigma not fused with the parapodal shield around the posterior margin of coxa IV; three pairs of pores on sternal shield; setae z3 on dorsum; setae J5 absent; dorsal shield with lateral and posterior portions darkened and submerged beneath striated cuticula; cinctures absent in chelae
...... Parichoronyssus
— Peritrematic shield posterior to the stigma fused to the parapodal shield around the posterior margin of coxa IV; less than three pores pairs on sternal shield; setae z3 absent; setae J5 present or absent; dorsal shield without portions submerged; cinctures may be present in chelae
...... (2)
2. Genu III with 9 setae; cinctures may be present on chelae
...... (3)
— Genu III with 10 setae; cinctures absent on chelae
...... (5)
3. Ventral process of palpal trochanter absent; cinctures absent on chelae; coxa II without anterior spur; tibia III with nine setae
...... Mitonyssoides
— Ventral process of palpal trochanter present; cinctures present on chelae; coxa II with anterior spur; tibia III with less than nine setae
...... (4)
4. Setae J5 absent; sternal glands absent on sternal shield; accessory setae absent on epigynal shield; ventral shields lack sculpturing
...... Radfordiella
— Setae J5 present and minute; sternal glands present, lateral to the first pair of pores and the glands are variable in structure on shield; accessory setae present or absent on epigynal shield; ventral shields with some distinct sculpturing
...... Macronyssus
5. Epigynal shield flanked by the genital setae inserted on the unarmed integument; chelicera basally swollen
...... Ophionyssus
— Epigynal shield with genital setae inserted on shield; chelicera basally not as above
...... (6)
6. Some or most dorsal and caudal setae of idiosoma barbed
...... (7)
— Barbed setae absent on idiosoma (or at most 1–2 pairs of caudal setae notched or feathered)
...... (10)
7. Ventral process of palpal trochanter absent; sternal shield with less than 3 pairs of setae
...... Thigmonyssus
— Ventral process of palpal trochanter present; sternal shield with 3 pairs of setae
...... (8)
8. Setae J3 present; J5 usually present (absent in C. cubensis Dusbábek 1969)]; some caudal setae are peg-like and with tip multiply barbed; setae on the legs smooth; sternal shield with a heavily sclerotized posterior band extending anteriad on the sides of shield; claw I much smaller than the others claws; leg II stout than the others legs
...... Chiroptonyssus
— Setae J3 and J5 absent; caudal setae are not peg-like and with single barbed tip; setae on the legs smooth or barbed; sternal shield lacking a sclerotized band; subequal claws and legs in width
...... (9)
9. Setae j3 present; all setae on legs smooth; sternal shield with smooth oval structures posterior to st1 and posteromedian to the first pair of pores; always three pairs of setae on sternal shield; coxa III with a posterior triangular, round-tipped spur
...... Lepronyssoides
— Setae j3 absent; usually legs with barbed setae, but a single barb; sternal shield lacking ovals structure; usually three pairs of setae on sternal shield, rarely st3 are off the shield (some O. sylviarum and O. hypertrichus Radovsky 2007); coxa III with a setiform posterior seta
...... Ornithonyssus
10. Dorsal shield divided
...... (11)
— Dorsal shield entire
...... (12)
11. Setae j3 and J5 absent; 1–2 pores pairs on sternal shield; sternal shield lacking sclerotized band; genu IV with two extra setae (11 setae)
...... Pellonyssus
— Setae j3 usually present (absent in e.g. S. evansi Delfinado 1960, S. benoiti Till and Evans 1964), often much reduced; J5 usually present, rarely absent; two pores pairs on sternal shield; sternal shield with a sclerotized band in posterior margin; genu IV with one extra setae (10 setae)
...... Steatonyssus
12. Setae J5 absent; dorsal shield always non-neotrichous; some coxae with ridges; claws subequal
...... Macronyssoides
— Setae J5 present; dorsal shield may be neotrichous; coxae without ridges; claws may be unequal
...... (13)
13. Dorsal shield neotrichous; setae J5 small, subequal to Z5; most 1–2 pairs of caudal setae notched or feathered; sternal shield lacking distinct sclerotization; without an anterior protusion on epigynal shield; claw I reduced relative to other claws
...... Mitonyssus
— Dorsal shield non-neotrichous; setae J5 minute; none of caudal setae notched or feathered; sternal shield with distinct X-shaped area; anterior edge of epigynal shield with a protusion; claws subequal
...... Chiasmanyssus n. gen.
Males
1. Peritrematic shield posterior to the stigma not fused, without the parapodal shield around the posterior margin of coxa IV; tarsus I with 2–4 strongly pedicellate distal setae; setae J5 absent; tibia III, genu III and IV without extra setae
...... Parichoronyssus
— Peritrematic shield posterior to the stigma fused, with the parapodal shield around the posterior margin of coxa IV; tarsus I with 0–1 pedicellate distal setae setae J5 present or absent; some tibia III, genu III and IV with extra setae
...... (2)
2. Genu III with 9 setae
...... (3)
— Genu III with 10 setae
...... (5)
3. Setae st1 arise in the presternal area; neotrichous on dorsal shield
...... Mitonyssoides
— Setae st1 arise in the esternal shield; non-neotrichous on dorsal shield
...... (4)
4. Setae pair J5 present and minute; ventral process of palpal trochanter with a strong longitudinal ridge; dorsal and/or caudal margin may bear some setae long and flagelliform; fixed chela with a tiny pilus dentilis; leg I slender than or equal to the other legs; all claws small; coxa II with anterior spur entire (except M. contrictus with bifid tip)
...... Macronyssus
— Setae pair J5 absent; without ventral process of palpal trochanter or at most with a line showing its position; dorsal and/or caudal margin never flagelliform; fixed chela without a tiny pilus dentilis; leg I and II stout but leg II may be slightly to much stronger than the other legs; all claws are strong; coxa II with anterior spur bifid or 2 subequal spur
...... Radfordiella
5. Some setae barbed on idiosoma at least caudally with 1–3 barbed setae
...... (6)
— All setae smooth (or 1–2 pairs of caudal setae notched or feathered) on idiosoma
...... (9)
6. Setae j1 and j2 off dorsal shield
...... Thigmonyssus
— Setae j1 and j2 on dorsal shield
...... (7)
7. Palpal femur without enlarged lateral setae and one pedicel; J5 usually present (absent C. cubensis, small); venter usually has sternogenital and ventrianal shields or rarely holoventral shield
...... Chiroptonyssus
— Palpal femur with spinose seta and on a lateral pedicel; J5 absent; venter always has a holoventral shield
...... (8)
8. Setae j3 absent; setae in the opisthonotal region, not including neotrichae behind J2; 1 or 2 enlarged lateral setae on palp femur, of which the anterior one on a prominent pedicel; all legs with some dorsal setae barbed
...... Ornithonyssus
— Setae j3 present; setae in the opisthonotal region, including about 3–4 neotrichae behind J2; palpal femur has only 1 enlarged lateral seta on pedicel; legs setae lack barbs
...... Lepronyssoides
9. Venter with sternogenital and anal shields, the ventral region unarmed; genital setae inserted on the unarmed integument flanking the sternogenital shield; Chelicera is basally swollen as in the female
...... Ophionyssus
— Venter with a holoventral shield or sternogenital and ventrianal shields; genital setae on the shield; Chelicera basally not as above
...... (10)
10. 1–2 pairs of caudal setae notched or feathered; claws I greatly reduced relative to other claws
...... Mitonyssus
— None setae as above; claws I not reduced relative to other claws
...... (11)
11. Genu IV with 2 extra setae (11 setae)
...... Pellonyssus
— Genu IV with 1 extra seta (10 setae)
...... (12)
12. Setae J5 absent; some setae on dorsal shield with expanded tip
...... Macronyssoides
— Setae J5 present or absent; none setae on dorsal shield with expanded tip
...... (13)
13. Setae J5 minute; j3 subequal to other setae on shield; anterior spur in coxa II absent; coxa III has an anterior seta setiform
...... Chiasmanyssus n. gen.
— Setae J5 when present small, not minute; setae j3 very reduced or absent; anterior spur in coxa II present with tip divided into several denticles; coxa III has an anterior seta ranging from normal setiform to regular spinose to spinose with a blade-like proximal expansion
...... Steatonyssus
Protonymphs
1. Setae j1 anterior to podonotal shield or may be on margin of podonotal shield
...... (2)
— Setae j1 on podonotal shield
...... (4)
2. Setae j2 off podonotal shield
...... Thigmonyssus
— Setae j2 on podonotal shield
...... (3)
3. Podonotal shield with 10 setae pairs; setae j3 present; J5 present and minute; pygidial shield with 6–7 setae pairs; 2 median setae between dorsal shield; normally 4 setae pairs between ventral shields but may be more than 10 setae pairs when neotrichous
...... Macronyssus
— Podonotal shield with 8–9 setal pairs; setae j3 absent; J5 absent; pygidial shield with 1–3 setae pairs; 3 median setae between dorsal shield; 2–3 setae pairs between ventral shields, never neotrichous
...... Pellonyssus
4. Usually no more than two median setal pairs between dorsal shields, a posterior pair (J3) may be present in integument, in which case it is shifted laterally
...... (5)
— About 3–8 or more median setal pairs between dorsal shields, setae J3 pair when present on integument, not laterally shifted
...... (9)
5. Chelicera with a swollen base; on tarsus I, seta s2 small much less than half the length of seta s1
...... Ophionyssus
— Chelicera with an approximately uniform base in width throughout its length; seta s2 on tarsus I not as above
...... (6)
6. Setae J5 present and minute; pygidial shield with six setae pairs
...... Chiasmanyssus n. gen.
— Setae J5 absent; pygidial shield no more than four setae pairs
...... (7)
7. Pygidial shield with two setae pairs; sensory field of tarsus 1 with 2–4 pedicillate distal setae; genital setae pair absent; two pairs flanking the anal shield; none extra seta on tibia III, genu III and IV
...... Parichoronyssus
— Pygidial shield with 3–4 setae pairs; sensory field of tarsus 1 with 0–1 pedicillate distal setae; genital setae pair present; 0–1 pairs flanking the anal shield; all tibia III, genu III and IV with one extra seta or at least genu IV with an extra seta
...... (8)
8. Setae j3 present, may be reduced but always represented by the trichopores; pygidial shield with a pair of enlarged anterolateral pores; pygidial shield normal or much reduced; none extra seta in tibia III and genu III but genu IV may be with one extra seta; claw I not larger than other legs; peritreme lacking a post-stigmal part
...... Radfordiella
— Setae j3 present and subequal other shield setae; pygidial shield lacking enlarged pores; pygidial shield always normal; one extra seta on tibia III, genu III and IV; claw I larger than those of the other legs; peritreme with a post-stigmal part
...... Macronyssoides
9. Three median setal pairs between dorsal shields (non-neotrichous)
...... (10)
— Four or more median setal pairs between dorsal shields (neotrichous)
...... (12)
10. All setae smooth on idiosoma
...... Steatonyssus
— Barbed setae on idiosoma, usually single barb
...... (11)
11. Podonotal shield with 10 pairs of setae; j3 absent; Z4 not reduced; all legs with some barbed setae
...... Ornithonyssus
— Podonotal shield with 11 pairs of setae; j3 present; Z4 reduced; all legs with smooth setae
...... Lepronyssoides
12. Venter with only 3 pairs of setae between the ventral shield
...... Mitonyssoides
— Venter with more than 3 pairs of setae between the ventral shields
...... (13)
13. Venter neotrichous with about 5 or 6 pairs of setae between shields (except C. venezolanus); some caudal setae on the pygidial shield peg-like and/or with short barbs; leg II much stouter than other legs; claws II much larger than other claws and claws I as claws III and IV
...... Chiroptonyssus
— Venter neotrichous with about 8–11 pairs of setae between shields; some caudal setae on the pygidial shield; leg II with normal thickness as the other legs; claws II subequal other claws but claws I much reduced or absent
...... Mitonyssus
Acknowledgements
We thank Pedro H. da Silva Conceição for preparing the slides and Dr. Leopoldo Ferreira de Oliveira Bernardi for taxonomic assistance and correction of the first draft of this manuscript. We also thank Dr. Juan B. Morales-Malacara for critical reading of the draft of this manuscript. Authors are in debt to anonymous referees and Dr. Farid Faraji (University of Amsterdam) for their invaluable suggestions and corrections The authors also thank the Program for Technological Development in Tools for Health-PDTISFIOCRUZ for use of its facilities, especially to Renata de B. R. Oliveira who assisted with sequencing and the company Carste Ciência e Meio Ambiente for depositing the specimens at UFMG AC. This study was supported by resources from FAPEMIG-VALE (Edital 07/2018-Research in Speleology). This study is part of the first author's PhD thesis in the post-graduation Program in Zoology-UFMG.
References
- Almeida J.C., Silva S.S.P., Serra-Freire N.M., Valim M.P. 2011. Ectoparasites (Insecta and Acari) associated with bats in Southeastern Brazil. J. Med. Entomol., 48:753-757. doi:10.1603/ME09133
- Augustson G.F. 1945. A new genus, new species of Dermanyssid mite (Acarina) from Texas. Bull. South Calif. Acad. Sci., 44(2): 46-48.
- Azevedo A.A., Linardi P.M., Coutinho M.T.Z. 2002. Acari ectoparasites of bats from Minas Gerais, Brazil. J. Med. Entomol., 39: 553-555. doi:10.1603/0022-2585-39.3.553
- Bassini-Silva R, Jacinavicius F.C., Huang-Bastos M., Dowling A.P., Barros Battesti D.M. 2020. A Checklist of Macronyssid Species (Mesostigmata: Macronyssidae) From Brazil, J. Med. Entomol. ,58(2): 625-633. doi:10.1093/jme/tjaa245
- Bassini-Silva R, Jacinavicius F.C., Joppert A.M., Dowling A.P., Barros Battesti D.M. 2019. New host association and locality record for Pellonyssus gorgasi Yunker and Radovsky, 1966 (Mesostigmata: macronyssidae) in the state of São Paulo, Brazil. Int. J. Acarology, 45: 235-237. doi:10.1080/01647954.2019.1604801
- Berlese A. 1888. Acari Austro-Americani quos collegit Alysius Balzam. Manipulus primus. Boll. Soc. Entomol. Ital., 20: 171-222.
- Canestrini G., Fanzago F. 1877. Intorno agli Acari Italiani. Atti del R. Inst. Veneto Sci. Lett. Arti. Ser., 5(4): 124-125.
- Clark G.M., Yunker C.E. 1956. A new genus and species of Dermanyssidae (Acarina: Mesostigmata) from the English sparrow, with observations on its life cycle. In Proc. Helminthol. Soc. Washington, 23: 93-101.
- Darriba D., Taboada G.L., Doallo R., Posada D. 2012. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods, 9(8): 772. doi:10.1038/nmeth.2109
- Domrow R. 1958. New and little known Australian Laelaptidae (Acarina). Proc. Linn. Soc. New South Wales, 82: 352-366.
- Edgar R. 2004. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res., 32(5): 1792-1797. doi:10.1093/nar/gkh340
- Evans G.O., Till W.M. 1979. Mesostigmatic mites of Britain and Ireland (Chelicerata: Acari: Parasitiformes). An introduction of their external morphology and classification. Tran. Zool. Soc. Lond., 35: 139-270. doi:10.1111/j.1096-3642.1979.tb00059.x
- Fonseca F. 1935a. Notas de Acarologia XIII. Novas espécies sul-americanas de parasitos do gênero Liponissus Kolenati. 1958 (Acarina: Liponissidae). Mem. Inst. Butantan, 9: 45-90.
- Fonseca F. 1935b. Notas de acarologia. XXII. Liponissus haematophagus, sp. n. (Acari. Liponissidae). Mem. Inst. Butantan, 10: 25-28.
- Fonseca F. 1939. Notas de Acarologia XXVII. Liponissus brasiliensis sp. n., parasita habitual de roedores e acidental do homem. Mem. Inst. Butantan, 12: 1-8.
- Fonseca F. 1941. Notas de Acarologia XXXII. Novas espécies brasileiras do gênero Liponissus Kolenati (Acari: Liponissidae). Mem. Inst. Butantan, 15: 103-118.
- Fonseca F. 1948. Monograph of the general and species of Macronyssidae Oudemans, 1936 (synom.: Liponyssidae Vitzthum, 1931) (Acari). Proc. Zool. Soc. London, 118: 249-334. doi:10.1111/j.1096-3642.1948.tb00378.x
- Fonseca F. 1954. Notas de Acarologia XXXVI. Aquisições novas para a fauna brasileira de ácaros hematófagos (Acari: Macronyssidae). Mem. Inst. Butantan, 1: 79-92.
- Fonseca F. 1960. Notes d'acarologie. XLV. Enquête acarologique a Pérou. Acarologia Tome II (I), 2: 1-34.
- Gillespie J.J., Johnston J.S., Cannone J.J., Gutell R.R. 2006. Characteristics of the nuclear (18S, 5.8S, 28S and 5S) and mitochondrial (12S and 16S) rRNA genes of Apis mellifera (Insecta: Hymenoptera): structure, organization, and retrotransposable elements. Insect Mol. Biol., 15(5): 657-686. doi:10.1111/j.1365-2583.2006.00689.x
- Guindon S., Dufayard J.F., Lefort V., Anisimova M., Hordijk W., Gascuel O. 2010. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol., 59(3): 307-321. doi:10.1093/sysbio/syq010
- Hall T.A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser., 41: 95-8.
- Herrin C.S., Radovsky F.J. 1974. Venezuelan Macronyssidae. 3.Chiroecetes lonchophylla, n.g., n.sp., from a long-tongued bat. J. Med. Entomol., 11: 347-351. doi:10.1093/jmedent/11.3.347
- Hirst S. 1913. On three new species of gamasid mites found on rats. Bull. Entomol. Res., 4(2): 119-124. doi:10.1017/S0007485300043029
- Kolenati F.A. 1858. Synopsis prodroma der auf Chiroptern als epizoen vorkommenden Lausmilben, Carida Kolenati. Wiener Entomol. Monatschr., 2: 4-7.
- Krita (2020) Krita Foundation. https://krita.org/en/
 .
. - Kumar S, Stecher G., Tamura. K. 2016. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol., 33(7): 1870-1874. doi:10.1093/molbev/msw054
- Mangold A.J., Bargues M.D. Mas-Coma S. 1998. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res., 84(6): 478-484. doi:10.1007/s004360050433
- Megnin P. 1884. Étude sur I'Ophionyssus natricis P. Gervais. Bull. Soc. Zool. France, 9: 107.
- Micherdzinski W. 1980. Eine Taxonomische analyse der Familie Macronyssidae Oudemans. 1936. I. Subfamilie Ornithonyssinae Lange. 1958 (Acarina: Mesostigmata). Państwowe Wydawn. Naukowe, Warsaw, 264 pp.
- Moras L.M., De Oliveira Bernardi L.F., Graciolli G., Gregorin R. 2013. Bat flies (Diptera: Streblidae, Nycteribiidae) and mites (Acari) associated with bats (Mammalia: Chiroptera) in a high-altitude region in southern Minas Gerais, Brazil. Acta Parasitol., 58(4): 556-63. doi:10.2478/s11686-013-0179-x
- Nieri-Bastos F.A, Labruna M.B., Marcili A., Durden L. A., Mendoza-Uribe L., Barros-Battesti D.M. 2011. Morphological and molecular analysis of Ornithonyssus spp. (Acari: Macronyssidae) from small terrestrial mammals in Brazil. Exp. Appl. Acarol., 55(4): 305. doi:10.1007/s10493-011-9475-z
- Oliveira G.M.B., Bassini-Silva R., Huang-Bastos M., Pereira J.S., Sponchiado J., Guimarães M.F., Dowling A.P.G., Labruna M.B., Barros-Battesti D.M., Jacinavicius F.C., Horta, M.C. 2020. Contribution about the knowledge of Lepronyssoides pereirai (Fonseca, 1935) (Mesostigmata: Macronyssidae): hosts and distribution. Int. J. Acarology, 46(5): 377-379. doi:10.1080/01647954.2020.1808060
- Orlova M.V., Kazakov D.V., Orlov O.L., Mishchenko V.A., Zhigalin A.V. 2017. The first data on the infestation of the parti-coloured bat, Vespertilio murinus (Chiroptera, Vespertilionidae), with gamasid mites, Steatonyssus spinosus (Mesostigmata, Gamasina, Macronyssidae). Russ. J. Theriol., 16: 66-73. doi:10.15298/rusjtheriol.16.1.06
- Oudemans A.C. 1936. Kritisch Historisch Oversicht der Acarologia. Leiden. 3A: 430pp.
- QGIS development team (org.) (2021) QGIS Geographic Information System. QGIS Association. http://www.qgis.org
- Radovsky F.J. 1966. Revision of the macronyssid and laelapid mites of bats: outline of classification with descriptions of new genera and new type species. J. Med. Entomol., 3(1): 93-99. doi:10.1093/jmedent/3.1.93
- Radovsky F.J. 1967. The Macronyssidae and Laelapidae (Acarina, Mesostigmata) parasitic on bats. Univ. Calif. Publ. Entomol., 46: 1-288.
- Radovsky F.J. 1969. Adaptive radiation in the parasitic Mesostigmata. Acarologia II: 450-483.
- Radovsky F.J. 2010. Revision of Genera of the Parasitic Mite Family Macronyssidae:(Mesostigmata Dermanyssoidea) of the World. West Bloomfield: Indira Publishing House, 288 pp.
- Radovsky F.J., Furman D.P. 1969. An unusual new genus and species of Macronyssidae (Acarina) parasitic on a disc-winged bat. J. Med. Entomol., 6(4): 385-393. doi:10.1093/jmedent/6.4.385
- 10.1093/jmedent/6.4.385 doi:10.1093/jmedent/6.4.385
- Ronquist F., Teslenko M., Van Der Mark P., Ayres D.L., Darling A., Höhna S., Larget B., Liu L., Suchard M.A., Huelsenbeck J.P. 2012. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol., 61(3): 539-542. doi:10.1093/sysbio/sys029
- Sambon L.W. 1928. The parasitic acarians of animals and the part they play in the causation of the eruptive fevers and other diseases of man: Preliminary considerations based upon an ecological study of typhus fever. Ann. Trop. Med. Parasit., 22(1): 67-132. doi:10.1080/00034983.1928.11684565
- Saunders R.C., Yunker C.E. 1973. Venezuelan Macronyssidae. II. Nycteronyssus desmodus, n. gen., n. sp., off a vampire bat. J. Med. Entomol., 10(4): 381-384. doi:10.1093/jmedent/10.4.381
- Saunders R.C., Yunker C.E. 1975. Venezuelan Macronyssidae IV. A new genus and species of rodent-parasitizing Ornithonyssinae (Acarina, Macronyssidae). J. Med. Entomol., 11(6): 756-759. doi:10.1093/jmedent/11.6.756
- Shepherd A.J., Narro S. P. 1983. The genus Ornithonyssus Sambon 1928 in the Ethiopian region: description of a new species and a redescription of O. voseinnesi (Zumpt and Till. 1953) (Acarina. Mesostigmata). Acarologia, 24(4): 347-353.
- Sidorchuk E.A., Vorontsov D.D. 2014. Computer-aided drawing system-Substitute for camera lucida. Acarologia, 54(2): 229-239. doi:10.1051/acarologia/20142130
- Silva C.D.L., Valim M.P., Graciolli G. 2017. Ácaros ectoparasitos de morcegos no estado de Mato Grosso do Sul, Brasil. Iheringia Ser. Zool. 107 (n. suppl.). ISSN 0073-4721. doi:10.1590/S0073-47212017000200211
- Smith S. de W., Bond J.E. 2003. An Analysis of the Secondary Structure of the Mitochondrial Large Subunit rRNA Gene (16S) in Spiders and Its Implications for Phylogenetic Reconstruction. J. Arachnol., 31: 44-54. doi:10.1636/0161-8202(2003)031[0044:AAOTSS]2.0.CO;2
- Till W. M., Evans G.O. 1964. The genus Steatonyssus Kolenati (Acari: Mesostigmata). Bull. Brit. Mus. (Nat. Hist.), Zool. 11(8): 511-582. doi:10.5962/bhl.part.4724
- Till W.M., Evans G.O. 1966. The genus Chelanyssus Zumpt and Till (Acari: Mesostigmata). Acarologia, 8(3): 401-414.
- Uchikawa K. 1991. Glauconyssus: a new genus associated with African bats (Acari: Macronyssidae). J. Med. Entomol., 28: 446-449. doi:10.1093/jmedent/28.3.446
- Whitaker Jr. J.O., Mumford R.E. 1977. Records of ectoparasites from Brazilian mammals. Entomol. News, 88(9-10): 255-258.
- Yunker C.E., Lukoschus F.S., Giesen K.M.T. 1990. Parasitic mites of Surinam: 24. The subfamily Ornithonyssinae, with descriptions of a new genus and three new species (Acari: Mesostigmata: Macronyssidae). Zool. Meded., 63(14): 169-186.
- Yunker C.E., Radovsky F.J. 1966. The Dermanyssid mites of Panama (Acarina, Dermanyssidae). In: Wenzel R.L., Tipton V.J. (Eds.). Ectoparasites of Panama. Field Museum of Natural History, Chicago, 88-103.
- Yunker C.E., Radovsky F.J. 1980. Parasitic mites of Surinam. XXXVI. A new genus and two new species of Neotropical Macronyssidae (Acari: Mesostigmata). J. Med. Entomol., 17(6): 545-554. doi:10.1093/jmedent/17.6.545
- Yunker C.E., Saunders R.C. 1973. Venezuelan Macronyssidae. I. The genera Acanthonyssus Yunker & Radovsky, and Argitis, n. gen. J. Med. Entomol., 10(4): 371-381. doi:10.1093/jmedent/10.4.371
- Zumpt F., Till W.M. 1953. The genus Liponyssus Kolenati in the Ethiopian region (Acarina: Laelaptidae) S. Afr. J. Med. Sci., 18: 5-12.



2021-02-08
Date accepted:
2021-05-18
Date published:
2021-05-27
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2021 Gomes-Almeida, Brenda K. and Pepato, Almir R.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)