



A new species of Amblydromalus Chant & McMurtry (Acari: Phytoseiidae), with notes on occurrence of genus in South America
Demite, Peterson Rodrigo1 ; Rezende, José Marcos2 ; Carvalho Dahmer, Priscila3 ; Cavalcante, Ana Cristina C.4 and Lofego, Antonio Carlos5
1✉ Instituto de Biociências, Universidade Federal de Mato Grosso, Cuiabá, Mato Grosso, Brazil.
2Departamento de Zoologia e Botânica, Instituto de Biociências, Letras e Ciências Exatas, Universidade Estadual Paulista, UNESP, São José do Rio Preto, São Paulo, Brazil.
3Instituto de Ciências Exatas e Tecnologia, Universidade Federal do Amazonas, UFAM, Itacoatiara, Amazonas, Brazil.
4Instituto de Ciências Exatas e Tecnologia, Universidade Federal do Amazonas, UFAM, Itacoatiara, Amazonas, Brazil.
5Departamento de Zoologia e Botânica, Instituto de Biociências, Letras e Ciências Exatas, Universidade Estadual Paulista, UNESP, São José do Rio Preto, São Paulo, Brazil.
2021 - Volume: 61 Issue: 3 pages: 527-537
https://doi.org/10.24349/acarologia/20214445ZooBank LSID: 0C03B33C-20A4-47E1-AB84-509B598E10A2
Original research
Keywords
Abstract
Introduction
The genus Amblydromalus Chant & McMurtry (Acari: Phytoseiidae) belongs to the tribe Euseiini Chant & McMurtry, subtribe Typhlodromalina Chant & McMurtry, with 23 valid species, the majority recorded in the Neotropical Region (Chant and McMurtry 2005, 2007, Demite et al. 2021). To date, all species of this genus are characterized by having 33 pairs of setae, with female idiosomal setal pattern 10A:9B/JV-3:ZV, the most common for Amblyseiinae (Chant and McMurtry 2005). Twelve species have been recorded from Brazil, being six of them recorded from the Brazilian Amazon region, namely Amblydromalus akiri Nuvoloni, Lofego, Rezende & Feres, A. insolitus Nuvoloni & Lofego, A. itacoatiarensis Demite, Cavalcante & Lofego, A. manihoti (Moraes), A. villacarmelensis (Moraes) and A. zannoui Famah Sourassou, Sarmento & Moraes (Moraes et al. 1994, Navia et al. 2005, Nuvoloni et al. 2015a, b, Vasconcelos and Silva 2015, Cruz et al. 2019, Demite et al. 2019). Additionally, Gondim Jr. et al. (2012) identified two morphospecies (Amblydromalus aff. arawak and Amblydromalus sp.) in the state of Roraima, however, their observation based on a few specimens.
This study aims to describe a new species of the genus Amblydromalus, the first of its kind which presents J1 setae, associated with plants in the states of Amazonas and Mato Grosso, Brazil.
Material and methods
Mites were collected from leaf samples from six native plants and one exotic fruit (rambutan) in two states from Brazil (Amazonas and Mato Grosso). In the state of Amazonas, the samplings were carried out in municipalities of Itacoatiara, Maués, Parintins, and Silves (east region of the state). Natural vegetation and residential area were sampled in the first three municipalities and in the last, respectively. In the state Mato Grosso, the samplings were carried out in Cotriguaçu municipality (north region of the state) in natural vegetation. All areas are within the domain of the Amazon Forest biome.
The phytoseiid specimens were sorted under a stereomicroscope (40x) and subsequently mounted on microscope slides in Hoyer's medium. They were examined under phase-contrast microscopy (Zeiss Axio Imager M3). The new species was illustrated using a drawing tube attached to the microscope; the images were processed with the software Adobe Illustrator CS6. Measurements of taxonomically relevant structures were done with the use of a graded eye-piece. The measurements of the holotype (female) are given in bold, followed by the average measurement and then by minimum and maximum values (in parentheses) for the holotype and paratypes.
The setal nomenclature adopted was that of Lindquist and Evans (1965) and Lindquist (1994), as adapted by Rowell et al. (1978) for the dorsum and by Chant and Yoshida-Shaul (1991) for the venter of the phytoseiids. The idiosomal setal pattern follows Chant and Yoshida-Shaul (1992). The notation of gland pores (solenostomes) or lyrifissures (poroids) is based on Athias-Henriot (1975). Terminology for the spermathecal apparatus follows that described by Beard (2001).
The analyses on A. akiri were done studying specimens of the type series, deposited in the collection of Acari of the Departmento de Zoologia e Botânica, da UNESP, Campus de São José do Rio Preto, São Paulo, Brazil.
The studies that report the occurrence of Amblydromalus species in South America (previous records) were obtained from the Phytoseiidae Database (Demite et al. 2021; http://www.lea.esalq.usp.br/phytoseiidae/ ![]() ). Records of species of the genus that contained ''cf.'' or ''af.'' were not considered for this study. The records of A. horatii (De Leon) in Brazil, Guyana, and Suriname by Byrne et al. (1983) and Murphy (1984) were considered as Amblydromalus arawak (De Leon), since that this species is a junior synonym of A. arawak (Denmark et al. 1999).
). Records of species of the genus that contained ''cf.'' or ''af.'' were not considered for this study. The records of A. horatii (De Leon) in Brazil, Guyana, and Suriname by Byrne et al. (1983) and Murphy (1984) were considered as Amblydromalus arawak (De Leon), since that this species is a junior synonym of A. arawak (Denmark et al. 1999).
Sytematics
Amblydromalus amazonicus n. sp. Demite, Rezende & Lofego
ZOOBANK: 6F1500CC-5935-41BA-A5F3-AA18B70EFD0F ![]()
(Figure 1)
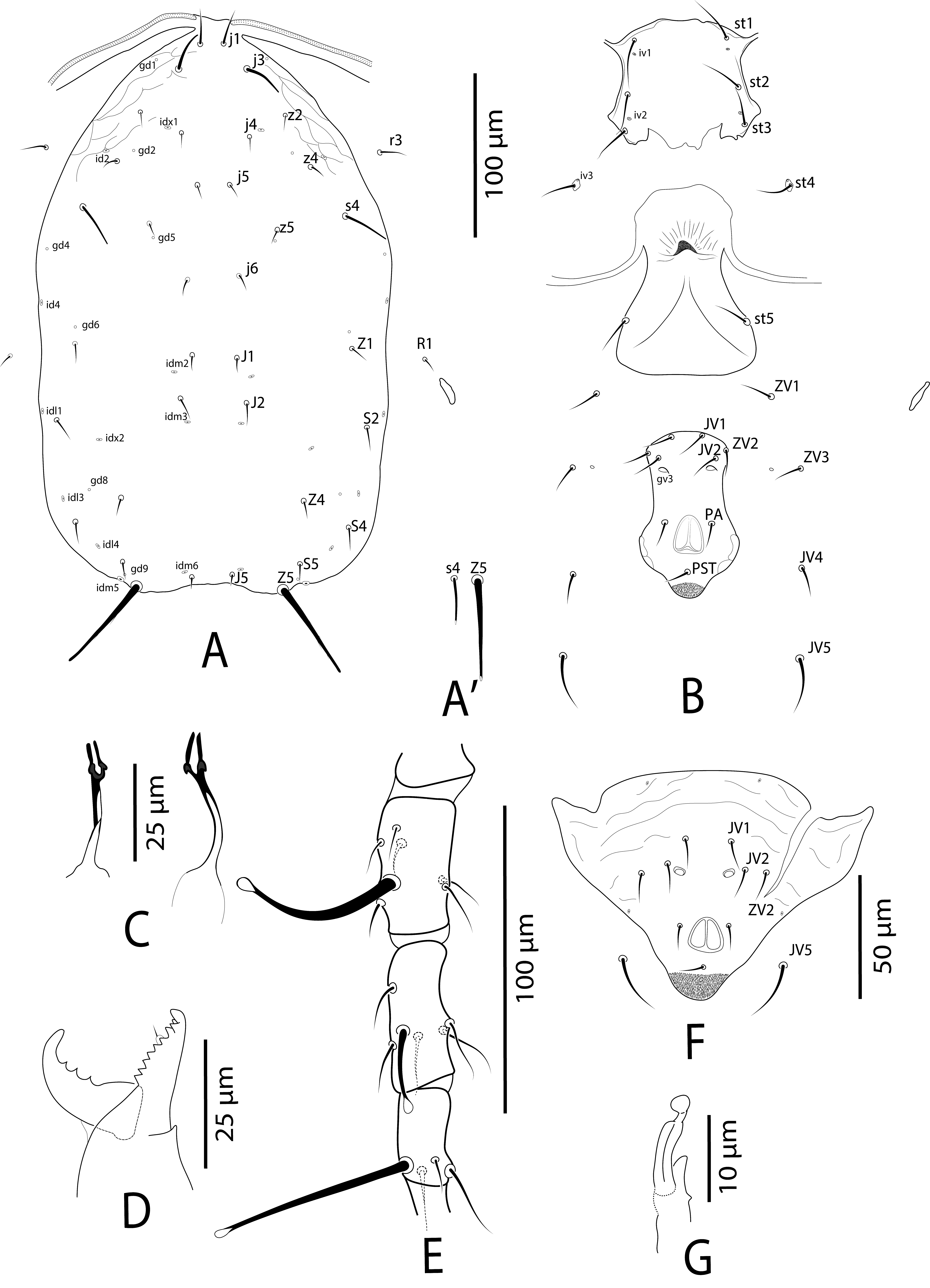
Diagnosis
Female with dorsal shield smooth, only with some striae, mainly in anterolateral region; all dorsal setae smooth and pointed, except Z5 slightly serrated; with seta J1 present. Sternal shield smooth, except lateral striae, with posterior margin trilobate; genital and ventrianal shields smooth; ventrianal shield vase-shaped, with a pair of crescentic preanal pores; all ventral setae smooth. Calyx tubular; atrium widening next to major duct and with forked aspect; major duct with thick wall next to the atrium. Macrosetae smooth with knobbed tips, present on genua of all legs, tibiae of legs III and IV and basitarsus of leg IV.
Female (n= 14)
Dorsum — (Figure 1A). Idiosomal setal pattern 10A: 10B/JV-3:ZV. Dorsal shield mostly smooth, only with some striae, mainly in the anterolateral region; 320 302 (280–323) long from anterior to posterior margins along midline, 195 198 (190–210) wide at level of s4; with 18 pairs of setae on dorsal shield; setae r3 and R1 inserted in the unsclerotized cuticle; with 11 pairs of lyrifissures and seven pairs of gland pores; setae lengths: j1 20 20 (17–23), j3 25 23 (21–26), j4 10 9 (7–10), j5 9 8 (7–9), j6 10 9 (8–10), J1 11 10 (8–11), J2 12 10 (9–12), J5 7 7 (5–7), z2 12 10 (9–12), z4 10 10 (8–11), z5 9 8 (6–9), Z1 12 11 (10–12), Z4 11 10 (9–12), Z5 60 57 (53–62), s4 26 22 (20–26), S2 14 13 (10–15), S4 14 12 (10–14), S5 10 10 (9–10), r3 15 13 (11–15), R1 10 9 (8–10). All dorsal setae smooth, except Z5 slightly serrated; in some specimens Z5 and s4 with tips ending in a small membrane (Figure 1A').
Venter — (Figure 1B). Sternal shield smooth, except lateral striae, with three pairs of setae and two pairs of lyrifissures (iv1 and iv2); distances between st1–st3 52 52 (49–60), st2–st2 64 63 (59–68); st4 and one lyrifissure (iv3) on metasternal plate. Genital shield smooth, distance between st5–st5 70 65 (59–73). With two pairs of metapodal plates. Ventrianal shield vase-shaped, smooth; 98 91 (83–98) long, 50 50 (45–54) wide at level of ZV2 and 60 57 (52–64) at level of anus; with three pairs of pre-anal setae (JV1, JV2 and ZV2) and a pair of crescentic pores (gv3) posterior to JV2. Four pairs of opisthogastric setae on unsclerotized cuticle (JV4, JV5, ZV1 and ZV3). Seta JV5 40 35 (30–40). All ventral setae smooth.
Peritreme — Extending to level of j1.
Spermatheca — (Figure 1C). Calyx tubular, 27 23 (18–27) (for some mounted specimens the calyx is visually bent); atrium widening next to major duct and with forked aspect; major duct with thick wall next to atrium.
Chelicera — (Figure 1D). Fixed digit 26 26 (23–30) long, with 9 9 (8–10) teeth in addition to apical hook and pilus dentilis; movable digit, 28 24 (23–28), with 3 4 (3–5) teeth in addition to apical hook.
Legs — (Figure 1E). Macrosetae smooth with knobbed tips; lengths: SgeI 27 27 (25–28), SgeII 27 27 (25–30), SgeIII 36 36 (34–39), StiIII 23 23 (21–25), SgeIV 53 53 (51–55), StiIV 27 28 (24–30), StIV 63 61 (55–63). Chaetotactic formula of genu II: 2-2/0-2/0-1 and genu III: 1-2/1-2/0-1.
Male (n= 4)
Dorsum — Dorsal shield ornamentation as in female, 224 (220–225) long, 155 (146–170) wide at level of s4; setae lengths: j1 17 (16–18), j3 21 (19–22), j4 8 (8–9), j5 6 (6–7), j6 8 (7–8), J1 8 (7–10), J2 9 (8–10), J5 5 (5–6), z2 8, z4 9 (8–9), z5 7 (6–7), Z1 9 (8–10), Z4 9 (8–10), Z5 42 (40–45), s4 17 (17–18), S2 10 (9–10), S4 9 (8–10), S5 7 (7–8), r3 9 (8–10), R1 7 (7–8). Setae r3 and R1 on dorsal shield. All dorsal setae smooth, except Z5 serrate.
Venter — (Figure 1F). Sternogenital shield smooth; ventrianal shield subtriangular, with striation anteriorly and laterally to pores; 94 (87–97) long and 128 (120–132) wide at anterior corners, with three pairs of pre-anal setae (JV1, JV2 and ZV2), two pairs of lyrifissures and one pair of pores postero-mesad of JV2; JV5 23 (22–24). All ventral setae smooth.
Peritreme — Extending to level of z2.
Chelicera — Fixed digit 20 long, with nine teeth in addition to hook and pilus dentilis; movable digit 17 long, with four teeth in addition to hook. Spermatodactyl (Figure 1G) with shaft 18 (16–19) long.
Legs — Macrosetae smooth with knobbed tips, except SgeI smooth and pointed; lengths: SgeI 20, SgeII 19 (18–20), SgeIII 22 (20–23), StiIII 18, SgeIV 36 (34–37), StiIV 22 (21–24), StIV 45 (43–47). Chaetotactic formulae of genu II and genu III as female.
Type specimens
Holotype female, three paratype females and three paratype males on Nephelium lappaceum L. (Sapindaceae), in Silves (02°54'49''S, 58°26'15''W), Amazonas state, 27 October 2019, R.B. Souza collector; two paratype females and one paratype male on Hymenaea sp. (Leguminosae) in Contriguaçu (09°51'00''S, 58°14'00''W), Mato Grosso state, 15 January 2015, J.M. Rezende & A.C. Lofego collectors; one paratype female on Pauroma guianensis Aubl. (Urticaceae), Itacoatiara (3°5'37''S, 58°27'31''W), Amazonas state, 24 November 2017, P.R. Demite & A.C.C. Cavalcante collectors; one paratype female on Hevea brasiliensis (Willd. ex A.Juss.) Müll.Arg. (Euphorbiaceae), Itacoatiara (2°59'41''S, 58°27'12''W), Amazonas state, 25 June 2019, A.C.C. Cavalcante, P.C. Dahmer & R.B. Souza collectors; one paratype female on Pouteria caimito (Ruiz & Pav.) Radlk. (Sapotaceae), in Itacoatiara (2°59'41''S, 58°27'12''W), Amazonas state, 26 September 2019, R.B. Souza collector; one paratype female on Bertholletia excelsa Bonpl. (Lecythidaceae), Parintins (2°40'20''S, 56°46'24''W), Amazonas state, 2 January 2020, M.P. Duque collector; three paratype females, one paratype male and one deutonymph on Leguminosae unidentified, Itacoatiara (03°00'46.8''S, 58°26'58.9''W), Amazonas state, 06 January 2020, P.C. Dahmer collector; one paratype female on Oenocarpus bacaba Mart. (Arecaceae), Maués (3°26'06''S, 57°50'51''W), Amazonas state, 08 March 2020, P.C. Dahmer collector. All type specimens are deposited in the collection of Acari of the Departmento de Zoologia e Botânica, da UNESP, Campus de São José do Rio Preto, São Paulo, Brazil.
Differential diagnosis
Amblydromalus amazonicus n. sp. is more similar to Amblydromalus akiri Nuvoloni, Lofego, Rezende & Feres, 2015b, by the atrium format, widening next to major duct and macroseta SgeIV knobbed. Also is similar to A. insolitus Nuvoloni & Lofego in Nuvoloni et al (2015a), A. itacoatiarensis Demite, Cavalcante & Lofego, 2019 and A. zannoui Famah Sourassou, Sarmento & Moraes, 2017 by atrium shape. In addition, A. akiri differs from A. amazonicus n. sp. by having s4 about 45% longest and StiV and StIV with blunt tips; A. insolitus differs by having j3 and s4 about 40% and 70% longest, respectively; A. itacoatiarensis by having j3, s4, SgeI and StiIV about 50%, 150%, 50% and 40% longest, respectively; A. zannoui by having j1, j3, s4, SgeI, SgeII, StiIV and StIV about 40%, 65%, 70%, 40%, 40%, 200% and 65%, respectively. Additionally, A. insolitus, A. itacoatiarensis and A. zannoui differs of A. amazonicus n. sp. by having all macrosetae pointed. Amblydromalus amazonicus n. sp. differs from all species of genus by the presence of seta J1.
Etymology
The specific name ''amazonicus'' refers to Amazon Forest biome.
Remarks
According to Chant and McMurtry (2005) mites of the genus Amblydromalus have the ratio s4: Z1< 3: 1. The ratio between these two setae in Amblydromalus amazonicus n. sp. was lower, being the type series average 2: 1 (2.2: 1 in the holotype; 1.8–2.4 in the type series specimens), what could lead this species to be classified as Ueckermannseius according to the definition of Chant and McMurtry (2007). However, the s4: Z1 < 3: 1 ratio has also been observed in other species classified as Amblydromalus: Amblydromalus tigrus (Denmark & Evans) (2.1: 1; holotype; Denmark et al. 1999); Amblydromalus ntandu (Pritchard & Baker) (2.1: 1; paratype; Moraes et al. 2006); Amblydromalus manihoti (Moraes) (2.7: 1; holotype; Moraes et al. 1994); Amblydromalus akiri (2.9: 1; type series; Nuvoloni et al. 2015b) and Amblydromalus arawak (De Leon) (2.9: 1; holotype; Moraes et al. 1994). In addition, in other populations of two of these species, different from what was observed in the type series, was observed ratio greater than 3: 1 between s4 and Z1. In material collected from A. ntundu from Cameroon and DR Congo the ratio was 3.1: 1 (Moraes et al. 2006). In the case of A. manihoti, a species frequently collected in South America, the s4: Z1 ratio ranged from 1.8: 1 (Rocha et al. 2015) to 4.1: 1 (Guanilo et al. 2008, Demite et al. 2017). In this way, the s4 / Z1 ration seems to be an unstable character and, therefore, would not be the best character to separate two genera. So, we consider this new species within the genus Amblydromalus, despite the s4: Z1 \textless3: 1 ratio, due it be morphologically closer to some Amblydromalus species, as indicated in differential diagnosis, than any other Ueckermannseius species. We believe that molecular studies or even integrative taxonomy should be performed to verify the phylogenetic relationships within the Typhlodromalina subtribe, and verify if Amblydromalus and Ueckermannseius are in fact two valid clades or if constitute a single natural group.
Within Amblydromalus, only for a single specimen, collected in Cameroon, was reported the presence of seta J1. This specimen was identified as A. swaga (Pritchard & Baker) by Moraes et al. (2006), although for others four specimens analyzed in the same study, including holotype, the presence of J1 was not observed, indicating that the presence of this seta may be an anomaly in A. swaga. Differently in A. amazonicus n. sp. the presence of J1 is a stable character, present in all specimens found, including females, males and deutonymph. Amblydromalus amazonicus n. sp. belongs to the limonicus species group, by having Z4 much shorter than 40% of the distance between its base and that of seta Z5, and shorter than the distance between its base and that of seta S4 (Chant and McMurtry 2005). Despite the presence of seta J1 on dorsal shield we decided to include A. amazonicus n. sp. within the limonicus species group than creating a new one. Within of Euseiini tribe, the seta J1 is recorded in Typhlodromalus fragosoi (Yoshida-Shaul and Chant 1991).
New morphological information on Amblydromalus akiri Nuvoloni, Lofego, Rezende & Feres
In the original description of A. akiri the chaetotactic formula annotate to genu III is 1-2/2-2/0-1, with a total of eight setae (Nuvoloni et al. 2015b). However, an observation on specimens of types series (holotype and paratypes) revealed that the specimens actually have seven setae. The chaetotactic formula should be correct as 1-2/1-2/0-1. The same difference on chaetotaxy was observed also on another species, A. insolitus Nuvoloni & Lofego described with eight setae on genu III (Nuvoloni et al. 2015a) and then Demite et al. (in press) verified that in fact these species bear seven setae on this leg segment, as in type series as on specimens collected in the municipality of Poconé (Pantanal biome), Mato Grosso state, Brazil.
Occurrences of Amblydromalus species in South America.
With the description of A. amazonicus n. sp., the genus currently has 24 described species. Of these 13 are reported on the South American subcontinent (Table 1). All 13 species are registered in Brazil, five described in the last six years (Nuvoloni et al. 2015a, 2015b, Zannou et al. 2017, Demite et al. 2019 and this study). On the other hand, no species of this genus has been registered in Argentina, Chile and Uruguay. This difference in the number of records between countries is mainly due to the lack of surveys of mites in the countries. The last five described species of this genus were collected in regions hitherto little explored (Central-West, Northeast and North regions of Brazil). In addition, the records indicate a probable preference of these mites for regions of a warmer climate, which could explain the absence of these mites in regions further south of the continent.


Key to Amblydromalus species (females) recorded from South America (updated from Demite et al. 2019)
1. Seta Z4 more than 70% as long as distance between its base and that of seta Z5, and distinctly longer than S4 and/or S5
...... congeae species group ... 2
— Seta Z4 much shorter than 40% of distance between its base and that of seta Z5, and at most as long as S4 and/or S5
...... limonicus species group ... 4
2. Seta Z4 relatively shorter, at most as long as distance between its base and base of Z5; insertion of S4 almost in transverse line with insertion of Z4
...... A. laetus (Chant & Baker, 1965)
— Seta Z4 relatively longer, longer than distance between its base and base of Z5; insertion of S4 well posterior to insertion of Z4
...... 3
3. Seta S2 longer than distance between its base and base of Z1
...... A. congeae (De Leon, 1965)
— Seta S2 about 0.5 as long as distance between its base and base of Z1
...... A. macroatrium Moraes, Barbosa & Castro, 2013
4. Seta J1 present
...... A. amazonicus n. sp.
— Seta J1 absent
...... 5
5. Macrosetae of leg IV capitate (SgeIV) or blunt (StiIV and StIV)
...... A. akiri Nuvoloni, Lofego, Rezende & Feres, 2015b
— Macrosetae of leg IV pointed
...... 6
6. Dorsal shield covered mainly by roundish ornamentation (weak), with a few anterolateral striae
...... A. insolitus Nuvoloni & Lofego in Nuvoloni et al. (2015a)
— Dorsal shield mainly smooth, except a few anterolateral striae
...... 7
7. Preanal seta JV1 almost in straight line with setae JV2 and ZV2
...... A. arawak (De Leon, 1966)
— Preanal seta JV1 located anteriorly the setae JV2 and ZV2 (mainly)
...... 8
8. Width of major duct when connecting to atrium greater than width of calyx when connecting to atrium
...... 9
— Width of major duct when connecting to atrium equal to or less than width of calyx when connecting to atrium
...... 10
9. Calyx of spermatheca elongate, with a median narrowing, about 24 µm in length
...... A. zannoui Famah Sourassou, Sarmento & Moraes, 2017
— Calyx of spermatheca tubular, without a median narrowing, about 14 µm in length
...... A. itacoatiarensis Demite, Cavalcante & Lofego, 2019
10. Seta Z4 about two times longer than J2 and Z1; insertion of J5 posterior to insertion of Z5
...... A. villacarmelensis (Moraes) in Moraes et al. (1994)
— Setae J2, Z1 and Z4 sub-equal in length; insertion of J5 in transverse or little anterior line with insertion of Z5
...... 11
11. Setae j3 and s4 sub-equal in length
...... A. manihoti (Moraes) in Moraes et al. (1994)
— Seta j3 shorter than s4 in length
...... 12
12. Seta Z5 longer than s4; calyx of spermatheca 15–20 µm; macroseta of tibia IV (StiIV) about 45 µm
...... A. rapax (De Leon, 1965)
— Seta Z5 shorter than s4; calyx of spermatheca 25–40 µm; macroseta of tibia IV (StiIV) longer than 60 µm
...... A. limonicus (Garman & McGregor, 1956)
Acknowledgments
We thank Isabel Reis Guesdon (Universidade Federal do Amazonas, Itacoatiara, Amazonas) for plant identification. Antonio C. Lofego received a research productivity fellowship from CNPq [Proc. no. 310617/2018-9]. Peterson R. Demite receives a scholarship (PNPD) from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). Priscila C. Dahmer recieved a schorlarship (POSGRAD) from the Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM).
References
- Aponte O., McMurtry J.A. 1993. Phytoseiid mites of Venezuela (Acari: Phytoseiidae). Int. J. Acarol., 19(2): 149-157. doi:10.1080/01647959308683974
- Athias-Henriot C. 1975. Nouvelles notes sur les Amblyseiini II. Le rélève organotaxique de la face dorsal adulte (Gamasides protoadeniques, Phytoseiidae). Acarologia, 17(1): 20-29.
- Bakker F.M., Klein M.E., Mesa N.C., Braun A.R. 1993. Saturation deficit tolerance spectra of phytophagous mites and their phytoseiid predators on cassava. Exp. Appl. Acarol., 17: 97-113. doi:10.1007/BF00156947
- Beard J.J. 2001. A review of Australian Neoseiulus Hughes and Typhlodromips De Leon (Acari: Phytoseiidae: Amblyseiinae). Invertebr. Taxon., 15(1); 73-158. doi:10.1071/IT99017
- Bonato O., Noronha A.C. da S., Moraes G.J. de. 1999. Distribution et échantillonnage des populations de Amblyseius manihoti Moraes (Acari, Phytoseiidae) sur manioc au Brésil. J. Appl. Entomol., 123: 541-546. doi:10.1046/j.1439-0418.1999.00421.x
- Braun A.R., Mesa N.C., Cuellar M.E., Melo E.L., Moraes G.J. de. 1993. Biosystematics of phytoseiid mites (Acari:Phytoseiidae) associated with cassava. Exp. Appl. Acarol., 17: 205-213. doi:10.1007/BF00118437
- Buosi R., Feres R.J.F., Oliveira A.R., Lofego A.C., Hernandes F.A. 2006. Ácaros plantícolas (Acari) da ''Estação Ecológica de Paulo de Faria", estado de São Paulo, Brasil. Biota Neotrop., 6(1): bn02006012006. doi:10.1590/S1676-06032006000100009
- Byrne D.H., Bellotti A.C., Guerrero J.M. 1983. The cassava mites. Trop. Pest Manage., 29(4): 378-394. doi:10.1080/09670878309370833
- Castro T.M.M.G. de, Moraes G.J. de. 2010. Diversity of phytoseiid mites (Acari: Mesostigmata: Phytoseiidae) in the Atlantic Forest of São Paulo. Syst. Biodivers., 8(2): 301-307. doi:10.1080/14772001003801375
- Chant D.A., Baker E.W. 1965. The Phytoseiidae (Acarina) of Central America. Mem. Entomol. Soc. Can., 41: 1-56. doi:10.4039/entm9741fv
- Chant D.A., McMurtry J.A. 2005. A review of the subfamily Amblyseiinae Muma (Acari: Phytoseiidae) Part VI. The tribe Euseiini n. tribe, subtribes Typhlodromalina n. subtribe, Euseiina n. subtribe, and Ricoseiina n. subtribe. Int. J. Acarol., 31(3): 187-224. doi:10.1080/01647950508684424
- Chant D.A., McMurtry J.A. 2007. Illustrated keys and diognoses for the genera and subgenera of the Phytoseiidae of the world (Acari: Mesostigmata). West Bloomfield: Indira Publishing House. pp. 219.
- Chant D.A., Yoshida-Shaul E. 1991. Adult ventral setal patterns in the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol., 17(3): 187-199. doi:10.1080/01647959108683906
- Chant D.A., Yoshida-Shaul E. 1992. Adult idiosomal setal patterns in the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol., 18(3): 177-193. doi:10.1080/01647959208683949
- Collier K.F.S., De Lima J.O.G., Albuquerque G.S. 2004. Predacious mites in papaya (Carica papaya L.) orchards: in search of a biological control agent of phytophagous mite pests. Neotrop. Entomol., 33(6): 799-803. doi:10.1590/S1519-566X2004000600020
- Cruz W.P. da, Krug C., Vasconcelos G.J.N. de, Moraes G.J. de. 2019. Mite (Arachnida: Acari) diversity and abundance on oil palms in the central region of the Brazilian Amazonia. Syst. Appl. Acarol., 24(9): 1736-1750. doi:10.11158/saa.24.9.10
- De Leon D. 1965. Phytoseiid mites from Puerto Rico with descriptions of new species (Acarina: Mesostigmata). Fla Entomol., 48(2): 121-131. doi:10.2307/3493102
- De Leon D. 1966. Phytoseiidae of British Guyana with keys to species (Acarina: Mesostigmata). Stud. Fauna Suriname other Guyanas, 8: 81-102.
- Demite P.R., Cavalcante A.C.C., Lofego A.C. 2019. A new species of Amblydromalus Chant & McMurtry (Acari: Mesostigmata: Phytoseiidae) from Brazil, with a key to Brazilian species of the genus. Syst. Appl. Acarol., 24(12): 2483-2491. doi:10.11158/saa.24.12.13
- Demite P.R., Dias M.A., Cavalcante A.C.C., Ramos M.V.V., Lofego A.C. 2017. Phytoseiid mites (Acari: Mesostigmata: Phytoseiidae) associated with Cerrado biome plants in Brazil, with description of a new species. Syst. Appl. Acarol., 22(12): 2141-2177. doi:10.11158/saa.22.12.9
- Demite P.R., Feres R.J.F., Lofego A.C., Oliveira A.R. 2009. Plant inhabiting mites (Acari) from the Cerrado biome of Mato Grosso State, Brazil. Zootaxa, 2061: 45-60. doi:10.11646/zootaxa.2061.1.3
- Demite P.R., Lofego A.C., Feres R.J.F. 2011. Phytoseiidae (Acari) in forest fragments in the State of São Paulo, Brazil. Zootaxa, 3086: 31-56. doi:10.11646/zootaxa.3086.1.2
- Demite P.R., Lofego A.C., Feres R.J.F. 2013. Mite (Acari; Arachnida) diversity of two native plants in fragments of a semideciduous seasonal forest in Brazil. Syst. Biodivers., 11(2): 141-148. doi:10.1080/14772000.2013.806368
- Demite P.R., Moraes G.J. de, McMurtry J.A., Denmark H.A., Castilho R. de C. 2021. Phytoseiidae Database. [internet] [10 March 2021]. Available from: http://www.lea.esalq.usp.br/phytoseiidae

- Demite P.R., Rezende J.M., Lofego A.C., Amaral F.S.R., Barreto M.R., Moraes G.J. de (in press). Phytoseiid mites (Acari: Mesostigmata: Phytoseiidae) from Mato Grosso State, Central-Western Brazil. An. Acad. Bras. Ciênc.
- Denmark H.A., Evans G.A., Aguilar H., Vargas C., Ochoa R. 1999. Phytoseiidae of Central America (Acari: Mesostigmata). West Bloomfield: Indira Publishing House. pp. 125.
- Eichelberger C.R., Johann L., Majolo F., Ferla N.J. 2011. Mites fluctuation population on peach tree (Prunus persica (L.) Batsch) and in associated plants. Rev. Bras. Frutic., 33(3): 765-773. doi:10.1590/S0100-29452011005000102
- Famah Sourassou N., Sarmento R.A., Moraes G.J. de. 2017. Description of a new species of the Amblydromalus limonicus (Acari: Phytoseiidae) species group based on morphological and molecular evidences. Int. J. Acarol., 43(5): 374-379 doi:10.1080/01647954.2017.1317021
- Farias A.R., Flechtmann C.H.W., Moraes G.J. de, McMurtry J.A. 1981. Predadores do acaro verde da mandioca, no Nordeste do Brasil. Pesqui. Agropec. Bras., 16(3): 313-317.
- Feres R.J.F., Buosi R., Daud R.D., Demite P.R. 2007. Padrões ecológicos da comunidade de ácaros em euforbiáceas de um fragmento de mata Estacional Semidecidual, no Estado de São Paulo. Biota Neotrop., 7(2): bn04907022007. doi:10.1590/S1676-06032007000200022
- Ferla N.J., Marchetti M.M., Gonçalves D. 2007. Ácaros predadores (Acari) associados à cultura do morango (Fragaria sp., Rosaceae) e plantas próximas no Estado do Rio Grande do Sul. Biota Neotrop., 7(2): bn01807022007. doi:10.1590/S1676-06032007000200012
- Ferla N.J., Moraes G.J. de. 2002. Ácaros predadores (Acari) em plantas nativas e cultivadas do Estado do Rio Grande do Sul, Brasil. Rev. Bras. Zool., 19(4): 1011-1031. doi:10.1590/S0101-81752002000400006
- Fiaboe K.K.M., Gondim Jr. M.G.C., Moraes G.J. de, Ogol C.K.P.O., Knapp M. 2007. Surveys for natural enemies of the tomato red spider mite Tetranychus evansi (Acari: Tetranychidae) in northeastern and southeastern Brazil. Zootaxa, 1395: 33-58. doi:10.11646/zootaxa.1395.1.2
- Furtado I.P., Moraes G.J. de, Kreiter S., Flechtmann C.H.W., Tixier M.-S., Knapp M. 2014. Plant inhabiting phytoseiid predators of Midwestern Brazil, with emphasis on those associated with the tomato red spider mite, Tetranychus evansi (Acari: Phytoseiidae, Tetranychidae). Acarologia, 54(4): 425-431. doi:10.1051/acarologia/20142138
- Furtado I.P., Moraes G.J. de, Kreiter S., Knapp M. 2006. Search for effective natural enemies of Tetranyhcus evansi in south and southeast Brazil. Exp. Appl. Acarol., 40: 157-174. doi:10.1007/s10493-006-9045-y
- Garman P., McGregor E.A. 1956. Four new predaceous mites (Acarina: Phytoseiidae). South. Calif. Acad. Scienc. Bull., 55: 7-13.
- Gondim Jr. M.G.C., Castro T.M.M.G., Marsaro Jr. A.L., Navia D., Melo J.W.S., Demite P.R., Moraes G. J. de. 2012. Can the red palm mite threaten the Amazon vegetation? Syst. Biodivers., 10(4), 527-535. doi:10.1080/14772000.2012.752415
- Gonçalves D., Silva G.L. da, Ferla N.J. 2013. Phytoseiid mites (Acari) associated with yerba mate in southern Brazil, with description of a new species. Zootaxa, 3746(2): 357-371. doi:10.11646/zootaxa.3746.2.6
- Guanilo A.D., Moraes G.J. de, Knapp M. 2008. Phytoseiid mites (Acari: Phytoseiidae) of the subfamily Amblyseiinae Muma from Peru, with description of four new species. Zootaxa, 1880: 1-47. doi:10.11646/zootaxa.1880.1.1
- Lindquist E.E. 1994. Some observations on the chaetotaxy of the caudal body region of gamasine mites (Acari: Mesostigmata), with a modified notation for some ventrolateral body setae. Acarologia, 35: 323-326.
- Lindquist E.E., Evans G.O. 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina. Mem. Entomol. Soc. Can., 47: 1-64. doi:10.4039/entm9747fv
- Mendonça A.L., Lofego A.C., Pott A., Daud R.D., Demite P.R. 2019. Phytoseiidae (Parasitiformes: Mesostigmata) from the Pantanal, Mato Grosso do Sul State, Brazil. Syst. Appl. Acarol., 24(4): 587-612. doi:10.11158/saa.24.4.6
- Miranda V.C., Azevedo E.B. de, Cruz W.P. da, Jorge S.J., Pedro-Neto M., Castilho R. de C., Tixier M.-S., Moraes G.J. de, Sarmento R. A. 2021. Potential of the predatory mite Amblydromalus zannoui to control pest mites on Jatropha curcas. Biocontrol. doi:10.1007/s10526-021-10080-z
- Moraes G.J. de, Alencar J.A. de, Lima J.L.S., Yaninek J.S., Delalibera Jr. I. 1993. Alternative plant habitats for common phytoseiid predators of the cassava green mite (Acari: Phytoseiidae, Tetranychidae) in northeast Brazil. Exp. Appl. Acarol., 17: 77-90. doi:10.1007/BF00156945
- Moraes G.J. de, Alencar J.A. de, Wenzel Neto F., Mergulhao S.M.R. 1990. Explorations for natural enemies of the cassava green mite in Brazil. In: Howeler R.H. (Ed.). Proceedings of the Eight Symposium of the International Society of Tropical Root Crops.; Bangkok. p. 35-353.
- Moraes G.J. de, Barbosa M.F. de C., Castro T.M.M.G. de. 2013. Phytoseiidae (Acari: Mesostigmata) from natural ecosystems in the State of São Paulo, Brazil. Zootaxa, 3700(3): 301-347. doi:10.11646/zootaxa.3700.3.1
- Moraes G.J. de, Denmark H.A., Guerrero J.M. 1982. Phytoseiid mites of Colombia (Acarina: Phytoseiidae). Int. J. Acarol., 8(1): 15-22. doi:10.1080/01647958208683273
- Moraes G.J. de, Mesa N.C., Braun A., Melo E.L. 1994. Definition of the Amblyseius limonicus species group (Acari: Phytoseiidae), with descriptions of two new species and new records. Int. J. Acarol., 20(3): 209-217. doi:10.1080/01647959408684019
- Moraes G.J. de, Zannou I.D., Oliveira A.R., Yaninek J.S., Hanna R. 2006. Phytoseiid mites of the subtribes Typhlodromalina and Euseiina (Acari: Phytoseiidae: Euseiini) from sub-Saharan Africa. Zootaxa, 1114: 1-52. doi:10.11646/zootaxa.1114.1.1
- Murphy S.T. 1984. Biological control of the cassava green mite (Mononychellus spp.) in east Africa. Integrated Pest Management of Cassava Green Mite. Proceedings Regional Training Workshop in East Africa; CIBC, Kenya. p. 55-61.
- Navia D., Moraes G.J. de, Lofego A.C., Flechtmann C.H.W. 2005. Acarofauna associada a frutos de coqueiro (Cocos nucifera L.) de algumas localidades das Américas. Neotrop. Entomol., 34(2): 349-354. doi:10.1590/S1519-566X2005000200026
- Noronha A.C. da S., Mesa N.C. 1990. Efeito de diferentes alimentos sobre a fecundidade Amblyseius limonicus (Acarina: Phytoseiidae). Rev. Bras. Mandioca, 9(1/2): 75-81.
- Noronha A.C. da S., Moraes G.J. de. 1989. Flutuação populacional do ácaro verde da mandioca e seus predadores fitoseideos (Acari: Tetranychidae, Phytoseiidae) em Cruz das Almas - Bahia. Rev. Bras. Mandioca, 8(2): 31-39.
- Noronha A.C. da S., Silva E.S., Argolo P.S. 2004. Ocorrência de Typhlodromalus aripo (DeLeon) (Acari, Phytoseiidae) em cultivares de mandioca. Magistra, 16(1): 54-57.
- Nuvoloni F.M., Lofego A.C., Castro E.B., Feres R.J.F. 2015a. Phytoseiidae (Acari: Mesostigmata) from rubber tree crops in the State of Bahia, Brazil, with description of two new species. Zootaxa, 3964(2): 260-274. doi:10.11646/zootaxa.3964.2.6
- Nuvoloni F.M., Lofego A.C., Rezende J.M., Feres R.J.F. 2015b. Phytoseiidae mites associated with Hevea spp. from the Amazon region: a hidden diversity under the canopy of native trees. Syst. Biodivers., 13(2): 182-206. doi:10.1080/14772000.2014.985344
- Pritchard A.E., Baker E.W. 1962. Mites of the family Phytoseiidae from Central Africa, with remarks on genera of the world. Hilgardia, 33: 205-309. doi:10.3733/hilg.v33n07p205
- Reis P.R. 1979. Ácaros da mandioca e seu controle. Inf. Agropecu., 5(59/60): 63-65.
- Rocha M. dos S., Silva G.L. da, Silva J.O. da, Freitas E.M. de, Ferla N.J. 2015. Phytoseiid mites (Acari: Phytoseiidae) in the northern coastal region of the Rio Grande do Sul State, Brazil. Neotrop. Biodivers, 1(1): 22-35. doi:10.1080/23766808.2015.1052230
- Rowell H.J., Chant D.A., Hansell R.I.C. 1978. The determination of setal homologies and setal patterns on the dorsal shield in the family Phytoseiidae (Acarina: Mesostigmata). Can. Entomol., 110: 859-876. doi:10.4039/Ent110859-8
- Samways M.J., Ciociola A.I. 1980. O complexo de artropodos da mandioca (Manihot esculenta Crantz) em Lavras, Minas Gerais, Brasil. Na. Soc. Entomol. Bras., 9(1): 3-10. doi:10.37486/0301-8059.v9i1.199
- Silva E.A., Reis P.R., Zacarias M.S., Marafeli P.P. 2010. Fitoseídeos (Acari: Phytoseiidae) associados a cafezais e fragmentos florestais vizinhos. Ciênc Agrotec., 34(5): 1146-1153.0 doi:10.1590/S1413-70542010000500010
- Souza I.V. de, Argolo P.S., Gondim Jr. M.G.C., Moraes G.J. de, Bittencourt M.A.L., Oliveira A.R. 2015. Phytoseiid mites from tropical fruit trees in Bahia State, Brazil (Acari, Phytoseiidae). Zookeys, 533: 99-131. doi:10.3897/zookeys.533.5981
- Toldi M., Freitas E.M. de, Silva V.L. da, Caumo M., Ferla J.J., Orlandi C.R., da Costa T., Johann L., Ferla N.J. (2021) Communities of predatory mites (Phytoseiidae and Stigmaeidae) in different environments of the Brazilian Pampa. Acarologia, 61(1): 20-30. doi:10.1080/01647954.2021.1915378
- Vasconcelos G.J.N. de, Silva N.M. da. 2015. Plant-inhabiting mites (Arachnida: Acari) in Amazonian agroecosystems. In: Sant'Anna B.S., Takeara R., Abegg M.A. (Eds.). Amazonian Resources: microbiota, fauna, and flora, New York, Nova Science Publishers. p. 99-113.
- Vasconcelos G.J. de, Silva F.R. da, Barbosa D.G.F., Gondim Jr. M.G.C., Moraes G.J. de. 2006. Diversidade de fitoseídeos (Acari: Phytoseiidae) em fruteiras tropicais no estado de Pernambuco, Brasil. Magistra, 18(2): 90-101.
- Yaseen M., Bennett F.D. 1976. Distribution, biology and population dynamics of the green cassava mite in the neotropics. Proceedings of the Fourth Symposium of the International Society of Tropical Root Crops; CIAT, Cali. p. 197-202.
- Yoshida-Shaul E., Chant D. A. 1991. Five new species of Phytoseiidae from Central and South America (Acari: Gamasina). Int. J. Acarol., 17(2): 93-102. doi:10.1080/01647959108683888
- Zacarias M.S., Moraes G.J. de. 2001. Phytoseiid mites (Acari) associated with rubber trees and other euphorbiaceous plants in southeastern Brazil. Neotrop. Entomol., 30(4): 579-586. doi:10.1590/S1519-566X2001000400011



2021-03-16
Date accepted:
2021-05-13
Date published:
2021-05-31
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2021 Demite, Peterson Rodrigo; Rezende, José Marcos; Carvalho Dahmer, Priscila; Cavalcante, Ana Cristina C. and Lofego, Antonio Carlos
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)