



A new species of the genus Opilioacarus With, 1902 (Acari: Opilioacarida) for the Iberian Peninsula
Moraza, María L.1 ; Prieto, Carlos E.2 and Balanzategui, Iñaki3
1✉ Universidad de Navarra, Facultad de Ciencias, Departamento de Biología Ambiental, Campus Universitario, 31080, Pamplona España.
2Universidad del País Vasco, Departamento de Zoología y Biología Celular Animal, Facultad de Ciencia y Tecnología, Apdo. 644, 48080-Bilbao España.
3Estación Experimental de Zonas Áridas (CSIC), Departamento de Ecología Funcional y Evolutiva, 04120-Almería España.
2021 - Volume: 61 Issue: 1 pages: 128-147
https://doi.org/10.24349/acarologia/20214422ZooBank LSID: 0DD47550-9CF2-409B-9387-EE87B2BCD139
Original research
Keywords
Abstract
Introduction
The Order Opilioacarida comprises one family of long-legged mites, brownish gray and with distinctive blue or purple pigment patterns on the boy and legs. These mites possess ancestral attributes such a segmented opisthosoma, some species have three pairs of lateral eyes and tarsi I-IV divided. They are large mites (1.5 to 2.5 mm in length) living in caves, under rocks and in litter, in semiarid environments, tropical forest and warm temperate regions of the world. Opilioacarid mites have been observed feeding on pollen, fungal spores and arthropods remains. In their ontogenetic development, include a prelarval instar able to move although it cannot feed (Walter and Harvey 2009).
The family Opilioacaridae currently includes 53 valid taxa (with two subspecies and three fossil species) distributed in 13 genera (Araújo et al. 2020). Although the family has been recorded around the World (Harvey 1996), the genus Opilioacarus With, 1902 is the only member in the Mediterranean region.
Opilioacarus segmentatus With, 1902 was the first Opilioacarid mite described. This species, together with two new species of the new genus Eucarus, currently O. italicus (With, 1904) and Adenacarus arabicus (With, 1904), was the base to create the suborder Notostigmata by With (1904). Opiloacarids were the great unknown group of mites until the works of Grandjean (1936), Redikorzev (1937), and Chamberlin and Mulaik (1942). In the 1960s these mites began to be taken into consideration (Naudo 1963; Brignoli 1967; Hammen 1968, 1969, 1977; Coineau and Hammen 1979; Leclerc 1989), and since the beginning of the current century, the works are numerous both describing new taxa (Juvara-Bals and Baltac 1977; Vázquez and Klompen 2002, 2009, 2010, 2015; Bernardi et al. 2012, 2014; Vázquez et al. 2014, 2015, 2016; Klompen and Vázquez 2015; Araújo et al. 2018a, b) and providing information on post-embryonic development (Klompen et al. 2015). However, information on the geographic distribution, biology and ecology of the group is scarce (Leclerc 1989; Harvey 1996; Walter and Proctor 1998). At present, the genus Opilioacarus includes two fossil species — O. aenigmus Dunlop et al. 2004 from the Baltic amber (Eocene) and O. groehni Dunlop & Bernardi, 2014 from the Cretaceous Burmese amber —, and three living species: O. segmentatus from Algeria, O. italicus from Sicily and O. brignolii Araújo & di Palma, 2018 from southern Italian peninsula and Sardinia (previously cited as O. italicus by Brignoli in 1967), although its authors do not ruled out that O. brignolii could be a junior synonym of O. italicus.
Opilioacarus italicus was firstly described based on a nymphal instar and Brignoli (1967) redescribed what he considered that species from Sardinia and Apulia, confirming its proximity to O. segmentatus. The later species, redescribed by Grandjean (1936) including information on different types of setae on the palp tarsus, was reported from Uzbekistan (Krivolutsky 1965) and Greece (Thaler and Knoflach 2002), although taking into account biogeographical considerations the ascription to O. segmentatus seems doubtful. On the other hand, Araújo et al. (2018b) ensure the type species of O. italicus is lost, consider it a nomen dubium because its description can be applied to almost all Opilioacaridae, making a proper identification impossible, and describe O. brignolii based on the material recorded by Brignoli (1967), assuming that it could be conspecific, thus a junior synonym of O. italicus.
Opilioacarus italicus and O. segmentatus have been separated by their different idiosomal ornamentation, abdominal shape, and leg proportions, and without a doubt their geographical distribution should be also considered important for the separation of both species.
Materials and methods
Specimens of this new species were directly collected by hand using a brush or collected using pitfall traps. Mites were preserved in 70% ethanol and later mounted using Hoyer's medium on microscope slides after digestion with Nesbitt's fluid and sealed with GLPT insulating varnish. Morphological observations, measurements, and illustrations were made using compound microscopes equipped with differential interference contrast and phase contrast optical systems, drawing tubes, and stage-calibrated eyepiece micrometers.
Setal notation for the subcapitulum and chelicera follows Hammen (1966); for palps and legs Grandjean (1936), Vázquez and Klompen (2002), and Araújo et al. (2018b); for sternal and genital areas follows Klompen et al. (2015), and Vázquez and Klompen (2015). Setal formula for leg segments follows Evans (1963) for Mesostigmata mites (number of anterolateral setae – number of dorsal setae / number of ventral setae – number of posterolateral setae).
Measurements of structures are given in micrometers (µm), indicating the ranges among specimens measured. Idiosoma and prodorsal shield lengths are along midline, from anterior margin of vertex to caudal margin. Leg lengths are from base of the trochanter to apex of the tarsus, excluding the pretarsus. Distinction of pore-like structures on the idiosomal integument are either poroids (lyrifissures) or glandular openings (solenostomes), as distinguished morphologically are depicted in circular form, poroids in elliptical or slit form.
Specimens of the new species are deposited in the Museum of Zoology, University of Navarra (MZUNAV), Pamplona (Spain), in the Arachnological Collection of the Department of Zoology and Animal Cell Biology (ZUPV), University of the Basque Country, Leioa (Spain), and in the Acari Collection of Experimental Station of Arid Zones (EEZA-CSIC), Almería (Spain).
Diagnosis of the genus Opilioacarus
The following combination of characters implements the diagnosis given by Araújo et al. (2018b): Prodorsum with two pairs of lateral eyes; opisthosomal setae restricted to the preanal segment with three to six dorsal setae and one or two pairs of lateroventral setae; abdominal segment with numerous lyrifissures and glands openings arranged in transverse rows; four pairs of stigmata. Leg I is less than three times the length of the idiosoma; telotarsus I with crown-like eupathidion (ζ1) close to a posterodosal group of six sensilla; shortest leg II lacks prefemur and bears a bifid dorsal acrotarsal solenidia. With's organ is membranous and discoid; gnathosomal dorsal solenidia pl4 short and blunt at the base of rutellum. Tritosternum with two setae on each sternapophysis. Adults palp-tarsus with four or five foliate setae d1 and one pectinate seta d2. Cheliceral movable digit with one tooth.
Diagnosis of Opilioacarus baeticus new species
ZOOBANK: 143C7A68-2830-4D27-BF59-A4882A12AA2C ![]()

Palp tarsus with five foliate setae d1, each with three lobes, and pectinate seta d2; sternal verrucae with st1 and four or five (females) or three or four (males) tapering ribbed setae; area between sternal and genital verrucae with two pairs of long, stout and ribbed, tapering setae (st2–st3) similar to st1 and st5 and three to six pairs of stout and ribbed setae in both sexes; pregenital verrucae with st5 and five to seven stout and ribbed setae in both sexes; female lacks pregenital setae, and with five eugenital short, tapering and ribbed setae; males with five tapering, short and smooth pregenital setae in a trapezoidal area and five genital setae in a lobulated area. Anal valves with 12–13 stout and ribbed setae each. Cheliceral movable digit of adults with one ventral fimbriate process; female palp-tarsus with 23 ch setae, male with 16–18 ch setae, both sexes with 14 sm setae. Preanal segment with five dorsal and one pair of lateroventral stout and ribbed setae. Legs IV longer than body length; basitarsus I with a basal dorsal seta short, thick and barbed; tibia II–IV coronida present; genu II–IV with a dorsodistal sensorial seta enlarged and barbed, femur II–IV with a dorsal longitudinal row of blunt setae s, and femur II–IV with dorsal longitudinal rows of short blunt setae.
Description
Based on female (n= 7) and male (n= 4) and female tritonymph (n=1).
Female idiosoma 2161–2445 µm long, 1000–1138 µm wide; male 1944–2000 µm long, 944–1111 µm wide; female tritonymph 1020 µm long, 510 µm wide.
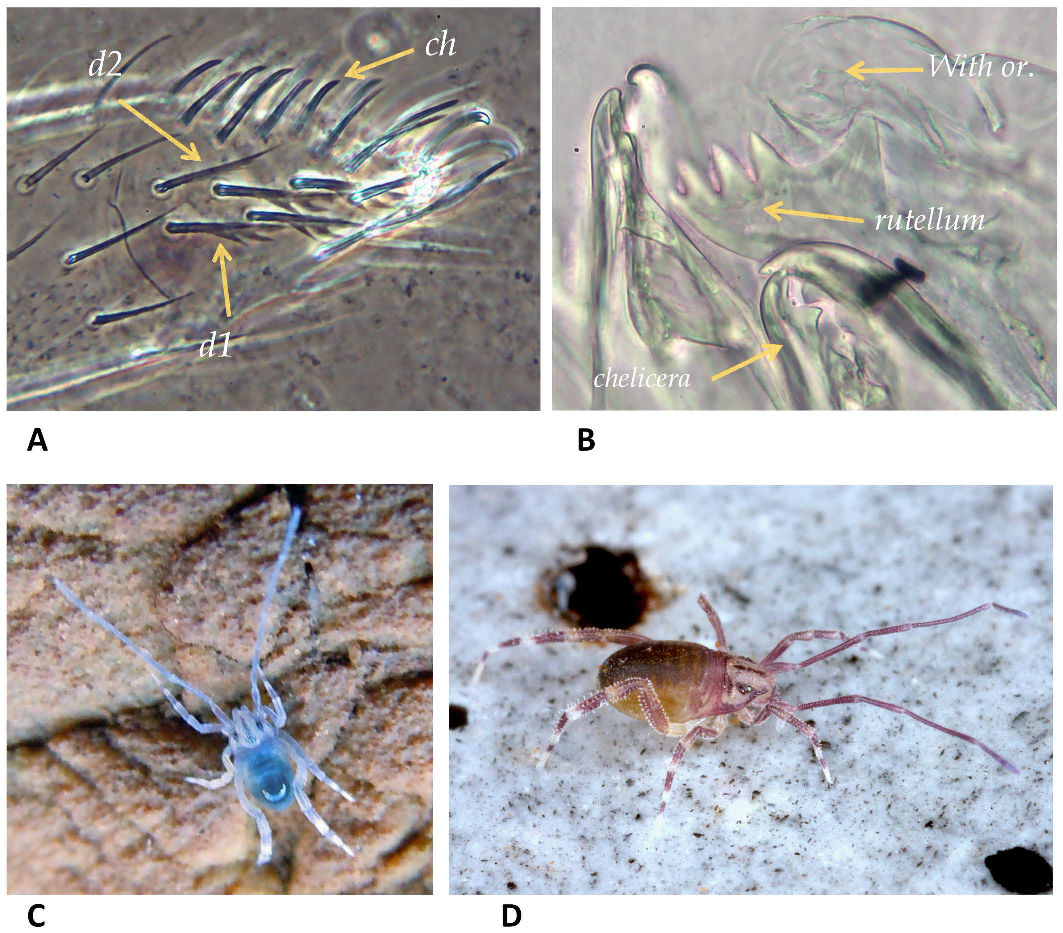
Gnathosoma — Subcapitulum. All paralabial setae present (Figure 2A–C): pl1 conical (19–23 µm long), obtuse, inserted near base of cb3 (69–89 µm long); pl2 (With's organ) discoid, marginally smooth (Figure 11B); pl3 (rutella) inserted dorsolaterally, 122 µm long in the male and 150 µm in the female long, with five distinct teeth in a row (Figure 11B); dorsal solenidia pl4 short and blunt (Figure 2B); all four pairs of circumbucal setae (cb1–4) with bifurcate tips (Figure 2D) and similar in size and length; female with three rounded tip setae vm1, vm2, lvm (Figure 2A) (53–58 µm long), eight pairs of pointed setae 33–45 µm long (lm1-2, vm3, vp1-3, lvp, ldm setae) (Figure 2B); male with one rounded tip seta vm1 (50 long); eight pairs of pointed setae (vm3, lm1-2, vp1-3, lvp, ldm seta (Figure 2). Female tritonymph only cb1-4, pl1-4, lm2 and vp1 setae present. Lateral lips with distinct canals (dl1, dl2) and their orifices (ogl1, ogl2). Hypostome with ventral and apical teeth, most basal teeth rib shape. Dorsal surface of subcapitulum with a medial and laterals coniculate areas (Figure 2B).

Chelicera (Figure 2E-G). Basal segment with one seta (cht1) in normal shape and size; a small denticulate paraxial surface. Cheliceral fixed digit with well-developed apical hook (Figure 2F), and three dorsal setae, ch1 72 µm long, ch2 and ch3 shorter, 25–30 µm long; dorsal and paraxial lyrifissures present; lateral denticle absent; movable digit with well-developed apical hook, ogc present, and one ventral membranous fimbriate process (Figure 2G).
Palps. Female (Figure 3A–L). Palp-tarsus with iα and iπ lyrifissures; five setae d1 foliate (Figure 3A, C, D) with three acute lobules and d2 pectinate (Figure 3E); four setae v1; four v2; three v3; three s (Figure 3E); 14 sm; 23 ch setae (Figure 3D); tarsal claws distal, with ventral margin smooth. Pal-tibia with subequal setae: dorsal setae thin and longer than ventral setae r; ca. 48 setae and four setae s (two dorsal and two ventral) (Figure 3F–H). Genu (Figure 3I, J) with 41 r and 9 palmate setae p (7r+1p – 21+2p / 10+1p – 2r+5p). Femur with 12 r and 15 p (Figure 3L). Trochanter with 6–8 r setae (Figure 3L). Male palp-tarsus similar to female, with five d1 foliate with three lobules and d2 barbed; 16 ch setae. Female tritonymph palp-tarsus (Figure 11A) with four foliate d1, one pectinate d2, three v1, two v2, two v3, three s, 4 sm and 15 ch setae.
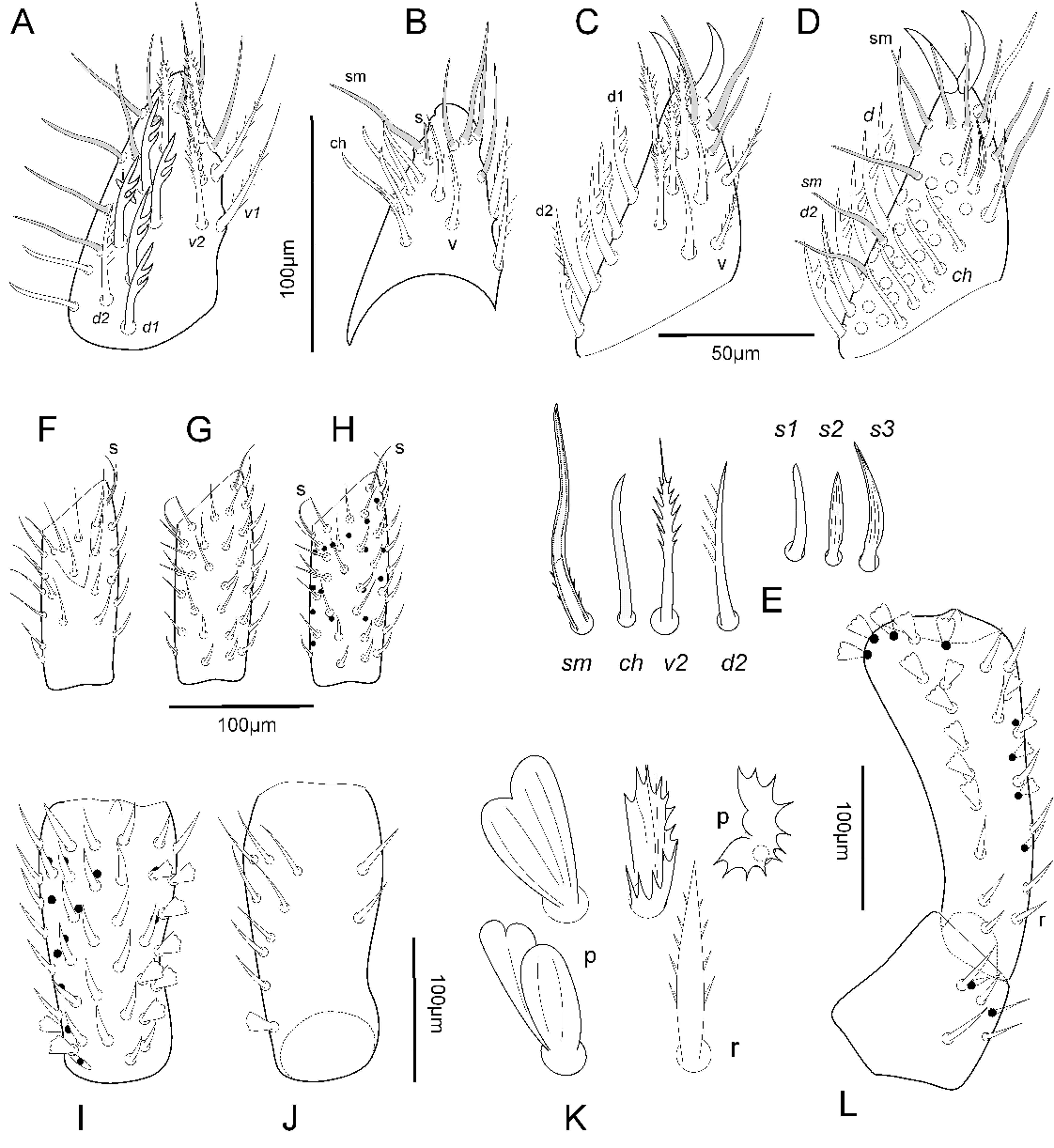
Idiosoma dorsum — Prodorsum. Prodorsal shield covered with round tubercles, pigmented around eyes and in the medial central region, with a dark blue frame, and posterior convex pigmented band (Figure 1B); shield with two pairs of eyes; stout, ribbed setae 16–23 µm long, and one pair of lyrifissures antiaxial to anterior eyes; shield with rounded or slightly acuminate anteriorly margin and a convex row of nine setae at the posterior region (Figure 1C); male 500–597 µm long, 508–778 µm wide at eyes level; female 583–694 µm long, 805–1000 µm wide.
Opisthonotum dark blue-violet with a sharp distinction between colored tergites and pale interarticulated membranes (Figures 1A, 11D); integument coniculate; nymphs may have a bright blue color (Figure 11C). The first two-three abdominal segments marked by distinct grooves and indistinct muscular spots, other segments only marked by distinct spots and an arrangement of lyrifissures and glands (Figure 1A). Except for preanal segment with a row of five dorsal setae and one pair of ventrolateral setae in both sexes (Figure 4F, G), other idiosomal segments lack setae, but with numerous lyrifissures and glands openings arranged in transverse rows. Four pairs of stigmata.
Idiosoma venter — Tritosternum. Each sternapophysis 222–250 µm long, with two setae, one smooth seta 31 µm long at tip and one 88–89 µm long, barbed, positioned more basally (Figure 4A); distal narrow region with smooth cuticle, basal region coniculate.
Sternal region. Sternal verrucae each with a slightly longer ribbed tapering seta st1 (53–57 µm long) and three to five tapering, ribbed setae in the female (Figure 4A) or three or four in the male (Figure 4B) or one in the female tritonymph (female 41–53 µm long, male 47–53 µm long, (Figure 4C). Setae st2 and st3 acute, stout, long, ribbed tapering (female 61–64 µm, male 44–50 µm), and two or three (female) or three (male and female tritonymph) pairs of stout ribbed setae (female 43–50 µm, male 38–45 µm long). Sternal region with three pairs of sternal lyrifissures (iv1–iv3).
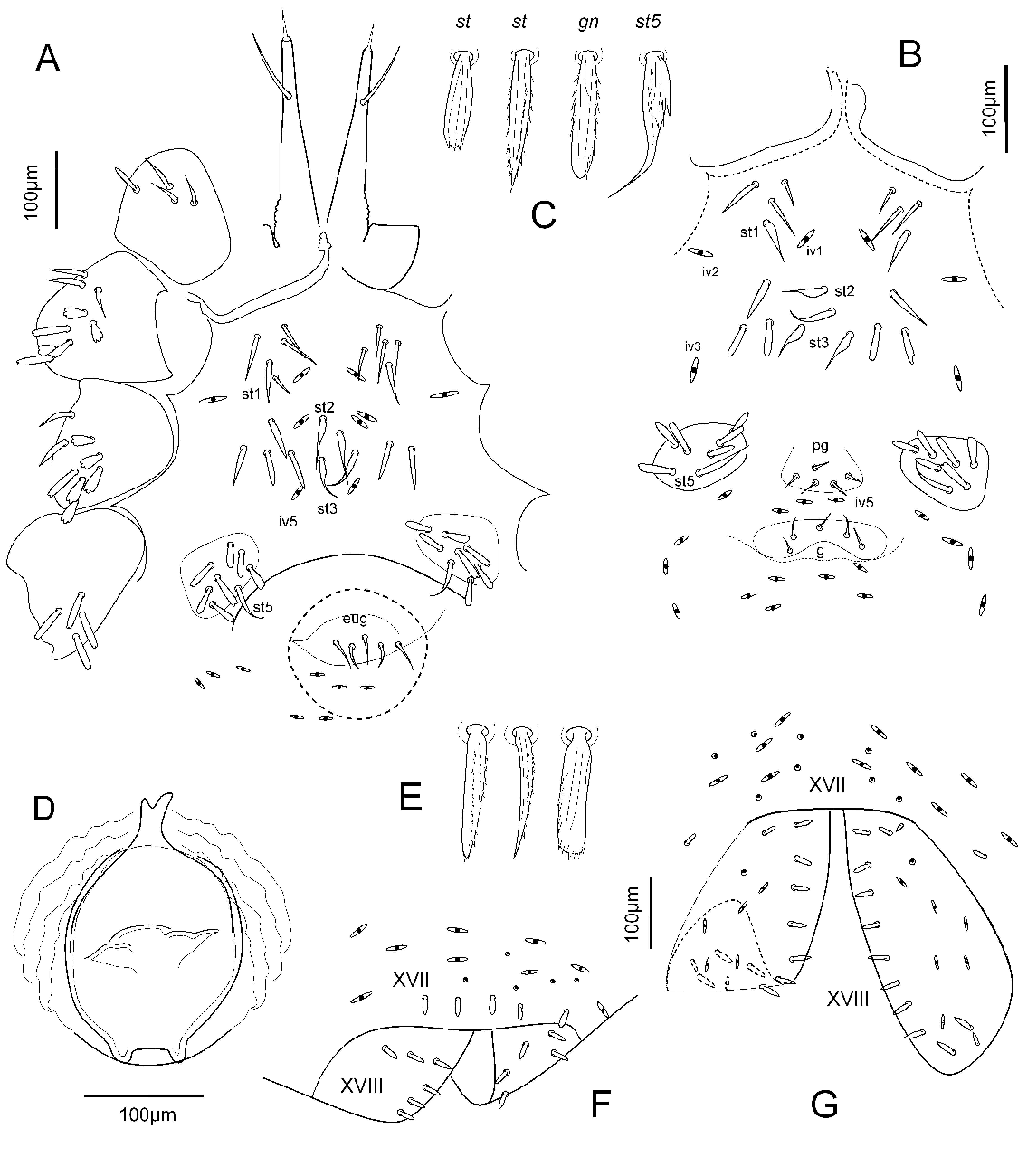
Pregenital area. Adult female and female tritonymph without setae and one pair of lyrifissures (iv4); male with five acute short setae (13–20 µm long) in a trapezoidal area and several lyrifissures. Pregenital capsules each with tapering, ribbed seta st5 (female 55–57 µm, male 36–40 µm long) and five to seven (female) or six to seven (male) or two (female tritonymph) stout ribbed setae (female 27–36 µm, male 27–39 µm long).
Genital area. Female without setae. Male genital area lobulated area with five acute, smooth setae, 19–21 µm long, and at least three lyrifissures. Female with five acute eugenital setae (27–36 µm long); ovipositor consisting of sack shape structure with two digitiform smooth projections (Figure 4D).
Preanal region. Preanal XVI segment without dorsal and ventral setae; preanal segment XVII with five dorsal setae (Figure 4F) and one pair of ventral setae.
Anal valves each with stout ribbed setae (Figure 4G), 12–13 in female and male, six or seven lyrifissures and one to six gland openings.
Legs — Legs with blue-violet stripes and different types of setae, apart from special sensilla on tarsus I (Figure 5B, C: palmate, stout ribbed setae with barbed margin, fan-shaped setae, occasionally with a tiny terminal tip (''papillate'' type ''p'' setae of different length) (Figure 6G); tapering, smooth setae ''s''; tapering and barbed setae, and smooth acute setae.

Leg I–IV lengths from trochanter to base of pretarsus: I 3930 µm, II 1911–2288 µm, III 1900–2055 µm, IV 2766–3044 µm. Coxae I–IV, female 3r+1p, 3r+6p, 2r+6p, 4p (Figure 4A); male 4r, 4r+7p, 1r+7p, 6p.
Legs I. Leg I longer than the body and longer than legs II, III and IV; leg I twice as long than the shortest leg III. Telotarsus I (Figure 5A-C) 336 µm long, with proximal cuticle ornate (approximately first two whorls of setae), other setae on smooth cuticle. Pretarsus with two well-developed sessile claws; Haller's organ with two sensilla in a cavity (Figure 5C) and distal posterodorsal cluster of seven sensilla (Figure 5B): sensillum ζ1 with crown-like tip inserted close to group of sensilla; ω1 and ω2 rod-like with blunt tip, ω3 and ω4 rod-like with filiform tip, ω5 conical, and ω6 spiked maze-like; three anterodorsal sensilla ω with filiform tips at the same level than ζ1; other long telotarsal setae with blunt tips.
Basitarsus I (Figure 5D) slightly longer than telotarsus, 358 µm long; three basal whorls with dorsal and lateral setae serrate, smooth ventral setae, and at least three short blunt solenidia; one barbed dorsal setae shorter and thin than other dorsal setae. Three distal whorls with smooth, blunt tips setae; soft articular tegument between telo- and basitarsus with two ventral lyrifissures.
Tibia (Figure 6A, B) (486 µm long, 61 µm wide at the distal region); distal whorl with 5p+1r setae; rod-like, blunt tip sensilla present in two rows (8 and 10 setae s); 44 setae r + 15 p setae (7 dorsal, 2 anterolateral, 3 posterolateral, 3 ventral) on segment; formula 10 – 10s+11+11+8s / 10 +9 – 10. Pre-tibia I (Figure 6C) (681 µm long, 56 µm wide at the distal region): rod-like setae in two rows (12+12 s); 10 palmate setae (4 dorsal, 4 lateral, 2 ventral).

Genu (Figure 6D) (661 µm long, 86 µm wide at the distal region); 8 setae p on distal region (1 4 / 2 1); rod-like, blunt sensilla present in two rows (7+9 s setae); formula 12p – 7s+(6r+13p) + (5r+13p)+ (1r+9s) / 9r+9r –16p.
Femur (Figure 6E) (854 µm long, 89 µm wide at distal region); distal whorl with 6 setae p and 1 r setae; other whorls with 6 setae; first 4 distal pairs of dorsal setae and 6 posterodorsal setae type r; except for two setae p other ventral setae acute; formula 18p – (4r+17p) +1r+ (10r+8p) / (17r+p) + 19r+1p – 28p. Prefemur (Figure 6F) with one whorl of 5 setae r.
Trochanter (Figure 6F) 454 µm long; distal region with a whorl of 10 stout, ribbed setae and proximal region with 2 setae r; except for 7 setae r type, other setae on the segment are palmate, p setae (Figure 6G).
Acrotarsus II–IV have four whorls of setae (including pretarsal solenidia) and pretarsus with smooth sessile claws, two plumose setae, and two smooth setae. Acrotarsus II (Figure 7D) 89–91 µm long, dorsal solenidia (ωp), lateral (ωd), and dorsal bifid solenidia present; first lateral pair (lv) pectinate, and other acrotarsal setae barbed (4– 6/8–4). Acrotarsus III–IV (Figure 7E) 69–75 µm and 77–99 µm long to base of pretarsus; dorsal bifid solenidia absent (4– 5/8–4).

Telotarsus II–IV (Figure 7A–C) 306 µm, 235 µm, 314 µm long respectively, with ibt and ibv, anterolateral, lateral, and smooth ventral setae with acute thin tips, and dorsal and posterolateral setae stout, long ribbed setae with blunt tips; only telotarsus IV with three dorsal papillate long setae. Telotarsi II–III (Figure 7A, B) with 7 complete whorls of setae (7– 7+7 / 9+9 –7 and 7– 7+7 / 7+7 – 8); telotarsus IV (Figure 7C) with 8 whorls (8– 7+7 / 9+9 – 8), dorsal chaetoma incomplete.
Basitarsus II–IV (Figure 7F, G, H), 417 µm, 314 µm, 573 µm long respectively; ibt, ibv present; distal ωd on the surface in legs II and III and in a cavity present in leg IV (Figure 7H) ; 5, 3 and 2 long distal solenidia between antero- and posterolateral papillate dorsal setae; coronidia present with at least 11, 10 and 20 setae s respectively and one basal g; all lateral setae palmate, specially enlarge in basitarsus IV (Figure 7H); distal pair of ventral setae r longer and stout than other; distal whorl of setae 1p– 2r/2r–1p, 1p–2p/2r–1p and 1p–3p+1s / 2r–1p respectively on legs II-IV; other basitarsal ventral setae long, smooth and acute.
Basitarsi with complex (more than six setae each) whorls of seta; chaetomic formula II–IV respectively excluding dorsal, blunt, long solenidia: 13– 8+9 / 8+9–16; 12–6+7 / 9+9–14; 13–8+7 / 9+10–10.
Genu II–IV (Figure 8A–C) 300 µm, 281 µm, 446 µm long respectively; lyrifissure ibv present; a dorsal longitudinal row of 6, 7 and 8 setae barbed, blunt solenidia s shorter than longer palmate setae p; other dorsal and lateral setae palmate p; 4, 8, 6 ventral setae with acute tips and other ventral setae papillate.

Femur II–IV (Figure 8D–F) elongate and flattened with basal lyrifissures present; 589 µm, 465 µm, 731 µm long respectively; two dorsal longitudinal rows of acute, thin setae, shorter or as long as papillate setae present (4+7, 6+10, 4+11 respectively), other setae palmate of different size, those in the lateral surfaces smaller.
Prefemur (Figure 9D, E). Leg II prefemur absent; prefemur III 173 µm and IV 296 µm long. Except for three (leg III) or two (IV) setae r, other dorsal, lateral and ventral setae palmate.

Trochanters II–IV (Figure 10A-C) 223 µm, 181 µm, 335 µm long respectively, with a dorsal lobulated protuberance; all ventral setae palmate; trochanter II with 4 laterodorsal blunt, barbed, ribbed setae increasing in length toward lateral side, and a longitudinal more lateral row of 3 setae r (Figure 10A); leg III with 1 pair of dorsodistal setae and 2 r basal setae (Figure 10B), and leg IV with 1 pair of dorsal barbed, ribbed setae r (Figure 10C).

Material examined
This species has been found in three localities, plus a photographic record, from southern and southeaster Spain. Holotype: one adult female [ZUPV·5908], Isleta del Moro: Barranco del Paraíso (Níjar, Almería); 30SWF835754, 70 m a.s.l.; 19·Mar.·2015, C. Prieto leg; dry riverbed, under stones. Paratypes: 13 specimens [MZUNAV] from the same locality and date; one female tritonymph [EEZA·854] Pozo de los Frailes: Boca de los Frailes (Níjar, Almería); 30SWF773737, 110 m a.s.l.; 13·Jul.·2018, J. Moya-Laraño leg.; pitfall trap in mediterranean semiarid grassland; one female [ZUPV·637], 5 specimens [MZUNAV], Arroyomolinos: C. Susto (Zahara, Cádiz); 30STF8876, 400 m a.s.l.; 25·Nov.·1988, C. Prieto leg; wasteland, under a large stone. One nymph [photo 299035 at Insectarium Virtual] Purias (Lorca, Murcia); 30SXG219589, 614 m a.s.l.; 08·Dec.·2011, Jose Carrillo leg.; mediterranean shrub on southern slope of rocky hillside.
Etymology
The specific epithet refers to the geographical name of the Betic mountain ranges of the Andalusian System where the specimens were found.
Geographical distribution
Opilioacarus segmentatus was previously reported from several localities in Algeria (With 1902; Grandjean 1936) and from several Greek Islands: Korfu (Silvestri 1905) in the Adriatic sea and Crete (Thaler and Knoflach 2002) and Kassos, Karpathos and Rhodes (Beron 1990) along the southern limits of the Aegean sea. There are also two records from Peloponese, Leonidion by Ludicke and Madel (1937) and Areopoli by Thaler and Knoflach (2002). The new Iberian species expand 800 km the distribution area of Opilioacarus westward. This suggests that the distribution could include the northern part of Morocco (the Rif) also because of the emerged connection between the African Rifean region and the Iberian Betic region, the Gibraltar arc, that closed the Mediterranean sea and drove the Mesinian salinity crisis, about 6 Mya. This event favoured biotic exchange between Africa and Iberia to several mammals (Agustí et al. 2006) and invertebrates adapted to arid land, as scorpions of the genus Buthus (Gantenbein 2004), and could favoured the colonisation of the Betic region by the genus Opilioacarus. But, in respect with Greek records, intermediate land masses as Sicily and the Italian Peninsula were occupied by other species (O. italicus and O. brignolii) and a biogeographical connection between the Aegean region and Algeria is not known, thus suggesting that greek records of O. segmentatus belong to a distinct species.

Taxonomic considerations
According to the original description of O. segmentatus, to its short redescription given by Grandjean in 1936 (who did not examined the types), and to data given by Araújo et al. (2018), several inconsistencies or lack of information are detected regarding the number of setae ch on palp-tarsus and pregenital setae.
The number of palpal setae d1 is five, character not mentioned by With (1902, 1904). The number of setae ch in palp-tarsus varies from the 16 setae illustrated by Grandjean in 1936 (see fig. 5A) to ''17+'' in males mentioned by Araújo et al. (2018).
The number of setae on pregenital capsules varies from three (Araújo et al., 2018) to five to eight indicated by With (see Plate V, figure 10) and Grandjean. With (1904) illustrated a male specimen (see Plate IV, figure 3) since pregenital setae can be observed between the capsules.
Only Araújo et al. (2018) indicate the presence of nine or 10 setae in anal valves. However, the authors mentioned above did not indicate the instar and sex of the specimen when giving this quantitative information.
When studying a female tritonymphal instar of the Iberian O. baeticus specimens, its characters direct us to identify it as O. brignolii (palps with four setae d1, 15 setae ch, and three setae on pregenital capsule (st5+2). However, adult of both sexes clearly differ from O. brignolii and characters of mature instars (both sexes) lead us to think about their belonging to a species close to O. segmentatus. We did not ask for the types of O. segmentatus because, as mentioned by Araújo et al. (2018), the syntypes deposited at ZMUC and MNHN did not allow them to make a proper re-description of O. segmentatus. Despite the similarities found in the descriptions of O. segmentatus given by With (1902, 1904) and Grandjean (1936) with O. baeticus n. sp. we assign these specimens to a new species and take the opportunity to make a description of the species that implement and provides new data from previous descriptions of the species of this genus. The great similarities may suggest that this new species could be a junior synonym of With's species.
Differences between O. baeticus n. sp., O. segmentatus and O. brignolii are indicated in the following tentative key to genera. This key does not include O. italicus because its description is incomplete disallowing a proper evaluation of characters.
Key to the world genera of the Opilioacaridae
Modified from Vázquez et al. 2014 and based on adult specimens.
1. Tritosternum flattened, with 4 to 5 setae on each sternapophysis (Arabia)
...... Adenacarus Van der Hammen
— Tritosternum with 2 setae on each sternapophysis
...... 2
2. Three pairs of eyes; With's organ biramous
...... 3
— Two pairs of eyes; With's organ hyaline, discoid
...... 4
3. Movable digit of chelicera with 1–2 denticles (Central Asia)
...... Paracarus Chamberlin & Mulaik
— Movable digit of chelicera with more than 3 denticles (Indochina)
...... Siamacarus Leclerc
4. With setae on all opisthosomal segments
...... 5
— Setae on opisthosoma restricted to pre-anal segment
...... 6
5. With 5 foliate setae on palp tarsus (SE-Africa)
...... Salfacarus Van der Hammen
— With 6 foliate setae on palp tarsus (Tropical Africa)
...... Panchaetes Naudo
6. Four to six foliate setae on palp tarsus
...... 7
— Eight to twelve foliate setae on palp tarsus
...... 10
7. Leg I three times the length of idiosoma (Central Africa)
...... Phalangiacarus Coineau & Hammen
— Leg I shorter than three times the length of idiosoma
...... 8
8. Three setae on pre-anal segment (1 dorsal, 2 lateroventral) (Americas)
...... Neocarus Chamberlin & Mulaik
— Six to eight setae on pre-anal segment (4–6 dorsal, 1–2 lateroventral)
...... 9
9. Movable digit of chelicera with 4 teeth (India)
...... Indiacarus Das & Bastawade
— Movable digit of chelicera with 1 tooth (Mediterranean region)
...... Opilioacarus With 12
10. Pre-anal segment with 1 dorsal setae
...... 11
— Pre-anal segment with 4–6 dorsal setae (Amazonian basin)
...... Amazonacarus Vázquez, Araújo & Feres
11. Tarsus I sensillum with ''crown-like'' tip distant from main group of sensory sensilla. Palp tibia r-type setae not enlarged (Americas); palp tarsus with more than 7 foliate setae
...... Caribeacarus Vázquez & Klompen
— Tarsus I sensillum not as above. Palp tibia with 14–18 thick, spiny and very large r-type setae which form a crest (Americas)
...... Brasilacarus Vázquez, Araújo & Feres
12. Palp-tarsus with 4 foliate setae d1 and 14–15 (female) and 11–14 (male) ch setae; male with 3 pregenital setae; female with 7 eugenital setae; female idiosoma length 910–1154 µm, male 824-1327 µm; leg I 1726-2249 µm long
...... O. brignolii Araújo & Di Palma
— Palp-tarsus with 5 foliate setae d1
...... 13
13. Palp-tarsus with more than 17 ch setae; both sexes pregenital capsule with st5 and 5 spinose setae; males with 3 or 5 pregenital setae; 9–10 pairs of anal setae; palp-trochanter with 2 pointed setae; palp-tibia with fan setae; largest female idiosoma length 2700 µm, smallest male 2200 µm long; leg I 3180–3480 µm long; leg IV 3290–3600 µm long
...... O. segmentatus With
— Palp-tarsus with 23 (female) and 16 (male) ch setae; both sexes pregenital capsule with st5 and 6–7 spinose setae; female with 0 and male with 5 pregenital setae; palp-trochanter with 6 pointed setae; palp-tibia without fan setae; female idiosoma length 2161–2750 µm, male 1940–2000+ µm; leg I 3930 µm long; leg IV 2766–3044 µm
...... O. baeticus n. sp.
Remarks
According to the dimensions given for adults specimens, O. segmentatus appears to be a species larger than O. brignolii (twice its length) and slightly smaller than O. baeticus n. sp.; O. italicus, known only by a tritonymph of 1.25 mm, thus larger than most adults specimens of O. brignolii.
Acknowledgements
This study was supported by research grants CGL2015-66192-R and PID2019-103863RB-I00 from the Spanish Ministry of Economy and Competitiveness and the Ministry of Science and Innovation respectively. Both to MMZ.
Ours sincere thanks to Jose Carrillo and Eduardo Mateos for providing the photographic material of this species, and to Jordi Moya and Eva de Mas for providing specimens from Almería and their suggestions and revision of the manuscript.
References
Agustí J., Garcés M., Krijgsman W. 2006. Evidence for African-Iberian exchanges during the Messinian in the Spanish mammalian record. Palaeogeography, Palaeoclimatology, Palaeoecology, 238(1-4): 5–14. doi:10.1016/j.palaeo.2006.03.013
Araújo M.S., Bichuette M.E., Bauchan G.R., Ochoa R., Feres R.J.F. 2018a. A new species of cave dwelling Neocarus (Acari: Opilioacaridae) from Bahia state, Brazil, with remarks on taxonomic characters. Zootaxa, 4402 (2): 303–322. doi:10.11646/zootaxa.4402.2.4
Araújo M., di Palma A., Feres J.F. 2018b. A new species of Opilioacarus With, 1902 (Acari: Opilioacaridae) from Italy, and a new diagnosis of the genus. Zootaxa, 4500 (1): 135–145. doi:10.11646/zootaxa.4500.1.9
Araújo, M.S., Di Palma, A., Feres, R.J.F. 2020. Catalog of the Opilioacarida (Acari: Parasitiformes). Zootaxa, 4895: 332–356. doi:10.11646/zootaxa.4895.3.2
Bernardi L.F.O., Zacarias M.S. Ferreira R.L. 2012. A new species of Neocarus Chamberlin & Mulaik, 1942 (Acari: Opilioacarida) from Brazilian caves and karst areas. Zootaxa, 3416: 53–68. doi:10.11646/zootaxa.3416.1.5
Bernardi L.F.O., Klompen H., Ferreira R.L. 2014. Neocarus caipora, a new mite species (Parasitiformes: Opilioacarida: Opilioacaridae) from Brazilian amazon caves. Acarologia, 54: 47–56. doi:10.1051/acarologia/20142113
Beron P.K. 1990. On the occurrence of Opilioacarus segmentatus With, 1903 (Arachnida, Opilioacarida) on the islands of Kassos, Karpatos and Rhodes (Greece). Act. Zool. Bul., 39.
Brignoli M. 1967. Su Opilioacarus italicus (With) (Acarina, Notostigmata). Fragm. Entomol., 5: 111–121.
Coineau Y., Van der Hammen L. 1979. The postembryonic development of Opilioacarida, with notes on new taxa and on a general model for the evolution. In: Piffl E. (Ed) Proceedings 4th international congress of acarology, Saalfelden (Austria), Akadémiai Kiadó, Budapest, pp. 437–441.
Chamberlin R.V., Mulaik S. 1942. On a new family in the Notostigmata. Proceedings of the Biological Society of Washington, 55: 125–132.
Das N.P.I., Bastawade D.B. 2007. The first report of the acarine suborder Opilioacarida from India, with description of new genus Indiacarus, and a new species, Indiacarus pratyushi. Acarologia, 47: 3–11.
Dunlop J.A., Bernardi L.F.O. 2014. An opilioacarid mite in Cretaceous Burmese amber. Naturwissenschaften, 101: 759–763. doi:10.1007/s00114-014-1212-0
Dunlop J.A., Wunderlich J., Poinar G.O. Jr. 2004. The first fossil opilioacariform mite (Acari: Opilioacariformes) and the first baltic amber camel spider (Solifugae). Trans. R. Soc. Edinburgh, Earth Sciences, 94: 261–273. doi:10.1017/S0263593300000663
Evans, G.O. (1963) Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bulletin of the British Museum (Natural History). Zoology, 10, 275–303. doi:10.5962/bhl.part.20528
Gantenbein B. 2004. The genetic population structure of Buthus occitanus (Scorpiones: Buthidae) across the Strait of Gibraltar: calibrating a molecular clock using nuclear allozyme variation. Biol. J. Linn. Soc., 81(4): 519–534. doi:10.1111/j.1095-8312.2003.00295.x
Grandjean F. 1936. Un acarien sysnthétique: Opilioacarus segmentatus With. Bull. Soc. Hist. Nat. Afrique N., 27: 413–444.
Hammen L. van der 1966. Studies on Opilioacarida (Arachnida) I. Description of Opilioacarus texanus (Chamberlin & Mulaik) and revised classification of the genera. Zool. Verhandelingen, 86: 3–80.
Hammen L. van der 1968. Studies on Opilioacarida (Arachnida). II. Redescription of Paracarus hexophthalmus (Redikorzev). Zool. Mededelingen, 43 (5): 57–76.
Hammen L. van der 1969. Studies on Opilioacarida (Arachnida) III. Opilioacarus platensis Silvestri, and Adenacarus arabicus (With). Zoologische Mededelingen 44 (8): 113–131.
Hammen L. van der 1977. Studies on Opilioacarida (Arachnidea) IV. The genera Panchaetes Naudo and Salfacarus gen. nov. Zool. Mededelingen, 51: 43–78.
Harvey M.S. 1996. Small arachnids and their value in Gondwanan biogeographic studies. In: Hopper S.D. Chappill J.A., Harvey M.S., George A.S. (Eds). Gondwanan heritage: past, present and future of the Western Australian Biota. Surrey Beatry & Sons, Chipping Norton. pp. 155–162.
Juvara-Bals I, Baltac M. 1977. Deux nouvelles espèces d'Opilioacarus (Acarina: Opilioacarida) de Cuba. In: Orghidan T, Núñez A Jiménez, Decou V, Negrea S, Bayés NV (Eds.). Résultats des expéditions biospéleogiques Cubano-Roumaines á Cuba. Academiei Republicii Socialiste Romania, Bucuresti, pp. 169–184.
Klompen H., Vázquez M.M., Bernardi L.F.O. 2015. Post-embryonic development in the mite suborder Opilioacarida, with notes on segmental homology in Parasitiformes (Arachnida). Exp. Appl. Acarol., 67: 183–207. doi:10.1007/s10493-015-9939-7
Krivolutsky D.A. 1965. A new finding of Opilioacarina in USSR. Russ. J. Zool., 44: 1413.
Leclerc P. 1989. Considerations paléogéographique á propos de la découverte en Thaïlande d'Opilioacariens nouveaux (Acari-Notostigmata). C. R. Soc. Biogéogr., 65: 162–174.
Ludicke M., Madel W. 1937. Biozönotische Studien in der griechischen Immergrünen Region. Zool. Jahrb. Abt. Syst. Ókol. Grogr. (Berlin), 69: 300.
Naudo M.H 1963. Acariens Notostigmata de l'Angola. Publicaçoes Culturais da Companhia de Diamantes de Angola, 63: 13–24.
Redikorzev V. 1937. Eine neue Opilioacarus-Art. Zool. Anz., 118: 10–12.
Silvestri F. 1905. Note Aracnologische. I-III. Redia, 2: 239–261 + plates XXI-XXIV.
Thaler K., Knoflach B. 2002. Neue Opilioacarus-Funde (Acari, Notostigmata) in Peleponnes und Ägäis (Griechenland). Entomol. Nachr. Ber., 46: 271–277.
Vázquez M.M., De Araújo M.S., Feres R.J.F. 2014. A new genus and two new species of Opilioacaridae (Acari: Parasitiformes) from Amazonia, Brazil with a key to world genera. Zootaxa, 3814 (2): 151–176. doi:10.11646/zootaxa.3814.2.1
Vázquez M.M., De Araújo, M.S., Feres R.J.F. 2015. Brasilacarus cocaris (Acari: Opilioacaridae), a new genus and species from Amazonia, Brazil. Zootaxa, 3915: 375–389. doi:10.11646/zootaxa.3915.3.3
Vázquez M.M., Araújo M.S., Feres R.J.F. 2016. A new species of Diplothyrus (Parasitiformes: Neothyridae) from Brazil. Acarologia, 56: 269–278. doi:10.1051/acarologia/20162243
Vázquez M.M., Klompen H. 2002. The family Opilioacaridae (Acari: Parasitiformes) in North and Central America, with description of four new species. Acarologia, 42: 299–322.
Vázquez M.M., Klompen H. 2009. New species of New World Opilioacaridae (Acari: Parasitiformes) with the description of a new genus from the Caribbean region. Zootaxa, 2061: 23–44. doi:10.11646/zootaxa.2061.1.2
Vázquez M.M., Klompen H. 2010. The genus Salfacarus (Acari: Opilioacarida) in Madagascar. Zootaxa, 2482: 1–21. doi:10.11646/zootaxa.2482.1.1
Vázquez M.M., Klompen H. 2015. The family Opilioacaridae (Parasitiformes: Opilioacarida) in Mexico, description of two new species and notes on biology and geographical distribution. Zootaxa, 3957 (5): 535–552. doi:10.11646/zootaxa.3957.5.3
Walter D.E., Proctor H.C. 1998. Feeding behaviour and phylogeny: observations on early derivative Acari. Exp. Appl. Acarol., 22 (1998): 39–50. doi:10.1023/A:1006033407957
With C.J. 1902. A new acaride Opilioacarus segmentatus. Forhandlingar vid Nordiska Naturforskareog Lakaremotet i Helsingfors den 7 till 12 Juli 1902 (Comptes Rendus du Congrès des Naturalistes et Médecins du Nord tenu à Helsingfors), Sektionen för Zoologi, 20: 4–5.
With C.J. 1904. The Notostigmata, a new suborder of Acari. Vidensk. Meddel. Dansk Naturhist. Foren. Kjøbenhavn, 137–191.
Walter D.E., Harvey M.S. 2009. Order Opilioacarida. In: Krantz G.W, Walter D.E. (Eds.). A Manual of Acarology. Texas Tech University Press, pp.104–105.



2020-09-17
Date accepted:
2021-02-23
Date published:
2021-03-04
Edited by:
Tsolakis, Haralabos

This work is licensed under a Creative Commons Attribution 4.0 International License
2021 Moraza, María L.; Prieto, Carlos E. and Balanzategui, Iñaki
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)