



First evidence of predation of the ant species Lasius alienus on the poultry red mite Dermanyssus gallinae
Kar, Sirri  1
; Akyildiz, Gurkan2
; Sirin, Deniz3
; Rodriguez, Sergio E.4
and Camlitepe, Yilmaz5
1
; Akyildiz, Gurkan2
; Sirin, Deniz3
; Rodriguez, Sergio E.4
and Camlitepe, Yilmaz5
1✉ Department of Biology, Tekirdag Namik Kemal University, Tekirdag, Turkey.
2Department of Biology, Tekirdag Namik Kemal University, Tekirdag, Turkey.
3Department of Biology, Tekirdag Namik Kemal University, Tekirdag, Turkey.
4Galveston National Laboratory, Department of Microbiology and Immunology, Institute for Human Infections and Immunity, University of Texas Medical Branch, Galveston, Texas, USA.
5Department of Biology, Trakya University, Edirne, Turkey.
2021 - Volume: 61 Issue: 1 pages: 115-120
https://doi.org/10.24349/acarologia/20214420Short note
Keywords
Abstract
Introduction
Dermanyssus gallinae (Poultry red mite, PRM) is a nidicolous, nocturnal and hematophagous ectoparasite that infests mainly chickens and other birds, and rarely other warm-blooded animals and humans (Roy and Chauve, 2010; George et al. 2015). PRM is the most common parasitic problem in commercial laying facilities worldwide (Tomley and Sparagano, 2018). Until today, many biological control methods have been attempted against PRM (Sparagano et al. 2014). In this context, some predator mites such as Androlaelaps casalis, Gaeolaelaps aculeifer, Stratiolaelaps scimitus and Cheyletus eruditus (Buffoni et al. 1997; Ali et al. 2012; Lesna et al. 2012) have been tested. However, an ideal mitigation goal has not been achieved yet (Mul et al. 2009; Sparagano et al. 2014; Knapp et al. 2018).
The ant species Lasius alienus (Förster, 1850) is one of the most abundant ant species in the Western Palearctic. It occurs mostly in natural open habitats, but is also found in light forest, forest edges and urban areas (wooded gardens, residential areas). It can reach nest densities up to 100 nests/100 m2 (Seifert, 2018). Nests of this species are usually found in the soil, under stones, and occasionally in woods or some other materials. Workers (2-4 mm in length) establish foraging trails on the ground and in trees. The number of workers in a colony, which is active in cold and warm months, can be more than 10,000. Although the predation of many ant species on various arthropod groups has been well documented (Sanders and van Veen, 2011; Campolo et al. 2015; Milligan et al. 2016; Morris and Perfecto, 2016), there is no detailed data about L. alienus. However, this species was reported to consume both dead and small live insects, gather plant nectar, and feed on honeydew secreted by aphids. Workers can also forage in dwellings in search of food (Collingwood, 1979; Seifert, 1992; Robinson, 2005; Dussutour and Simpson, 2009).
This research was undertaken to look for evidence of possible predator activity of L. alienus on D. gallinae.
Materials and Methods
Background
In the study, a laboratory population of D. gallinae was used, which was established from the field samples collected from Turkish farms in 2015 and identified following Roy et al. (2009) and Di Palma et al. (2012). The mites were maintained in a parasite investigation unit (40o59''N, 27o34'E; altitude: 17 m), of which parts were set up as a backyard coop complex in a woodland area. This coop complex contained a free-range area (4 x 5 m) and a roosting area (1x1.5 m) for chickens, with soil and concrete ground, respectively. The main component of the unit was enclosed by a wire mesh that allows the passage of ants.
The mites were placed on the solid top reservoir (25 x 50 cm size; 8 cm depth) of a four-legged (h: 20 cm) wooden roosting stand (Supplementary material 1). The reservoir was supplied with wood pieces and dry pine needles to provide hiding places for the mites. The stand was placed in the roosting area of the unit as an additional roosting site. Four to six adult hens were continuously present in the unit during the study period and were fed ad libitum.
In a first attempt to establish the mites in the facility, the population disappeared within the first week of introduction. Although ants' foraging trails leading to the top of the stand were observed, we did not realize that the ants may lead to the loss of the mite population. In the second attempt, we noticed that the mites were captured from their hiding sites by the ants during the daytime. Investigation of the area around the study units revealed natural occurrence of colonies of ants that we identified as L. alienus following Seifert (1992).
Demonstration of ant predation on PRM
Experiment 1
Foraging ants occurring naturally in the environment around the experimental unit in the summer period were attracted to the top of a white painted wooden table (75x100 cm, h: 80 cm) placed within the unit, using a piece of cotton impregnated with diluted commercial sugar and watermelon placed in a Petri dish (Supplementary material 2). The next day, two foraging trails originating from two distinct ant colonies led to the supplied food, and ca. 100 ants were observed on the Petri dish at any given time. Then, four new Petri dishes were placed (T0) around the initial dish (Figure 1A). The four new dishes contained respectively: (i) pieces of dry hen feces colonized by ca. 100 adults (fed female: 1-1.5 mm) and other stages (egg, larva, and nymph) of D. gallinae, (ii) mite-free dry hen feces, (iii) commercial sugar diluted with watermelon juice, and (iv) nothing (negative control). All the dishes were sealed with a thin cloth strainer to prevent the entry of ants and the exit of mites (Figure 1). Ant behavior was observed for one hour, starting immediately after setting the four Petri dishes (T0), and rechecked after 24 hours.

During the 1-hour observation period at T0, photographs were taken at the 5th, 10th, 20th, 30th, 40th, 50th, and 60th minutes. The ants which climbed on the four new Petri dishes, were counted on the pictures. Video footage were used to record the time spent by each ant on the dishes.
Experiment 2
In order to observe the direct interest of the ants in mites, ants were attracted to the top of the white-painted wooden table as described in Experiment 1. Hen-bedding material containing live mites at various biological stages was placed next to the sugar-containing Petri dish and was readily invaded by foraging ants. Ant behavior was observed, and a video has been shot (Supplementary material 2). This experiment was repeated monthly during the warmer months to observe whether the interest of ants in mites was persistent across the season of ant activity.
Experiment 3
To observe the interest of the ants in different biological stages of PRM, the same setup as in Experiment 2 was established again. A pile of dry hen feces colonized by mites was divided into two equal pieces, in such a way as to include an approximately similar number of biological forms of D. gallinae (white stages (eggs, larvae, unfed nymphs), and \textasciitilde100 fed reddish nymphs and adults). One of the pieces was placed next to the sugar-containing Petri dish invaded by ants. The other piece, which was used as a negative control, was kept at -20 oC to hold the mites inactive. After 24 hours, the piece of the feces presented to the ants and the other piece kept in the freezer as a negative control were examined and compared under the stereomicroscope (Figure 2).
Results and discussion
Our first experiment suggested that the ants were most attracted by the dish containing dry hen feces colonized by D. gallinae (Table 1, Figure 1A, Supplementary material 2). Furthermore, during the 1-hour observation period at T0, it was observed that more than ten ants attempted to open the strainer on the top of the Petri dish containing the feces colonized by D. gallinae (Figure1B, Supplementary material 2). This type of effort was also observed in the dish containing mite-free hen feces on 3 ants during the 1-hour observation period at T0. As the strainer prevented direct contact between ants and PRM, this experiment suggests that ants are attracted by PRM and/or hen feces at a distance. At the 24th hour time point of the experiments; no particular interest of the ants was observed in any of the four Petri dishes.
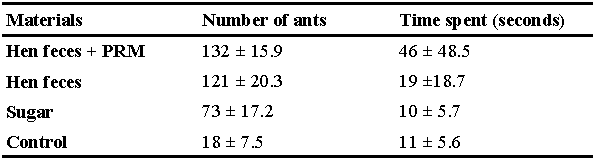

During Experiment 2, ant movements directed to the sugar-containing Petri dish changed immediately after placing the hen-bedding material containing mites. The ants started to move quickly and irregularly and began to collect mites, carrying them subsequently to their nests (Figure 1C, Supplementary material 2). Ant workers bent their gaster and likely sprayed formic acid when catching live adult mites, but rarely when catching larvae and nymphs (Figure 1D, Supplementary material 2). The repetitions of this experiment during the warmer months showed that the interest of ants in mites was persistent across the season of ant activity. Although a quantitative measurement could not be done, no qualitative change was observed in this interest during the warmer months.
In Experiment 3, the stereomicroscope examination performed at T+24th hour showed that the workers of L. alienus took away the eggs, larvae, nymphs, adults, and even eggshells of D. gallinae from the piece presented to them for twenty-four hours (Figure 2).

The localization of D. gallinae in the upper parts of the poultry housings has been considered a restrictive factor for the effectiveness of some predatory mite species on PRM, as those predatory mites are mostly located in manure, on the bottom of housings (Lesna et al. 2012). In our study, we observed that ant workers were able to find and eliminate all forms of PRM within a range of several meters from their colony, even at the top of the stand, including from dried chicken feces where PRM frequently hide. This suggests that L. alienus workers would be efficient at finding mites in the different parts of the hen house. Furthermore, no ant was seen on the chickens or their food, and no chicken was noticed to eat the ants or interfere with them. So, there is little risk that this ant species would be a vector of poultry pathogen or parasite. In fact, some ant species are known to be the intermediate host of some poultry parasites (Taylor et al. 2016), but there is no report on the vector potential of L. alienus for any vertebrates. It was reported that the workers of some ant species (e.g., the fire ant Solenopsis invicta) could sting and bite chickens (Tomberlin and Drees, 2007). However, L. alienus does not have a sting, and, unlike its close relative species L. niger, it is known as a non-aggressive species (Collingwood, 1979; Robinson, 2005).
We showed that L. alienus is a potential predator on all biological stages of PRM. This prey-predator interaction is worth investigating more thoroughly as it might lead to an effective biological control strategy.
Supplementary data
Supplementary material 1
General view of the wooden roosting stand used for the propagation of D. gallinae.
Supplementary material 2
Video showing L. alienus workers trying to open strainer to get inside the Petri dish that contained hen feces colonized by the poultry red mite Dermanyssus gallinae (PRM) in experiment 1 (Part I) and hunting PRM in experiment 2 (Part II).
acarologia_4420_Supplementary-material-1.tif
acarologia_4420_Supplementary-material-2.mp4
References
Ali W., George D.R., Shiel R.S., Sparagano O.A., Guy J.H. 2012. Laboratory screening of potential predators of the poultry red mite (Dermanyssus gallinae) and assessment of Hypoaspis miles performance under varying biotic and abiotic conditions. Vet. Parasitol., 187: 341-344. doi:10.1016/j.vetpar.2012.01.014
Buffoni G., Di Cola G., Baumgartner J., Maurer V. 1997. The local dynamics of acarine predator-prey (Cheyletus eruditus-Dermanyssus gallinae) populations: identification of a lumped parameter model. Mitt. Schweiz. Entomol. Ges., 70: 345-359. doi:10.5169/seals-402682
Campolo O., Palmeri V., Malacrinò A., Laudani F., Castracani C., Mori A., Grasso D.A. 2015. Interaction between ants and the Mediterranean fruit fly: new insights for biological control. Biol. Control., 90: 120-127. doi:10.1016/j.biocontrol.2015.06.004
Collingwood C.A. 1979. The Formicidae (Hymenoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand., 8: 1-174.
Di Palma A., Giangaspero A., Cafiero M.A., Germinara G.S. 2012. A gallery of the key characters to ease identification of Dermanyssus gallinae (Acari: Gamasida: Dermanyssidae) and allow differentiation from Ornithonyssus sylviarum (Acari: Gamasida: Macronyssidae). Parasit. Vectors., 5: 104. doi:10.1186/1756-3305-5-104
Dussutour A., Simpson S.J. 2009. Communal nutrition in ants. Curr. Biol., 19: 740-744. doi:10.1016/j.cub.2009.03.015
George D.R., Finn R.D., Graham K.M., Mul M.F., Maurer V., Valiente Moro C., Sparagano O.A.E. 2015. Should the poultry red mite Dermanyssus gallinae be of wider concern for veterinary and medical science? Parasit. Vectors., 8: 178. doi:10.1186/s13071-015-0768-7
Knapp M., Van Houten Y., Van Baal E., Groot T. 2018. Use of predatory mites incommercial biocontrol: current status and future prospects. Acarologia, 58: 72-82. doi:10.24349/acarologia/20184275
Lesna I., Sabelis M.W., van Niekerk T.G.C.M., Komdeur J. 2012. Laboratory tests for controlling poultry red mites (Dermanyssus gallinae) with predatory mites in small \$'\$laying hen\$'\$ cages. Exp. Appl. Acarol., 58: 371-383. doi:10.1007/s10493-012-9596-z
Milligan M.C., Johnson M.D., Garfinkel M., Smith C.J., Njoroge P. 2016. Quantifying pest control services by birds and ants in Kenyan coffee farms. Biol. Conserv., 194: 58-65. doi:10.1016/j.biocon.2015.11.028
Morris J.R., Perfecto I. 2016. Testing the potential for ant predation of immature coffee berry borer (Hypothenemus hampei) life stages. Agr. Ecosyst. Environ., 233: 224-228. doi:10.1016/j.agee.2016.09.018
Mul M., Van Niekerk T., Chirico J., Maurer V., Kilpinen O., Sparagano O., Thind B., Zoons J., Moore D., Bell B., Gjevre A.G., Chauve C. 2009. Control methods for Dermanyssus gallinae in systems for laying hens: results of an international seminar. World's Poultry Sci. J., 65: 589-600. doi:10.1017/S0043933909000403
Roy L., Chauve C. 2010. The genus Dermanyssus (Mesostigmata: Dermanyssidae): history and species characterization. In: Sabelis, M.W. and Bruin, J. (Eds.), Trends in Acarology: Proceedings of the 12th International Congress. Springer Science, Business Media, pp. 49-55. doi:10.1007/978-90-481-9837-5_8
Roy L., Dowling A.P.G., Chauve C.M., Buronfosse T. 2009. Delimiting species boundaries within Dermanyssus Dugès, 1834 (Acari: Dermanyssidae) using a total evidence approach. Mol. Phylogenet. Evol., 50: 446-470. doi:10.1016/j.ympev.2008.11.012
Sanders D., van Veen F.J.F. 2011. Ecosystem engineering and predation: the multi-trophic impact of two ant species. J. Anim. Ecol., 80: 569-576. doi:10.1111/j.1365-2656.2010.01796.x
Seifert B. 1992. A taxonomic revision of the palaearctic members of the ant subgenus Lasius s. str. (Hymenoptera: Formicidae). Abh. Ber. Naturkundemus. Görlitz., 66: 1-67. doi:10.5281/zenodo.24614
Seifert B. 2018. The ants of Central and North Europe. Germany: Lutra Verlags- und Vertriebsgeseklschaft. pp. 408.
Sparagano O.A.E., George D.R., Harrington D.W.J., Giangaspero A. 2014. Significance and control of the poultry red mite, Dermanyssus gallinae. Annu. Rev. Entomol., 59: 447-466. doi:10.1146/annurev-ento-011613-162101
Taylor M.A., Coop R.L., Wall R.L. 2016. Veterinary Parasitology. 4th edition. Wiley Blackwell Publishing, Oxford, UK, p.1032.
Tomberlin J.K., Drees B. 2007. Poultry Pest Management. Texas A&M Agrilife Extention. E-445.
Tomley F.M., Sparagano O. 2018. Spotlight on avian pathology: red mite, a serious emergent problem in layer hens. Avian Pathol., 47: 533-535. doi:10.1080/03079457.2018.1490493



2020-02-24
Date accepted:
2021-01-27
Date published:
2021-02-05
Edited by:
Roy, Lise

This work is licensed under a Creative Commons Attribution 4.0 International License
2021 Kar, Sirri; Akyildiz, Gurkan; Sirin, Deniz; Rodriguez, Sergio E. and Camlitepe, Yilmaz
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)