



The first report on the oribatid mites (Acari: Oribatida) in tundra of the Chunatundra Mountains on the Kola Peninsula, Russia
Leonov, Vladislav D.1
1A.N. Severtsov Institute of Ecology and Evolution RAS, 119071, Russia.
2020 - Volume: 60 Issue: 4 pages: 722-734
https://doi.org/10.24349/acarologia/20204398Original research
Keywords
Abstract
Introduction
During previous investigations of the tundra communities on the Kola Peninsula, a high number of oribatid mite species was found. Until now, 89 species from 47 genera and 27 families were known for the mountain tundra of the Kola Peninsula (Liskovaya 2011; Zenkova and Melekhina 2014; Leonov and Rakhleeva 2015; Leonov et al. 2015; Leonov and Rakhleeva 2020). However, compared to more investigated regions, the known number of oribatid species in the mountain tundra of the Kola Peninsula is still far from expected. In the Scandinavian Peninsula, 145 species, 69 genera, and 42 families of oribatid mites were found in the alpine and arctic/alpine habitats (Heggen 2010), 123 oribatid species belonging to 39 families were found in the treeless habitats of Taimyr Peninsula (Makarova 2015), and 151 species, 83 genera, 46 families were found in the western North American Low Arctic (Behan-Pelletier 1999). In addition, the known fauna of the oribatid mites in the mountain tundra habitats of the Kola Peninsula comprises significantly fewer species than the fauna of oribatid mites in plain tundra on the Kola Peninsula, which includes140 species belonging to 73 genera and 39 families (Krivolutsky 1966; Liskovaya 2011; Leonov and Rakhleeva 2015). Tundra is a diverse complex of communities that includes different types of habitats, development of which depends on many environmental factors — temperature, character of snow cover, etc. (Chernov and Matveyeva 1997; Körner 2003; Nagy and Grabherr 2009). Even minimal changes in the plant communities in tundra may lead to extremely great changes in the oribatid mite communities (Minor et al. 2016a; Seniczak and Plichta 1978; Seniczak et al. 2014). Thus, the investigation of the different types of tundra habitats is important for a better understanding of the structure and complexity of the tundra biome.
The aims of this research were to obtain data on the fauna and abundance of the oribatid mites from the main types of the mountain tundra habitats of the Chunatundra Mountains and to compare explored habitats with each other and with the literature data.
Materials and methods
Study area
The Chunatundra Mountains are located in the central part of the Kola Peninsula, west of the Khibiny Mountains (Figure 1). The average annual air temperature is -1 °C. The region belongs to the area with a predominance of precipitation over evaporation. In the mountains, precipitation amounts 600–800 mm per year and exceeds 1 000 mm at the peaks of the largest mountains. The climate type is subarctic marine (Agarkova et al. 2008).

In the mountains of the Kola Peninsula, the following vegetation zones are present: the spruce-pine-birch belt occupies areas up to 250–480 m a.s.l.; above this, the belt of crooked birch forest spreads over several tens of meters; the belt of dwarf shrubs extends up to 600–700 m a.s.l.; the alpine lichen tundra belt occurs up to 850–900 m a.s.l.; and the belt of sparse vegetation (represented by small shrubs and lichens) occupies the tops of the mountains and plateaus (Stanyukovich 1973).
Study sites
Three tundra plots and one sphagnum bog within the belt of the mountain tundra were studied. The coordinates and altitudes of the explored habitats were obtained using a GPS/GLONASS receiver Garmin eTrex 30.
1. Lichen tundra plot at an altitude of 638 m a.s.l. (ChT-LT-638; 67°41.015' N, 32°32.579' E) is located on the flat plateau of the mountain. Vegetation covers about 50% and is developed on the rocks and in the cavities between them. Flavocetraria nivalis dominates in vegetation cover. Cetraria islandica is numerous. Empetrum hermaphroditum, Vaccinium vitis-idaea, Arctostaphylos uva-ursi grow within lichen thalluses and do not exceed lichens in height. There are clumps of Juncus trifidus and single representatives of Carex sp. The soil (hereinafter, the names of the soils are given using WRB (IUSS Working Group WRB 2015)) is Entic Podzol (Skeletic).
2. Lichen tundra plot at an altitude of 466 m a.s.l. (ChT-LT-466; 67°39.662' N, 32°35.720' E). A closed cover of Flavocetraria nivalis is developed with rare patches of Cetraria islandica and Alectoria ochroleuca. Dwarf shrubs Vaccinium vitis-idaea, Phyllodoce caerulea, Empetrum hermaphroditum and shrub Betula nana grow only within lichen thalluses and do not exceed them in height. Betula nana occurs sporadically and sometimes formed thick pillow-like thickets. Representatives of Festuca ovina are rare. The soil is Entic Podzol.
3. Dwarf shrub tundra plot at an altitude of 419 m a.s.l. (ChT-DST-419; 67°39.725' N, 32°36.050' E). The dwarf shrub cover is extremely dense and forms a multi-layer system. Betula nana and Salix lapponum dominate in the upper layer, Vaccinium myrtillus grows under them. The lower layer is represented by mosses and lichens penetrated by Empetrum hermaphroditum. The vegetation of this habitat includes lichens (Cetraria islandica, Cladonia gracilis), mosses (Pleurozium schreberi), grasses (Festuca ovina, Cornus suecica), dwarf shrubs (Vaccinium myrtillus, Phyllodoce caerulea, Empetrum hermaphroditum), shrubs (Betula nana, Salix lapponum) and single, small and crooked representatives of Betula sp. Soil is Histic Entic Gleyic Podzol.
4. Sphagnum bog at an altitude of 417 m a.s.l. (ChT-SB-417; 67°39.567' N, 32°36.370' E). The bog is developed within the tundra belt on a flattened side of the mountain range. The dominant vegetation is represented by a typical wetland species — Sphagnum sp. and Eriophorum sp. Also, mosses (Pleurozium schreberi, Polytrichum sp.), grasses (Carex sp., Bartsia alpine), dwarf shrubs (Vaccinium myrtillus, Empetrum hermaphroditum, Rubus chamaemorus) and shrubs (Ledum palustre, Salix sp.) grow in this habitat.
Sampling and sample processing
In each studied habitat, 10 samples were taken with a corer (25 cm\textsuperscript{2}) up to 10 cm depth (included topsoil, litter and soil-cover vegetation — mosses, lichen and dwarf shrub parts) between July 24 and July 26, 2015. The samples were placed in multilayer paper bags which were put in the cardboard boxes with cells for each sample. The samples were delivered to Moscow in three days for further processing. Microarthropods were extracted using Tullgren funnels in the laboratory without additional heating for 10 days (Potapov and Kuznetsova 2011).
Identification of mites
Adult oribatid mites were identified to the species level using the identification keys of Ghilarov (1975), Weigmann (2006) and Bayartogtokh (2010). Ptyctimous oribatid mites were identified using Niedbała (2011). Juveniles were checked, but information about the juveniles was not used in the statistical procedures. The valid species status and synonymy are based on the nomenclature of Weigmann (2006).
Data analysis
Mean values, standard deviations, and standard errors were calculated using Microsoft Office Excel 2010. Permutational MANOVA (PerMANOVA) was performed to establish a significant difference between the oribatid mite communities of the studied habitats (Anderson 2001). SIMPER procedure (similarity percentages) was used for determining which species contribute to the dissimilarity between the investigated habitats (Clarke 1993). Bray-Curtis similarity measure and the relative abundance (%) of species in samples were used for the PerMANOVA and SIMPER tests. Calculations were made in PAST 3.26b (Hammer et al. 2001).
The oribatid communities in the explored habitats were compared using multidimensional scaling and the discriminant functional analysis according to Tiunov and Scheu (2000). Based on arcsine transformed values of species relative abundance in samples, the similarity matrix was calculated using the Bray-Curtis similarity measure. The similarity matrix was analyzed using multidimensional scaling. The minimum number of meaningful dimensions was evaluated by comparing actual stress values with the theoretical exponential function of stress. A 3-dimensions solution showed the best result. The coordinates of the samples in the 3-dimensions space were used for discriminant function analysis. Two significant discriminant functions (canonical roots) were obtained. To interpret the extracted canonical roots in terms of changes in the relative abundance of oribatid species Spearman rank correlations were calculated between the canonical scores of canonical roots and relative abundance of species in the individual samples. These calculations were made using Statistica 6.0 (StatSoft, Tulsa).
Results
A total of 3 776 specimens of adult oribatid mites were collected and identified to species level, and 2 052 specimens of juveniles were identified to family level. Seventy species and one subspecies belonging to 37 genera and 24 families were found in the explored habitats (Table 1). Compared to the previous studies, 9 species and one subspecies of oribatid mites were found on the Kola Peninsula for the first time: Brachychthonius impressus, Neobrachychthonius marginatus magnus, Sellnickochthonius furcatus, Atropacarus cf. genavensis, Trimalaconothrus foveolatus, Kunstidamaeus cf. diversipilis, Suctobelbella similis, Hydrozetes lacustris, Ceratozetes parvulus, and family Mucronothridae, represented by Mucronothrus nasalis (Liskovaya 2011; Zenkova and Melekhina 2014; Leonov and Rakhleeva 2015; Leonov et al. 2015; Leonov and Rakhleeva 2020).
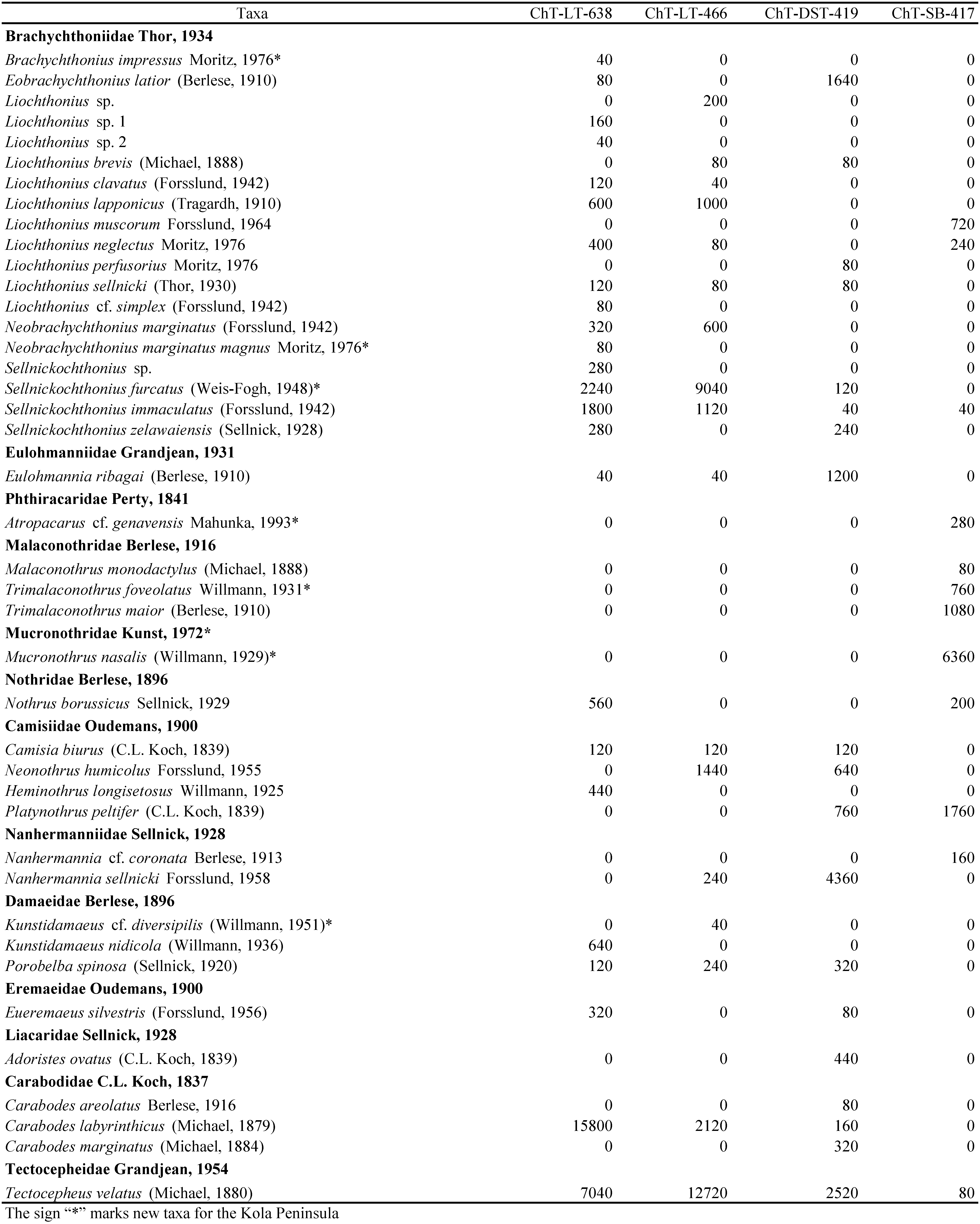


The families Brachychthoniidae, Suctobelbidae, and Oppiidae were the most diverse in the discovered local fauna. Eighteen species and one subspecies of Brachychthoniidae (26.8% of the total fauna), 9 species of Suctobelbidae (12.7% of the total fauna), and 7 species of Oppiidae (11.3% of the total fauna) were found (Table 1). The lichen and dwarf shrub tundra differed considerably in the fauna of the oribatid mites. The explored sphagnum bog was characterized by a specific fauna of oribatid mites. Mucronothrus nasalis, which was found there, is one of the most adapted species of oribatid mites to aquatic habitats (Behan-Pelletier and Eamer 2007). In this investigation, species of sphagnum bog was an important component of biodiversity: 13 species of the discovered local fauna (18%) were found only in this habitat.
Among tundra habitats, the abundance of oribatid mites was lower in the lichen tundra and increased in the dwarf shrub tundra. The sphagnum bog was characterized by the lowest abundance among all explored biotopes. Adult oribatid mites were more numerous than juveniles in the tundra habitats in this investigation (Table 1). In contrast, in the sphagnum bog, juvenile oribatid mites were more numerous than adults through the high abundance of both Malaconothridae and Mucronothridae instars (Table 2).

According to the PerMANOVA test, differences observed between assemblages of oribatid mites of all explored habitats were statistically significant (Figure 2). Based on the results of SIMPER test, they were due to the different composition of complexes of dominant species: the five most numerous species determined more than 50% of the cumulative difference between oribatid mite assemblages of the explored habitats.
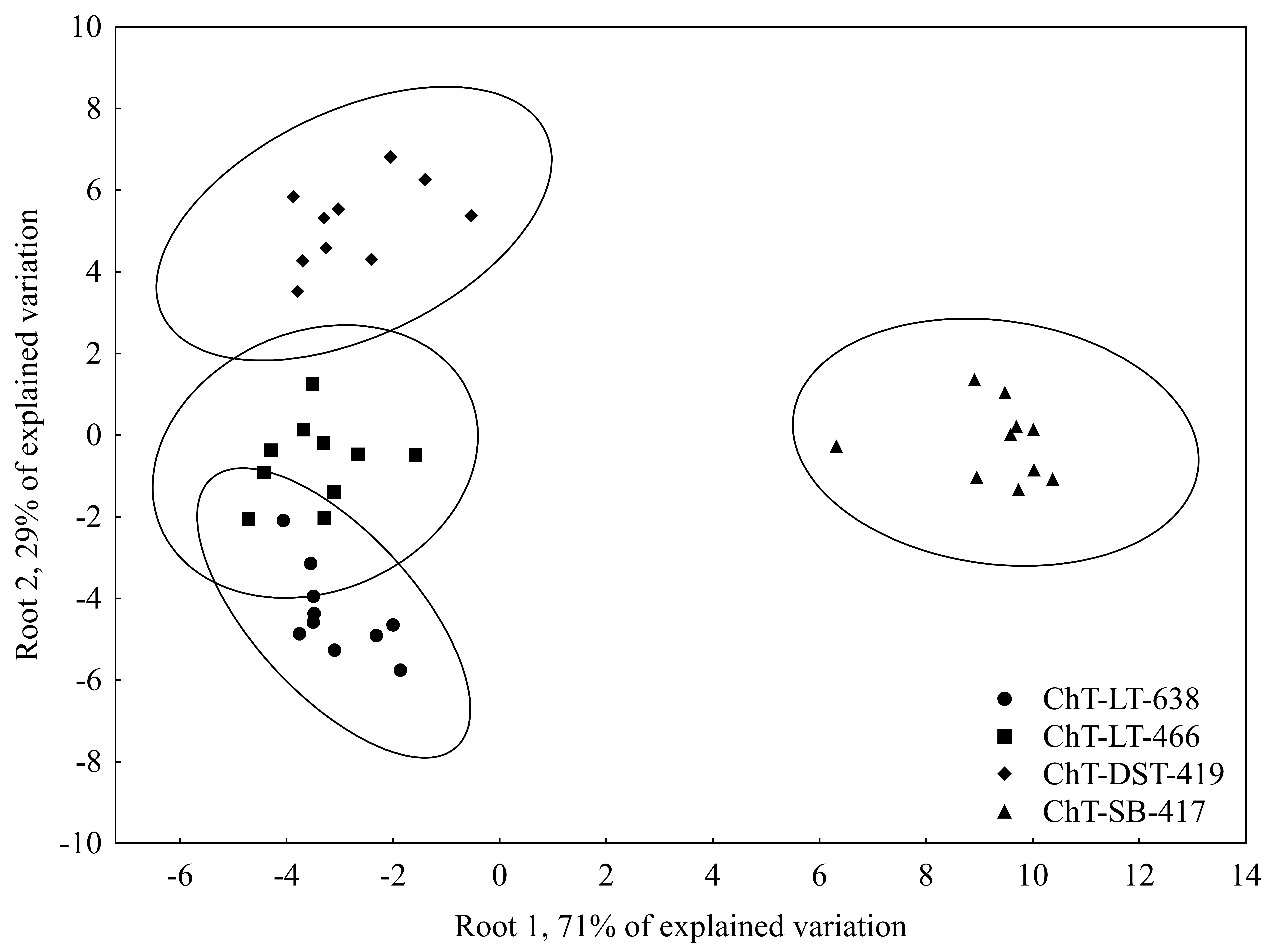
In the lichen tundra, Carabodes labyrinthicus, Tectocepheus velatus, and Sellnickochthonius furcatus dominated. The relative abundance of these species and also Nothrus borussicus, Mycobates sarekensis and six species of the family Brachychthoniidae significantly decreased from the lichen tundra to the dwarf shrub tundra (Table 3). Small-size species of the families Suctobelbidae and Oppiidae dominated in the dwarf shrub tundra. They decreased in the relative abundance from the dwarf shrub tundra to the lichen tundra, as well as species Nanhermannia sellnicki, Chamobates borealis, Eobrachychthonius latior, Hemileius initialis (and some other species). A specific complex of dominant species was found in the sphagnum bog, represented by species of wet habitats: Mucronothrus nasalis, Limnozetes ciliatus, Platynothrus peltifer, Trimalaconothrus maior, and Limnozetes cf. rugosus. The relative abundance of these oribatid species increased significantly with the highest values of the Spearman rank correlation from the tundra habitats to the sphagnum bog. Along with this, species which were the most characteristic of the lichen and dwarf shrub tundra significantly decreased in the relative abundance (Table 3).

Discussion
The discovered local oribatid mite fauna of the Chunatundra Mountains is characterized by a high diversity of the families Brachychthoniidae, Suctobelbidae, and Oppiidae (Table 1). The families Brachychthoniidae and Oppiidae often are the most diverse in the tundra communities (Ananieva et al. 1973, 1979; Thomas and MacLean Jr 1988; Grishina et al. 1998; Behan-Pelletier 1999, Makarova 2015) and polar deserts (McAlpine 1965; Bulavintsev and Babenko 1983; Makarova 2002). Families Ceratozetidae and Camisiidae, which are an important component of the faunae in the tundra communities (Thomas and MacLean Jr 1988; Sidorchuk 2009; Heggen 2010; Coulson et al. 2014; Makarova 2015), had low diversity in the present study: only four species of families Ceratozetidae and Camisiidae (each of them comprises by 5.6% of the fauna) were found. These families are most diverse in the East European tundra (Melekhina 2020).
Despite the relatively geographical closeness and some similarities in the natural conditions, the oribatid mite fauna of the Chunatundra Mountains differs from the local oribatid faunae of alpine and arctic/alpine habitats of the South, North-West and North-East Scandinavian Peninsula (Heggen 2010). In these local faunae, Camisiidae, Damaeidae, Ceratozetidae and Oppiidae are the most diverse. The families Achipteriidae and Ceratoppiidae, well represented in the alpine and arctic/alpine habitats in the Scandinavian Peninsula (Heggen 2010), were not found in this investigation.
Like in the Lovozersky Mountains (Leonov and Rakhleeva 2020), a large number of species of the family Suctobelbidae was found in the local fauna of the Chunatundra Mountains. In contrast to Chunatundra Mountains, the family Suctobelbidae has low diversity in the local faunae of the Scandinavian Peninsula (Heggen 2010). A high diversity of the family Suctobelbidae is not common for plain tundra (Makarova 2015; Melekhina and Zinovyeva 2012; MacLean et al. 1978; Thomas and MacLean Jr 1988) and mountain tundra (Sidorchuk 2009), in contrast to the taiga zone (Krivolutsky et al. 1999; Laskova 2001; Melekhina 2004). The occurrence of Suctobelbidae might be typical for communities of mountain tundra that are surrounded by mountain forests, which may be a source of the increasing diversity of the oribatid mites and particularly the family Suctobelbidae. It could also be a specific feature of the oribatid mite fauna of the Kola Peninsula which is developed under the relative mildness conditions of the oceanic type of climate. The mild conditions may lead to the relative species richness of the local faunae of oribatid mites on the Kola Peninsula.
The abundance of oribatid mites in the explored habitats (Table 1) corresponds to the values described for tundra communities in previous investigations (Thomas and MacLean Jr 1988; Melekhina and Zinovyeva 2012; Leonov and Rakhleeva 2020). In the sphagnum bog, the abundance of oribatid mites is comparatively high for such severe environmental conditions compared to some other wetland habitats located in a temperate climate (Zaitsev 2013; Lehmitz 2014; Minor et al. 2016b). Discovered values of the total (adult+juvenile) oribatid mite abundance are comparable with that in the mires of western Norway (Seniczak et al. 2010; Seniczak et al. 2019b; Solhøy 1979) but these values are lower than the density of the oribatid mites in the bogs of Poland (Seniczak et al. 2019a).
Like in the previously investigated plain and mountain tundra of the Kola Peninsula (Leonov and Rakhleeva 2015; Leonov et al. 2015; Leonov and Rakhleeva 2020), lichen and dwarf shrub tundra differ significantly in the composition of oribatid mite community. Carabodes labyrinthicus, Nothrus borussicus, Sellnickochthonius immaculatus, Mycobates sarekensis, and Tectocepheus velatus are more typical for the lichen tundra. Species of Mycobates are known as characteristic of saxicolous mosses above the upper forest line and species of Mycobates and Carabodes are closely related to the lichens (Materna 2000). Mycobates sarekensis occurs in both higher alpine zones and glacier-forelands in alpine habitats of the Scandinavian Peninsula (Heggen 2010).
In this investigation, species of Carabodes have different preferences to environmental conditions. Carabodes labyrinthicus is strongly associated with the lichen tundra, whereas two other species of Carabodes are found only in the dwarf shrub tundra, where C. labyrinthicus decreases significantly in the abundance. This trend was observed in the previous investigations of the oribatid mites in the Lovozersky Mountains, Khibiny Mountains and in the plain tundra of the Kola Peninsula (Leonov and Rakhleeva 2015; Leonov et al. 2015; Leonov and Rakhleeva 2020). Carabodes labyrinthicus was found as a permafrost indicator in sub-Arctic palsa mires (Markkula 2014; Markkula et al. 2018).
Species of the Suctobelbidae and Oppiidae are most abundant in dwarf shrub tundra. A similar pattern was observed in the previous investigations of the oribatid mites in the mountain and plain tundra of the Kola Peninsula (Leonov and Rakhleeva 2015; Leonov et al. 2015; Leonov and Rakhleeva 2020). Species of these families were more common in a mountain forest habitat than above tree-line in the Central Alps (Fischer and Schatz 2013). The Suctobelbidae and Oppiidae species were characteristic for saxicolous mosses below the upper forest-line in the Krokonoše Mountains in Czech Republic (Materna 2000).
The distribution of Nanhermannia sellnicki is similar to the results obtained in previous investigations of the tundra communities on the Kola Peninsula (Leonov and Rakhleeva 2015; Leonov et al. 2015; Leonov and Rakhleeva 2020). However, this species may show changing ecological preferences in different environmental conditions. Nanhermannia sellnicki was the most abundant species in the bogs of the western part of Norway, where the climate is mild and temperate with an average annual temperature of 6.8 °C (Seniczak et al. 2019b). Chamobates borealis, which was strongly associated with the wet conditions in the Lovozersky Mountains (Leonov and Rakhleeva 2020) and was found as a permafrost indicator in sub-Arctic palsa mires (Markkula 2014; Markkula et al. 2018), is significantly correlated with the tundra conditions and with dwarf shrub tundra in this research. This difference might be caused by different climate conditions of the comparing regions. The ecological preferences of these species require further refinement.
Acknowledgements
I thank A.A. Rakhleeva (Soil Science Faculty, Lomonosov Moscow State University) for the supervision of this investigation. I thank O.L. Makarova and A.B. Babenko (IPEE RAS) for consultations and the discussion of the results. I am very grateful to E.A. Sidorchuk (PIN RAS) for the help and teaching in the identification of the oribatid mites. The author thanks A.V. Tiunov, Corresponding Member of the RAS (A.N. Severtsov Institute of Ecology and Evolution RAS), for the advice in part of statistical processing of the data. I am very grateful to the reviewers whose advice and remarks helped to improve the manuscript. The author would like to thank the staff of the Lapland Nature Reserve for their assistance in conducting this investigation. The author thanks the Russian Foundation for Basic Research for the support of this investigation (Project No. 14-04-31754).
References
Agarkova T.V. et al. 2008. Kola Encyclopedia. Apatity: KSC RAS. pp. 600.
Ananieva S.I., Krivolutsky D.A., Chernov Yu.I. 1973. The oribatid mites (Oribatei) of subzones of typical tundra of western Taimyr. In: Tikhomiriv B.A. (Ed.) Biogeocenoses of the Taimyr tundra and their productivity. Leningrad: Nauka. p. 148–151.
Ananieva S.I., Krivolutsky D.A., Chernov Yu.I. 1979. Oribatid mites (Oribatei) in the subzone of arctic tundra in the northeast of Taimyr. In: Aleksandrova V.D., Matveeva N.V. (Eds). Arctic tundra and polar deserts of Taimyr. Leningrad: Nauka. p. 144–147.
Anderson M.J. 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecol., 26(1): 32–46. doi:10.1046/j.1442-9993.2001.01070.x
Bayartogtokh B. 2010. Oribatid Mites of Mongolia (Acari: Oribatida). Moscow: KMK Scientific Press. pp. 372.
Behan-Pelletier V.M. 1999. Oribatid mite fauna of northern ecosystems: a product of evolutionary adaptations or physiological constraints. In: Needham, Mitchell, Horn and Welbourn (Eds). Acarology IX: Proceedings of the International Congress. p. 87–105.
Behan-Pelletier V.M., Eamer B. 2007. Aquatic Oribatida: adaptations, constraints, distribution and ecology. In: Acarology XI: Proceedings of the International Congress. p. 71–82.
Bulavintsev V.I., Babenko A.B. 1983. Soil-dwelling invertebrates in the eastern sector of Franz Josef Land. Zool. Zh., 62(7): 1114–1116.
Chernov Yu.I., Matveyeva N.V. 1997. Arctic ecosystems in Russia. In: Wielgolaski F.E. (Ed.). Ecosystems of the World. 3. Polar and Alpine Tundra. Amsterdam: Elsevier Science B.V. p. 361–507.
Clarke K.R. 1993. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol., 18(1): 117–143. doi:10.1111/j.1442-9993.1993.tb00438.x
Coulson S.J., Convey P., Aakra K. et al. 2014. The terrestrial and freshwater invertebrate biodiversity of the archipelagoes of the Barents Sea; Svalbard, Franz Josef Land and Novaya Zemlya. Soil. Biol. Biochem., 68: 440–470. doi:10.1016/j.soilbio.2013.10.006
Fischer B.M., Schatz H. 2013. Biodiversity of oribatid mites (Acari: Oribatida) along an altitudinal gradient in the Central Alps. Zootaxa, 3626(4): 429–454. doi:10.11646/zootaxa.3626.4.2
Ghilarov M.S. 1975. The keys for identification of the soil-dwelling mites. Sarcoptiformes. Moscow: Nauka. pp. 491.
Grishina L.G., Babenko A.B., Chernov Yu.I. 1998. The Oribatid Mites (Sarcoptiformes, Oribatei) of Taimyr Peninsula Western Coast. Zoodiversity, 32(1-2): 116–118.
Hammer Ø., Harper D.A.T., Ryan P.D. 2001. Paleontological Statistics Software: Package for Education and Data Analysis. Palaeontol. Electron, 4(1): 1–9.
Heggen M.P. 2010. Oribatid mites of Alpine Fennoscandia. Nor. J. Entomol., 57(1): 38–70.
IUSS Working Group WRB. 2015. World Reference Base for Soil Resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. Rome: FAO. pp. 191.
Körner C. 2003. Alpine plant life: functional plant ecology of high mountain ecosystems. Berlin, Heidelberg: Springer. pp. 349. doi:10.1007/978-3-642-18970-8
Krivolutsky D.A. 1966. The oribatid mites in the tundra soils. Pedobiologia, 6(3): 277–280.
Krivolutsky D.A., Zaitsev A.S., Laskova L.M. 1999. Geography of the oribatid mite biodiversity in the European North of Russia. Petrozavodsk: KarRC RAS. pp. 36.
Laskova L.M. 2001. Biodiversity of oribatid mites of Karelia. Biogeography of Karelia. Ser. Biology, 2: 125–132.
Lehmitz R. 2014. The oribatid mite community of a German peatland in 1987 and 2012 — effects of anthropogenic desiccation and afforestation. Soil. Org., 86(2): 131–145.
Leonov V.D., Rakhleeva A.A. 2015. New information on similarity and difference between highland and plain tundra of the Kola Peninsula based on oribatid mite data (Acari: Oribatida). Euroasian Entomol. J., 14(5): 489–499.
Leonov V.D., Rakhleeva A.A., Sidorchuk E.A. 2015. Distribution of Oribatides Mites (Acari: Oribatida) along an Altitudinal Profile of Mount Vud'yavrchorr (the Khibiny Mountains). Eurasian Soil Sci., 48(11): 1257–1267. doi:10.1134/S1064229315110101
Leonov V.D., Rakhleeva A.A. 2020. The first report on oribatid mites in tundra belts of the Lovozersky Mountains on the Kola Peninsula, Russia. Acarologia, 60(2): 301–316.
Liskovaya A.A. 2011. Faunal and ecological diversity of oribatid mites (Acariformes: Oribatei) in the ecosystems of the Kola North [PhD Thesis]. Petrozavodsk: Petrozavodsk State University. pp. 178.
MacLean S.F., Behan V., Fjellberg A. 1978. Soil Acari and Collembola from Chaun Bay, Northern Chukotka. Arct. Alp. Res., 10(3): 559–568. doi:10.2307/1550679
Makarova O.L. 2002. Acarocenoses (Acariformes, Parasitiformes) in polar deserts: 1. Mite assemblages of the Severnaya Zemlya Archipelago: structure of fauna and abundance. Entomol. Rev., 82(7): 839–856.
Makarova O.L. 2015. The fauna of free-living mites (Acari) of Greenland. Entomol. Rev., 95(1): 108-125. doi:10.1134/S0013873815010133
Markkula I. 2014. Permafrost dynamics structure species compositions of oribatid mite (Acari: Oribatida) communities in sub-Arctic palsa mires. Polar Res., 33(1): 22926. doi:10.3402/polar.v33.22926
Markkula I., Oksanen P., Kuhry P. 2018. Indicator value of oribatid mites in determining past permafrost dynamics in northern European sub-Arctic peatlands. Boreas, 47(3): 884–896. doi:10.1111/bor.12312
Materna J. 2000. Oribatid communities (Acari: Oribatida) inhabiting saxicolous mosses and lichens in the Krkonoše Mts. (Czech Republic). Pedobiologia, 44(1): 40–62. doi:10.1078/S0031-4056(04)70027-X
McAlpine J.F. 1965. Insects and related terrestrial invertebrates of Ellef Ringnes Island. Arctic, 18: 73–103. doi:10.14430/arctic3455
Melekhina E.N. 2004. Faunistic diversity of oribatid mites (Oribatida) of the taiga zone of the Komi Republic. Vestnik Insituta biologii Komi NC UrO RAN, 5: 6-9.
Melekhina E.N. 2020. Analysis of oribatid fauna of the east european tundra with first reported data of Subpolar Urals. Divers., 12(6): 235. doi:10.3390/d12060235
Melekhina E.N., Zinovyeva A.N. 2012. First data on oribatid mites (Acari: Oribatida) of the Pay-Khoy Ridge (Yugor Peninsula). Proc. Komi Sci. Cent. Ural Division of the Russ. Acad. Sci., 2(10): 42–50.
Minor M.A., Babenko A.B., Ermilov S.G., Khaustov A.A., Makarova O.L. 2016a. Effects of cushion plants on high-altitude soil microarthropod communities: cushions increase abundance and diversity of mites (Acari), but not springtails (Collembola). Arct. Antarct. Alp. Res., 48(3): 485-500. doi:10.1657/AAAR0015-064
Minor M.A., Ermilov S.G., Philippov D.A., Prokin A.A. 2016b. Relative importance of local habitat complexity and regional factors for assemblages of oribatid mites (Acari: Oribatida) in Sphagnum peat bogs. Exp. Appl. Acarol., 70(3): 275–286. doi:10.1007/s10493-016-0075-9
Nagy L., Grabherr G. 2009. The biology of alpine habitats. Oxford, New York: Oxford University Press on Demand. pp 336.
Niedbała W., 2011. Ptyctimous mites (Acari, Oribatida) of the Palaearctic Region. Systematic part. Warszawa: Natura optima dux Foundation, 472 p.
Potapov M.B., Kuznetsova N.A. 2011. Methods of research of microarthropods communities. Moscow: KMK Scientific Press. pp. 88.
Seniczak A., Seniczak S., Graczyk R., Waldon-Rudzionek B., Nowicka A., Pacek, S. 2019a. Seasonal Dynamics of Oribatid Mites (Acari, Oribatida) in a Bog in Poland. Wetlands, 39(4): 853–864. doi:10.1007/s13157-019-01125-2
Seniczak A., Seniczak S., Iturrondobeitia J.C., Solhøy T., Flatberg K.I. 2019b. Diverse Sphagnum Mosses Support Rich Moss Mite Communities (Acari, Oribatida) in Mires of Western Norway. Wetlands. doi:10.1007/s13157-019-01236-w
Seniczak A., Solhøy T., Seniczak S., Riva-Caballero, A.D.L. 2010. Species composition and abundance of the oribatid fauna (Acari, Oribatida) at two lakes in the Fløyen area, Bergen, Norway. Biol. Lett., 47(1): 11–19. doi:10.2478/v10120-009-0014-0
Seniczak S., Plichta W. 1978. Structural dependence of moss mite populations (Acari, Oribatei) on patchiness of vegetation in moss lichen tundra at the north coast of Hornsund, West Spitsbergen). Pedobiologia, 18(8): 145–152.
Seniczak S., Seniczak A., Gwiazdowicz D.J., Coulson S.J. 2014. Community structure of oribatid and gamasid mites (Acari) in moss-grass tundra in Svalbard (Spitsbergen, Norway). Arct. Antarct. Alp. Res., 46(3): 591–599. doi:10.1657/1938-4246-46.3.591
Sidorchuk E.A. 2009. New data on the fauna of oribatid mites (Acari, Oribatida) from the Polar Urals. Entmol. Rev., 89: 554–563. doi:10.1134/S0013873809050054
Solhøy T. 1979. Oribatids (Acari) from an oligotrophic bog in western Norway. Fauna Norv. Ser. B., 26: 91–94.
Stanyukovich K.V. 1973. Vegetation of the USSR mountains (botanical and geographical sketch). Dushanbe: Donish. pp. 416.
Thomas R.H., MacLean Jr S.F. 1988. Community structure in soil Acari along a latitudinal transect of tundra sites in northern Alaska. Pedobiologia, 31: 113–138.
Tiunov A.V., Scheu S. 2000. Microfungal communities in soil, litter and casts of Lumbricus terrestris L. (Lumbricidae): a laboratory experiment. Appl. Soil Ecol., 14(1): 17–26. doi:10.1016/S0929-1393(99)00050-5
Weigmann G. 2006. Hornmilben (Oribatida): Acari, Actinochaetida. Keltern: Goecke & Evers. pp. 520.
Zaitsev A.S. 2013. Oribatid mite communities (Acari: Oribatida) in different habitats of the Polistovsky Nature Reserve (Pskov Region, Russia). Estonian J. Ecol., 62(4): 276–286. doi:10.3176/eco.2013.4.04
Zenkova I.V., Melekhina E.N. 2014. The Oribatid mites (Acari: Oribatida) of the Khibiny Mountains. In: Evdokimova G.A., Vandysh O.I. (Eds). Environmental problems of the northern regions and ways of their solution. Apatity: Kola Science Center of the Russian Academy of Sciences. p. 135–140.



2020-06-28
Date accepted:
2020-10-02
Date published:
2020-10-05
Edited by:
Baumann, Julia

This work is licensed under a Creative Commons Attribution 4.0 International License
2020 Leonov, Vladislav D.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)