



Distribution and biological features of Typhlodromus (Anthoseius) recki (Acari: Phytoseiidae) on Tetranychus urticae, T. evansi (Acari: Tetranychidae) and Aculops lycopersici (Acari: Eriophyidae)
Tixier, Marie Stephane  1
; Douin, Martial2
; Rocio, Oliva3
; Gonzalez, Lucia4
; Pount, Benjamin5
and Kreiter, Serge
6
1
; Douin, Martial2
; Rocio, Oliva3
; Gonzalez, Lucia4
; Pount, Benjamin5
and Kreiter, Serge
6
1✉ CBGP, Institut Agro, INRA, CIRAD, IRD, Univ. Montpellier, France, 755 Avenue du Campus Agropolis, CS 30016, 34988 Montferrier-sur-Lez cedex, France.
2CBGP, Institut Agro, INRA, CIRAD, IRD, Univ. Montpellier, France, 755 Avenue du Campus Agropolis, CS 30016, 34988 Montferrier-sur-Lez cedex, France.
3Universidad de Almería, Carretera Sacramento s/n, 04120 La Cañada de San Urbano, Almería, Spain.
4CBGP, Institut Agro, INRA, CIRAD, IRD, Univ. Montpellier, France, 755 Avenue du Campus Agropolis, CS 30016, 34988 Montferrier-sur-Lez cedex, France.
5CBGP, Institut Agro, INRA, CIRAD, IRD, Univ. Montpellier, France, 755 Avenue du Campus Agropolis, CS 30016, 34988 Montferrier-sur-Lez cedex, France.
6CBGP, Institut Agro, INRA, CIRAD, IRD, Univ. Montpellier, France, 755 Avenue du Campus Agropolis, CS 30016, 34988 Montferrier-sur-Lez cedex, France.
2020 - Volume: 60 Issue: 4 pages: 684-697
https://doi.org/10.24349/acarologia/20204396Original research
Keywords
Abstract
Introduction
Mites of the family Phytoseiidae are predators, and some species are used for controlling pest mites and small insects, in various crops all over the world (Gerson et al. 2003; McMurtry et al. 2013). This family contains 2,521 valid species (Demite et al. 2020) but only a few are presently used in biological control, probably because of little knowledge on the biological features of most species. We thus hypothesise that much more species than the 20-30 species presently known for their predatory efficiency could be exploited for biological control issues (Van Lenteren 2012; Van Lenteren et al. 2017). Phytoseiidae mites are naturally occurring in crops and uncultivated areas (Moraes et al. 1986). Most Phytoseiidae species are generalist predators (McMutry & Croft 1997; McMurtry et al. 2013); they feed on prey and also on pollen, plant exudates, sometimes plant tissues, fungi, etc. Their occurrence and diversity are strongly affected by plant phylloplane features (Karban et al. 1995; Kreiter et al. 2002; Schmidt 2004; Tixier 2018). Tomato plants have specific morphological and chemical characteristics that can act as a selective filter on Phytoseiidae communities. Trichomes on tomato leaves and stems hinder mite dispersal and development, Phytoseiidae get stuck on the sticky trichomes and die (Kennedy 2003; Simmons and Gurr 2005; Sato et al. 2011; Van Houten et al. 2013a, b). Alkaloids contained in tomato and ingested by prey adapted to this plant, can also be toxic for non-adapted predators (Koller et al. 2007; Ferrero et al. 2014a, b). However, some studies report the presence of Phytoseiidae on tomatoes (i.e. Moraes et al. 1986; Tixier et al. 2020). Studies on Phytoseiulus longipes Evans show that strains of this species collected on Solanaceae in Argentina and Brazil are adapted to tomato and can control both Tetranychus urticae (Koch) and T. evansi Baker & Pritchard on this crop (Furtado et al. 2007; Ferrero et al. 2014a, b). However, as P. longipes is endemic to the Neotropical region, the introduction of this species in Europe is confronted with regulatory and environmental problems. In a recent study, we showed that Typhlodromus (Anthoseius) recki Wainstein is one of the species most frequently reported on Solanaceae in Europe (Tixier et al. 2020). It is also observed on plants with high trichome density, or presenting spiny and sticky trichomes (especially on plants of the families Lamiaceae and Boraginacae) (Moraes et al. 1986; Swirski and Amitai 1997; Tixier et al. 2020). Little is known on the biology of T. (A.) recki; it has been reported on tomatoes, feeding on mites and thrips, in Turkey and Iran (Hajizadeh et al. 2006; Kumral and Cobanoglu 2015a, b). In the present study, a series of experiments was carried out to characterise the feeding and dispersal habits of T. (A.) recki. Furthermore, its current distribution was also analysed.
Materials and methods
Distribution of Typhlodromus (Anthoseius) recki
Information on the geographic distribution and plant occurrence was retrieved from 59 publications (from 1958 to 2018) reporting T. (A.) recki occurrence (database available on request).
Plant Nomenclature
We followed the Angiosperm Phylogeny Group III's nomenclature (2009) for family, genus and species names.
Feeding habits of Typhlodromus (Anthoseius) recki in the laboratory
The aim was to characterise the ability of T. (A.) recki to feed on three prey species. Three populations of T. (A.) recki collected in France on Solanaceae and Lamiaceae were considered (Table 1). Two ''control'' species, commonly used in biological control (Knapp et al. 2018), both collected on Solanaceae in France (Table 1), were also studied: Neoseiulus californicus (McGregor) and N. cucumeris (Oudemans). The predatory mites were maintained in rearing units, as described by McMurtry and Scriven (1965). Pollen of Typha sp. was added daily as food source.


Feeding ability was assessed on three prey species: (i) T. evansi collected on tomato, Solanum lycopersicum L., in a greenhouse at Saint-Jeannet (Alpes-Maritimes, South-East France) in October 2007 and then reared in the laboratory on Solanum nigrum L., (ii) T. urticae collected in Montpellier in 2010 and maintained since then on Phaseolus vulgaris L. (var. Contender) in a greenhouse and (iii) Aculops lycopersici (Tryon) sent by Koppert BV (Netherlands) in 2017 and then reared on entire plants of S. nigrum in the laboratory.
Three treatments were tested: (i) T. urticae on bean leaf discs, (ii) T. evansi on S. nigrum leaf discs and (iii) A. lycopersici on S. nigrum leaf discs. As T. urticae does not develop well on S. nigrum, it was impossible to carry out the tests on this latter plant. Three tetranychid females (T. evansi or T. urticae) were placed on a leaf disc (2.5 cm in diameter) of S. nigrum or P. vulgaris var. Contender, disposed on humid cotton in a Petri dish (10 cm in diameter) to avoid mite escape. After two days, the Tetranychidae females were removed, the number of eggs laid was counted and a predatory female (young gravid female issued from the rearing units) introduced in the experimental arena. The experimental conditions were 25 °C, 70% relative humidity and 16D: 8N photoperiod. The number of prey and predator eggs was counted each day during four days as a rapid test to screen the predator efficiency.
For A. lycopersici experiments, S. nigrum leaf discs (2.5 cm in diameter), highly infested with this pest, were placed on humid cotton in a Petri dish (10 cm in diameter). One predatory female (young gravid female issued from the rearing units) was introduced and the number of eggs laid by the predator was counted daily during four days. The experimental conditions were the same as for the Tetranychidae experiments. For each prey and predator population / species treatment, 15 replicates were performed.
Analysis of variance (ANOVA) followed by mean comparison tests (Newman-Keuls) were carried out to compare the mean daily fecundity and mean daily egg consumption of the populations / species considered for each food treatment. The analyses were performed using Statistica (Statsoft France, 2010).
Dispersal abilities of Typhlodromus (Anthoseius) recki in the laboratory
The experiment aims at determining ''in vitro'' conditions how T. (A.) recki disperses along tomato stems. The population of T. (A.) recki used was that collected in Saint-Guilhem-le-Désert (Hérault, South of France) on Mentha suaveolens Ehrhart (Table 1). This population was chosen because of its higher fecundity compared to the other strains, when fed on A. lycopersici. The experimental design consisted of a detached tomato stem (var. Marmande) (10 cm in length) placed in a vertical position within a box filled with agar (Figure 1). Agar was maintained moistened (wet cotton strips placed around the agar layer) to avoid mite escape and keep the leaf discs and the stem turgescent. A leaf tomato disc with pollen or A. lycopersici was placed at one extremity of the stem, whereas a tomato leaf disc without food was placed at the other extremity. Shelters (wool yarns and cardboard) were disposed on the leaf discs, as nests where Phytoseiidae use to lay eggs. The boxes were turned each day to avoid phototropism effects. Fifteen replicates were carried out for each treatment (pollen or A. lycopersici). For each replication, one young gravid female of T. (A.) recki (issued from rearing units) was placed in the middle of the stem. The position of the female, the number of eggs and immatures on each leaf disc was counted for seven days. Student t-tests were carried out to compare the mean number of eggs laid on leaf discs with and without food.

Assessment of the ability of T. (A.) recki to disperse from mint to tomato under laboratory conditions
In a recent survey, high densities of T. (A.) recki were observed on M. suaveolens (Tixier et al. 2020). Thus, we wondered about the use of this plant as a reservoir (banker plant) for T. (A.) recki. This experiment aims at determining under laboratory conditions if T. (A.) recki would disperse from mint to tomato. The population used was the same as that considered for the stem dispersal experiment (Saint Guilhem-Le-Désert). The mint leaves were collected at Saint Guilhem-Le-Désert. Five females of T. evansi were deposited for three days on a tomato leaf disc (var. Marmande, 2.5 cm in diameter). The tomato leaf disc was cut into two equal parts. Each tomato half-disc was placed adjacently to a half-leaf disc of mint (same dimension as the tomato half-disc) containing two females of T. (A.) recki. The mint leaf discs were checked to avoid prey presence. The number of T. evansi (females, eggs and immatures) and the number of Phytoseiidae (females, eggs and immatures) on each half-disc were daily counted during 7 days and 15 replications were performed. Student t-tests were carried out to compare the number of T. evansi and T. (A.) recki on the two half discs through the time. A Chi2 test was carried out to compare the percentages of females, eggs and eggs+immatures on each half-disc, through the time. The analyses were performed using Statistica (Statsoft France 2010).
Results and discussion
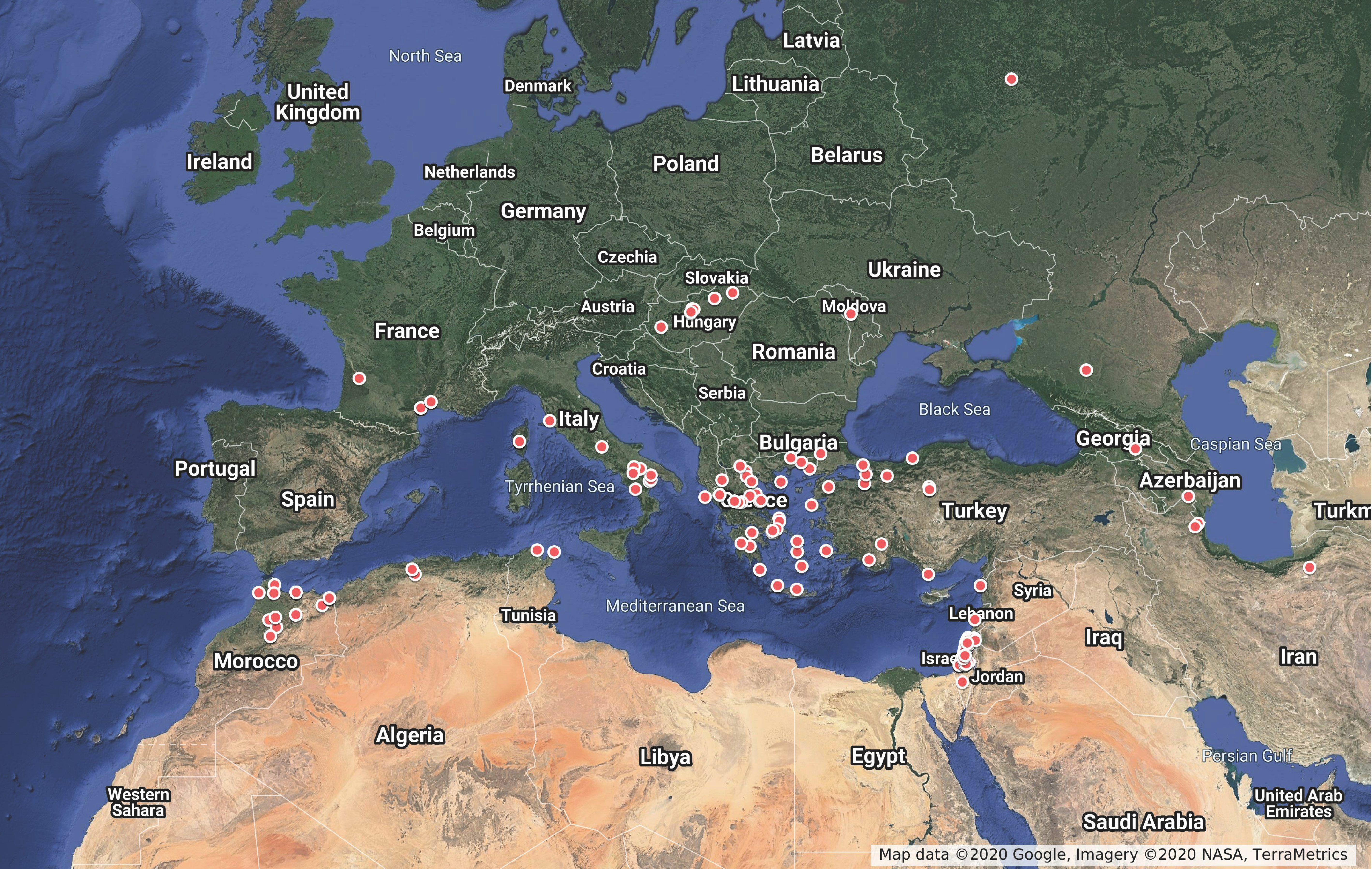
Distribution of Typhlodromus (Anthoseius) recki
Typhlodromus (A.) recki is reported from 22 countries, mainly in the Southern West-Palearctic region (Figure 2) under Mediterranean climate. It is reported 421 times on 213 plant species, belonging to 138 genera and 51 families. It is recorded less than 5 times on 37 plant families, and the highest record rates were observed on six plant families (56% of reports): Asteraceae (61 reports), Lamiaceae (50 reports), Rosaceae (40 reports), Boraginaceae (36 reports), Fabaceae (26 reports) and Solanaceae (25 reports). This might suggest close relationships between this species and those plants. However, because of no close phylogenetic relationship among these plant families, such occurrence seems not related to evolutionary aspects, as also stated by Tixier (2018) and Tixier et al. (2020). Typhlodromus (A.) recki is reported less than 5 times on 123 plant genera. The highest number of reports (19% of total reports) is observed on species of the genera Solanum (19 reports), Echium (17 reports), Vitis (14 reports), Prunus (12 reports) and Salvia (11 reports). Again, these genera are not phylogenetically close, and T. (A.) recki occurrence seems not related to plant support evolution. Those data provide elements on the ''probability'' to retrieve T. (A.) recki on some plant species, although we are aware that the recorded occurrences do not exactly reflect the real distribution.
Feeding habits of Typhlodromus (Anthoseius) recki in the laboratory
Tetranychus urticae and bean
The average number of eggs laid over 4 days by females of T. (A.) recki, N. cucumeris and N. californicus was significantly different (P = 0.0008) (Table 2). This was due to the significantly higher fecundity of N. californicus (1.4 eggs/ day/ female) as no difference was observed between the three populations of T. (A.) recki and N. cucumeris. Those differences were not significant at days 1 (P=0.07) and 2 (P=0.06) but were significant at days 3 (P<0.001) and 4 (P=0.001) (Table 4, Figure 3a). For days 3 and 4, the fecundity of N. californicus was significantly higher than that of N. cucumeris and T. (A.) recki. Differences in fecundity through the time were only observed for N. californicus, a higher fecundity being observed at days 3 and 4 than at days 1 and 2 (Table 4). Even if not significant, we can note that among the T. (A.) recki populations, the highest daily fecundity was observed for the population collected on Datura stramonium L. (0.95 eggs/ day/ female) and the lowest for the population collected on S. lycopersicum L. (0.57 eggs/ day/ female) (Table 2).

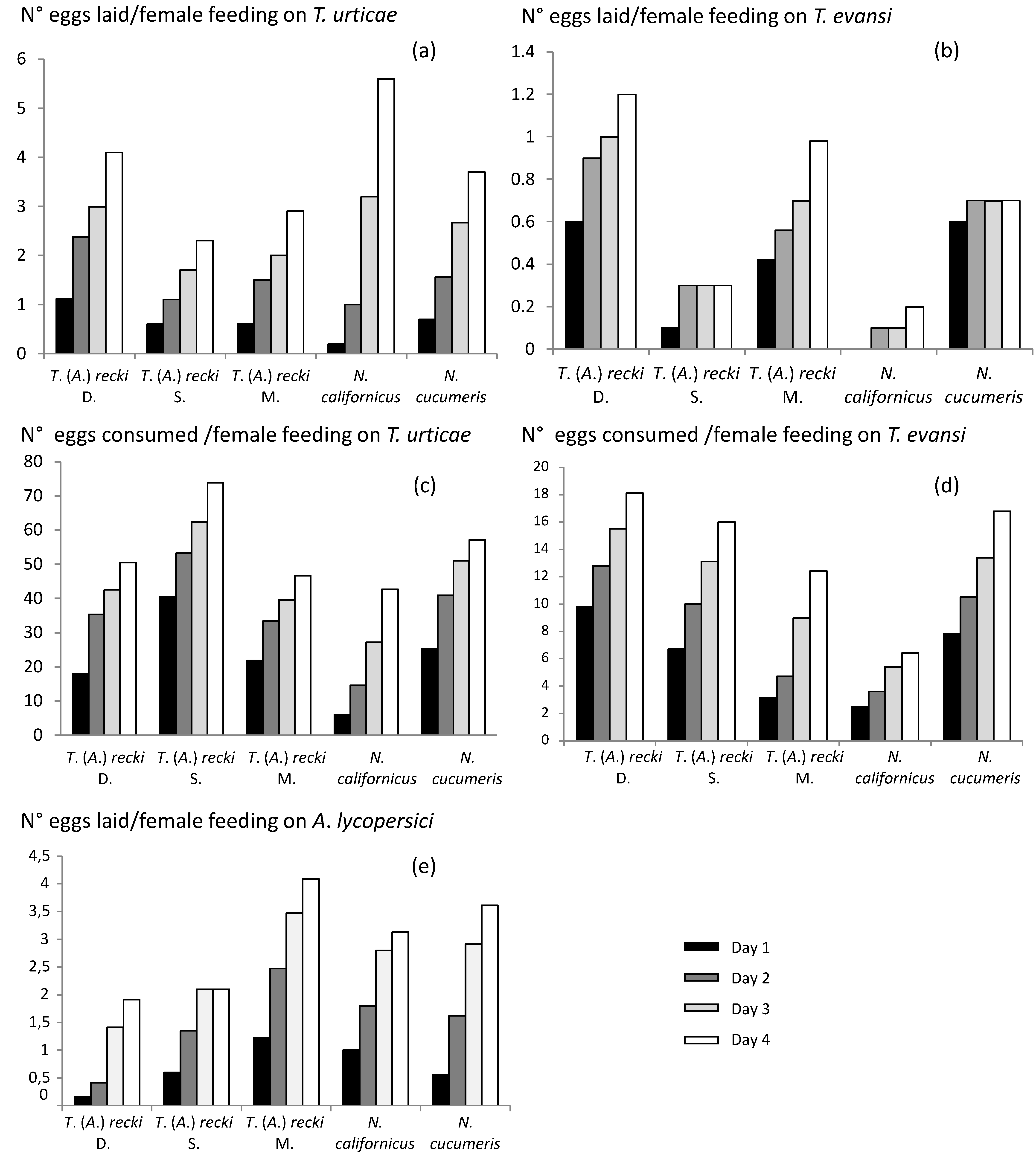
The number of T. urticae eggs consumed by the populations/species was significantly different (P = 0.02) (Table 3). The highest mean consumption was observed for T. (A.) recki collected on S. lycopersicum (18.47 eggs consumed / day) and the lowest for N. californicus (10.67 eggs consumed / day) and T. (A.) recki collected on M. suaveolens (11.69 eggs consumed / day). The numbers of eggs consumed were significantly different at day 1 (P<0.001), day 3 (P=0.05) and day 4 (P<0.001) (Table 4). At day 1, T. (A.) recki from S. lycopersicum showed the highest consumption (40.5 eggs consumed) and N. californicus the lowest (6.0 eggs consumed). At day 3 and 4, the highest values were observed for N. californicus (Table 4). Egg consumption significantly decreased from day 1 to day 4, except for N. californicus for which they increased (Table 4). The mean number of eggs consumed during 4 days was the highest (73.9) for T. (A.) recki from S. lycopersicum. For the other populations of T. (A.) recki, this consumption was about 50 eggs (Figure 3c).

There are no data on biological features of T. (A.) recki in literature. Differences between the three populations were observed but they were relatively low, except for the population collected on S. lycopersicum that consumed significantly more T. urticae eggs than the two others. Globally, T. (A.) recki has low transformation rates: many eggs of T. urticae need to be consumed to produce one Phytoseiidae egg (especially for the population collected on S. lycopersicum). These results are in accordance with biological features (observed in experimental conditions comparable to the present ones: 18 °C-25%C, 65%-75% HR and 16D:8N - 18D:6N) of other generalist Phytoseiidae species. For instance, Kasap (2010) showed for Kampimodromus aberrans (Oudemans), a low consumption rate (2.85 eggs / day) of T. urticae and a mean fecundity of one egg per day. Zemek (1993) indicated a low fecundity for Typhlodromus (Typhlodromus) pyri Scheuten when fed with T. urticae (0.72 eggs / day). Basheer et al. (2014) observed for Typhlodromus (Typhlodromus) athiasae Porath & Swirski, a daily consumption of 10 T. urticae eggs and a mean fecundity of 1.42 eggs/day. The consumption and oviposition rates reported in the literature for N. californicus are variable depending on the populations considered and the environmental conditions. Globally, the mean fecundity is about 2 eggs/female/day (Gotoh et al. 2004; Lebdi-Grissa et al. 2005; Canlas et al. 2006; Rhamani et al. 2009; Marafeli et al. 2014), but lower values were observed by Hatherly et al. (2005) (0.62 eggs/day) and Castagnoli et al. (2001) (1.46 eggs / day). The fecundity here observed (1.61 eggs/day) show an intermediary value; such differences might be due to strain effects (Castagnoli et al. 2001). Regarding T. urticae egg consumption, the present results (10.7 eggs consumed/ female/ day) are similar to those obtained for several strains of N. californicus by Castagnoli et al. (2001) (9.09 - 11.41) and a bit lower than the data obtained by Canlas et al. (2006). For N. cucumeris, the fecundity here observed is much lower than that reported by Al-Azzazy et al. (2018). The daily T. urticae egg consumption (14.52) is in accordance with values obtained by Li & Zhang (2016) and Zheng et al. (2017) and higher than those reported by Al-Azzazy et al. (2018).

Tetranychus evansi on Solanum nigrum
The number of eggs laid by the three populations of T. (A.) recki, N. cucumeris and N. californicus was significantly different (P = 0.016) (Table 2). The highest fecundity was observed for T. (A.) recki from D. stramonium L. (0.3 eggs / female/ day) and the lowest for N. californicus (0.05 eggs / female/ day) (Table 2). Significant differences were only observed at day 1 (P=0.01) (Table 4, figure 3b), where the highest fecundity was observed for T. (A.) recki from D. stramonium and N. cucumeris and the lowest for N. californicus. For all the species, the mean fecundity decreases through the time (Table 4). The mean number of T. evansi eggs consumed per day was significantly different (P=0.01). The highest values were observed for T. (A.) recki collected on D. stramonium, T. (A.) recki collected on S. lycopersicum and N. cucumeris, and the lowest for N. californicus (Table 3). No significant difference was observed at each day (Table 4). For all species, the daily consumption significantly decreased through the time (Table 4). The cumulated number of eggs consumed during four days ranged from 6.4 (N. californicus) to 18.1 (T. [A] recki from D. stramonium) (Figure 3d). These results, as in Escudero and Ferragut (2005), Moraes & McMurtry (1985) and Koller et al. (2007), show the low mean fecundity of N. californicus when fed on T. evansi, even lower than the values obtained in these publications (0.79 eggs/ female/ day, 0.5 eggs/ female/ day and 0.56 eggs/female/day, respectively). The low mean fecundity observed for N. cucumeris and the three populations of T. (A.) recki are in accordance with the results obtained for eight Phytoseiidae species by Moraes and McMurtry (1985). For T. evansi egg consumption, data in literature are scarce. Moraes and McMurtry (1986) reported a lower mean consumption by Phytoseiulus persimilis Athias-Henriot (1.4 - 1.7 eggs/ female/day) than the one here obtained for N. cucumeris and T. (A.) recki (3.11 - 4.52 eggs/female/day).
Aculops lycopersici on Solanum nigrum
The mean fecundity of the populations / species was not significantly different (P = 0.055) (Table 2), but we can note that the highest fecundity for T. (A.) recki collected on M. suaveolens (1.00 eggs/ female) and the lowest for T. (A.) recki collected on D. stramonium (0.35 eggs/ female). Significant differences were only significant at day 1 (P=0.02) (Table 4, Figure 3e). The values obtained were much lower than the ones reported for N. cucumeris when fed with A. lycopersici by Al-Azzazy et al. (2018) (2.12 eggs/ female/ day), for Euseius concordis (Chant) by Moraes and Lima (1983) (1.7 eggs/ female/ day), Amblydromalus limonicus Garman & McGregor (Van Houten et al. 2013a, b) (3.6 eggs/ female/ day) and Amblyseius swirskii Athias-Henriot (Momen and Abdel-Khalek 2008; Park et al. 2010; Van Houten et al. 2013a, b) (2.00, 1.7 and 2.4 eggs/ female/ day, respectively). Castagnoli et al. (2003) reported a decreasing rm from F1 to F3 generations (0.21 to 0.08) for N. californicus feeding on A. lycopersici. Some preliminary measurements (data not shown) indicate a consumption of 50 eriophyids mites / day by T. (A.) recki (population from M. suaveolens). This value is much higher than the one reported by Al-Azzizy et al. (2018) for N. cucumeris (consumption ranging from 12.82 to 21.69 A. lycopersici per day).
Considering each predator separately, T. evansi was the less favourable food source. However, no distinction between the populations/species was observed when the prey was T. urticae or A. lycopersici. Only, a higher fecundity was observed for N. californicus fed on T. urticae than on A. lycopersici (Table 2).
Dispersal abilities of Typhlodromus (Anthoseius) recki along tomato stem in laboratory studies
Females dispersed along the stem bridge whatever the food source (pollen or A. lycopersici). A low proportion was observed on the stem. They essentially moved during the first day (Figure 4). Afterwards, their movement was less frequent. The females were preferentially found on the discs with food, both for pollen and A. lycopersici treatments (Table 5). The number of eggs laid on the leaf disc with food was significantly higher than those laid on the leaf disc without food (P < 0.0001 - pollen disc : 1.50 eggs/female/day, disc without pollen: 0.25 eggs/ female/ day; P = 0.03 - disc with A. lycopersici : 1.09 eggs/ female/ day, disc without A. lycopersici: 0.40 eggs/ female/ day). No significant difference in fecundity was observed for leaf discs with A. lycopersici and pollen (P = 0.13). For a same replicate, eggs were sometimes observed on both leaf discs suggesting that the female was able to re-cross the stem bridge. Even if those results are encouraging, cut tomato stem have different physical and chemical properties than not-cut ones. Thus, further experiments on entire plants are needed to conclude on the ability T. (A.) recki to disperse along the stem, as shown for other Phytoseiidae species (P. longipes and Phytoseiulus macropilis (Banks)) by Sato et al. (2011).


Assessment of the ability of T. (A.) recki to move from mint to tomato leaves under laboratory conditions
One day after the beginning of the experiments, females of T. (A.) recki dispersed to the tomato half-disc infested with T. evansi (Table 6). Overall, the number of females and the percentage of occupation of each half-disc (mint vs. tomato) were not significantly different. However, at days 3 and 6, the number of females of the tomato half-disc was higher than on the mint half-disc and the percentage of females on tomato half-disc was higher than on mint at days 2, 3 and 6 (Figure 5).

The number of eggs+ immatures of T. (A.) recki was significantly greater on the mint half-disc at day 1 and day 2. At day 3, no significant difference was observed between the two treatments, whereas at days 6 and 7, the number of eggs + immatures were much greater on the tomato half-disc (Table 6). At day 7, all the eggs of T. (A.) recki were laid on the tomato disc. The percentage of eggs+immatures was significantly higher on mint half-discs at day 1 and 2, whereas it was significantly higher on tomato half-discs at day 7 (Figure 5).

The number of T. evansi females decreased significantly through time (from 5 females / leaf disc at day 0 to 1.72 at day 7). Several hypotheses can explain this observation: (i) female escaped (but this is unlikely as they would be observed on moistened cotton), (ii) female were predated even if predation usually occur on eggs and immature stages and/or (iii) female died (but cadavers were not retrieved). The number of eggs and immatures of T. evansi increased until the day 3 and then decreased but no significant difference was observed through the time (P=0.32) (Table 6).
Conclusion
This study presents first data on the biology of T. (A.) recki. It confirms that this species is a generalist predator, with biological features close to those of other efficient generalist predators as K. aberrans and T. (T.) pyri. Its mean consumption of T. urticae is similar to N. californicus, which is considered as an efficient predator of spider mites (i.e. Knapp et al. 2018). Furthermore, it seems to perform better on T. evansi than the generalist species already tested. It is similarly attracted by prey and pollen, confirming its generalist statute. It is the first time that this species is tested on A. lycopersici, one of the most important pests in tomato crops (Trottin-Caudal et al. 2003; Duso et al. 2009). These preliminary results are encouraging considering both biological features and dispersal abilities. However, as laboratory observations do not entirely reflect real situations, trials on entire plants are required. Furthermore, because of the biological characteristics of this generalist predator, such as for K. aberrans and T. (T.) pyri, biodiversity conservation biological control should be favoured, in line with the banker plant strategy already tried in tomato greenhouses to control white flies (i.e. Frank 2010). Several characteristics of T. (A.) recki (endemic species of Europe, current distribution, its occurrence on Solanaceae and the fact that it is attracted by prey) constitute elements to further investigate its predation ability at plant and crop level.
Acknowledgements
We thank the French national program (Ecophyto – Project ACAROSOL) that granted in part the surveys carried out. We are also grateful to the two anonymous reviewers for their accurate and relevant comments.
References
Al-Azzazy M., Suloiman M., Al-Rehiayani M., Nagdy F. A. 2018. Life tables of the predatory mite Neoseiulus cucumeris (Acari: Phytoseiidae) on two pest mites as prey, Aculops lycopersici and Tetranychus urticae. Archives of Phytopathology and Plant Protection 51 (11-12) : 637-648. doi:10.1080/03235408.2018.1507013
Basheer A., Saker I., Dahiah H., Mofjeh M. 2014. Life Table Parameters of Typhlodromus (Typhlodromus) athiasae Porath and Swirski (Gamasida: Phytoseiidae), Predator of the Two-Spotted Spider Mite, Tetranychus urticae (Koch) (Actinedida: Tetranychidae). Egyptian Journal of Pest Control, 24(2): 373-377.
Castagnoli M., Simoni S., Liguori M. 2003. Evaluation of Neoseiulus californicus (McGregor) (Acari: Phytoseiidae) as a candidate for the control of Aculops lycopersici (Tyron) (Acari Eriophyoidea): a preliminary study. Redia, 86: 97-100.
Castagnoli M., Simoni S., Nachman G. 2001. Short-term changes in consumption and oviposition rates of Neoseiulus californicus strains (Acari: Phytoseiidae) after a diet shift. Exp. Appl. Acarol., 25: 969-983. doi:10.1023/A:1020639301898
Canlas L.J., Mamano H., Ochiai M., Takeda M. 2006. Biology and predation of the strain Japanese of Neoseiulus californicus (McGregor) (Acari: Phytoseiidae). Syst. Appl. Acarol., 11 (2): 141-157. doi:10.11158/saa.11.2.2
Demite P.R., Moraes G.J. de, McMurtry J.A., Denmark H.A., Castilho R.C. 2020. Phytoseiidae Database. Available from: url{www.lea.esalq.usp.br/phytoseiidae} (accessed 15/IV/2020)
Duso C., Castagnoli M., Simoni S., Angeli G. 2009. The impact of eriophyoids on crops: Recent issues on Aculus schlechtendali, Calepitrimerus vitis and Aculops lycopersici. Exp. Appl. Acarol., 51(1-3): 151-68. doi:10.1007/978-90-481-9562-6_8
Escudero L.A., Ferragut F. 2005. Life-history of predatory mites Neoseiulus californicus and Phytoseiulus persimilis (Acari: Phytoseiidae) on four spider mite species as prey, with special reference to Tetranychus evansi (Acari: Tetranychidae). Biol Control, 32(3): 378-384. doi:10.1016/j.biocontrol.2004.12.010
Ferrero M., Tixier M.-S., Kreiter S. 2014a. From specificity to specialisation. I. Comparative life histories of three populations of Phytoseiulus longipes Evans (Acari, Mesostigmata, Phytoseiidae) depending on prey species and plant supports. Exp. Appl. Acarol., 62: 313-324. doi:10.1007/s10493-013-9745-z
Ferrero M., Tixier M.-S., Kreiter S. 2014b. From specificity to specialization. II. Choice tests and attractiveness of different prey species, prey stages and plant supports for two populations of Phytoseiulus longipes Evans [Acari, Mesostigmata, Phytoseiidae]. Exp. Appl. Acarol., 62: 325-335. doi:10.1007/s10493-013-9746-y
Frank S.D. 2010. Biological control of arthropod pests using banker plant systems: Past progress and future directions. Biol. Contr., 52 : 8-16 doi:10.1016/j.biocontrol.2009.09.011
Furtado I.P., Moraes G.J.de, Kreiter S., Tixier M.-S., Knapp M. 2007.Potential of a Brazilian population of the predatory mite Phytoseiulus longipes as a biological control agent of Tetranychus evansi (Acari: Phytoseiidae, Tetranychidae). Biol. Control, 42: 139-147. doi:10.1016/j.biocontrol.2007.04.016
Gerson U., Smiley R.L., Ochoa T. 2003. Mites (Acari) for Pest Control. Blackwell Science, Oxford, UK, 539pp. doi:10.1002/9780470750995
Gotoh T., Yamaguchi K., Mori K. 2004. Effect of temperature on life history of the predatory mite Amblyseius (Neoseiulus) californicus (Acari: Phytoseiidae). Exp. Appl. Acarol., 32(1-2):15-30. doi:10.1023/B:APPA.0000018192.91930.49
Hajizadeh H., Kamali K., Faraji F. 2006. Predatory mite fauna of Phytoseiidae of northwest Iran (Acari: Mesostigmata). Turk. J. Zool., 34(4): 497-508.
Hatherly I., Bale J. S., Walters K.F.A. 2005. Intraguild predation and feeding preferences in three species of phytoseiid mite used for biological control. Exp. Appl. Acarol., 37(1-2): 43-55. doi:10.1007/s10493-005-0358-z
Karban R., English-Loeb G., Walker M.A., Thaler J. 1995. Abundance of phytoseiid mites on Vitis species: effects of leaf hairs, domatia, prey abundance and plant phylogeny. Exp. Appl. Acarol., 19(4): 189-197. doi:10.1007/BF00130822
Kasap I. 2010. Prey-stage preference and population increase of the predaceous mite Kampimodromus aberrans (Oudemans) (Acari: Phytoseiidae) on Tetranychus urticae Koch (Acari: Tetranychidae) under laboratory conditions. Int. J. Acarol., 36 (6): 473-481. doi:10.1080/01647950903575341
Kennedy G.G. 2003. Tomato, pest, parasitoids, and predators: tritrophic interactions involving the genus Lycopersicon. Ann. Rev. Entomol., 48: 51-72. doi:10.1146/annurev.ento.48.091801.112733
Knapp M., Van Houten Y., Van Baal E., Grout T. 2018. Use of predatory mites in commercial biocontrol: current status and future prospects. Acarologia, 58: 72-82. doi:10.24349/acarologia/20184275
Koller M., Knapp M., Schausberger P. 2007. Direct and indirect adverse effects of tomato on the predatory mite Neoseiulus californicus feeding on the spider mite Tetranychus evansi. Entomol. Experim. Appl., 125: 297-305. doi:10.1111/j.1570-7458.2007.00625.x
Kreiter S., Tixier M.-S., Croft B.A., Auger P., Barret D. 2002. Plants and leaf characteristics influencing the predaceous mite Kampimodromus aberrans (Acari: Phytoseiidae) in habitats surrounding vineyards. Environ. Entomol., 31: 648-660. doi:10.1603/0046-225X-31.4.648
Kumral N.A., Cobanoglu S. 2015a. The potential of the nightshade plants (Solanaceae) as reservoir plants for pest and predatory mites. Turk. J. Entomol., 39(1): 91-108. doi:10.16970/ted.55042
Kumral N.A., Cobanoglu S. 2015b. A reservoir weed for mites: Datura stramonium L. (Solanaceae) in the vicinity of cultivated solanaceous plants in Turkey. Int. J. Acarol., 41: 563-573. doi:10.1080/01647954.2015.1084043
Lebdi-Grissa K., Van Impe G., Lebrun P. 2005. Paramètres biologiques et démographiques de Neoseiulus californicus (acari : phytoseiidae) à differentes temperatures. Acarologia, 45 (1): 13-22.
Li G.Y., Zhang Z.K. 2016. Some factors affecting the development, survival and prey consumption of Neoseiulus cucumeris (Acari: Phytoseiidae) feeding on Tetranychus urticae eggs (Acari: Tetranychidae) Syst. Appl. Acarol., 21(5): 555-566. doi:10.11158/saa.21.5.1
Marafeli P.P., Reis P.R., Silveira E.C., Souza-Pimentel G.C., Toledo M.A. 2014. Life history of Neoseiulus californicus (McGregor, 1954) (Acari: Phytoseiidae) fed with castor bean (Ricinus communis L.) pollen in laboratory conditions. Braz. J. Biol., 74(3): 691-697. doi:10.1590/bjb.2014.0079
McMurtry J.A., Croft B.A. 1997. Life-styles of phytoseiid mites and their roles in biological control. Annu. Rev. Entomol., 42: 291-321. doi:10.1146/annurev.ento.42.1.291
McMurtry J.A., Moraes G.J. De, Sourassou N.F. 2013. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol., 18(4): 297-320. doi:10.11158/saa.18.4.1
McMurtry J.A., Scriven G.T. 1965. Insectary production of phytoseiid mites. J. Econ. Entomol., 58(2): 282-284. doi:10.1093/jee/58.2.282
Momen F.M., Abdel-Khalek A. 2008. Effect of the tomato russet mite Aculops lycopersici (Acari: Eriophyidae) on the development and reproduction of three predatory phytoseiid mites. Int. J. Trop. Insect Sci., 28: 53-57. doi:10.1017/S1742758408942594
Moraes G.J., Lima H.C. 1983. Biology of Euseius concordis (Chant) (Acarina: Phytoseiidae) a predator of the tomato russet mite. Acarologia, 24: 251-255.
Moraes G.J. de, McMurtry J.A. 1985. Comparison of Tetranychus evansi and T. urticae (Acari: Tetranychidae) as prey for eight species of phytoseiid mites. Entomophaga, 30(4): 393-397. doi:10.1007/BF02372345
Moraes G.J.de, McMurtry J.A. 1986.Suitability of the spider mite Tetranychus evansi as prey for Phytoseiulus persimilis. Entomol. Exp. Appl. 40: 109-115. doi:10.1111/j.1570-7458.1986.tb00490.x
Moraes G.J. de, McMurtry J.A., Denmark H.A. 1986. A catalog of the mite family Phytoseiidae. References to taxonomy, synonymy, distribution and habitat-EMBRAPA - DDT, 353 pp.
Park H.H., Shipp L., Buitenhuis R. 2010. Predation, development, and oviposition by the predatory mite Amblyseius swirskii (Acari: Phytoseiidae) on tomato russet mite (Acari: Eriophyidae). J. Econ. Entomol., 103: 563-569. doi:10.1603/EC09161
Rahmani H., Fathipour Y. Kamali K. 2009. Life history and population growth parameters of Neoseiulus californicus (Acari: Phytoseiidae) fed on Thrips tabaci (Thysanoptera: Thripidae) in laboratory conditions. Syst. Appl. Acarol., 14: 91-100. doi:10.11158/saa.14.2.2
Schmidt R.A. 2014. Leaf structures affect predatory mites (Acari: Phytoseiidae) and biological control: a review. Exp. Appl. Acarol., 62: 1-17. doi:10.1007/s10493-013-9730-6
Sato M.M., Moraes G.J., Haddad M.L., Weseka V.W. 2011. Effect of trichomes on the predation of Tetranychus urticae (Acari: Tetranychidae) by Phytoseiulus macropilis (Acari: Phytoseiidae) on tomato, and the interference of webbing. Exp. Appl. Acarol., 54: 21-32. doi:10.1007/s10493-011-9426-8
Simmons A.T., Gurr G.M. 2005. Trichomes of Lycopersicon species and their hybrids: effects on pests and natural enemies. Agric. Forest Entomol., 7: 265-276. doi:10.1111/j.1461-9555.2005.00271.x
StatSoft France 2010. STATISTICA (logiciel d'analyse des données), version 9.1. http://www.statsoft.fr
 .
.Swirski E., Amitai S. 1997. Annotated list of phytoseiid mites (Mesostigmata: Phytoseiidae) in Israel. Isr. J. Entomol., 31: 21-46.
The Angiosperm Phylogeny Group. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botan. J. Linn. Soc., 161, 105-121. doi:10.1111/j.1095-8339.2009.00996.x
Tixier M.-S. 2018. Predatory mites (Acari: Phytoseiidae) in agro-ecosystems and conservation biological control: a review and explorative approach for forecasting plant-predatory mite interactions and mite dispersal. Front. Ecol. Evol. 6: 192. doi:10.3389/fevo.2018.00192
Tixier M.-S., Kreiter S., Douin M. 2020. Phytoseiidae (Acari: Mesostigmata) on plants of the family Solanaceae: results of a survey in South of France and a review of world biodiversity. Exp. Appl. Acarol. In press. doi:10.1007/s10493-020-00507-0
Trottin-Caudal Y., Fournier C., Leyre J.-M. 2003. Biological control of Aculops lycopersici (Massee) using the predatory mites Neoseiulus californicus McGregor and Neoseiulus cucumeris (Oudemans) on tomato greenhouse crops. In: Proceedings, international symposium on Greenhouse Tomato: integrated crop protection and organic production, 17-19 September 2003. Avignon, France: 153-157.
Van Houten Y., Glas J.J., Hoogerbrugge H., Rothe J., Bolckmans K. , Simoni S., van Arkel J., Alba J.M., Kant M.J., Sabelis M.W. 2013a. Herbivory-associated degradation of tomato trichomes and its impact on biological control of Aculops lycopersici. Exp. Appl. Acarol., 60: 127-138. doi:10.1007/s10493-012-9638-6
Van Houten Y., Knapp M., Hoogerbrugge H., Bolckmans K. 2013b. The potential of Amblyseius swirskii as biocontrol agent for Aculops lycopersici on tomatoes. IOBC-WPRS Bulletin, 93: 51-57.
Van Lenteren J.C. 2012. The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. BioControl, 57:1-20. DOI 10.1007/s10526-011-9395-1 doi:10.1007/s10526-011-9395-1
Van Lenteren J.C., Bolckmans K., Köhl J., Ravensberg W.J., Urbaneja A. 2017. Biological control using invertebrates and microorganisms: plenty of new opportunities. BioControl, 63 : 39-59 (2018). doi:10.1007/s10526-017-9801-4
Zemek R. 1993. Characteristics of development and reproduction in Typhlodromus pyri on Tetranychus urticae and Cecidophyopsis ribis. II. Progeny of overwintered females. Exp. Appl. Acarol., 17(11): 847-858. doi:10.1007/BF00225858
\biblio{Zheng Y., De Clercq P., Song Z.-W., Li D.-S., Zhang B.-X. 2017. Functional response of two Neoseiulus species preying on Tetranychus urticae Koch. Syst. Appl. Acarol., 22(7): 1059-1068. \doibib{doi:10.11158/saa.22.7.13
} }



2020-05-20
Date accepted:
2020-09-28
Date published:
2020-09-30
Edited by:
Tsolakis, Haralabos

This work is licensed under a Creative Commons Attribution 4.0 International License
2020 Tixier, Marie Stephane; Douin, Martial; Rocio, Oliva; Gonzalez, Lucia; Pount, Benjamin and Kreiter, Serge
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)