



Development, reproduction and life table parameters of Tetranychus turkestani (Acari: Tetranychidae) on three different host plants
Bazazzadeh, Fereshteh1 ; Shishehbor, Parviz2 ; Esfandiari, Mehdi3 and Farahi, Sara4
1Department of Plant Protection, Faculty of Agriculture, Shahid Chamran University of Ahvaz, Ahvaz, Iran.
2✉ Department of Plant Protection, Faculty of Agriculture, Shahid Chamran University of Ahvaz, Ahvaz, Iran.
3Department of Plant Protection, Faculty of Agriculture, Shahid Chamran University of Ahvaz, Ahvaz, Iran.
4Department of Plant Protection, Faculty of Agriculture, Shahid Chamran University of Ahvaz, Ahvaz, Iran.
2020 - Volume: 60 Issue: 3 pages: 643-655
https://doi.org/10.24349/acarologia/20204393Original research
Keywords
Abstract
Introduction
The strawberry spider mite, Tetranychus turkestani Ugarov & Nikolski (also known as Tetranychus atlanticus McGregor), is one of the most important agricultural pests in the world (Jeppson et al. 1975; Bolland et al. 1998). It is a polyphagous species and feeds on 271 plant species belonging to more than 65 plant families (Migeon and Dorkled 2020). This phytophagous mite is distributed throughout the world and causes yield losses in cultivated crops (field and glasshouse crops) by sucking the cell contents (Jeppson et al. 1975; Helle and Sabelis 1985; Zhang 2003; Hoy 2011).
Feeding activity of T. turkestani leads to the appearance of white chlorotic spots on upper side of leaves with copious webbing on the underside of the leaves. In serious infestations, leaves may drop and flowering may be noticeably reduced. The short life span and high reproductive potential cause a fast-growing population that allows the mite to attain economic injury level in appropriate condition, resulting in a rapid decline of host plant yield (Carey and Bradley 1982; Tomczyk and Kropczynska 1985; Sohrabi and Shishehbor 2008; Karami Jamour et al. 2012).
The effect of host plant morphological and chemical features on biological characteristics of tetranychid mite species has been indicated frequently (Tomczyk and Kropczynska 1985; Awmack and Leather 2002). Host plant species or cultivars are among the most important factors that affect development time, survival, longevity and fecundity of spider mites (den Boom et al. 2003; Vasquez et al. 2008). Information on the impact of Leguminosae host plants on the development and reproductive performance of T. turkestani is limited; in fact, the only documented report was that of Sohrabi and Shishehbor (2008), who reported that longevity of T. turkestani varied among three Leguminosae plant species (cowpea, green gram, pinto bean).
Several important field crops belonging to the Fabaceae, Cucurbitaceae and Malvaceae plant families are attacked by T. turkestani, among which Fabaceae (Leguminosae) plant species (commonly known as the legume, pea or bean) are the most important host plants for this mite pest (Jeppson et al. 1975). However, little is known about the development, survival, reproduction and life table parameters of T. turkestani on Fabaceae host plants. Development of efficient strategies for controlling strawberry spider mite will require knowledge of its biological relationships with various host plants. Among these, an important component will be an understanding of host suitability. Hence, the aim of this study was to better understand the influence of some Leguminosae plant species on life history parameters of T. turkestani by constructing the life tables of this spider mite on three commercial Leguminosae plants.
Materials and methods
Plant materials
Three plant species of Fabaceae were used in this study: Cowpea (Vigna unguiculata L.), white bean (Phaseolus lunatus L.) and red bean (Phaseolus calcaratus Roxb.). Seeds were planted in plastic pots (20 cm diameter, 10 cm height) in a mixture of compost (Nutrifood Company, Iran) and sawdust (1:1 ratio) and kept in wooden cages (120 × 60 × 60 cm) covered by white nylon mesh (with an aperture of 120 µm). Plants were maintained in the laboratory at 25 ± 1 °C, 65 ± 5% RH and 16:8 (L:D) with illumination (4000 lux) provided by fluorescent lamps. Studies were initiated when cowpea, white bean and red bean were three, four and four weeks old, respectively. The heights of these plants at these ages were ≈ 30, 25 and 25 cm for cowpea, white bean and red bean, respectively. Only the second to fourth fully expanded leaves (from the growing terminal) were selected. The leaves were replaced every three to four days throughout the study.
Spider mite colony
Tetranychus turkestani was collected from morning glory (Convolvulus arvensis L.) leaves at Shahid Chamran University of Ahvaz, Ahvaz (Iran) (31°18'21''N 48°39'33''E), and used to start rearing the colony. This stock colony was separately maintained on cowpea, white bean and red bean plants. Infested plants were kept in wooden framed cages under laboratory conditions described above. Plants were maintained until they were severely damaged by the spider mites; new plants being added when needed. Synchronous colonies of T. turkestani were provided by rearing five generations (which lasted 80, 65 and 68 days) on cowpea, white bean and red bean, respectively.
Preimaginal developmental time, survival and sex ratio
Plastic boxes (14 × 11 × 4 cm) described by Sohrabi and Shishehbor (2008) were used as test arenas. Three layers of cotton mat, with the same size, soaked in water and placed in the box. A detached leaf of each plant species was placed lower side uppermost on the cotton mats in each arena. A narrow strip of tissue paper was placed on periphery of each leaf. For individual tests, the leaf surface was subdivided into two ca. equal areas using the same barrier. The soaked cotton mat and tissue papers kept the leaves fresh and prevented the escape of tested mites. For each experiment, one mated adult female from each stock colony was transferred with the aid of a fine camel hair brush (000) to the arenas and was allowed to lay eggs. After a 24 hours oviposition period, only one newly laid egg was left on each leaf arena and the mite and the excess eggs were removed. Sixty eggs were used for each treatment. The plastic boxes were kept in an incubator at 25 ± 1 °C, 65 ± 5% RH and 16:8 (L:D); a new leaf was prepared and the mites were transferred at the first sign of leaf deterioration. Egg to adult developmental time, survival and sex ratio were checked twice daily, under a dissecting microscope at magnification up to 100 x. The presence of an exuvium was used as the criterion for successful molting. For calculation purposes, we assumed that molting or death occurred at the midpoint between two successive observations (Karami Jamour and Shishehbor, 2012). Individuals trapped in the wet tissue paper surrounding the leaf arena were excluded from data analysis.
Adult longevity and fecundity
Adults used for longevity and fecundity experiments were reared from eggs obtained from previous experiments. Newly molted adult female and male of T. turkestani (age < 24 h old) were paired and transferred into the new test arenas, so each arena contained one female and one male. Egg laying and longevity were recorded twice daily. Every three days, mites were transferred to a new arena containing a detached leaf until the female died. Males that died or escaped from the experimental unit were replaced by young ones. Females trapped in the wet tissue or dead because of improper handling were excluded from data analysis.
Statistical analysis
The age-stage specific survival rates (Sxj ) (where x is the age and j is the stage), age-stage specific fecundity (fxj ), age-specific survival rates (lx ), age-specific fecundity (mx ), and the population parameters net reproductive rate (R0 ), intrinsic rate of increase (rm ), finite rate of increase (λ) and mean generation time (T) were calculated using an age-stage, two-sex life table (Chi and Liu 1985; Chi 1988). The age-stage life expectancy (exj ) was calculated according to Chi and Su (2006). The net reproductive rate (R0 ) is the total number of offspring produced by an average individual during its lifetime and was calculated as:
\[R_0=\sum_{x=0}^{\infty} l_x m_x\]
The intrinsic rate of increase is estimated by using iterative bisection method from:
\[\sum_{x=0}^{\infty} e^{-r(x+1)} l_x m_x=1\]
with age indexed from 0 (Goodman 1982). The mean generation time is defined as the length of time that a population needs to increase by R0 - fold of its size at the stable age-stage distribution, i.e., erT = R0 or λT = R0 . Therefore, it is calculated as T= (ln R0 )/rm . The gross reproductive rate (GRR) is calculated as Σ mx . The reproductive value (vxj ) considered as the expectation of future offspring of individuals of age x and stage j (Fisher 1930). To estimate the means, variances and standard errors of the population parameters of T. turkestani, bootstrap techniques were used (Efron and Tibshirani 1993) with 100,000 bootstrap samples for more stable and precise estimates. The computer program TWOSEX-MSChart (Chi 2017) was used to perform the analysis and estimate the life table parameters. A paired bootstrap test was also used to compare the differences in life history parameters among host plant species.
Results
The female and male immature developmental time are shown in Tables 1 and 2, respectively. Significant differences were observed between immature developmental duration of females (Table 1, P < 0.05). Same results obtained for males (Table 2, P < 0.05). Mean total development duration of T. turkestani females and males from egg to adult were about 1.5 days longer on white bean and red bean compared to cowpea. However, there were no significant differences in the mean developmental time of female and male T. turkestani on white bean and red bean.
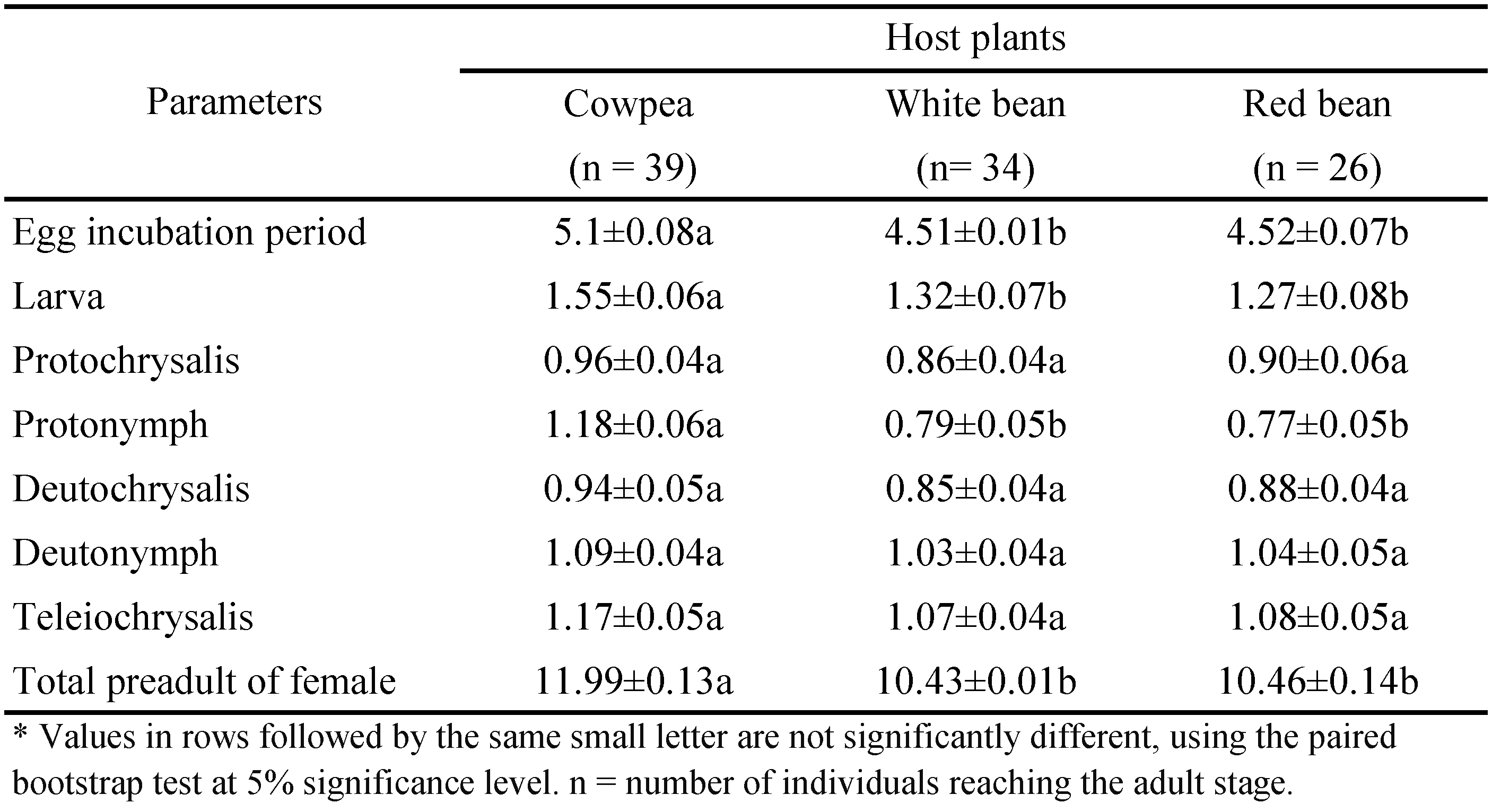

Host plant had no significant effect on the total preadult survivorship of T. turkestani (P < 0.05). The highest percentage of mite preadult survival was found on cowpea (80 ± 0.05), followed by white bean (76.67 ± 0.05) and red bean (75 ± 0.05). Host plant significantly affected female progeny percentage (P < 0.05). The highest percentage of female progeny was recorded from T. turkestani reared on cowpea (80% ± 0.43) followed by white bean (73% ± 0.47) and red bean (58% ± 0.65).

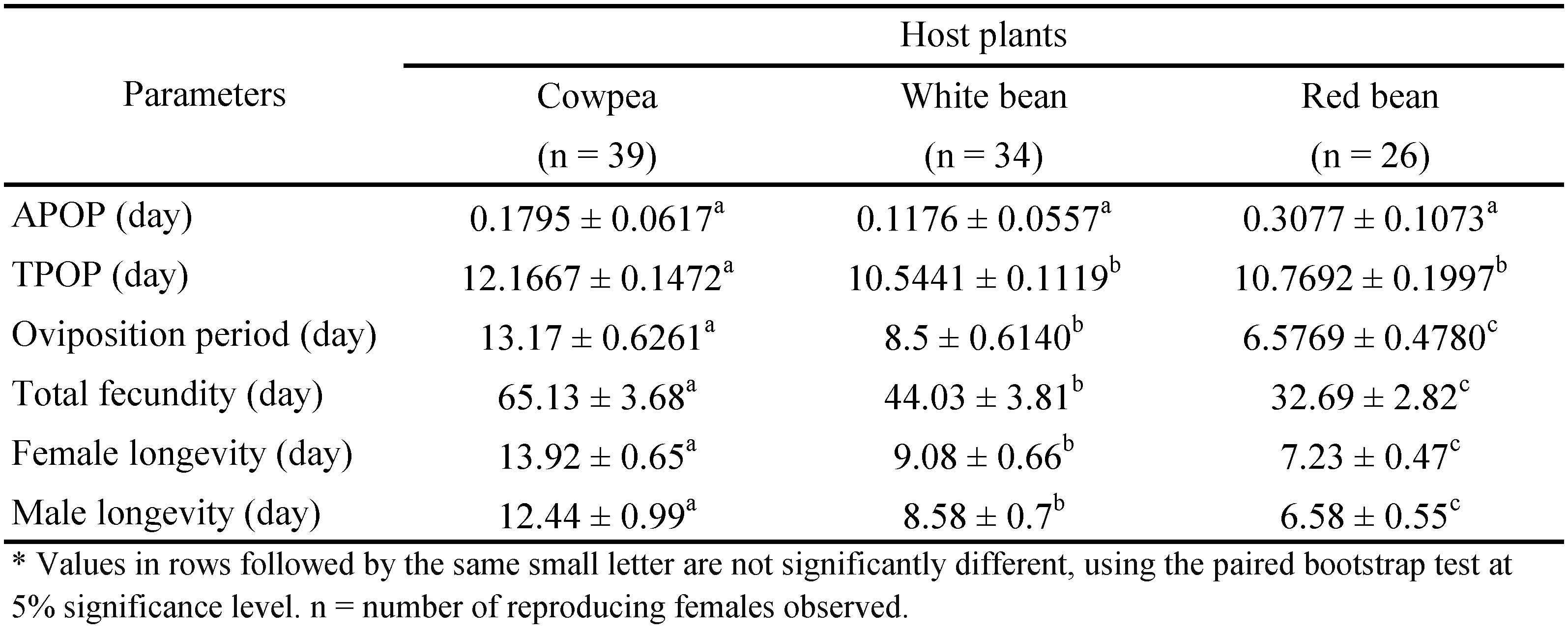
The mean duration of preoviposition (from adult emergence to egg laying, APOP) did not differ significantly among the three host plants (Table 3). In the mean duration of total preoviposition (from egg development to egg laying, TPOP), the difference was found only between cowpea and the two beans. However, oviposition duration (from first to last egg laid) and female and male longevity differed significantly among host plant species tested (Table 3).
The age-stage-specific survival rate (Sxj) shows the probability that an egg can survive to age x and stage j (Figure 1). Significant overlaps were observed between stages in all three reproductive cohort of T. turkestani due to variable developmental rates occurring among individuals. The ability to observe the beginning and ending of subsequent stages (i. e., egg, larva, etc. stages) in the survival curve for each stage is an advantage of using the age-stage, two sex life table. We took the non-feeding resting stages (protochrysalis, deutochrysalis and teliochrysalis) out of the figures 1-4 to avoid presenting busy charts.

The age-specific survival rate (lx ), age-specific fecundity (mx ), age-specific maternity (lxmx ) and age-stage specific fecundity (fx ) were calculated and are shown in Figure 2. The age-specific survival rate demonstrates a simplified overview of the survival history when all stages are pooled (Figure 2). The lx is affected by bean plant species. Oviposition period varied from 6.57 (red bean) to 8.50 (white bean) and 13.17 (cowpea) and differed significantly between three host plants (Table 3).

Host plant had no significant effect on mean daily fecundity. However, mean total fecundity of T. turkestani was significantly different among the three host plants (Table 3). The highest mean total number of eggs was laid on cowpea (65.13 ± 3.68 eggs/female).
The age-stage-specific life expectancy (exj ) is the lifespan remaining for an individual of age x and stage j. The age-stage specific life expectancies of T. turkestani on different host plants species are plotted in Figure 3. The life expectancy of a newborn egg was about 14, 16 and 21 in mites reared on red bean, white bean and cowpea, respectively. The maximum life expectancy of all stages of T. turkestani was recorded in cowpea. Life expectancy decreased gradually with age, since the study was conducted in the laboratory and thus unaffected.

The age stage reproductive value (vxj ) of T. turkestani represents the contribution of an individual to the future population (Figure 4). The reproductive value of a newborn (v01 ) is the finite rate itself. For example, a newly emerged female (age 15 d) has a reproductive value of 24.54 in cowpea. On the other hand, a 11-d old female has a lower reproductive value, 18.60 in red bean. The reproductive curve (vxj ) of T. turkestani shows the highest value in mites reared on cowpea. Life table statistics of T. turkestani clearly differed among host plants (Table 4). Mites reared on cowpea had a significantly higher net reproductive rate (R0 ) than those reared on white bean and red bean. However, host plant species had no significant effect on intrinsic rate of increase (rm ) and finite rate of increase (Table 4).


Discussion
Different host plants significantly affected the biology of T. turkestani. The strawberry spider mite was able to reproduce on all host plants studied; however, the cowpea was more suitable for its development and reproduction. Several features of the host plant, such as morphology, nutritional values, and secondary chemical compounds, may have affected the biological reaction of T. turkestani. However, the effect of these factors upon development and reproduction of this species need to be studied in detail. Morphological and chemical characteristics of the plants, as well as nitrogen levels, are known to be related to mite development and reproduction (Tomczyk and Kropczynska 1985).
Results of our experiment showed that host plant can affect development time of T. turkestani. Developmental duration of T. turkestani female ranged from 10.43 days on white bean to 11.99 days on cowpea. Developmental time of female T. turkestani was reported to be 10.80 days on alfalfa (Andres 1957), 10.70 days on cotton (Carey and Bradley 1982), 9.90 days on eggplant (Nemati et al. 2005), 12.17, 11.79, 10.81 days on cowpea, green gram, pinto bean, respectively (Sohrabi and Shishehbor 2008), 8.92 days on kidney bean (Latifi et al. 2010) and 10.27 days on cucumber (Karami-Jamour and Shishehbor 2012) which are similar to our results. However, lower developmental time of 7.70 days was reported by Cagle (1956) for T. turkestani on cotton at the same temperature (25 °C) (Table 5). Differences in the mite strain, host plant and experimental condition may provide an explanation for shorter developmental time.
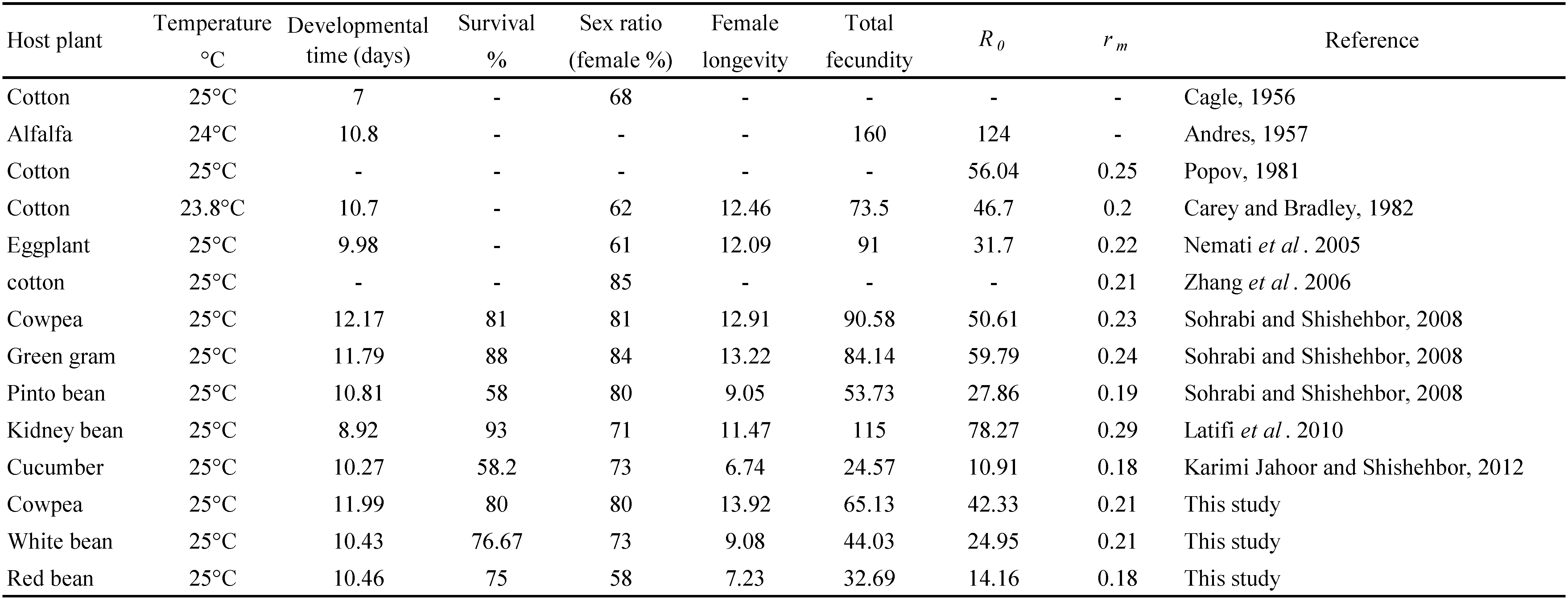
The egg to adult development duration of males of T.turkestani on all of the host plant species tested was shorter than the respective duration of females. A similar trend has also been reported for T. turkestani on cowpea, green gram and pinto bean (Sohrabi and Shishehbor 2008), on kidney bean (Latifi et al. 2010) and other tetranychid species such as Eutetranychus orientalis (Klein) (Imani and Shishehbor 2009), Tetranychus urticae Koch (Riahi et al. 2011) and Tetranychus pacificus McGregor (Carey and Bradley 1982).
Survival of T. turkestani on different host plants has been documented in the literature. Preimaginal survival of T. turkestani was reported to be 82 and 89 % on cowpea and green gram, respectively (Sohrabi and Shishehbor 2008), 93 % on kidney bean (Latifi et al. 2010) at the same temperature (25 °C) which are similar to the results obtained in the present study. However, lower survival of 58% was reported by Karami Jamour and Shishehbor (2012) on cucumber. These disparities may be explained by dissimilarities in host plant suitability for T. turkestani along with differences in experimental conditions.
The sex ratio was strongly biased for females. These data are in line with the findings of Sohrabi and Shishehbor (2008) who reported 91, 80 and 84% females for T.turkestani on cowpea, green gram and pinto bean, respectively. Sex ratio of other genera of tetranychid mites are known to be female biased (Huffaker et al. 1969). Helle and Pijnacker (1985) also reported that the ratio of 1:3 male:female is often found in many amphigonic tetranychid species.
The longevity of females of T. turkestani on all three host plants tested was longer than the respective longevity of males. A similar trend has also been reported for T. turkestani on eggplant (Nemati et al. 2005), on cowpea, green gram and pinto bean (Sohrabi and Shishehbor 2008), on kidney bean (Latifi et al. 2010), on cucumber (Karami Jamour and Shishehbor 2012) (Table 5) and other tetranychid species such as E. orientalis (Imani and Shishehbor 2009), and T. urticae (Riahi et al. 2011).
Only a few studies have been carried out to assess the biology of T. turkestani on Leguminose plant species. Mean total fecundity of T. turkestani was reported to be 90.58, 84.14 and 53.73 eggs/female on cowpea, green gram and pinto bean, respectively (Sohrabi and Shishebor 2008) and 115 eggs/female on kidney bean (Latifi et al. 2010) (Table 5). The values found in both studies are higher than those found in the current study, i. e., 65.13, 44.03 and 32.69 eggs/female on cowpea, white bean and red bean, respectively. Differences in ecological factors, namely, strain of mites, host plant species, as well as measurement methods may provide an explanation for higher fecundity of T. turkestani reported in other studies compared to our results.
At 25 °C, intrinsic rate of increase (rm ) of T. turkestani was reported to be 0.20 on cotton (Carey and Bradley 1982), 0.22 on eggplant (Nemati et al. 2005), 0.21 on cotton (Zhang et al. 2006), 0.23, 0.24, 0.19 on cowpea, green gram and pinto bean, respectively (Sohrabi and Shishehbor 2008) and 0.18 on cucumber (Karami Jamour and Shishehbor 2012) which are similar to our results (Table 5). However other laboratory studies have reported a variety of rm values for this species. The value of rm of T. turkestani was found to be 0.25 on cotton (Popov 1981) and 0.29 on kidney bean (Latifi et al. 2010) which are higher than the values obtained in the present study, i. e., 0.21, 0.21, 0.18 on cowpea, white bean and red bean, respectively, reflecting lower juvenile mortality, higher fecundity and longer adult life span in latter studies. Differences in ecological elements, such as strain of mite and species of host plant, in addition to data analysis method, may provide an explanation for higher rm .
The results of the current study indicated that T. turkestani performance was best on cowpea, worst on red bean, and intermediate on white bean. With rapid development and high survival rates, T. turkestani can cause significant damage to many economically important Leguminosae crops. If the ecological conditions are suitable, a female T. turkestani can complete its development in about 10 days, and produce up to 65 eggs. Knowledge of the biology of a mite is important for improving programs for pest management, predicting the population appearance and density, and for determining the timing of insecticide applications, as well as release of natural enemies in pest management.
Acknowledgements
This study was financially supported by the research deputy of Shahid Chamran University of Ahvaz, Ahvaz, Iran.
References
Andres L.A. 1957. An ecological study of three species of Tetranychidae (Acarina: Tetranychidae) and their response to temperature and humidity [PhD Thesis] Berkeley; University of California. pp. 49.
Awmack C.S., Leather R.S. 2002. Host plant quality and fecundity in herbivorous insects. Ann. Rev. Entomol., 47: 817-844. doi:10.1146/annurev.ento.47.091201.145300 ![]()
Bolland H.R., Gutierrez J., Flechtmann C.H.W. 1998. World catalogue of the spider mite family (Acari: Tetranychidae), with reference to taxonomy, synonymy, host plants and distribution. Brill academic Publisher Leiden. pp. 392.
Cagle L.R. 1956. Life history of the spider mite, Tetranychus atlanticus McG. Virginia Agric. Exp. Station. Tech. Bull., 124, pp. 22.
Carey J.R., Bradley J.W. 1982. Developmental rate, vital schedules, sex ratios, and life tables of Tetranychus urticae, T. turkestani and T. pacificus (Acarina: Tetranychidae) on cotton. Acarologia, 23(4): 333-345.
Chi H. 1988. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol., 17: 26-34. doi:10.1093/ee/17.1.26 ![]()
Chi H. 2017. TWOSEX-MS Chart: a computer program for the age-stage, two-sex life table analysis [Internet]. Available from: http://140.120.197.173/Ecology/ ![]()
Chi H., Liu H. 1985. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sinica, 24: 225-240.
Chi H., Su H.Y. 2006. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 35:10-22. doi:10.1603/0046-225X-35.1.10 ![]()
Efron B., Tibshirani R.J. 1993. An introduction to the bootstrap. New York, Chapman & Hall, pp. 456. doi:10.1007/978-1-4899-4541-9 ![]()
Fisher R.A. 1930. The genetical theory of natural selection. Oxford, Clarendon Press. pp. 318. doi:10.5962/bhl.title.27468 ![]()
Goodman D. 1982. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat., 119: 803-823. doi:10.1086/283956 ![]()
Helle W., Pijnacker L. P. 1985. Parthenogenesis, choromosomes and sex. In: Helle W. Sabelis M. W. (eds). Spider mites: their biology, natural enemies and control. Volume 1A. Elsevier Science Publisher. p. 129-139.
Helle W. Sabelis M. W. 1985. Spider mites: their biology, natural enemies and control. Volume 1A. Elsevier Science Publisher.
Hoy M.A. 2011. Agricultural acarology: introduction to integrated mite management. New York, CRC Press.
Huffaker C.B., Vrie M.V., McMurtry J.A. 1969. The ecology of tetranychid mites and their natural control. Ann. Rev. Entomol., 14: 125-174. doi:10.1146/annurev.en.14.010169.001013 ![]()
Imani Z., Shishehbor P. 2009. Effect of temperature on life history and life table of Eutetranychus orientalis (Klein) (Acari: Tetranychidae). Syst. Appl. Acarol., 14: 11-18. doi:10.11158/saa.14.1.2 ![]()
Jeppson L. R., Keifer H. H., Baker E. W. 1975. Mites injurious to economic plants. Berkeley, University of California Press, pp. 614.
Karami Jamour T., Shishehbor P. 2012. Development and life table parameters of Tetranychus turkestani (Acari: Tetranychidae) at different constant temperatures. Acarologia, 52(2): 113-122. doi:10.1051/acarologia/20122037 ![]()
Karami Jamour T., Shishehbor P., Mossadegh M. S. 2012. Biology and life table parameters of strawberry spider mite, Tetranychus turkestani studied on three cucurbit plants. Iranian J. Plant Prot. Sci., 43(1): 23-31. (in Persian).
Latifi M., Azmayeshfard P., Kharazipakdel A., Saboory A. R., Alahyari H. 2010. Effect of three constant temperatures on life table parameters of Tetranychus turkestani Ugarov and Nikolski (Prostigmata: Tetranychidae). Iranian J. Plant Prot. Sci., (Ir. H. Agric. Sci.), 41: 51-59.
Migeon A., Dorkled F. 2020. Spider Mites Web: a comprehensive database for the Tetranychidae. Available from: http://www1.montpellier.inra.fr/CBGP/spmweb ![]() (Accessed 20/08/2020)
(Accessed 20/08/2020)
Nemati A.R., Soleimannejadian A., Shishehbor P., Kamali, K. 2005. Evaluation of the effect of temperature on biological parameters of Tetranychus turkestani Ugarov and Nikolski (Acari: Tetranychidae). Plant Prot., (Sci. J. Agric.), 29: 209-222.
Popov S.Y. 1981. Survival tables and biological parameters of the spider mite Tetranychus turkestani. Izvestya Timiryazevskoi Selskokhozyaistvennoi Akademii, 1: 124-133.
Riahi E., Nemati A., Shishehbor P., Saeidi Z. 2011. Population growth parameters of the two-spotted spider mite, Tetranychus urticae, on three peach varieties in Iran. Acarologia, 51(4): 473-480. doi:10.1051/acarologia/20112029 ![]()
Sohrabi F., Shishehbor, P. 2008. Effect of host plant and temperature on growth and reproduction of the strawberry spider mite Tetranychus turkestani Ugarov and Nikolski (Acari: Tetranychidae). Syst. Appl. Acarol., 13: 26-32. doi:10.11158/saa.13.1.2 ![]()
Tomczyk A., Kropczynska D. 1985. Effects of host plants. In: Helle W. Sabelis M. W. (eds). Spider mites: their biology, natural enemies and control. Volume 1A. Elsevier Science Publisher. pp. 317-330.
den Boom C.E.M. van, van Beek T. A., Dicke M. 2003. Differences among plant species in acceptance by the spider mite tetranychus urticae Koch. J. Appl. Entomol., 127: 177-183. doi:10.1046/j.1439-0418.2003.00726.x ![]()
Vasquez C., Aponte O., Morales J., Sanabria M. E., Garcia G. 2008. Biological studies of Oligonychus punicae (Acari: Tetranychidae) on grapevine cultivars. Exp. Appl. Acarol., 45: 59-69. doi:10.1007/s10493-008-9154-x ![]()
Zhang Z. Q. 2003. Mites of greenhouses: identification, biology and control. CABI Publishing, pp. 244. doi:10.1079/9780851995908.0000 ![]()
Zhang Y.X., Ji J., Wang F.T., Chen X., Chen F. 2006. The fecundity potential of Tetranychus turkestani Ugarov and Nikolski (Acari: Tetranychidae). Acta Phylophylactica Sinica, 33(4): 379-383.



2020-03-17
Date accepted:
2020-09-16
Date published:
2020-09-22
Edited by:
Tsolakis, Haralabos

This work is licensed under a Creative Commons Attribution 4.0 International License
2020 Bazazzadeh, Fereshteh; Shishehbor, Parviz; Esfandiari, Mehdi and Farahi, Sara
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)