



Contribution to the knowledge of the oribatid mite genus Perxylobates (Acari, Oribatida, Haplozetidae)
Ermilov, Sergey G.1 and Liao, Jhih-Rong2
1✉ Institute of Environmental and Agricultural Biology (X-BIO), Tyumen State University, Tyumen, Russia.
2Department of Entomology, National Taiwan University, Taipei, Taiwan.
2020 - Volume: 60 Issue: 3 pages: 612-621
https://doi.org/10.24349/acarologia/20204392ZooBank LSID: B053A157-5E77-49B1-AE1D-AA2A18D24C76
Original research
Keywords
Abstract
Introduction
The oribatid mite genus Perxylobates of the family Haplozetidae (Acari, Oribatida) was proposed by Hammer (1972) with Xylobates vermiseta Balogh and Mahunka, 1968 as type species. According to Subías (online version 2020), the genus comprises 15 species, which are distributed in the Pantropical and Subtropical regions.
During taxonomic identification of oribatid mites from farmland with Ricinus communis in Taiwan, we found a new species, belonging to Perxylobates. The main goal of our paper is to describe and illustrate this new species. Hitherto, one perxylobatid mite species was registered in Taiwan, P. coreanus Choi and Aoki, 1993 (Ermilov and Liao 2018).
The additional goals of our paper are to revise generic diagnosis and to provide an identification key to known species of Perxylobates, and to discuss the systematic placement of Perxylobates mahunkai Bayoumi, 1980.
Materials and methods
Specimens — Substrate samples containing oribatid mites were collected from the Biological Control Branch, Miaoli District Agricultural Research and Extension Station (Fig. 1) in Taiwan. Mites were extracted from samples into 75% ethanol using Berlese's funnels with electric lamps in laboratory conditions during 10 days.

As mentioned below in detail, types are distributed among two institutions: the National Taiwan University, Taipei, Taiwan (NTU); the Tyumen State University Museum of Zoology, Tyumen, Russia (TSUMZ).
Observation and documentation — Specimens were mounted in lactic acid on temporary cavity slides for measurement and illustration. Body length was measured in lateral view, from the tip of the rostrum to the posterior edge of the notogaster. Notogastral width refers to the maximum width of the notogaster in dorsal view (behind pteromorphs). Lengths of body setae were measured in lateral aspect. All body measurements are presented in micrometers. Formulas for leg setation are given in parentheses according to the sequence trochanter-femur-genu-tibia-tarsus (famulus included). Formulas for leg solenidia are given in square brackets according to the sequence genu-tibia-tarsus.
Drawings were made with a camera lucida using a Leica transmission light microscope ''Leica DM 2500''.
Terminology — Morphological terminology used in this paper follows that of F. Grandjean: see Travé & Vachon (1975) for references, Norton (1977) for leg setal nomenclature, and Norton & Behan–Pelletier (2009), for overview.
Abbreviations —Prodorsum: lam = lamella; slam = sublamella; Al = sublamellar porose area; tu = tutorium; plc = prodorsal lateral carina; ro, le, in, bs, ex = rostral, lamellar, interlamellar, bothridial and exobothridial setae, respectively; bos = bothridial scale; D = dorsophragma; P = pleurophragma. Notogaster: c, la, lm, lp, h, p = notogastral setae; Aa, A1, A2, A3 = notogastral porose areas; ia, im, ip, ih, ips = notogastral lyrifissures; gla = opisthonotal gland opening. Gnathosoma: a, m, h = subcapitular setae; or = adoral seta; d, l, sup, inf, cm, acm, ul, su, lt, vt = palp setae; ω = palp solenidion; cha, chb = cheliceral setae; Tg = Trägårdh's organ. Epimeral and lateral podosomal regions: 1a, 1b, 1c, 2a, 3a, 3b, 3c, 4a, 4b = epimeral setae; PdI, PdII = pedotecta I, II, respectively; dis = discidium; con = concavity; cp = circumpedal carina. Anogenital region: g, ag, an, ad = genital, aggenital, anal and adanal setae, respectively; iad = adanal lyrifissure; Amar = marginal porose area; po = preanal organ; ψ, τ = setae of ovipositor. Legs: Tr, Fe, Ge, Ti, Ta = leg trochanter, femur, genu, tibia, tarsus, respectively; pa = leg porose area; ω, σ, φ = leg solenidia; ɛ = leg famulus; d, l, v, ev, bv, ft, tc, it, p, u, a, s, pv, pl = leg setae.
Systematics
Generic diagnosis of Perxylobates
Adult — Sexual species with dimorphism absent. Body size: Small to medium (length about 330–550). Integument: Body surface smooth or foveolate. Prodorsum: Rostrum rounded or slightly truncate. Lamella long, narrow, well separated, without cusp and tooth distally. True translamella absent. Prolamella absent or present. Sublamella and sublamellar porose area present. Tutorium ridge-like. Rostral, lamellar, interlamellar and exobothridial setae developed, usually setiform (rarely ro and/or le thickened), barbed or smooth; ro inserted dorsolaterally on the rostrum, le on end of lamella or on prodorsal surface medial to lam, in in interbothridial region. Bothridial seta long, setiform or with unilateral head, ciliate. Bothridium cup-shaped, with anterolateral scale. Dorsosejugal porose area Ad not observed. Dorsophragma elongate longitudinally. Notogaster: Anterior margin of notogaster completely absent. Pteromorph movable, large, curved ventrally. Octotaxic system with three or four (if A3 present) pairs of rounded porose areas. With 10 pairs of short, setiform notogastral setae. Dorsosejugal porose area not observed. Gnathosoma. Subcapitulum diarthric. Palp with setation 0–2–1–3–9(+ω). Solenidion of palptarsus connected to eupathidium, located on cylindrical tubercle. Axillary saccule absent. Trägårdh's organ of chelicera elongate triangular. Lateral podosomal and epimeral regions: Pedotecta I and II represented by small lamina. Genal tooth and custodium absent. Discidium and circumpedal carina present. Humeral porose areas Am and Ah not observed. Typical epimeral setal formula 3–1–3–2(or 3). Anogenital region: Five (rarely four) pairs of genital, one pair of aggenital, two pairs of anal and three pairs of adanal setae. Adanal seta ad1 posterior, ad2 lateral, ad3 anterior to anal aperture. Adanal lyrifissure located close and lateral to anal plate. Marginal porose area present, narrowly band-like. Legs: All legs monodactylous. Porose area present dorsoparaxially on all femora and on trochanters III, IV.
Juvenile instars — Not known.
Perxylobates hakkai n. sp.
ZOOBANK: 5E8F86F8-520E-4408-821C-B0807ACC1F9C ![]()
(Figures 2–4)
Diagnosis — Body size 332–398 × 190–215. Lamella long, about 2/3 length of prodorsum. Rostral, lamellar and interlamellar setae of medium length, setiform, barbed, distance ro–le distinctly shorter than le–in, le pressed to prodorsal surface. Bothridial seta long, with unilaterally dilated head, ciliate. Exobothridial seta minute. Notogastral setae short, setiform, thin, smooth. Four pairs of rounded porose areas present. Epimeral setal formula 3–1–3–2, seta 1c minute, other epimeral and anogenital setae short, setiform, thin, roughened. Five pairs of genital setae. With semi-oval concavity posteriorly to acetabulum IV. Femur III and genu IV with two and one seta, respectively.
Description — Measurements – Body length 365 (holotype), 332–398 (four paratypes); body width 199 (holotype), 190–215 (four paratypes). Females indistinctly larger than males: 365–398 × 199–215 versus 332–365 × 190–199.

Integument – Body light brown. Surface of body and all legs microporose (visible in dissected specimens under high magnification, × 1000). Antiaxial side of femur III striate.
Prodorsum (Figs 2a, 2c) – Rostrum rounded. Lamella long, about 2/3 length of prodorsum. Prolamella absent. Sublamella about 1/3 length of lamella. Sublamellar porose area (18–22 × 10–12) oval, poorly visible. Tutorium of medium size, about 1/3 length of lamella. Prodorsal lateral carina distinct. Rostral (20–28), lamellar (16–24) and interlamellar (24–30) setae setiform, barbed, distance ro–le distinctly shorter than le–in, in erect, le located slightly medial to lamellar end, pressed to prodorsal surface. Bothridial seta (69–77) with long stalk and short, unilaterally dilated head, shortly ciliate. Anterior bothridial scale well developed. Exobothridial seta (2) minute. Sejugal porose area not observed.
Notogaster (Figs 2a, 2c, 3a) – Pteromorph triangular, rounded laterally, with well developed hinge. Ten pairs of notogastral setae (8–10) setiform, thin, smooth. Four pairs of rounded porose areas present, Aa (8–10) larger than A1–A3 (4–8), A3 located medial to p1. All notogastral lyrifissures, opisthonotal gland openings and circumgastric scissure distinct. Circumgastric sigillar band not observed.
Gnathosoma (Figs 3b-d) – Subcapitulum size 86–94 × 73–77. Subcapitular setae a (18–20) and m (18–20) setiform, slightly barbed, h (22–24) setiform, sparsely shortly ciliate, m thinnest. Adoral seta (10–12) setiform, barbed. Palp (53–57) with typical formula. Seta inf thickened, shortly ciliate. Postpalpal seta (4) spiniform, smooth. Chelicera (98–102) with two setiform, barbed setae (cha 41, chb 20–24).

Epimeral and lateral podosomal regions (Figs 2b, 2c) – Epimeral setal formula 3–1–3–2. Epimeral setae 4c absent, 1c minute (2), others (1b, 3c 14–16, 1a, 2a, 3a, 3b, 4a, 4b 10–12) setiform, thin, roughened. Pedotectum II divided into two scales. Circumpedal carina long, directed to pedotectum II. Discidium triangular. A semi-oval concavity located posteriorly to acetabulum IV.
Anogenital region (Figs 2b, 2c, 3a, 3e) – Five pairs of genital (g1 12, g2–g5, 8–10), one pair of aggenital (12), two pairs of anal (12) and three pairs of adanal (12) setae setiform, thin, roughened. Adanal lyrifissure distinct. Marginal porose area complete, narrowly band-like. Ovipositor elongated (110 × 41), blade (49) shorter than length of distal section (beyond middle fold; 61). Each of the three blades with four smooth setae, ψ1 ≈ τ1 (24–28) setiform, ψ2 ≈ τa ≈ τb ≈ τc (10–12) thorn-like. Six coronal setae not observed.

Legs (Figs 4a-c) – Monodactylous. Claw of all tarsi strong, slightly barbed on dorsal side, with tubercle ventrobasally. Tarsus I with longitudinal ridge distodorsally. Tibiae I and II with tubercle proximoventrally. Dorsoparaxial porose area on femora I-IV and on trochanters III, IV distinct. Proximoventral porose area on all tarsi and distoventral porose area on all tibiae not observed. Formulas of leg setation and solenidia: I (1–5–3–4–20) [1–2–2], II (1–5–3–4–15) [1–1–2], III (2–2–1–3–15) [1–1–0], IV (1–2–1–3–12) [0–1–0]; homology of setae and solenidia indicated in Table 1. Famulus of tarsus I short, erect, slightly dilated apically, inserted between solenidion ω2 and seta ft''. Femur III and genu IV with reduced setation, with two and one seta, respectively (seta l' absent). Some ventral setae (a, s, pv) on tarsi I–III thickened, with strong unilateral spines. Solenidion ω1 on tarsus I, ω1 and ω2 on tarsus II and σ on genu III bacilliform, other solenidia setiform.
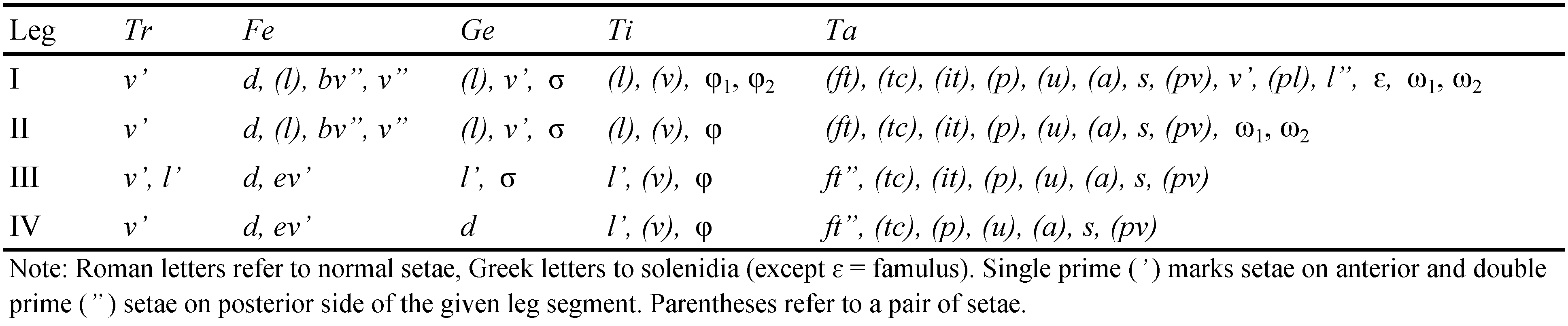

Material examined — Holotype (male) and four paratypes (one male and three females): Taiwan, Miaoli County, Dahu Township, Biological Control Branch, Miaoli District Agricultural Research and Extension Station, 24°25.416' N, 120°52.200' E, 285 m a.s.l., farm, soil under Ricinus communis (Fig. 1), 21.III.2018 (J.R. Liao & H.C. Lee).
Type deposition — The holotype is deposited in the collection of the NTU; four paratypes are deposited in the collection of the TSUMZ.
Etymology — The species name hakkai refers to the Hakka. Taiwanese Hakka people is the main population who lives in the type locality Miaoli County, Taiwan.
Remarks — Distinctive characters of the new species with the other species of the genus Perxylobates can be found in the identification key below.
Discussion
Perxylobates mahunkai was described by Bayoumi (1980) from Egypt and included in Perxylobates. However, this species differs from other representatives of the genus by the presence of a developed anterior notogastral margin (versus anterior notogastral margin completely absent in Perxylobates – generic characteristic). All morphological traits of P. mahunkai correspond to those of the genus Protoribates Berlese, 1908 (e.g. Weigmann et al. 1993; Bayartogtokh 2010), therefore we propose the following combination: Protoribates mahunkai (Bayoumi, 1980) n. comb.
Perxylobates mayuloeus Corpuz-Raros, 1979 from the Philippines was included in Setoxylobates Balogh & Mahunka, 1967 by Subías (2004), however, we support its initial position in Perxylobates (this was discussed earlier in Ermilov and Liao 2020).
Hence, at present, the genus Perxylobates currently comprises 15 species.
Key to known species of Perxylobates
We exclude Perxylobates longissimus (Warburton, 1912) (Seychelles) from the key because this species has been very briefly and insufficiently described.
1. Bothridial seta with developed head
...... 2
— Bothridial seta setiform
...... 4
2. Lamellar seta inserted medial to end of lamella and distanced from this; rostrum truncate; body size: 339 × 194
...... Perxylobates mayuloeus Corpuz-Raros, 1979. Distribution: Philippines.
— Lamellar seta inserted very close to end of lamella; rostrum rounded
...... 3
3. Five pairs of genital setae; epimeral seta 1c minute; epimeral and aggenital setae roughened; body size: 332–398 × 190–215
...... Perxylobates hakkai n. sp. Distribution: Taiwan.
— Four pairs of genital setae; epimeral seta 1c comparatively long; epimeral and aggenital setae heavily barbed; body size: 340–413 × 174–214
...... Perxylobates coreanus Choi and Aoki, 1993. Distribution: Corea, Taiwan.
4. Epimere I with strong neotrichy (eight to nine pairs of setae); body size: 350–425 × 205–265
...... Perxylobates sinensis Wen, Aoki and Wang, 1984. Distribution: Southeast China.
— Epimere I without neotrichy (three pairs of setae)
...... 5
5. Lamellar seta thickened
...... 6
— Lamellar seta setiform
...... 9
6. Interlamellar seta long (not shorter than lamellar seta); epimeral setae 1b and 1c heavily barbed; body size: 445–495 × 210–230
...... Perxylobates paravermiseta Mahunka, 1976. Distribution: Hong-Kong.
— Interlamellar seta of medium length (distinctly shorter than lamellar seta); epimeral setae 1b and 1c not heavily barbed
...... 7
7. Insertions of lamellar setae located very close to each other; rostrum truncate; body size: 398–442 × 199–221
...... Perxylobates vietnamensis (Jeleva and Vu, 1987). Distribution: Vietnam.
— Insertions of lamellar setae distanced from each other; rostrum rounded
...... 8
8. Rostral seta thickened; body surface not foveolate; body size: 381–448 × 166–215
...... Perxylobates crassisetosus Ermilov and Anichkin, 2011. Distribution: Vietnam.
— Rostral seta setiform; body surface foveolate; body size: 338–363 × 156–175
...... Perxylobates vermiseta (Balogh and Mahunka, 1968) (see also Hammer 1972). Distribution: Pantropical region.
9. Lamella as long as length of prodorsum; tutorium reaching rostrum; anterior margin of pteromorph with tooth; body size: 348–415 × 166–199
...... Perxylobates thanhoaensis Ermilov, Vũ, Trinh and Dao, 2011. Distribution: Vietnam.
— Lamella 1/2 or 2/3 length of prodorsum; tutorium distinctly not reaching rostrum; anterior margin of pteromorph without tooth
...... 10
10. Lamellar seta minute or absent, inserted on end of lamella; all setae of epimere I bushy; body length: 340
...... Perxylobates sinlimes (Hammer, 1971). Distribution: Fiji, Antilles.
— Lamellar seta comparatively long, inserted medial to end of lamella; all setae of epimere I not bushy
...... 11
11. All setae of epimere I simple; distance ro-le equal le-in; body size: 393–451 × 172–213
...... Perxylobates brevisetus Mahunka, 1988. Distribution: Vietnam, Philippines.
— All or some setae of epimere I heavily barbed; distance ro-le distinctly shorter than le-in
...... 12
12. Interlamellar seta distinctly shorter than lamellar seta; body length: 390
...... Perxylobates barbatus Hammer, 1972. Distribution: Tahiti, Java.
— Interlamellar seta not shorter than lamellar seta
...... 13
13. Subcapitular seta h and epimeral seta 1b heavily barbed, 1a and 1c simple; body size: 470–542 × 219–267
...... Perxylobates taidinchani Mahunka, 1976. Distribution: Hong-Kong, Vietnam.
—S ubcapitular seta h and all setae of epimere I (1a, 1b, 1c) heavily barbed
...... 14
14. Lamellar seta distinctly shorter than interlamellar seta; body size: 402 × 205
...... Perxylobates obesus Bayoumi, 1977. Distribution: Egypt.
— Lamellar and interlamellar setae similar in length; body size: 445–470 × 217–239
...... Perxylobates guehoi Mahunka, 1978. Distribution: Mauritius, Vietnam.
Acknowledgements
We cordially thank H.C. Lee (National Taiwan University, Taipei, Taiwan) and Miaoli District Agricultural Research and Extension Station for assistance in collecting material; C.C. Ko (National Taiwan University, Taipei, Taiwan) for supporting the study; and Dr. Julia Baumann (University of Graz, Graz, Austria) and two anonymous reviewers for valuable comments.
References
Balogh J., Mahunka S. 1967. New oribatids (Acari) from Vietnam. Acta Zool. Acad. Sci. Hung., 13(1-2): 39-74.
Balogh J., Mahunka S. 1968. New oribatids (Acari) from Indonesian soils. Opusc. Zool. Budapest, 8(2): 341-346.
Bayartogtokh B. 2010. Oribatid mites of Mongolia (Acari: Oribatida). Moscow, KMK, 372 pp.
Bayoumi B.M. 1977. Two new oribatid mites (Acari) from Egypt. Ann. Hist. Nat. Mus. Nat. Hung., 69: 297-299.
Bayoumi B.M. 1980. A new Perxylobates species from Egypt (Acari: Oribatida). Folia Ent. Hung., 33(2): 21-22.
Berlese A. 1908. Elenco di generi e specie nuove di Acari. Redia, 5: 1-15.
Choi S.S., Aoki J. 1993. Three new species of oribatid mites (Acarina, Oribatei) from Korea. Kor. Journ. Ent., 23(1): 23-29.
Corpuz-Raros L.A. (1979) Philippine Oribatei (Acarina). I. Preliminary list of species and descriptions of forty new species. Philip. Agr., 62(1): 1-82.
Ermilov S.G., Anichkin A.E. 2011. Three new species of Haplozetidae (Acari: Oribatida) from Vietnam. Acarologia, 51(1): 43-59. doi:10.1051/acarologia/20111992 ![]()
Ermilov S.G., Liao J.-R. 2018. Additions to the oribatid mite fauna of Taiwan, with description of a new species of the genus Lohmannia (Acari, Oribatida). Syst. Appl. Acarol., 23(5): 1004-1020. doi:10.11158/saa.23.5.15 ![]()
Ermilov S.G., Liao J.-R. 2020. Taxonomic study of the genus Setoxylobates (Acari, Oribatida, Haplozetidae) and some related taxa. Syst. Appl. Acarol., 25(8): 1516-1525. doi:10.11158/saa.25.8.13 ![]()
Ermilov S.G., Vu Q.M., Trinh T.T., Dao D.T. 2011. Perxylobates thanhoaensis, a new species of oribatid mite from Vietnam (Acari: Oribatida: Haplozetidae). Int. Journ. Acarol. 37(2): 161-166. doi:10.1051/acarologia/20111992 ![]()
Hammer M. 1971. On some oribatids from Viti Levu, the Fiji Islands. Det Kong. Dansk. Vidensk. Selsk. Biol. Skr., 16(6): 1-60.
Hammer M. 1972. Tahiti. Investigations on the oribatid fauna of Tahiti, and on some oribatids found on the Atoll Rangiroa. Dansk. Vidensk. Selsk. Biol. Skr., 19(3): 1-66.
Jeleva M., Vu M.Q. 1987. New oribatids (Oribatei, Acari) from the northern part of Vietnam. Acta Zool. Bulg., 33, 10-18.
Mahunka S. 1976. Neue und interessante milben aus dem Genfer Museum XVIII. Oribatiden aus Hong-Kong (Acari). Acarologia, 18(2): 360-372.
Mahunka S. 1978. Neue und interessante milben aus dem Genfer museum XXVII. A first survey of the oribatid (Acari) fauna of Mauritius, Reunion and the Seychelles I. Rev. suisse Zool., 85(1): 177-236. doi:10.5962/bhl.part.82228 ![]()
Mahunka S. 1988. A survey of the Oribatid Fauna (Acari) of Vietnam, II. Acta Zool. Hung., 34(2-3): 215-246.
Norton R.A. 1977. A review of F. Grandjean's system of leg chaetotaxy in the Oribatei (Acari) and its application to the family Damaeidae. In: Dindal D.L. (Ed.). Biology of oribatid mites. Syracuse: SUNY College of Environmental Science and Forestry. pp. 33-61.
Norton R.A., Behan-Pelletier V.M. 2009. Oribatida. Chapter 15. In: Krantz G.W., Walter D.E. (Eds.). A Manual of Acarology. Lubbock: Texas Tech University Press. pp. 430-564.
Subías L.S. 2020. Listado sistemático, sinonímico y biogeográfico de los ácaros oribátidos (Acariformes: Oribatida) del mundo (excepto fósiles) (15ª actualización). Online version accessed in January 2020, 527 pp.; http://bba.bioucm.es/cont/docs/RO_1.pdf ![]()
Travé J., Vachon M. 1975. François Grandjean. 1882-1975 (Notice biographique et bibliographique). Acarologia, 17(1): 1-19.
Warburton C. 1912. The Percy Sladen Trust Expedition to the Indian Ocean in 1905. XX. The Acarina of the Seychelles. Trans. Lin. Soc. London, 2nd ser., Zool., 15: 349-360. doi:10.1111/j.1096-3642.1912.tb00105.x ![]()
Weigmann G., Miko L. & Nannelli R. 1993. Redescription of Protoribates dentatus (Berlese, 1883) with remarks about the genus Protoribates (Acarina, Oribatida). Redia, 76(1): 39-55.
Wen Z., Aoki J., Wang X. 1984. Studies on the oribatid mites of China I. The first report on the oribatids from M.T. Sheshan, Shanghai. Contributions Shanghai Institute of Entomology, 4: 295-307.



2020-07-01
Date accepted:
2020-08-26
Date published:
2020-09-02
Edited by:
Baumann, Julia

This work is licensed under a Creative Commons Attribution 4.0 International License
2020 Ermilov, Sergey G. and Liao, Jhih-Rong
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)