



The family Opilioacaridae (Acari: Parasitiformes) in Argentina, with description of two new species
Vázquez, Maria Magdalena1 ; Bernardi, Leopoldo Ferreira de Oliveira2 and Klompen, Hans3
1Universidad de Quintana Roo, Division de Ciencias e Ingenierias, Chetumal, Quintana Roo, Mexico.
2Bolsista do Programa Nacional de Pós-Doutorado (PNPD/CAPES), Departamento de Entomologia, Universidade Federal de Lavras (UFLA), Lavras, Minas Gerais, Brasil.
3✉ Acarology Laboratory, Ohio State University, 1315 Kinnear Road, Columbus, Ohio 43212-1192, U.S.A.
2020 - Volume: 60 Issue: 3 pages: 505-519
https://doi.org/10.24349/acarologia/20204380ZooBank LSID: 522A7ACE-B100-4B92-8A80-72E17A6F59B7
Original research
Keywords
Abstract
Introduction
The first species of Opilioacaridae described from the New World was Opilioacarus platensis Silvestri 1905, based on seven specimens collected from Salto, Uruguay (31.4167°S 57.0000°W) and Posadas, Misiones, Argentina (27.3667°S 55.9000°W) (Silvestri, 1905). The species was later reclassified as Neocarus platensis (Vázquez and Klompen, 2009). As expected given its age, the description of N. platensis lacks some detail in terms of relevant morphological characters, specifically for the ovipositor, but a more troubling item is that the author also failed to designate a holotype among the specimen series studied. This could be important because Silvestri apparently did not recognize any differences among the specimens from the two localities, an issue amplified by Van der Hammen, who redescribed O. platensis based on material from a third site, Sinimbu (listed as Sinumbu), Rio Grande do Sul, Brazil (29.500°S, 52.5000°W) (Hammen, 1969). Studies of Opilioacaridae in Mexico, Madagascar, and Brazil (Vázquez and Klompen, 2002; 2009; 2010; Bernardi et al., 2012; Bernardi et al., 2013; Araújo et al., 2018; Bernardi and Borges-Filho, 2018) have shown that many species of Opilioacaridae have quite localized distributions. On the other hand, the north temperate species N. texanus Chamberlin and Mulaik, 1942, may have a very broad range, from Central Texas, west to Arizona (both U.S.A.), and south to Guerrero, Mexico (Vázquez and Klompen, 2015), sites separated by more than 1000 km. This leaves two unanswered questions: 1) are the specimens collected from these three disparate south temperate localities (separated from each other by 410–470 km) indeed conspecific (mirroring the broad distribution of N. texanus in North America), or do they represent more than one species? 2) What is the species level diversity of Opilioacaridae in the temperate regions of South America?
The focus of this study is solely on the second question. To this end we examined material in the holdings of the Museo Argentino de Ciencias Naturales ''Bernardino Rivadavia'', Buenos Aires, Argentina. This material includes specimens of two species that are different from N. platensis (all localities), suggesting a modest radiation of Neocarus in Argentina. The goal of this study is to describe these two new species.
Materials and methods
Most material was studied as slide-mounted specimens. For this purpose, specimens were dissected, cleared in lactic acid and mounted on slides using Hoyer’s medium (Walter and Krantz, 2009). Due to the considerable size of these mites, specimens were dissected, with different parts of a single mite mounted on several different slides or using multiple cover slips on a single slide. Terminology for the palp tarsal sensilla follows Grandjean (1936) as modified by Vázquez and Klompen (2002), for the sternitogenital region we follow Klompen et al. (2015), and for the leg sensory structures Grandjean (1936), Van der Hammen (1966), and Araújo et al. (2018). Characters were processed in the data matrix development mode of vSysLab (Johnson, 2010) and were exported as proto-species descriptions.
Drawings were prepared using a Zeiss Axioscope 3 phase contrast microscope, connected to a drawing tube, measurements were taken using an ocular micrometer and are presented in micrometers (μm). Most measurements are summarized in Table 1.


Collections were made by hand, with specimens collected from under stones or rotting bark and preserved in alcohol. Many of the newly examined specimens were collected in 1953–1954, but they were generally in good condition. Retroactive georeferencing for older collection sites was based on GoogleEarth.
Specimen depository abbreviations: MACN: Museo Argentino de Ciencias Naturales ''Bernardino Rivadavia'', Buenos Aires, Argentina; OSAL: Ohio State University Acarology Collection, Columbus, Ohio, U.S.A. Instar abbreviations: F= female; M= male.
Taxonomy
Genus Neocarus Chamberlin and Mulaik, 1942
All specimens examined in this study are assigned to the genus Neocarus Chamberlin and Mulaik, 1942. Generic assignment is based on the following characteristics. In the adults 3 setae on the penultimate body segment (0 or <3 in all Old World taxa and Amazonacarus Vázquez et al., 2014; shared with Caribeacarus Vázquez and Klompen, 2009 and Brasilacarus Vázquez et al., 2015), 4–6 foliate setae on the palp tarsus (<7 in Caribeacarus and Brasilacarus), eupathidium zeta–1 (ζ1, the sensillum with a ''crown-like'' tip) in the main sensillar group of tarsus I (usually distal in Caribeacarus), and shiny fleshy setae with a whip-like tip absent from the palps (present in Brasilacarus).
Within Neocarus the Argentinean specimens share a few characteristics suggesting that they may be related: palp tarsus in adults carries 6 large, foliate sensilla with 4 prominent lobes (usually 3 lobes) and both pairs of genital glands in the males are distinct, large, and similar in size (generally indistinct in cleared specimens, or anterior pair much smaller than posterior).
Neocarus misiones n. sp.
ZOOBANK: 144FCF66-D9A1-4C22-AE7E-F45E462FD7B1 ![]()
Figures 1–4, 5A–B, 6–7, 8A
Diagnosis — Palp genu with \textasciitilde8 p-type setae, palp tarsus with 6 foliate setae with 4 rounded lobes each, and 15 ch-type sensilla. Sexual dimorphism in setation of prodorsal shield absent. Sternal setae st2 and st3 with attenuate tips. Pregenital region in male with 8–9 setae, mostly thick, ribbed and blunt, but a few lightly ribbed and tapering. Pregenital region in female without setae, genital region with 12–13 thick, lightly ribbed and barbed, tapering setae. Genital region in male with 7–9 barbed, lightly ribbed, tapering setae. Ovipositor without terminal sensilla but with three very distinct rounded terminal lobes and a bilobed membranous cover.
Description — Description based on 4 females and 4 males. Immatures not available.
Gnathosoma

Chelicera (Figure 1A). Basal segment in adults with 1 seta (cht), fixed digit with 3, one of those (ch1') large, thick. Seta cht on basal segment of male chelicera shorter than seta ch1'' on fixed digit. Setae ch2', ch2'' and cht in females and males simple, with an attenuated tip, lightly serrate. Number of the ventral denticles on movable digit in both female and male one (Figure 1A, arrow). Axial scale-like processes on movable digit in both female and male absent.
Subcapitulum (Figure 1B). All four pairs of paralabial setae present: pl1 relatively large, conical, With’s organ (pl2) membranous and discoid with a biramous core; rutellum (pl3) with one row of 5 teeth, inserted dorso-lateral; pl4 small, inserted dorsal. With 4 circumbuccal (cb) and 8–11 median and subcapitular (vm, lvm, ldm, vp, lvp) setae. Seta vm1 on the male subcapitulum present. Setae vm1 plus one more lateral setal pair with blunt tips in both sexes, all other subcapitular setae with attenuate tips. Females generally with more subcapitular setae (10–11) than males (8–9). Lateral lips in all instars with distinct canals (ogl1 and ogl2).

Palp (Figure 2). Trochanter with 4 large, ribbed, tapering setae (= r-type); femur with 9–13 papilliform (= p-type) and 15 r-type setae; genu with 8 p-type and 29 r-type setae; tibia with 30 long, lightly serrate and pointed setae, 27 r-type setae, and 2 long, thin, smooth sensilla. Tibia ventrally with one small indistinct gland; similar glands not observed for other palpal segments. Tibia and tarsus partially fused. Tarsus with 6 foliate (d-type), 8 v, 15 ch, and 9 sm sensilla, plus 3 solenidia (= s-type sensilla). Foliate sensilla each with 4 lobes with rounded, not filiform, tips. Modified sm3-type sensilla on male palp tarsus absent. Lyrifissures iα and iπ (not drawn) present. Pretarsus in shape of a pair of well-developed, smooth, sessile claws.
Idiosoma
Color. Dark blue and violet stripes on both body and legs. Body often with brownish background reflecting ingested food.

Dorsum. Prodorsal shield in adults with 142(M)–164(F) setae (N=1 for both) and two pairs of lateral eyes. One pair of prodorsal lyrifissures present (Fig 3A, grey arrow). Setae somewhat rounded and ''puffed-up'' (Figure 3, detail). Sexual dimorphism in setal shape or setal arrangement minimal (setae in female appear slightly larger). Dorsal idiosoma between the shield and the preanal segment without setae, but with numerous lyrifissures arranged in transverse rows. Setation of preanal segment limited to 1 dorsal, and 2 ventro-lateral setae. Anal valves with 10–16 setae each, with females showing higher numbers.

Sternitogenital region (Figure 4). Sternal verrucae in adults each with 3–4 large, serrate and pointed, and 1 composite (St1) seta. Setae St1 subequal in size to St5. Setae St2 and St3 in females and males barbed, tapering to a fine tip. Remaining sternal region with 4–6 pairs of stout, ribbed, setae with rounded tips. Pregenital capsules each with 1 long, tapering seta (St5) and 4–6 stout, ribbed, blunt-tipped setae. Pregenital and genital areas in female with, respectively, 0 and 12–13 sturdy, lightly ribbed and barbed, tapering setae (Figure 5A). Male with 8–9 setae in pregenital region, mostly thick, ribbed and blunt, but a few lightly ribbed and tapering. Genital region 7–9 barbed, lightly ribbed, tapering setae (Figure 5B–C). Ovipositor (Figure 6) without terminal setiform sensilla, but with three very distinct roundish terminal lobes. Without well sclerotized internal structures. Terminal area in one female (the only one where this area was fully visible) covered by a bilobed membrane/tectum (Figure 6A, arrow). Male genital valves rectangular to curved, not triangular. Male glands well developed, both pairs of similar size (Figure 7).



Legs
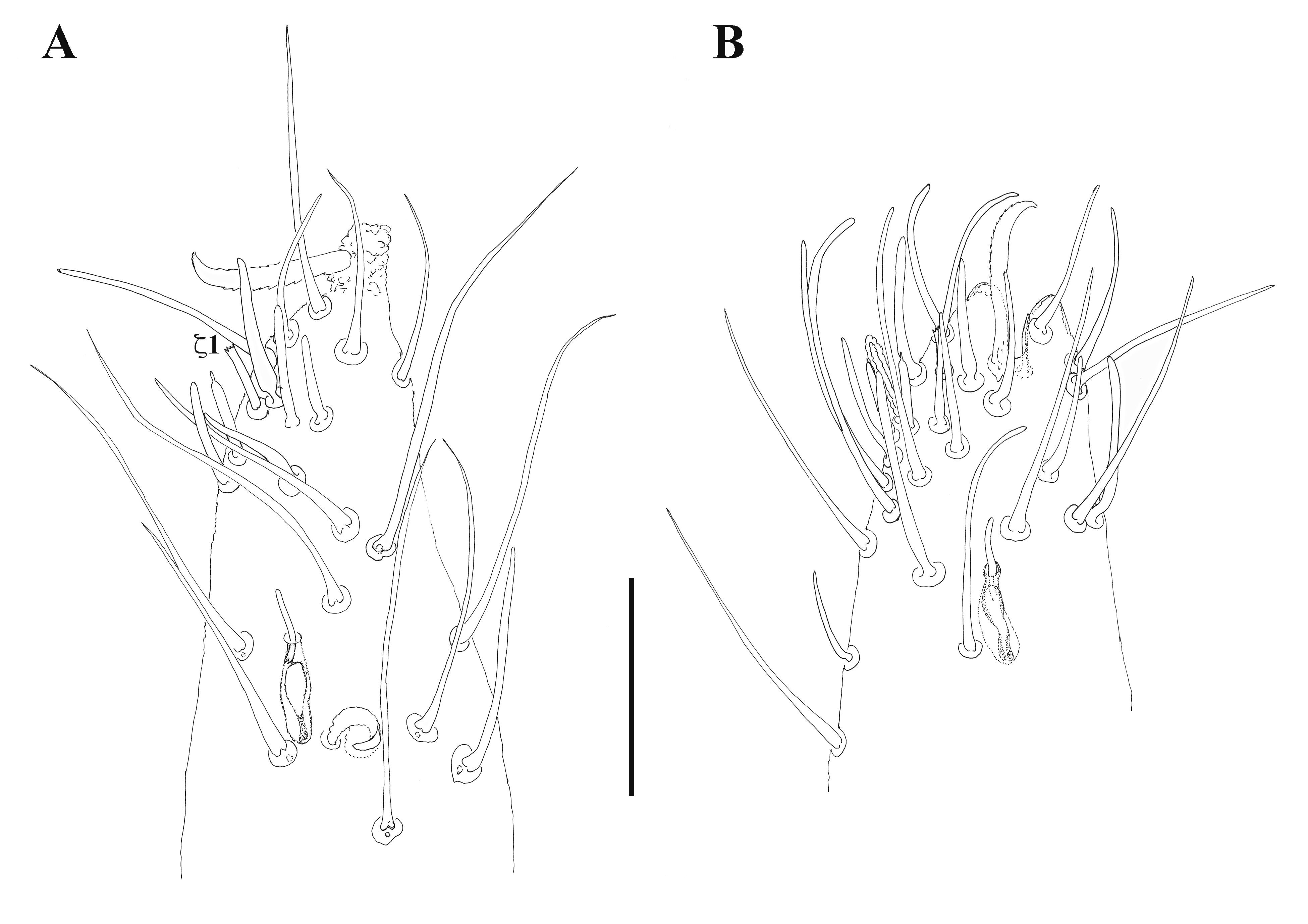
Length of legs I–IV in females, respectively, 3937–4200, 2185–2302, 2106–2367, and 3240–3390 (N=3), for males slightly shorter, respectively 3114–4162, 2025–2315, 2184, and 3065 (N=1–3). Ratio of legs I to idiosoma in female approximately 1.8–1.9, legs IV to idiosoma about 1.5–1.7. Eupathidium ζ1 on tarsus I inserted in dorsal sensory field; simple, without enlarged tip (Figure 8A). Smooth setae on tibia, genu and femur I absent in the male. Solenidion ωa on legs II positioned on acrotarsus; ωa on tarsi III–IV absent. Solenidion ωd on basitarsi II–IV inserted in apical portion, partially in a cup inside the segment. Tip of setae ld of ambulacra II–IV in male smooth and attenuate. Ventral portion of acrotarsi II–IV with 3 pairs of setae; lateral portion with 2 pairs distinctly lateral, plus 1 pair of ventro-lateral and one pair of dorso-lateral setae. Setae lv of acrotarsi II–IV with one, rarely two, small barbs. Papilliform setae on dorsal portion of the basitarsi II–III present; thin, smooth setae present in male, absent in female. Coronidia present on basitarsi II–IV in all adults, absent on tibiae and genua II–IV. The condition of the specimens (poor clearing) prevented making accurate counts of the smooth setae and coronidia.
Collection information — Type depository. Holotype female, MACN-AR 41730 (5 slides), MACN. Paratypes in OSAL (MACN-AR 4031/7) and MACN (all other).
Locality data. Holotype female, MACN-AR 41730: Argentina, Misiones, Iguazu, Puerto Libertad, 173m, 25.9189°S 54.5818°W, Oct 1954, coll. Schiapelli and De Carlo, no data on habitat. Same collection data: one female, MACN-AR 28898/3 (4 slides), one male MACN-AR 28898/2 (3 slides). Same locality, Oct 1953, coll. De Carlo, Schiapelli, Viana and Galiano, ex area with palm trees: two females, MACN-AR 4031/5 (2 slides), MACN-AR 4031/6 (1 slide), two males, MACN-AR 4031/7, MACN-AR 4031/8 (1 slide each).
Etymology — The specific name is derived from the primary collection locality, Misiones province, Argentina.
Comparative notes — This comparison is limited to described Neocarus species from South America, with exception of N. ojasti Lehtinen, 1980. The description of N. ojastii does not provide sufficient detail for an adequate comparison. Comparisons with N. platensis are based on the original description by Silvestri (1905), the re-description by Van der Hammen (1969) based on Brazilian material, and comments and notes by Marcel Santos de Araújo based on examination of the type series.
Neocarus misiones differs from N. coronatus Araújo et al., 2018 by the absence of distinct sexual differentiation in the setation of the prodorsal shield, and the presence of 6, rather than 4, d setae on the palp tarsus; from N. potiguar Bernardi et al., 2012, N. proteus Bernardi et al., 2013, and N. platensis by the absence, vs. presence, of pregenital setae in the female (Hammen, 1969; Araújo, pers. comm.). It differs from N. caipora Bernardi et al., 2014 and N. spelaion Bernardi and Borges-Filho, 2018 by the different shape of the genital setae in the female (sturdy and ribbed vs. thin and smooth), and the presence of 4, rather than 3 lobes on the d setae on the palp tarsus (N. caipora only).
Neocarus entrerios n. sp.
ZOOBANK: 19D08729-13CF-4DC3-8EDF-9BFED58D6B9E ![]()
Figures 5D–F, 8B, 9–13
Diagnosis — Palp genu without p-type setae, tarsus with 6 pairs of foliate setae with four rounded lobes each and 20 ch-type sensilla. Sexual dimorphism in setation of prodorsal shield distinct. Sternal setae St2 and St3 with attenuate tips. Pregenital area in males area with 6–10 stout, ribbed setae with relatively blunt tips, in genital area with 8–10 thinner, ribbed and tapering, setae. Females lacking pregenital setae, in genital area with 6–12 ribbed and tapering setae. Ovipositor lacking terminal sensilla, but with a very distinct pair of internal papillate hooks.
Description — Based on 4 females and 3 males. Immatures unknown.
Gnathosoma
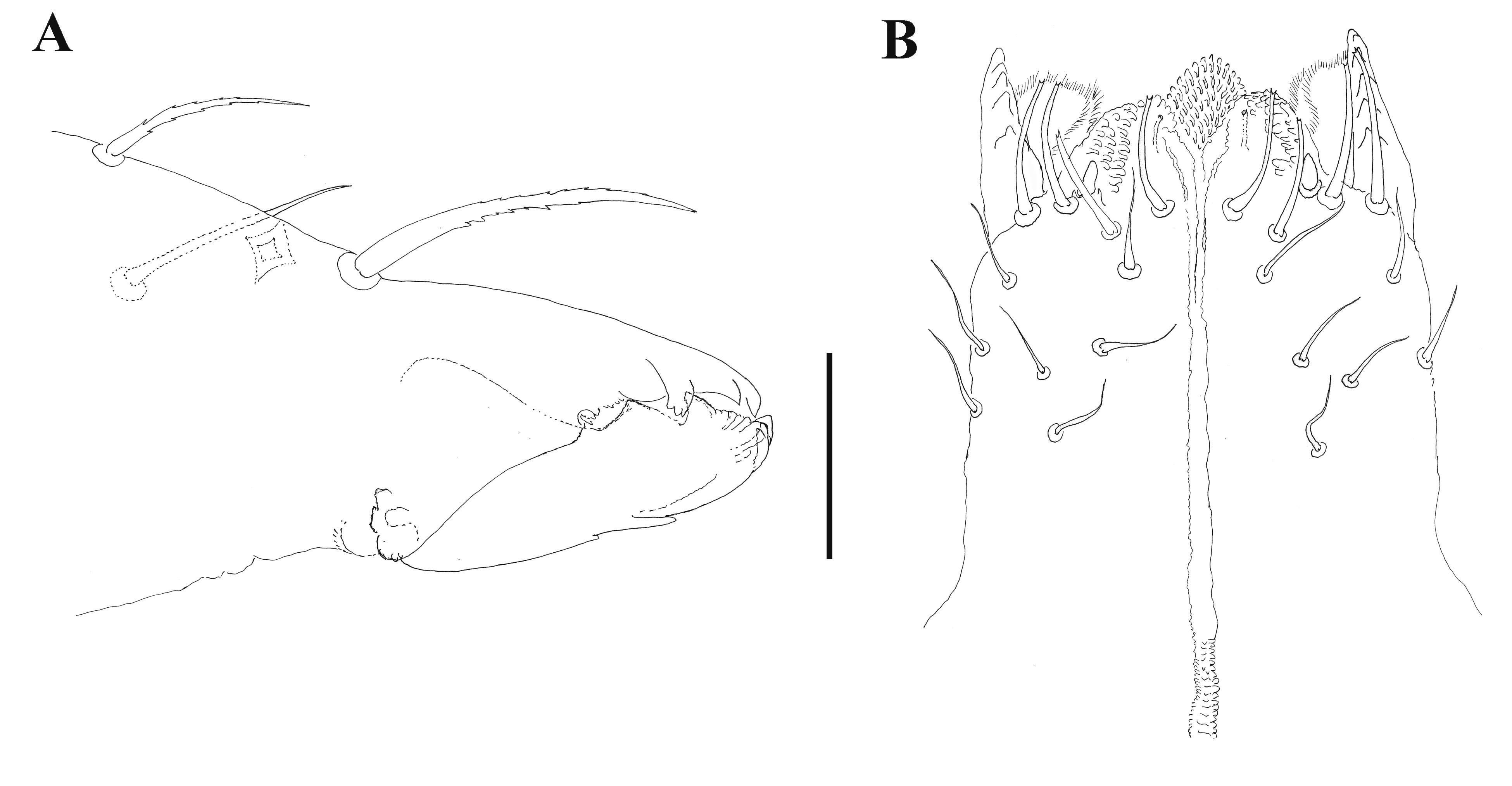
Chelicera (Figure 9A). Basal segment in adults with 1 seta, fixed digit with 3, one of which (ch1'') distinctly larger. Seta cht on basal segment of male chelicera shorter than seta ch1’’ on fixed digit. Setae ch2’, ch2’’ and cht in males simple, with an attenuated tip. One large, somewhat blunt, ventral denticle on movable digit in all adults. Axial scale-like processes on movable digit in both adults absent.
Subcapitulum (Figure 9B). All four pairs of paralabial setae present: pl1 relatively large, conical, With’s organ (pl2) membranous and discoid with a biramous core; rutellum (pl3) with one row of 5 teeth, inserted dorso-lateral; pl4 small, inserted dorsal. With 4 circumbuccal (cb) and 6–7 median and subcapitular (vm, lvm, ldm, vp, lvp) setae. Seta vm1 on the male subcapitulum present. Lateral lips with distinct canals (ogl1 and ogl2).
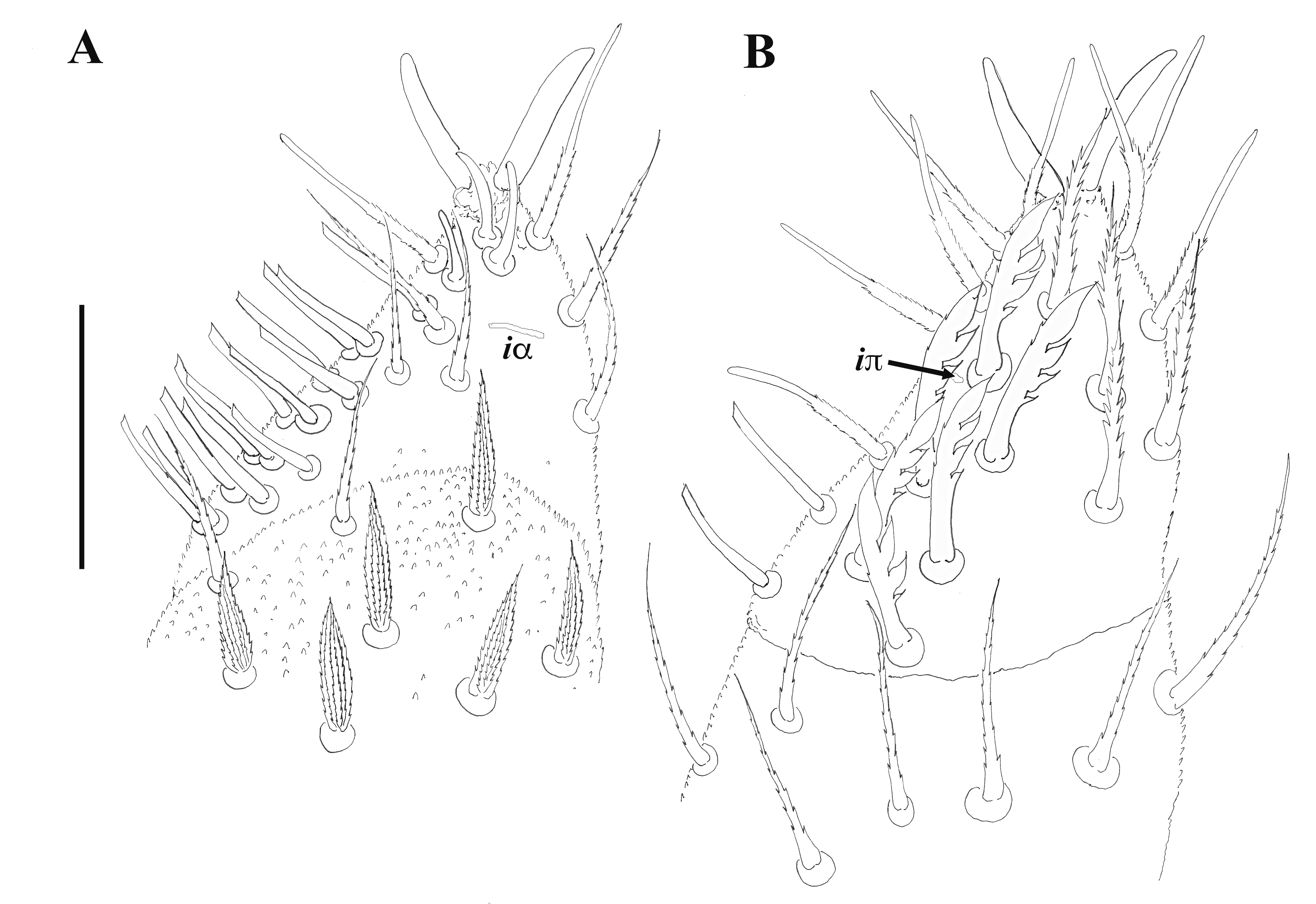
Palp (Figure 10). Trochanter with 3–4 ribbed, tapering setae (= r-type); femur with 4–9 (6–9 in F; 3–6 in M) papilliform (= p-type) and 13 r-type setae; genu with 0 p-type and 43 r-type setae; tibia with 21 long, lightly serrate and pointed setae, 38 r-type setae, and 2 long, thin, and smooth sensilla. Tibia and genu ventrally with a small indistinct gland. Tibia and tarsus partially fused. Tarsus with 6 foliate (d-type), 10 v, 20 ch, and 10–12 sm sensilla, plus 3 solenidia (= s-type sensilla). Foliate setae each with 4 lobes and rounded, not filiform, tips. Modified sm3-type sensilla on male palp tarsus absent. Lyrifissures iα and iπ distinct. Pretarsus in shape of a pair of well-developed sessile claws.
Idiosoma
Color: Violet-blue with the usual banding pattern. Color observed for alcohol preserved specimens only.

Dorsum. Prodorsal shield with two pairs of lateral eyes. One pair of prodorsal lyrifissures present. Setation in females and males consisting of, respectively, 186–204 and 218–242 setae. Sexual dimorphism in anterior portion of prodorsal shield (between anterior margin and lyrifissures) distinct, with a dense grouping of 56–62 setae in males (Figure 11A) vs. 22–24 in females (Figure 11B). Setal density in anterior area of females similar to that on the remaining shield. All setae somewhat rounded in appearance and ''puffed-up'' (Figure 11, detail). Dorsal idiosoma between the prodorsal shield and the preanal segment without setae, but with numerous lyrifissures arranged in transverse rows. Setation preanal segment limited to 1 dorsal, and 2 ventro-lateral setae. Anal valves with 12–16 stout, ribbed setae (14–16 in females; 12–16 in males).

Sternitogenital region (Figure 12). Sternal verrucae in adults each with 2–3 large pointed, and 1 composite (St1) setae. Setae St1 subequal in size to St5. Setae St2 and St3 in females and males barbed, tapering to a fine tip. Remaining sternal region with 4–6 pairs of stout, ribbed setae with blunt tips. Pregenital capsules each with 1 long, tapering seta (St5) and 6–8 (female) or 5–6 (male) stout, ribbed setae. Pregenital and genital areas in female with, respectively, 0 and 6–12 smooth, tapering setae with rounded tip (Figure 5D), male with, respectively, 6–10 stout, ribbed, and relatively blunt-tipped setae and 8–10 ribbed and tapering setae (Figure 5E–F). Ovipositor without terminal setiform sensilla, but ventrally with a pair of papillate, retrorse spines connected to a papillate ridge (Figure 13A, arrow). More dorsally two sets of sclerotized ridges, with the middle one connecting to form a W-shaped structure (Figure 13B, arrow). Male genital valves rectangular or curved, not triangular.

Legs
Length of legs I–IV in females and males very similar, combined as ''adults''. Average length and range, respectively, 3045 (2851–3269), 1596 (1490–1708), 1587 (1462–1691), and 2543 (2435–2789) (N=10, 10, 9, and 11). Ratio of legs I to idiosoma 1.4–1.6, legs IV to idiosoma 1.1–1.4. Eupathidium ζ1 on tarsus I inserted in dorsal sensory field; simple, without enlarged tip (Figure 8B). Solenidion ωa on legs II positioned on acrotarsus; ωa on tarsi III–IV absent. Solenidion ωd on basitarsi II–IV inserted apically, not sunk into the segment. Tip setae dl on ambulacra II–IV in adults smooth and attenuate. Ventral portion of acrotarsi II–IV with 3 pairs of setae; lateral portion with 2 pairs distinctly lateral, plus 1 pair of ventro-lateral and one pair of dorso-lateral setae. Setae lv of acrotarsi II–IV with one small barb, rarely two. Papilliform setae on dorsal portion of the basitarsi II–III present. Thin, smooth setae on basitarsus, tibia, genu, and (occasionally) femur I absent in female, present in male. Numbers of these setae highly variable among individuals (basitarsus 11–16, tibia 26–46, genu 0–13; N=3). Coronidia present on basitarsi II–IV of all adults (basitarsus II 8–13; III 10–12, IV 15–18; N=4), absent on tibiae and genua II–IV.
Collection information — Type depository. Holotype female, MACN-AR 41735. Paratypes in OSAL (OSAL 114547) and MACN (all other).
Locality data. Holotype female, MACN-AR 41735: Argentina, Entre Rios, Dept. Colón, Arroyo Palmar, Parque Nacional El Palmar, 26m, 31.8930°S 58.2384°W, 7 Aug 2011, Ramírez, M. J. et al., ex pasture with palm trees, low riparian forest. Same collection data: three females (MACN-AR 28895/5-6, OSAL 114547), three males (MACN-AR 28895/1–3).
Etymology — The specific name is derived from the primary collection locality, Entre Rios province, Argentina.
Comparative notes — Neocarus entrerios differs from N. potiguar, N. proteus, and N. platensis from Brazil by the absence of pregenital setae in the female. It differs from N. coronatus by the presence of 6 (vs. 4) foliate setae on the palp tarsus; from N. potiguar, N. caipora, N. platensis (from Argentina and Uruguay) and N. misiones by the absence (vs. presence) of p-type setae on the palp genu (N. proteus and N. spelaion are somewhat intermediate as they carry small numbers (1–7) of p-type setae on the palp genu), and from N. spelaion by the uniformity (vs. variability) in shape of the pregenital and genital setae in the male. Neocarus entrerios differs from N. misiones by the shape and number of genital setae in the female, the structure of the ovipositor, and the presence (vs. absence) of sexual differentiation in the setae on the prodorsal shield.
Discussion
The diversity of Opilioacaridae in Argentina is likely to be higher than indicated by the three species (including the two new ones) currently recorded from that country. We have been able to study a few specimens from other localities, and it appears that these may represent additional species. Unfortunately, all collections involve singletons, insufficient for an adequate description. The collection data for these specimens is listed to provide a better record of known diversity in the group in Argentina. Notably, most records so far are all from the northern and eastern border areas (Figure 14), suggesting Opilioacaridae may not penetrate deep into Argentina.

Records of undescribed Neocarus spp. from Argentina:
Argentina, Misiones, P.N. Iguazu, Area de la Garganta del Diablo, 185m, 25.7000°S 54.4500°W, 19–20 Jan 2005, Grismado, C., Lopardo, L., Piacentini, L., Quaglino, A. and Rubio, G., collection event no. MACN-AR 28896; 1 female.
Argentina: Misiones, Dto Cainguás, Parque Provincial Salto Encantado, 26.9500°S 54.8724°W, 10–12 Jan 2005, Grismado, C., Lopardo, L., Piacentini, L., Quaglino, A. and Rubio, G., ex collection event no. MACN-AR 28897; 1 unidentified instar.
Argentina, Salta, Salvador Mazza (formerly Pocitos), 625m, 22.0560°S 63.6870°W, 22 Nov 1954, ex collection event no. MACN-AR 28899; 1 female.
Acknowledgements
We thank Drs. Martin J. Ramírez and Andrés Porta, Museo Argentino de Ciencias Naturales ''Bernardino Rivadavia'' for making the material available, and Marcel Santos de Araújo for his comments on the type specimens of N. platensis. We thank Consorcio de Universidades Mexicana (CUMEX) for support to MMV during her stay at Ohio State University, and Capes for the granting of the doutorate funding (PDSE 1202-12-0 and doctorate scholarship) to LFOB.
References
Araújo M.S.d., Bichuette M.E., Bauchan G.R., Ochoa R., Feres R.J.F. 2018. A new species of cave dwelling Neocarus (Acari: Opilioacaridae) from Bahia state, Brazil, with remarks on taxonomic characters. Zootaxa, 4402: 303-322. doi:10.11646/zootaxa.4402.2.4 ![]()
Bernardi L.F.d.O., Borges-Filho E.L. 2018. Neocarus spelaion sp. n. (Parasitiformes, Opilioacaridae), a new species of cave dwelling Neocarus from Minas Gerais state, Brazil. Subterranean Biol., 27: 1-16. doi:10.3897/subtbiol.27.25777 ![]()
Bernardi L.F.d.O., Klompen H., Zacarias M.S., Ferreira R.L. 2013. A new species of Neocarus Chamberlin & Mulaik, 1942 (Opilioacarida: Opilioacaridae) from Brazil, with remarks on postembryonic development. Zookeys, 358: 69-89. doi:10.3897/zookeys.358.6384 ![]()
Bernardi L.F.d.O., Zacarias M.S., Ferreira R.L. 2012. A new species of Neocarus Chamberlin & Mulaik, 1942 (Acari: Opilioacarida) from Brazilian caves and karst areas. Zootaxa, 3416: 53-68. doi:10.11646/zootaxa.3416.1.5 ![]()
Grandjean F. 1936. Un acarien synthétique: Opilioacarus segmentatus With. Bull. Soc. Hist. Nat. Afr. Nord, 27: 413-444.
Hammen L., van der. 1966. Studies on Opilioacarida (Arachnida). I. Description of Opilioacarus texanus (Chamberlin & Mulaik) and revised classification of the genera. Zool. Verhand., Leiden, 86: 3-80.
Hammen L., Van der. 1969. Studies on Opilioacarida (Arachnida). III. Opilioacarus platensis Silvestri, and Adenacarus arabicus (With). Zool. Meded., Leiden, 44: 113-131.
Johnson N.F. 2010. Future taxonomy today: new tools applied to accelerate the taxonomic process. In: Polaszek A., (Ed). Systema Naturae 250: The Linnaean Ark. London: CRC Press Taylor & Francis Group. p. 137-147. doi:10.1201/EBK1420095012-c13 ![]()
Klompen H., Vázquez M.M., Bernardi L.F.d.O. 2015. Post-embryonic development in the mite suborder Opilioacarida, with notes on segmental homology in Parasitiformes. Exp. Appl. Acarol., 67: 183-207. doi:10.1007/s10493-015-9939-7 ![]()
Silvestri F. 1905. Note Aracnologische. I-III. Redia, 2: 239-261 + plates XXI-XXIV.
Vázquez M.M., Klompen H. 2002. The family Opilioacaridae (Acari: Parasitiformes) in North and Central America, with description of four new species. Acarologia, 42: 299-322.
Vázquez M.M., Klompen H. 2009. New species of New World Opilioacaridae (Acari: Parasitiformes) with the description of a new genus from the Caribbean region. Zootaxa, 2061: 23-44. doi:10.11646/zootaxa.2061.1.2 ![]()
Vázquez M.M., Klompen H. 2010. The genus Salfacarus (Acari: Opilioacarida) in Madagascar. Zootaxa, 2482: 1-21. doi:10.11646/zootaxa.2482.1.1 ![]()
Vázquez M.M., Klompen H. 2015. The family Opilioacaridae (Parasitiformes: Opilioacarida) in Mexico, description of two new species and notes on biology and geographical distribution. Zootaxa, 3957: 535-552. doi:10.11646/zootaxa.3957.5.3 ![]()
Walter D.E., Krantz G.W. 2009. Collecting, rearing and preparing specimens. In: Krantz G.W., Walter D.E., (Eds). A manual of acarology. Lubbock, TX: Texas Tech University Press. p. 83-95.



2019-11-26
Date accepted:
2020-06-03
Date published:
2020-06-16
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2020 Vázquez, Maria Magdalena; Bernardi, Leopoldo Ferreira de Oliveira and Klompen, Hans
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)