



A new species of the genus Lasioseius (Acari: Blattisociidae) inhabiting litter of secondary rainforest in Sumatra, Indonesia
Quintero-Gutiérrez, Edwin Javier  1
; Sandmann, Dorothee
2
; Cómbita-Heredia, Orlando
3
; Klarner, Bernhard
4
; Widyastuti, Rahayu5
and Scheu, Stefan
6
1
; Sandmann, Dorothee
2
; Cómbita-Heredia, Orlando
3
; Klarner, Bernhard
4
; Widyastuti, Rahayu5
and Scheu, Stefan
6
1✉ Instituto Colombiano Agropecuario (ICA), Subgerencia de Análisis y Diagnóstico, Grupo Red de Análisis y Diagnóstico Fitosanitario, Laboratorio de Análisis y Diagnóstico Fitosanitario de Caldas, Carrera 30 # 65-15, Manizales, Colombia.
2University of Göttingen, J.F. Blumenbach Institute of Zoology and Anthropology, Untere Karspüle 2, 37073 Göttingen, Germany.
3Acarology Laboratory, Ohio State University, 1315 Kinnear Rd., Columbus, OH 43212, U.S.A & Centro de Investigación en Acarología, Bogotá, Colombia.
4University of Göttingen, J.F. Blumenbach Institute of Zoology and Anthropology, Untere Karspüle 2, 37073 Göttingen, Germany.
5Institut Pertanian Bogor, Department of Soil Science and Land Resources, Damarga Campus, Bogor 16680, Indonesia.
6University of Göttingen, J.F. Blumenbach Institute of Zoology and Anthropology, Untere Karspüle 2, 37073 Göttingen, Germany & University of Göttingen, Centre of Biodiversity and Sustainable Land Use, Büsgenweg 1, 37077 Göttingen, Germany.
2020 - Volume: 60 Issue: 2 pages: 338-352
https://doi.org/10.24349/acarologia/20204371ZooBank LSID: 6BB13B33-4183-44C5-8216-25B6FC42FF5C
Original research
Keywords
Abstract
Introduction
Currently, the mite family Blattisociidae (Mesostigmata: Ascoidea) comprises 14 globally distributed genera with Lasioseius Berlese 1916 as the most diverse and rich genus. Lasioseius includes a total of 206 described species that have been found in different habitats such as soil, grass, moss and litter, but also associated with insects and on plants (Moraes et al. 2016). Some species have been widely studied as potential biological control agents (Moraes et al. 2015).
Up to date, nine species from five genera of Blattisociidae have been reported for Indonesia i.e., Blattisocius dentriticus (Berlese, 1918) from Ambon Island, Cheiroseius alstoni (Evans and Hyatt 1960) from Bogor, Cheiroseius parapodicus (Berlese, 1916), Hoploseius cometa (Berlese, 1910), Lasioseius sublaevis Berlese, 1916, and Platyseius mollicomus (Berlese, 1916) from Java, Lasioseius alter Vitzthum, 1925, Lasioseius frontalis Evans and Sheals, 1959 and Lasioseius polydesmophilus Evans and Sheals, 1959 from Sumatra. Only C. alstoni and L. alter have been collected from soil in decaying vegetation and rotting vegetable debris respectively. All the other species have been found associated with insects.
The main goal of this paper is to describe one new species of the genus Lasioseius (Lasioseius) based on adult specimens (female and male) collected from litter of secondary rainforest in Bukit Duabelas National Park, Jambi Province, Sumatra, Indonesia.
Materials and methods
This study forms part of a large project investigating arthropods of Indonesia, within the framework of the interdisciplinary project ''Ecological and socioeconomic functions of tropical lowland rainforest transformation systems (Sumatra, Indonesia)'' – EFForTS. For more details on the study region and the experimental design see Drescher et al. (2016).
Mites were extracted from soil samples of 16 x 16 cm taken from the core plots of the project with a spade in the Bukit Duabelas National Park, Jambi Province, Sumatra (for exact localities and habitats see ''Material examined'' section). Litter and topsoil to a depth of 5 cm were extracted separately using the high gradient canister method described by Kempson et al. (1963). All specimens were collected in November 2013 by B. Klarner. Mites were initially stored in 70% ethanol, then cleared in 55% lactic acid, in some cases gnathosoma was dissected from idiosoma, and slide preparations were done with Hoyer's medium.
Photographs and measurements were made using a Nikon Eclipse Ci microscope equipped with phase contrast and connected to a digital camera (Nikon, High-definition Color Camera Head DS-Fi2) controlled by (Nikon, Camera Control Sight DS-L3). Most images were taken in stacks with the focal depth controlled manually. Selected images were combined using Zerene Stacker Version 1.04. In some cases, images captured from different regions of the body were combined using the `photomerge' function in Adobe Photoshop, version 2015 (16.0 or 20150529.r.88; Adobe Systems Inc., San Jose, USA). Digital drawings were prepared with Adobe Illustrator, version CC 2015 (19.0.0), based on mite photographs.
All measurements are given in micrometers (μm) and included in the descriptions as a range (minimum–maximum). Lengths of shields were measured along their midlines; the level in which widths were measured are indicated in brackets. Leg measurements were taken from the proximal margin of the coxa, along of the midline of each segment, to the tip of the claw. Notations of body structures and idiosomal chaetotaxy generally follow Lindquist and Evans (1965). Leg chaetotaxy follows Evans (1963) and Evans and Till (1965). Notations for pore-like structures (gland pores and poroids/lyrifissures) follow the system of Athias-Henriot (1971) for ventral idiosoma and peritrematal shield, and Athias-Henriot (1975) for the dorsal idiosoma.
The holotype (female) and one paratype (male) are deposited in LIPI (Indonesian Institute of Science), Cibinong, Indonesia; the other paratypes in SMNG (Senckenberg Museum), Görlitz, Germany and OSAL (Ohio State Acarology Collection), Columbus, USA.
Results
Systematics
The genus Lasiouseuis includes two subgenera Lasioseius (Lasioseius) Berlese, 1916 and Lasioseius (Endopodalius) Christian and Karg, 2006 (Moraza and Linquist 2011). The subgenus Lasioseius (Lasioseius) includes the following four currently recognized species-groups: the phytoseioides-group (Lindquist, 1964) with twenty species, the porulosus-group (Walter and Lindquist, 1997) with seven species, the floridensis-group (Mineiro et al., 2009) with four species, and the cassidini-group with four species (Moraza and Lindquist, 2018).
Based on the key to the world genera and subgenera of Blattisociidae (Moraza and Linquist 2011) and on the diagnosis provided by Moraza and Lindquist (2018) for the species-groups, the species described here forms part of the subgenus Lasioseius (Lasioseius) and the floridensis-group by the combination of the following characters: females with most of the dorsal shield setae tricarinate and moderately long, many of them nearly as long as longitudinal intervals between their bases; posthumeral seta r4 short and simple, similar to r2, and at most half as long as neighboring tricarinate seta s4; tarsus II with one midlateral seta pl2 elongated and reaching nearly to base of pretarsus; leg IV with no tarsal setae longer than tibial segment; podonotal region of female dorsal shield with 21 pairs of setae, including r4 but excluding r5; and sternal shield with an anteromedial patch of reticula. The floridensis-group currently includes L. floridensis Berlese 1916, L. queenslandicus (Womersley, 1956), L. sugawarai (Ehara, 1964), and L. latinoamericanus Mineiro et al., 2009.
Blattisociidae
Lasioseius Berlese
Lasiosieus Berlese 1916: 33; Lindquist and Evans 1965: 40; Christian and Karg 2006: 105; Lindquist and Moraza 2010: 4.
Lasioseius orangrimbae n. sp. Quintero-Gutiérrez and Sandmann
ZOOBANK: B3F5F066-ABEB-4C0E-9B19-8AE00F85DF4E ![]()
Figures 1–9, 10a
Diagnosis
Female — Dorsal shield with reticulate surface, bearing 36 pairs of setae that are relatively long and tricarinate, except z1, s1, r2, r4, J5 which are smooth and aciculate, Z3–Z5, S4 and S5 tricarinate and with serrate margin. Soft integument bearing only seven relatively short pairs of setae r5, r6, R2–6; seta R1 and UR series absents. Ventral shields with all setae aciculate and smooth, sternal shield mostly smooth, central region slightly punctate, sparse lateral striations, and with two parallel anteromedian lines between st1 and st2; st1 in the sternal shield. Genital shield punctate (difficult to discern) with posterior margin truncate; four transversely aligned and elongated platelets between genital and ventrianal shield. Ventrianal shield strongly reticulate, bearing six pairs of preanal setae ZV2–ZV3, JV1–JV3, JV4 10–11; posterior margin slightly narrow at level of JV4. Only three pairs of setae ZV1, ZV4 and JV5 on opisthogastric integument, JV5 at least twice longer than ZV1 and ZV4. Two pairs of semi-rounded metapodal elements. Spermatheca with a short major duct leading to a weakly sclerotized shallowly funnel-shaped calyx as long as wide from which a long and thin minor duct arise. Tarsus II with pl2 macrosetae.
Male — Dorsal shield with 38 pairs of setae including r5–6 (r6 inserted sometimes on unsclerotized lateral cuticle), unsclerotized lateral cuticle with the setae R-UR series absents. Sternogenital shield with five setae st1–st5, ventrianal shield bearing six pairs of setae with six pairs of preanal setae, seta JV5 longer than other preanal setae; ZV5 in soft opisthogastric
Description
Female (Figures 1–6, 10a) (n = 5)

Idiosomal dorsum — (Figure 1). Dorsal shield 340–363 long, 219–239 wide (at level of seta S1), covering partially the idiosoma, strongly reticulate, except between the bases of Z5 and Z4 being slightly reticulate and with some granulations, reticulations less distinct centrally due to dense internal muscle insertions, lateral margins of soft integument exposed. Dorsal shield bearing 36 pairs of setae of similar length and shape; most setae relatively long and tricarinate, except pairs z1, s1, r2, r4, J5 which are smooth and aciculate; pairs of setae Z3–Z5, S4 and S5 tricarinate with serrate margin. Unsclerotized lateral cuticle bearing only seven pairs of relatively short setae of similar length r5, r6, R2-6, and a pair of lyrifissures (RP) between R3 and R4; R1 and UR series absents. Length of setae: j1 20–21, j2 18–19, j3–j4 25–26, j5 22–23, j6 23–24, z1 7–9, z2 20–2, z3 30, z4 27, z5 22, z6 22-23, s1 11–12, s2 7–9, s3 27–28, s4–s6 29–30, r2 7–8, r3 28–29, r4 7–8, r5 9–10, r6 7, J1 23–24, J2–J4 24–25, J5 11–12, Z1 28–29, Z2 29–30, Z3 36, Z4 46–47, Z5 41–42, S1–S3 25–27, R2–R3 9–10, R4–R6 11–12. Dorsal shield with 22 pairs of pore-like structures, including six pairs of gland openings (gd1–2, gd4, gd6, gd7 and gd9) and 16 pairs of poroids.

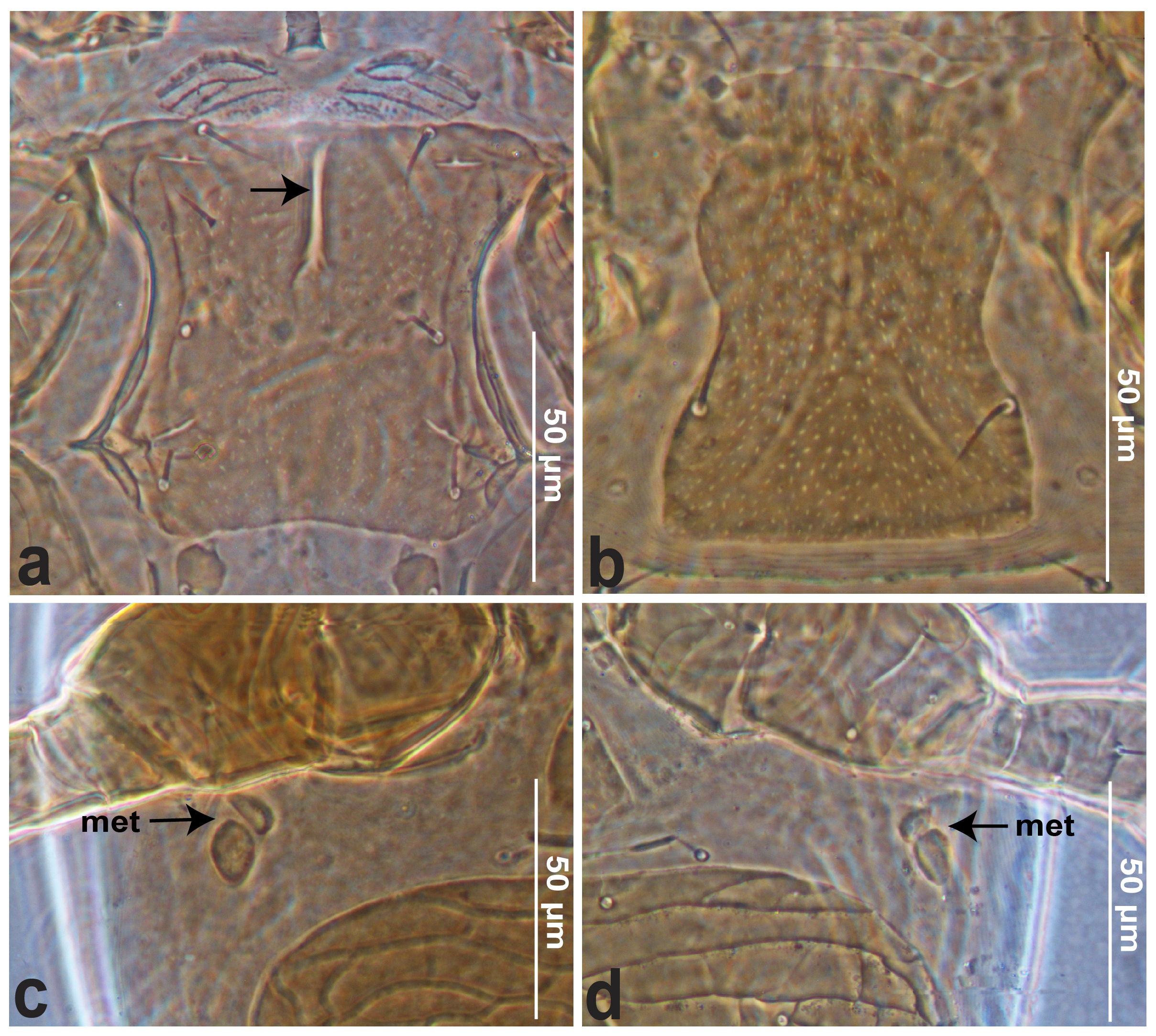
Idiosomal venter — (Figures 2, 3a–d). All setae aciculate and smooth. Tritosternum with columnar base and pair of completely free pilose laciniae. Pre-sternal region little sclerotized, punctate and transversely striate (Figures 3a). Sternal shield 253–260 long, 170–174 wide (at level of st2) mostly smooth, central region slightly punctate, difficult to discern (Figures 3a), with sparse lateral striations and with two parallel anteromedian lines between st1 and st2 (Figures 3a, see arrow), bearing three pairs of setae st1–st3 (15–16) and two pairs of slit-like poroids iv1–2, oblique. Posterior portion of endopodal plate represented by a triradiate fragment between coxae III–IV. Metasternal plates roundish, with st4 (16) and a pair of lyrifissures (iv3). Genital shield 72–75 long, 55–58 wide (at level of st5), evenly punctated, but difficult to discern (Figure 3b); posterior margin truncate, shield bearing one pair of setae st5 (15–16), slit-like poroids iv5 on unsclerotized cuticle and posterolaterad of st5; in front of the posterior margin four transversely aligned elongate platelets. Exopodal shield narrow, between coxa I–IV long and fused posteriorly (at level to the coxa IV) with the peritremal–parapodal shields. Peritrematal shield smooth, anteriorly fused with dorsal shield at level of the setae s1, fused posteriorly with the exopodal–parapodal shields; bearing pairs of poroids id3, gd3, id7, gdp and ip. Ventrianal shield wider 312–316 (at level of ZV3) than long 194–204, posterior margin slightly narrow at level of JV4; strongly reticulate and punctate anterior to the postanal seta; shield with six pairs of setae ZV2 15–16, ZV3 7, JV1 11–12, JV2 15–16, JV3 12–13 and JV4 10–11 additionally to the circumanal setae (postanal seta slightly longer 21–22 than para-anal setae 15–16). Further, with one pair of marginal pores (gv3) posterolaterad of para-anal setae and two pairs of poroids (ivo). Soft opisthogastric cuticle bearing only three pairs of setae, JV5 20-21 twice longer than ZV1 and ZV4 10–11, with two pairs of semi-rounded metapodal platelets (met) well developed (Figures 3c–d).

Gnathosoma — (Figures 4a–c). Subcapitulum (Figure 4a) corniculi well sclerotized, elongated and horn-like. Internal malae lanceolate, densely fimbriated, with apices slightly beyond of the corniculi. Deutosternum with eight transverse rows of denticles with first row (most distal), from second to seventh rows with 10-15 denticles, the first seven lines are connected by the lateral subparallel deutosternal lines, eighth line (most proximal) extending beyond lateral deutosternal lines and with 20–23 denticles; with smooth lines laterad of delineating deutosternal lines at region between fifth and sixth transverse lines; subcapitular setae smooth and aciculate, h1 and h3 32 longer than h2 23 and pc 29. Chelicerae (Figure 4b). Fixed digit with 13 small teeth plus one distal hook-like tooth, and a setiform pilus dentilus; dorsal and antiaxial lyrifissures as well as dorsal setae distinct; mobile digit with three large teeth and one large distal tooth. Gnathotectum (Figure. 4c) with three anterior projections, two serrate lateral projections and one acuminate middle process longer than the lateral ones (not serrate).

Legs — (Figures 5a–d) Lengths: I 303–307, II 255–261, III 220–224, IV 298–304. Chaetotaxy/setation—Coxae: I: 0, 0/1, 0/1, 0; II: 0, 0/1, 0/1, 0; III: 0, 0/1, 0/1, 0; IV: 0, 0/0, 0/1, 0; trochanters: I: 1, 0/1,1/2, 1; II: 1, 0/1, 0/2, 1; III: 1, 1/1, 0/2, 0; IV: 1, 1/1, 0/2, 0; femora: I: 2, 3/1, 2/2, 2; II: 2, 3/1, 2/2, 1; III: 1, 2/1, 1/0, 1: IV: 1, 2/1, 1/0, 1; genua: I: 2, 3/2, 3/1, 2; II: 2, 3/1, 2/1, 2; III: 2, 2/1, 2/1, 1; IV: 2, 2/1, 3/0, 1; tibia: I: 2, 3/2, 3/1, 2; II: 2, 2/1, 2/1, 2; III: 2, 1/1, 2/1, 1; IV: 2, 1/1, 3/1, 2; tarsi I: not counted, II: 18; III: 18; IV: 18. All legs I–IV with pretarsi, including a pair of claws and a pulvillus with median lobe rounded. Legs with all setae aciculate and smooth. Legs I-IV with most of the dorsal setae in femur–tibia slightly thicker than ventral setae. The most of setae in the tarsus I–IV moderately longer and/or stout than other segments. Tarsi II-IV with seta md 3.2 times longer than mv. Leg II with al setae in femur–tibia shorter than pl setae (al1 in femur II is long). Tarsus II with pl2 macrosetae (5.2 times longer than al2) (Figure 10a) and pd2 moderately longer than ad2. Legs III–IV with pl setae in femur–tibia shorter than pl setae. Tarsus IV with al2 slightly longer than pl2.

Spermathecal apparatus — (Figure 6). With a short major duct leading to a weakly sclerotized shallowly funnel-shaped calyx as long as wide from which a long 33 and thin minor duct arise with terminus duct indiscernible in specimens at hand.
Male (Figures 7–9) (n = 3)

Idiosomal dorsum — (Figures 7a–b). Dorsal shield 268–275 long, 178–180 wide at level of setae S1, chaetotaxy as in female but bearing two more pairs of setae (r5–6, Figure 7a) for a total 38 pairs of setae (r6 inserted on unsclerotized lateral cuticle in one of the three males, Figure 7b), other setae from R-series absent; dorsal setae slightly shorter than in female. Poroidotaxy, adenotaxy and ornamentation identical to that of female.

Idiosomal venter — (Figure 8). Similar to female except the following: Sternigenital shield 130–134 long, 58–61 wide at level between st2; bearing five pairs of simple setae (st1–5). Ventrianal shield with six pairs of setae (JV1–3, JV5, ZV1–2), excluding three circum-anal setae, JV5 longer than other preanal setae; ZV5 in soft opisthogastric integument.

Gnathosoma — (Figures 9a–b). As in female except the following: Chelicerae (Figure 9a); fixed digit with 12 small teeth plus one distally hook-like tooth and a setiform pilus dentilus; dorsal and antiaxial lyrifissures and dorsal setae distinct; mobile digit with a single tooth slightly directed forward and distally hook-like; with a slightly curved spermadactyl. Gnathotectum (Figure 9b) anteromedian region of epistome convex, with variably denticulate extensions.
Legs — Similar to those of female. Lengths: I 255–260, II 232–236, III 182–190, IV 264– 268. Tarsus II with pl2 macrosetae.

Type material and depository
Indonesia, Sumatra, holotype and all paratypes from Bukit Duabelas National Park, litter from secondary rainforest, research site BF1a, S 01°59'42.5'`, E 102°45'08.1'', 83 m a.s.l. Holotype: • 1♀, deposited at LIPI (OSAL 00124869); Paratypes: • 2♀ deposited at SMNG (OSAL 00124865 SMNG 2020 / 61895/1, OSAL 00124867 SMNG 2020 / 61895/3). • 2♀ deposited at OSAL, (OSAL 00124868, 00124870). • 1♂ deposited at LIPI, (OSAL 00124864). • 1♂ deposited at SMNG (OSAL 00124866 SMNG 2020 / 61895/2). • 1♂ deposited at OSAL (OSAL 00124863). All specimens were collected in November 2013 by B. Klarner. Additional photos of the species (ECOTAX_ID: 424404) are deposited in Ecotaxonomy database at www.ecotaxonomy.org.
Etymology
The epithet orangrimbae refers to forest-dwelling people ''Orang Rimba'' living in the sampling region.
Remarks
Differential characters: Lasioseius orangrimbae n. sp., significantly differs from the other species in the floridensis-group by the smaller body size in females and males (Females: 268–275 long, 178–180 wide, Males: 268–275 long, 178–180 wide) compared to L. floridensis (F: 433–450 long, 238–255 wide, M: 300–325 long, 190–208 wide), L. latinoamericanus (F: 402–422 long, 205–212 wide, M:302–315 long, 168–171 wide), L. queenslandicus (F: 430 long, 260 wide, M: 317-320 long, wide not reported), and L. sugawarai (F: 430 long, 270 wide, M: unknow). Additionally, the spermathecal apparatus in females of L. orangrimbae n. sp., has a sclerotized, funnel-shaped calyx that is as long as wide without conspicuous vesicle, in L. floridensis the spermathecal apparatus is sclerotized, shallowly funnel-shaped calyx with a conspicuous vesicle and a long, relatively thick minor duct; in L. latinoamericanus the spermathecal apparatus has a densely sclerotized region (embolus) with a slender funnel-shaped calyx with a conspicuous long vesicle and a very thin minor duct; in L. sugawarai the spermathecal apparatus is small, weakly sclerotized, bulbous and without discernible minor duct, and in L. queenslandicus the spermathecal apparatus is bifurcated after leaving the solenostome with a weakly sclerotized, tubular calyx and a relatively short thick minor.
Also, females of L. orangrimbae n. sp. differ from the those of the other species in the floridensis-group in having a total of seven pairs of marginal setae on soft cuticle (r5–6, R2–6) with R1 and UR setae absent, while L. floridensis, L. latinoamericanus and L. queenslandicus have eight pairs of marginal setae (r5–6, R1–6) with setae UR present, and L. sugawarai has eleven pairs of marginal setae (r5–6, R1–7) with two pair of UR setae. In addition, females L. orangrimbae n. sp., have only three pairs of setae in the opisthogastric cuticle (JV5, ZV1, ZV4), compared to L. floridensis and L. latinoamericanus which have six pairs (JV4–5, ZV3–5) and (ZV1, ZV3–4, JV4–5), respectively; L. latinoamericanus has seven pairs (ZV1, ZV3–ZV5, JV4–JV5, and a pair of submarginal UR) and L. sugawarai has five pairs (JV4–5, ZV1, ZV3–4). Finally, males of L. orangrimbae n. sp., do not have marginal setae (except r6 in one of the three examined males), whereas males in L. floridensis have five pairs of marginal setae (R2–6), L. queenslandicus two pairs (R2, R5) and L. latinoamericanus six pairs (R1–6).
Discussion
Five species of Lasioseius have been previously described from Sumatra L. sublaevis L. alter, L. frontalis, L. polydesmophilus and L. angustus. For L. sublaevis it is not possible to compare or stablish the subgenus or species group because the description is very short. Additionally, Christian and Karg (2006) did not include this species in their study. Lasioseius alter, L. frontalis, and L. polydesmophilus are not closely related to L. orangrimbae n. sp., because they belong to the subgenus Lasioseius (Endopodalius) Christian and Karg, 2006. Finally, L. angustus was described based only on males and also seems to belong to the subgenus Lasioseius (Endopodalius).
When examining our specimens with the key to the species of Lasioseius provided by Christian and Karg (2006) we ended in a similar way as reported in Britto et al. (2011). These authors reported that when they determined their specimens of L. floridensis with this key, it led them to the subgenus Crinidens, then to key 6 for their ometes-complex having five pairs of setae on the ventrianal shield. In our case ending in the subgenus Crinidens for having most dorsal setae tricarinate, then to the Lasioseius-ometisimilis-complex key 5 for having 6 setae on the ventrianal shield, and at the end to the cuplet 21-22 to the species L. laciniatus Christian and Karg, 2006, and L. tricuspidis Christian and Karg, 2006 for having the following characters: a normal number of dorsal setae (not reduced), two pairs of metapodal plates, margins of the tectum with three clear ''branches'' with medial ''branch'' as long as the lateral ones which have serrations, most of the dorsal setae tricarinate and dorsal setae not as long to reach the base of the next setae, sternal seta st1 on the sternal shield, ventrianal shield slightly wider than longer, post anal seta not long. However, the tectum shape in L. laciniatus and L. tricuspidis do not resemble the one of L. orangrimbae n. sp., in L. orangrimbae n. sp. the lateral processes of the tectum are serrate with the middle process longer than the lateral ones, whereas in L. laciniatus the lateral processes of the tectum are serrate with the middle one shorter than the lateral ones, and in L. tricuspidis the lateral processes are three-pronged with the middle one little longer than the lateral ones. Additionally, L. orangrimbae has a well-developed anteromedial patch in the sternal shield, which is absent in L. laciniatus and L. tricuspidis.
Finally, we would like to emphasize that L. orangrimbae n. sp., fits well the characters stated in Moraza and Lindquist (2018: 68, entry 4) for the floridensis-group ''Dorsal shield with post-humeral seta r4 short, simple, similar to r2, and not more than half as long as tricarinate seta s4 neighboring it medially; most dorsal setae tricarinate and moderately long, nearly as long as longitudinal intervals between their bases; anterior pair of sternal setae on weakly sclerotized presternal area, just before anterior margin of sternal shield; sternal shield with anteromedial patch of reticula; tarsus II with one midlateral seta pl2 elongated and reaching nearly to base of pretarsus'', except the character ''anterior pair of sternal setae on weakly sclerotized presternal area''. However, when we compared our specimens with the key to the species of Lasioseius in Christian and Karg (2006), we noticed that some species in the subgenus Lasioseius (Crinidens) sensu Christian and Karg, 2006 especially the species in the part of the key 5 (Lasioseius-ometisimilis-complex) number 11(14) that states ''The first pair of sternal setae localized anteriorly to the shield'' may fall in the floridensis-group because they all fit the characters of the group but some of them do not have the ''sternal shield with anteromedial patch of reticula'', and it is not clear if all species of Lasioseius-ometisimilis-complex also have ''tarsus II with one midlateral seta pl2 elongated and reaching nearly to base of pretarsus''. Examining holotype images of L. laciniatus and L. tricuspidis in VIRMISCO (Decker et al. 2018), we noticed that L. laciniatus and L. tricuspidis have the macroseta pl2 in tarsus II (Figures 10b–c). Therefore, it is not clear why the floridensis- group does not include species of the subgenus Lasioseius (Crinidens) sensu Christian and Karg (2006), or where those species would fall under the species-group key presented in Moraza and Lindquist (2018).
All the previous suggests that the subgenus Lasioseius (Crinidens) sensu Christian and Karg (2006) needs to be revised considering if certain species may belong to the floridensis-group, or if with a slight shift of characters the floridensis-group may be split or expanded to include other species.
Acknowledgements
Thanks to Bruce Halliday and Diana Rueda-Ramirez for help with literature. Thanks to the State Ministry of Research and Technology of Indonesia (RISTEK) for the research permit, the Indonesian Institute of Sciences (LIPI) and Ministry of Forestry (PHKA) for the collection permit, the village heads, local site owners, PT REKI and Bukit Duabelas National Park for granting access to their properties, and the many colleagues and helpers for support in the field. Financial support was provided by the German Research Foundation (DFG) in the framework of the collaborative German – Indonesian research project CRC990 (EFForTS).
References
Athias-Henriot, C. (1971) La divergence néotaxique des Gamasides (Arachnides). Bull. Sci. Bourgogne, 28: 93-106.
Athias-Henriot, C. (1975) Nouvelles notes sur les Amblyseiini. 2. Le relevé organotaxique de la face dorsale adulte (gamasides protoadéniques, Phytoseiidae). Acarologia, 17: 20-29.
Berlese, A. (1910) Brevi diagnosi di generi e specie nuovi di Acari. Redia, 6: 346-388.
Berlese, A. (1916) Centuria prima di Acari nuovi. Redia, 12: 19-67.
Berlese, A. (1918) Centuria quarta di Acari nuovi. Redia, 13: 115-192.
Britto, E.P.J., Lindquist, E.E., Moraes, G.J. (2011) Redescription of Lasioseius floridensis Berlese, 1916 (Acari: Mesostigmata: Blattisociidae), with notes on closely related species. Zootaxa, 15: 1-15. doi:10.11646/zootaxa.2905.1.1 ![]()
Christian, A., Karg, W. (2006) The predatory mite genus Lasioseius Berlese , 1916 (Acari, Gamasina). Abhandlungen und Berichte des Natrkundemuseums Görlitz, 77(2): 99-250.
Decker, P., Christian, A., Xylander, W.E.R. (2018) VIRMISCO – the virtual microscope slide collection. In: P. Stoev and G. D. Edgecombe (Eds), Proceedings of the 17th International Congress of Myriapodology. Zookeys, Krabi, Thailand, pp. 271-282. doi:10.3897/zookeys.741.22284 ![]()
Drescher, J., Rembold, K., Allen, K., Beckscha, P., Buchori, D., Clough, Y., Faust, H., Fauzi, A.M., Gunawan, D., Hertel, D., Irawan, B., Jaya, I.N.S., Klarner, B., Kleinn, C., Knohl, A., Kotowska, M.M., Krashevska, V., Krishna, V., Leuschner, C., Lorenz, W., Meijide, A., Melati, D., Steinebach, S., Tjoa, A., Tscharntke, T., Wick, B., Wiegand, K., Kreft, H., Scheu, S. (2016) Ecological and socio-economic functions across tropical land use systems after rainforest conversion. Phylosophicla Trans. B, 371: 20150275. doi:10.1098/rstb.2015.0275 ![]()
Ehara, S. (1964) Some mites of the families Phytoseiidae and Blattisocidae from Japan (Acarina: Mesostigmata). J. Fac. Sci. Hokkaido Univ. Ser. VI, Zool., 15: 378-394.
Evans, G.O. (1963) Observations on the chaetotaxy of the legs in the free-living Gamasina (Acari: Mesostigmata). Bull. Br. Museum (Natural Hist., 10: 275-303. doi:10.5962/bhl.part.20528 ![]()
Evans, G.O., Hyatt, K.H. (1960) A revision of the Platyseiinae (Mesostigmata: Aceosejidae) based on material in the collections of the British Museum (Natural History). Bull. Br. Museum (Natural Hist. Zool., 6(2): 27-101. doi:10.5962/bhl.part.26844 ![]()
Evans, G.O., Sheals, J.G. (1959) Three new mesostigmatic mites associated with millipedes in Indonesia. Entomol. Berichte, 19: 107-111.
Evans, G.O., Till, W.M. (1965) Studies on the British Dermanyssidae (Acari: Mesostigmata). Part 1. External morphology. Bull. Br. Museum (Natural Hist. Zool., 13: 247-294. doi:10.5962/bhl.part.16752 ![]()
Kempson, D., Lloyd, M., Ghelardi, R. (1963) A new extractor for woodland litter. Pedobiologia (Jena)., 3: 1-21.
Lindquist, E.E. (1994) Some observations on the chaetotaxy of the caudal body region of gamasine mites (Acari, Mesostigmata), with a modified notation for some ventrolateral body setae. Acarologia, 35(4): 323-326.
Lindquist, E.E., Evans, G.O. (1965) Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Mem. Entomol. Soc. Canada, 97(S47): 5-66. doi:10.4039/entm9747fv ![]()
Lindquist, E.E. (1964) A revision of mites of the subfamily Blattisocinae (Acarina: Blattisocidae) in America north of Mexico. University of California, Berkeley - University Microfilms, Inc., Ann Arbor, Michigan.
Lindquist, E.E., Moraza, M.L. (2010) Revised diagnosis of the family Blattisociidae (Acari: Mesostigmata: Phytoseioidea), with a key to its genera and description of a new fungus-inhabiting genus from Costa Rica. Zootaxa, 21: 1-21. doi:10.11646/zootaxa.2479.1.1 ![]()
Mineiro, J.L. de C., Lindquist, E.E., De Moraes, G.J. (2009) Edaphic ascid mites (Acari: Mesostigmata: Ascidae) from the state of São Paulo, Brazil, with description of five new species. Zootaxa, 2024: 1-32. doi:10.11646/zootaxa.2024.1.1 ![]()
Moraes, G.J., Britto, E., Mineiro, J., Halliday, B. (2016) Catalogue of the mite families Ascidae Voigts & Oudemans, Blattisociidae Garman and Melicharidae Hirschmann (Acari: Mesostigmata). Zootaxa, 4112(1): 1-299. doi:10.11646/zootaxa.4112.1.1 ![]()
Moraes, G.J., Venancio, R., Dos Santos, V.L.V, Paschoal, A.D. (2015) Potential of Ascidae, Blattisociidae and Melicharidae (Acari: Mesostigmata) as biological control agents of pest organisms BT - Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms. In: D. Carrillo, G. J. de Moraes, and J. E. Peña (Eds), Springer International Publishing, Cham, pp. 33-75. doi:10.1007/978-3-319-15042-0_2 ![]()
Moraza, M.L., Lindquist, E.E. (2018) A new species-group with new species of the genus Lasioseius (Acari: Mesostigmata: Blattisociidae) associated with neotropical hispine beetles in furled leaves of Heliconia. Acarologia, 58(1): 62-98. doi:10.24349/acarologia/20184227 ![]()
Moraza, M.L., Linquist, E.E. (2011) A new genus of fungus-inhabiting blattisociid mites (Acari: Mesostigmata: Phytoseioidea) from Middle America, with a key to genera and subgenera of the subfamily Blattisociinae. Zootaxa, 2758: 1-25. doi:10.11646/zootaxa.2758.1.1 ![]()
Vitzthum, G.H. (1925) Fauna sumatrensis. Acarinae. Suppl. Entomol., No. 11: 1-79.
Walter, D.E., Lindquist, E.E. (1997) Australian species of Lasioseius (Acari: Mesostigmata: Ascidae): the porulosus group and other species from rainforest canopies. Invertebr. Taxon., 11: 522-547. doi:10.1071/IT96003 ![]()
Womersley, H. (1956) On some new Acarina-Mesostigmata from Australia, New Zealand and New Guinea. J. Linn. Soc. London, Zool., 42: 505-599. doi:10.1111/j.1096-3642.1956.tb02218.x ![]()



2020-01-30
Date accepted:
2020-04-22
Date published:
2020-04-27
Edited by:
Faraji, Farid

This work is licensed under a Creative Commons Attribution 4.0 International License
2020 Quintero-Gutiérrez, Edwin Javier; Sandmann, Dorothee; Cómbita-Heredia, Orlando; Klarner, Bernhard; Widyastuti, Rahayu and Scheu, Stefan
 Download article Download low definition
Download article Download low definitionDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)