



The first report on oribatid mites in tundra belts of the Lovozersky Mountains on the Kola Peninsula, Russia
Leonov, Vladislav D.1 and Rakhleeva, Anna A.2
1✉ A.N. Severtsov Institute of Ecology and Evolution RAS, 119071, Russia.
2Soil Science Faculty, Lomonosov Moscow State University, 119991, Russia.
2020 - Volume: 60 Issue: 2 pages: 301-316
https://doi.org/10.24349/acarologia/20204369Original research
Keywords
Abstract
Introduction
The oribatid mite communities in mountain tundra habitats of the Kola Peninsula remain poorly studied. They have been studied only in the Khibiny Mountains, where 50 species, 34 genera, and 23 families of oribatid mites have been found (Zenkova and Melekhina 2014; Leonov and Rakhleeva 2015; Leonov et al. 2015; Leonov 2017). In two other large mountain massifs of the Kola Peninsula (the Chunatundra and the Lovozersky Mountains), oribatid mites have not been studied to date. The oribatid mite fauna of the plain tundra on the Kola Peninsula is better investigated, with 140 species known, belonging to 73 genera and 39 families (Krivolutsky 1966; Liskovaya 2011; Leonov and Rakhleeva 2015; Leonov 2017).
In the Scandinavian Peninsula, a total of 145 species, 69 genera, and 42 families of oribatid mites have been found only in the alpine habitats (Heggen 2010). Meanwhile, 151 species of oribatid mites are known to inhabit the western North American Low Arctic (Behan-Pelletier 1999a). Based on the aforementioned data, a considerable species richness is expected to be found in the mountain tundra habitats of the Kola Peninsula.
An important aspect of the tundra community research is the study of oribatid mites in different types of tundra, which include lichen tundra, dwarf shrub tundra and wetted tundra conditions. Due to differences in the vegetation of these tundra types, differences in the environmental conditions for oribatid mites exist. Thus, we can expect a major difference in the fauna and abundance of oribatid mites in these habitats.
The aims of this research are to obtain initial data on the fauna and abundance of oribatid mites of the Lovozersky Mountains and to compare them between the explored tundra habitats and with the literature data.
Materials and methods
Study area
The Lovozersky Mountains are located in the central part of the Kola Peninsula, east of the Khibiny Mountains (Figure 1). The average annual air temperature there is approximately -1\,°C. The region belongs to an area with a predominance of precipitation over evaporation. In areas with complex terrain, precipitation amounts to 600–800 mm. Precipitation exceeds 1 000 mm at the peaks of the largest mountain ranges. The climate type is subarctic marine (Agarkova et al. 2008).
In the mountains of the Kola Peninsula the following vegetation zones are present: the spruce-pine-birch belt occupies areas up to 250–480 m a.s.l.; above this, the belt of birch crooked forest spreads over several tens of meters; the belt of dwarf shrubs extends up to 600–700 m a.s.l.; the alpine lichen tundra belt occurs up to 850–900 m a.s.l.; and the belt of sparse vegetation covers the tops of the mountains and plateaus (Stanyukovich 1973).
Study sites
The research was conducted in the Seydyavvr State Nature Reserve (Figure 1). Five mountain tundra plots on a south-facing slope were explored in the northern part of the Lovozersky Mountains. The plots formed an altitude transect from the plateau between the Kuamdespahk and Kuivchorr Mountains to Lake Seydozero along the Suolui River. The following coordinates and altitudes of the plots were obtained using a GPS/GLONASS Garmin eTrex 30 receiver:

• 1) Grass-moss-lichen tundra plot: altitude of 740 m a.s.l. (plot GMLT-740; 67°51.118' N, 34°53.623' E). Vegetation cover is closed. The main plant and lichen species are Salix sp., Dryas octopetala, Empetrum hermaphroditum, Phyllodoce caerulea, Cetrariella delisei, Cladonia stellaris, Sphaerophorus globosus, Pleurozium schreberi, Juncus trifidus, and Carex sp. Grasses (single sedges and bumps of Juncus trifidus) grow mainly among lichen thalluses. Cetrariella delisei dominates among lichens. Cladonia stellaris is numerous. Pleurozium schreberi is a dominant species among mosses. The proportion of mosses in the vegetation is small compared to lichens. The Marchantiophytas occupy areas with no other vegetation. The dwarf shrub layer is not developed: shrubs take the form of trailing among the lichen thalluses. Some shrubs exhibit cushion growth forms. The soil type is Leptosol (hereafter, soil names are given using the World Reference Base for Soil Resources, WRB, IUSS Working Group WRB 2015).
• 2) Lichen tundra plot: altitude of 740 m a.s.l. (plot LT-740; 67°51.118' N, 34°53.623' E). Vegetation cover is closed. The main plant and lichen species are Salix sp., Betula nana, Vaccinium vitis-idaea, Flavocetraria nivalis, Cladonia stellaris, Sphaerophorus globosus, Pleurozium schreberi, and Carex sp. Flavocetraria nivalis forms a dense, closed lichen cover with the inclusion of Pleurozium schreberi. Salix sp. and Betula nana are not exceeded in height by the lichen thalluses and do not develop a shrub cover. Sparse grasses are represented by sedge species. The soil type is Leptosol.
• 3) Grass-lichen tundra plot: altitude of 721 m a.s.l. (plot GLT-721; 67°51.020' N, 34°53.547' E). This plot is vegetated by Carex spp., Salix sp., Salix cf. polaris, Phyllodoce caerulea, Vaccinium vitis-idaea, Cladonia stellaris, Cetrariella delisei. Vegetation cover is a mosaic represented by evenly distributed grasses (cereals, sedges), mosses, and lichens. Dwarf shrubs do not exceed the height of the lichen. The soil type is Entic Podzol.
• 4) Wetted tundra plot: altitude of 702 m a.s.l. (plot WT-702; 67°50.950' N, 34°53.595' E). This is a heavily wetted site of the mountain tundra which is fed by underground watercourses discharged to the surface and continuing their way open down the slope (the spring of the Suolui River). The vegetation cover is a dense, closed cover of Bryophyta and Marchantiophyta (without Sphagnum mosses) with some dicotyledonous and monocotyledonous plants developed. It is dominated by Dicranum sp., Sarmentypnum sarmentosum, Gymnomitrion concinnatum, Carex sp., Bartsia alpina, and Ranunculus acri. The shrub layer is represented by Salix sp.
• 5) Dwarf shrub tundra plot: altitude of 428 m a.s.l. (plot DST-428; 67°50.300' N, 34°53.913' E). Vegetation cover is closed. The main plant and lichen species are Betula nana, Salix sp., Juniperus communis, Vaccinium myrtillus, V. uliginosum, Cetraria islandica, and Pleurozium schreberi. Closed shrub cover is not developed. Betula nana and Salix sp. are represented by detached plants that are much higher than the dwarf shrubs. Vaccinium myrtillus and V. uliginosum with rare horsetails, small grasses and herbs developed a distinct and closed vegetation layer. The soil cover is represented by mosses and lichens with a dominance of Pleurozium schreberi and Cetraria islandica. The soil is Entic Podzol.
Sampling and sample processing
In each studied habitat 10 samples were taken with a corer (25 cm2) up to 10 cm depth (included topsoil, litter and soil-cover vegetation — mosses, lichen and dwarf shrub parts) between Jul. 30, 2014 and Aug. 1, 2014. The samples were placed in multilayer paper bags and transported to Moscow (moving time — three days) to carry out further processing. Microarthropods were extracted using a Tullgren funnel in the laboratory without additional heating for 10 days (Potapov and Kuznetsova 2011).
Mite identification
Adult oribatid mites were identified to the species level using the identification keys (Gilyarov 1975; Weigmann 2006; Bayartogtokh 2010; Niedbała 2011). We used additional literature on the systematics of oribatid mites, descriptions and redescriptions of species, keys to individual genera and families of mites, regional keys, and keys for individual groups (Shaldybina 1972; Norton 1977; Behan-Pelletier 1985; Behan-Pelletier 1986; Woas 1986; Pavlichenko 1994; Chinone 2003; Miko and Mourek 2008; Miko 2010; Miko et al. 2011; Mourek et al. 2011; Beck et al. 2014; Niedbała 2014). We checked juveniles to identify fauna more completely, but we did not use information about juveniles in further analyzes because of impossibility to identify most of them to species level. The valid species status and synonymy are based on the nomenclature of Weigmann (2006).
Data analysis
Mean values, standard deviations, and standard errors were calculated using Microsoft Office Excel 2010. The oribatid communities in the explored plots were compared using multidimensional scaling and discriminant functional analysis according to Tiunov and Scheu (2000, 2005). Based on the presence of species in individual samples and relative abundance in them, the similarity matrices between samples were calculated using the Jaccard and Bray-Curtis similarity measures (Bray and Curtis 1957; Krebs 1999; Magurran 2003). The similarity matrices were analyzed using multidimensional scaling. The minimum number of meaningful dimensions was evaluated by comparing actual stress values with the theoretical exponential function of stress. A 3D solution for both Jaccard and Bray-Curtis matrices showed the best results. The coordinates of the samples in 3D space were used for discriminant function analysis. Two significant discriminant functions (canonical Roots) were obtained. To interpret the extracted canonical Roots in terms of both occurrence in samples and abundance of oribatid species, Spearman rank correlations were calculated between the canonical scores of canonical Roots and both presence/absence and relative abundance of oribatid species in individual samples. Calculations were made in PAST 3.26b (Hammer et al. 2001) and Statistica 6.0 (StatSoft, Tulsa).
The Engelmann classification (Engelmann 1978) was used to describe the classes of oribatid mite dominance: eudominants (40–100%), dominants (12.5–39.9%), subdominants (4–12.4%), recedents (1.3–3.9%) and subrecedents (less than 1.3%) of mean abundance in the investigated habitats.
Results



A total of 5,535 specimens of adult oribatid mites were collected and identified. Overall, 71 species, 38 genera, and 24 families of oribatid mites were found in the Lovozersky Mountains (Table 1). As a result, the known oribatid mite fauna of the Kola Peninsula has increased by eight species (Liochthonius perfusorius, L. perelegans, Sellnickochthonius immaculatus, Phthiracarus cf. membranifer, Trimalaconothrus angulatus, Suctobelbella cf. prominens, Mycobates sarekensis, Oribatula amblyptera) compared to previous studies (Liskovaya 2011; Zenkova et al. 2011; Zenkova and Melekhina 2014; Leonov and Rakhleeva 2015; Leonov et al. 2015). One specimen of Camisia cf. invenusta in the tritonymph stage was found in plot GMLT-740, though adults of this species were not found.
Three oribatid families — Brachychthoniidae, Oppiidae, and Suctobelbidae — were most abundant in species. Each of these was represented by 11 species, which comprised 15.3% of the total discovered fauna. Camisiidae (5 species) and Ceratozetidae (4 species) contributed 6.9% and 5.6% to the local fauna, respectively.

The explored lichen and dwarf shrub tundra differed considerably in the composition and structure of oribatid mite community. Species of the family Brachychthoniidae constituted the majority of the lichen tundra fauna (8 species, 26.7% of the fauna). The family Suctobelbidae — second in the number of species in lichen tundra — had four species (13.3% of the fauna). All the remaining families included only one or two species. In the dwarf shrub tundra, the family Suctobelbidae had the largest representation (11 species, 19.6% of the fauna). The family Oppiidae exhibited the second-highest number of species (9 species, 16.1% of the fauna), while the family Brachychthoniidae included seven species (12.5% of the fauna).
The wetted tundra plot was characterized by ordinary fauna that is similar in composition to the fauna of the lichen tundra plots. The Brachychthoniidae remained the most abundant in species (3 species, 15% of the fauna), while the Oppiidae that is typically well represented in tundra communities included only two species (6.7% of the fauna) — Oppiella acuminata and O. nova.
The abundance of oribatid mites increased from lichen to dwarf shrub tundra. The lowest oribatid abundance was observed in the wetted tundra plot (Table 1).
Substantial differences were observed in the dominance structure of oribatid mite communities across the studied habitats (Table 2). Lichen tundra was characterized by a small number of dominant species with high levels of dominance in the explored plots. No dominant species were discovered in the dwarf shrub tundra, but 10 subdominant species were noted in this habitat. A specific complex of dominant species was observed in the wetted tundra plot. The strong dominance of T. angulatus and P. peltifer displays a specific type of oribatid dominance structure that is typical for wetlands. Notably, P. peltifer was not observed in the dominant complexes of the other explored tundra habitats and reached high values of dominance only in wetted tundra.
There was a significant difference in the structure of oribatid faunas of the explored habitats (Figure 2). Changes in environmental conditions for oribatid mites from lichen tundra to dwarf shrub tundra (canonical Root 1) led to a statistically significant decrease in the occurrence of M. sarekensis, O. amblyptera, T. velatus, C. labyrinthicus in the studied plots. The occurrence of many species, especially of the families Oppiidae and Suctobelbidae, increased. Species of these families had the highest Spearman R correlation values with canonical Root 1 (Table 3).
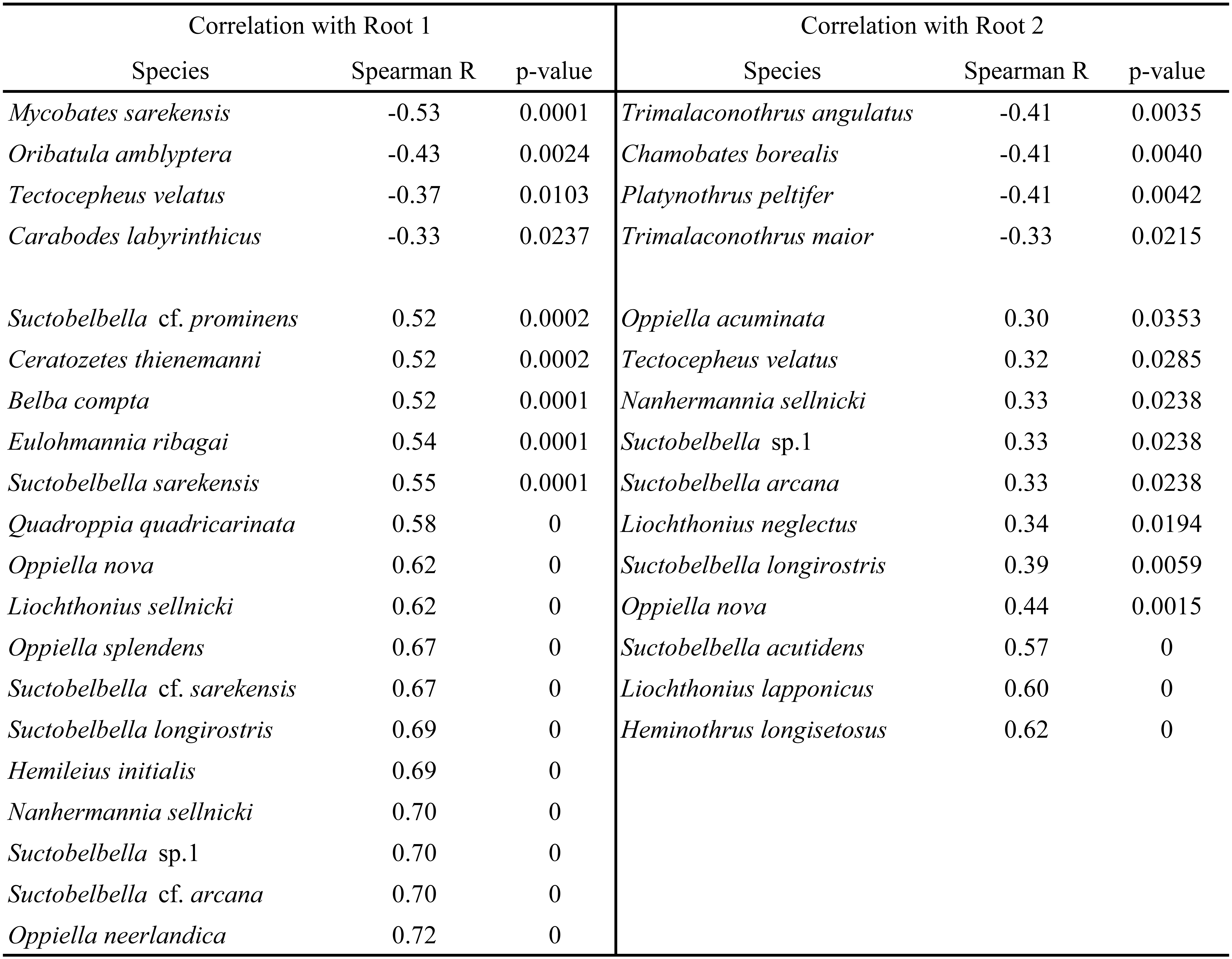

The occurrence of typical wetland oribatid species — T. angulatus, T. maior and P. peltifer (Behan-Pelletier and Eamer 2007) — decreased significantly from the wetted tundra plot to automorphic tundra habitats (canonical Root 2). The occurrence of species belonging to the families Suctobelbidae, Oppiidae and Brachychthoniidae significantly increased. The occurrence of H. longisetosus had the highest positive correlation with Root 2 (Table 3).
Results similar to those described above for fauna were obtained using the relative abundance of species in the studied biotopes (Figure 3). From lichen tundra to dwarf shrub tundra (canonical Root 1) the proportion of species T. velatus, O. amblyptera, C. labyrinthicus, M. sarekensis decreased significantly (Table 4). There was a statistical increase of relative abundance of many species, especially those of the family Suctobelbidae, with the highest values of Spearman R (Table 4). These results are similar to the results obtained exclusively through the analysis of the fauna in the explored habitats (Table 3). But in the case of fauna, just one species, O. neerlandica, had the highest positive correlation with dwarf shrub tundra condition. In the case of relative abundance, O. neerlandica, N. sellnicki, Suctobelbella sp.1 and Suctobelbella arcana demonstrated the highest positive correlation of relative abundance with changes in conditions from lichen tundra to dwarf shrub tundra. The relative abundance of typical wetland oribatid species decreased from the wetted tundra plot to automorphic tundra habitats (Table 4).


Discussion
Based on the analyses carried out above, we can distinguish species complexes specific for different types of tundra. T. velatus, O. amblyptera, C. labyrinthicus, M. sarekensis are more typical for the lichen tundra in this research. Species of the Mycobates and Oribatula are known from previous studies as characteristic for saxicolous mosses above the upper forest line. Species of the Mycobates and Carabodes genera are closely connected with lichens (Materna 2000). M. sarekensis was most common in both higher alpine zones and glacier-forelands in alpine Fennoscandia, T. velatus was related to glacier-forelands (Heggen 2010). T. velatus is a species which thrives in the most severe or disturbed conditions (Zyromska-Rudzka 1977; Kevan et al. 1995; Skubała 1997). Tectocepheus velatus was found as a dominant species (Tikhonov 2003; Sidorchuk 2009) or most occurring species (Thomas and MacLean Jr 1988) in tundra habitats.
The species of Suctobelbidae and Oppiidae were most common and abundant in dwarf shrub tundra of the Lovozersky Mountains and appeared in the complex of dominant species only in this habitat (Table 2). In the Central Alps, species of these families were most common in a mountain forest habitat than above tree-line (Fischer and Schatz 2013). Oppiella splendens, O. neerlandica, O. nova and Suctobelbella subtrigona were limited in altitudinal distribution by low-alpine and sub-alpine belts in the alpine Fennoscandia (Heggen 2010). The Suctobelbidae and Oppiidae species were characteristic for saxicolous mosses below the upper forest-line in the Krokonoše Mountains, Czech Republic (Materna 2000).
According to the law of relative constancy (Walter 1970), wetted tundra should be the most severe habitat in the tundra conditions. It is unexpected to see such strong correlation of relative abundance of Ch. borealis with wet environment (Table 4). Probably, this is one of few species of oribatid mites that are tolerant to wet conditions in the tundra. The relative abundance of species Suctobelbidae, Oppiidae and Brachychthoniidae increased significantly from the wetted tundra to automorphic tundra habitats. H. longisetosus demonstrates the avoidance of wetting under the tundra conditions (Table 3, 4). In alpine Fennoscandia, H. longisetosus was limited by altitude in its distribution (Heggen 2010).
The discovered oribatid mite fauna of the Lovozersky Mountains is typical for the oribatid mite fauna of the European North of Russia if we consider this region as a complex of boreal and tundra habitats (Melekhina 2011). There were some similarities and differences between the oribatid fauna in the explored tundra plots and other tundra and polar desert regions (Table 5).
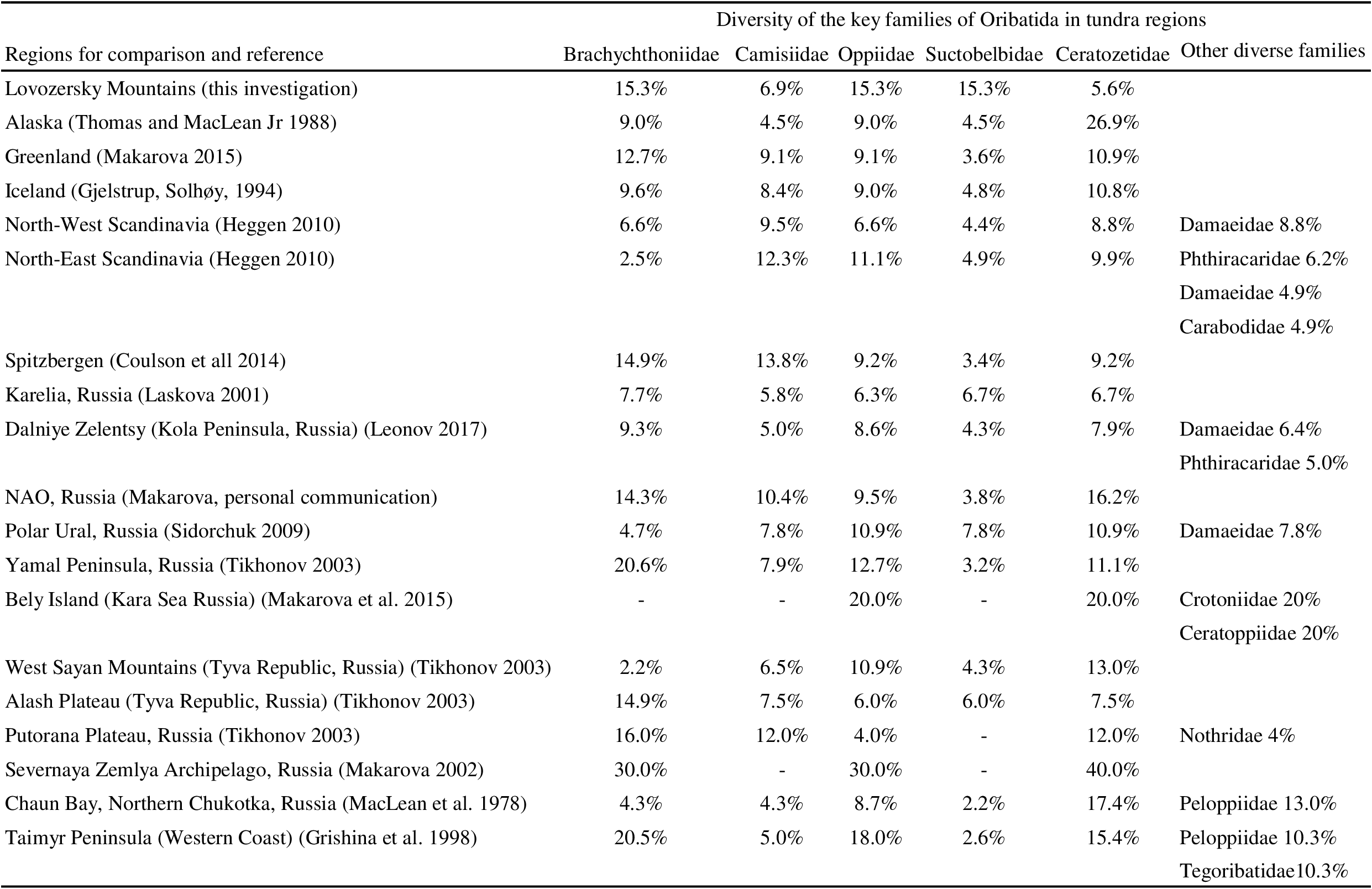
The families Brachychthoniidae, Oppiidae, and Ceratozetidae are usually the most diverse groups in the known tundra faunas (Table 5). It is known that the Ceratozetidae species, especially large ones, predominate in fauna or abundance in the local faunas of oribatid mites in tundra and polar deserts (Hammer 1954; Ananieva et al. 1979; Kalyakin et al. 1998; Behan-Pelletier 1999b; Makarova 2002; Krivolutsky et al. 2003; Ryabinin 2009). However, no species of this family were found in any studied lichen tundra plots, though 15 species of the family Ceratozetidae were noted in the plain lichen tundra of the Kola Peninsula and six species in the lichen tundra of Mount Vudyavrchorr, three of which (Diapterobates humeralis, Fuscozetes sp. and T. cf. trimaculatus) have a large body size (Leonov et al. 2015). Low diversity of Ceratozetidae is not typical for tundra (Table 5).
The discovered high diversity of the family Suctobelbidae in the fauna is not common for the previously explored tundra. In the typical tundra of the Chaunskaya Bay (Chukotka), the family Suctobelbidae was represented by only one species, Suctobelbella sp. (MacLean et al. 1978). Only three species of the family Suctobelbidae (S. acutidens, S. sarekensis, S. setosoclavata) and two species of the family Oppiidae (O. nova, O. translamellata) have been found at the latitudinal transect in Northern Alaska (21 habitats were investigated) (Thomas and MacLean Jr 1988). Despite the high diversity of Oribatida in Greenland Island, only 4 species of the family Suctobelbidae have been found (Makarova 2015). A high diversity of the family Suctobelbidae was found in boreal forests of Karelia (Laskova 2001) and in the mountain tundra of the Polar Urals (Sidorchuk 2009). The increasing role of the family Suctobelbidae is shown not only when comparing the total explored fauna of the Lovozersky Mountains with the faunas of other large regions, but also in the explored dwarf shrub tundra versus the lichen tundra (Tables 1, 2). This means that the high occurrence of the family Suctobelbidae and its high proportion in abundance may be an indicator of milder environmental conditions in arctic/subarctic habitats. This may be typical for a local mountain tundra which is surrounded by the underlying belts of the mountain forests being a source of the increasingly diverse family Suctobelbidae. But the last hypothesis seems to be confirmed only in the Polar Urals (Sidorchuk 2009).
Species found in wetted tundra enriched the species diversity of oribatid mites in the explored region. Liochthonius perfusorius, T. angulatus, T. maior, Oppiella sp.3, E. edwardsi species were added to the fauna of automorphic tundra habitats. However, this did not exclusively occur due to wetland species. Only T. angulatus and T. maior can be strictly considered as wetland species (Behan-Pelletier and Eamer 2007).
There was a similarity in oribatid abundance between the lichen tundra in the Lovozersky Mountains and those known from the literature for mountain and plain lichen tundra (Table 6). The densities of oribatid mites in the studied lichen tundra were similar to previously recorded values for the Khibiny Mountains (Vudyavrchorr Mountain). However, oribatid densities were considerably lower than noted for the plain lichen tundra of the Kola Peninsula (Leonov and Rakhleeva 2015). The recorded abundance in the dwarf shrub tundra of the Lovozersky Mountains does not look typical for most tundra communities, although similar high abundance was noted in previous studies of the mountain tundra of the Pai-Khoi Ridge, Putorana Plateau, and plain tundra of Alaska (Thomas and MacLean Jr 1988; Tikhonov 2003; Melekhina and Zinovyeva 2012). Oribatid abundance in the dwarf shrub tundra was close to that of the plain dwarf shrub tundra of the Kola Peninsula (Leonov and Rakhleeva 2015).

The abundance of oribatid mites in the dwarf shrub tundra was similar to that of forest communities (Table 6). Dwarf shrubs and cushion plants significantly modify the conditions under them (Seniczak and Plichta 1978; Seniczak et al. 2014) and provide to the oribatid mites with thicker litter that is more structurally diverse and has a richer feed base, which may mitigate the effect of severe environmental conditions (Minor et al. 2016a). The observed patterns of fauna (low diversity of the family Ceratozetidae and high diversity of the family Suctobelbidae) and high abundance in dwarf shrub tundra may be explained by the ambivalent nature of the oribatid community in the explored region, which combines boreal and arctic features. The fauna of the explored region is likely to be boreal in its genesis (reduced boreal due to the severity of conditions in the mountain tundra). Moreover, relative mildness of the Kola Peninsula climate due to oceanicity may lead to relative species richness (in comparison with zonal tundra).
The oribatid mite abundance in wetted tundra was considerably lower than in other plots of the Lovozersky Mountains (Table 1). It corresponds to that of oligotrophic bog habitats known from the literature (Weigmann, 1991; Zaitsev 2013; Minor et al. 2016b).
Despite the existence of some knowledge gaps regarding oribatid mite communities in the subarctic mountains and tundra regions, the fragmentation of the existing research, and the heterogeneity of data in the literature, some trends in the characteristic features of oribatid communities in the plain and mountain tundra have emerged. Oribatid communities in the tundra habitats of the Lovozersky Mountains are similar to other tundra regions in the high diversity of the families Brachychthoniidae and Oppiidae in the total fauna and abundances of oribatid mites in the explored lichen tundra are typical for other tundra regions. A high diversity of the family Suctobelbidae in the fauna, a small number of large thick-shelled oribatid mite species of the family Ceratozetidae, their low abundance, and the high abundance of oribatid mites in the dwarf shrub tundra are the features which distinguish the tundra habitats in our investigation from typical tundra environments elsewhere.
Acknowledgments
The authors are very grateful to O.L. Makarova and A.B. Babenko (A.N. Severtsov Institute of Ecology and Evolution RAS) for consultations and discussions regarding the results. The authors would like to thank E.A. Sidorchuk (PIN RAS) for her help and advice in the identification of oribatid mites. The authors thank A.V. Tiunov, Corresponding Member of the RAS (A.N. Severtsov Institute of Ecology and Evolution RAS), for advice in part of statistical processing of the data.
The authors thank the following participants of the expedition during which the material was collected: M.S. Bizin and K.O. Prokopyeva, graduates of the Faculty of Soil Science of the Lomonosov Moscow State University, as well as the staff of the Seydyavvr State Nature Reserve of Regional Significance for their assistance in conducting the expedition.
The authors are very grateful to the reviewers whose advice and remarks helped to improve the manuscript.
This research was supported by the Russian Foundation for Basic Research (Project No. 14-04-31754).
References
Agarkova T.V. et al. 2008. Kola Encyclopedia. Apatity: KSC RAS. pp. 600.
Ananieva S.I., Krivolutsky D.A., Chernov Yu.I. 1979. Oribatid mites (Oribatei) in the subzone of arctic tundra in the northeast of Taimyr. In: Aleksandrova V.D., Matveeva N.V. (Eds). Arctic tundra and polar deserts of Taimyr. Leningrad: Nauka. p. 144-147.
Bayartogtokh B. 2010. Oribatid Mites of Mongolia (Acari: Oribatida). Moscow: KMK Scientific Press. pp. 372.
Beck L., Horak F., Woas S. 2014. Zur Taxonomie der Gattung Phthiracarus PERTY, 1841 (Acari, Oribatida) in Südwestdeutschland. Carolinea, 72: 109-132.
Behan-Pelletier V.M. 1985. Ceratozetidae of the western North American Arctic. Can. Entomol., 117(11): 1287-1366. doi:10.4039/Ent1171287-11 ![]()
Behan-Pelletier V.M. 1999a. Oribatid mite fauna of northern ecosystems: a product of evolutionary adaptations or physiological constraints. In: Needham, Mitchell, Horn and Welbourn (Eds). Acarology IX: Proceedings of the International Congress. p. 87-105.
Behan-Pelletier V.M. 1999b. Oribatid mite biodiversity in agroecosystems: role for bioindication. Agric. Ecosyst. Environ., 74(1): 411-423. doi:10.1016/S0167-8809(99)00046-8 ![]()
Behan-Pelletier V.M., Eamer B. 2007. Aquatic Oribatida: adaptations, constraints, distribution and ecology. In: Acarology XI: Proceedings of the International Congress. p. 71-82.
Behan-Pelletier V.M.1986. Ceratozetidae (Acari: Oribatei) of the Western North American Subarctic. Can. Entomol., 118(10): 991-1057. doi:10.4039/Ent118991-10 ![]()
Bray J.R., Curtis J.T. 1957. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr., 27(4): 325-349. doi:10.2307/1942268 ![]()
Byzova Yu.B., Uvarov A.V., Gubina V.G. et al. 1986. Soil invertebrates of the White Sea islands of the Kandalaksha nature reserve. Moscow: Nauka. pp. 311.
Chernov Yu.I., Ananieva S.I., Khayurova E.P. 1971. A complex of soil invertebrates in the spotted tundra of the Western Taimyr. In: Tikhomirov B.A. (Ed). Biogeocenoses and their productivity of the Taimyr tundra. Leningrad: Nauka. p. 198-211.
Chinone S. 2003. Classification of the soil mites of the family Suctobelbidae (Oribatida) of Japan. Edaphologia, 72: 1-110.
Coulson S.J., Convey P., Aakra K. et al. 2014. The terrestrial and freshwater invertebrate biodiversity of the archipelagoes of the Barents Sea; Svalbard, Franz Josef Land and Novaya Zemlya. Soil. Biol. Biochem., 68: 440-470. doi:10.1016/j.soilbio.2013.10.006 ![]()
Engelmann H.-D. 1978. Zur Dominanzklassifizierung von Bodenarthropoden. Pedobiologia, 18: 378-380. doi:10.1016/0011-2275(78)90090-5 ![]()
Fischer B.M., Schatz H. 2013. Biodiversity of oribatid mites (Acari: Oribatida) along an altitudinal gradient in the Central Alps. Zootaxa. 3626(4): 429-454. doi:10.11646/zootaxa.3626.4.2 ![]()
Gilyarov M.S. 1975. The keys for identification of the soil-dwelling mites. Sarcoptiformes. Moscow: Nauka. pp. 491.
Gjelstrup P., Solhøy T. 1994. Zoology of Iceland. Oribatid mites (Acari). Steenstrupia. Zoological Museum University of Copenhagen. Vol. III. 57e: 1-78.
Grishina L.G., Babenko A.B., Chernov Yu.I. 1998. The Oribatid Mites (Sarcoptiformes, Oribatei) of Taimyr Peninsula Western Coast. Vestnik Zoologii, 32(1-2): 116-118.
Hammer M.S.J. 1954. Collemboles and Oribatids from Peary Land (North Greenland). København: Bianco Lunos Bogtrykkeri A-S. pp. 24.
Hammer Ø., Harper D.A.T., Ryan P.D. 2001. Paleontological Statistics Software: Package for Education and Data Analysis // Palaeontol. Electron, 4(1): 1-9.
Heggen M.P. 2010. Oribatid mites of Alpine Fennoscandia. Nor. J. Entomol., 57(1): 38-70.
IUSS Working Group WRB. 2015. World Reference Base for Soil Resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. Rome: FAO. pp. 191.
Kalyakin V.N., Krivolutsky D.A., Zamesova E.Yu. 1998. On the state of knowledge of terrestrial ecosystems of the Novaya Zemlya. Moscow Univ. Geography Bull., 6: 18-22.
Kevan P.G., Forbes B.C., Kevan S.M., Behan-Pelletier V. 1995. Vehicle tracks on High Arctic tundra: Their effects on the soil, vegetation, and soil arthropods. J. Appl. Ecol. 32: 655-667. doi:10.2307/2404660 ![]()
Krebs C.J. 1999. Ecological Methodology. CA: Addison-Wesley Educational Publishers, Inc. pp. 620.
Krivolutsky D.A. 1966. The oribatid mites in the tundra soils. Pedobiologia, 6(3): 277-280.
Krivolutsky D.A., Drozdov N.N., Lebedeva N.V., Kalyakin V.N. 2003. Geography of soil microarthropods on the Arctic islands. Moscow Univ. Geography Bull., 6: 33-40.
Laskova L.M. 2001. Biodiversity of oribatid mites of Karelia. Biogeography of Karelia. Ser. Biology, 2: 125-132.
Leonov V.D. 2017. The oribatid mites (Acari: Oribatida) of the tundra soils on the Kola Peninsula [PhD Thesis]. Moscow: Soil Science Faculty of the Lomonosov Moscow State University. pp. 205.
Leonov V.D., Rakhleeva A.A. 2015. New information on similarity and difference between highland and plain tundra of the Kola Peninsula based on oribatid mite data (Acari: Oribatida). Euroasian Entomol. J., 14(5): 489-499.
Leonov V.D., Rakhleeva A.A., Sidorchuk E.A. 2015. Distribution of Oribatides Mites (Acari: Oribatida) along an Altitudinal Profile of Mount Vud'yavrchorr (the Khibiny Mountains). Eurasian Soil Sci., 48(11): 1257-1267. doi:10.1134/S1064229315110101 ![]()
Liskovaya A.A. 2011. Faunal and ecological diversity of oribatid mites (Acariformes: Oribatei) in the ecosystems of the Kola North [Phd Thesis]. Petrozavodsk: Petrozavodsk State University. pp. 178.
MacLean S.F., Behan V., Fjellberg A. 1978. Soil Acari and Collembola from Chaun Bay, Northern Chukotka. Arct. Alp. Res., 10(3): 559-568. doi:10.2307/1550679 ![]()
Magurran A.E. 2003. Measuring biological diversity. John Wiley & Sons. pp. 264.
Makarova O.L. 2002. Acarocenoses (Acariformes, Parasitiformes) in polar deserts: 1. Mite assemblages of the Severnaya Zemlya Archipelago: structure of fauna and abundance. Entomol. Rev., 82(7): 839-856.
Makarova O.L. 2015. The fauna of free-living mites (Acari) of Greenland. Entomol. Rev., 95(1): 108-125. doi:10.1134/S0013873815010133 ![]()
Makarova O.L., Ermilov S.G., Yurtaev A.A., Mansurov R.I. 2015. The first data on the soil mites (Acari) of the Arctic Belyi Island (Northern Yamal, the Kara sea). Entomol. Rev., 95(6): 805-810. doi:10.1134/S0013873815060147 ![]()
Materna J. 2000. Oribatid communities (Acari: Oribatida) inhabiting saxicolous mosses and lichens in the Krkonoše Mts. (Czech Republic). Pedobiologia, 44(1): 40-62. doi:10.1078/S0031-4056(04)70027-X ![]()
Melekhina E.N. 2011. Taxonomic diversity and arealogy of oribatid mites (Oribatei) of the European North of Russia. Proc. Komi Sci. Cent. Ural Division of the Russ. Acad. Sci., 2(6): 30-37.
Melekhina E.N., Zinovyeva A.N. 2012. First data on oribatid mites (Acari: Oribatida) of the Pay-Khoy Ridge (Yugor Peninsula). Proc. Komi Sci. Cent. Ural Division of the Russ. Acad. Sci., 2(10): 42-50.
Miko L. 2010. Taxonomy of European Damaeidae (Acari: Oribatida) III. Species of the Kunstidamaeus tenuipes (Michael, 1885) group, with a description of Kunstidamaeus fraterculus n. sp. from East Slovakia. Zootaxa, 2327: 51-64. doi:10.11646/zootaxa.2327.1.4 ![]()
Miko L., Ermilov S.G., Smelyansky I.E. 2011. Taxonomy of European Damaeidae (Acari: Oribatida) VI. The oribatid mite genus Parabelbella: Redescription of P. elisabhetae and synonymy of Akrodamaeus. Zootaxa, 3140: 38-48. doi:10.11646/zootaxa.3140.1.3 ![]()
Miko L., Mourek J. 2008. Taxonomy of European Damaeidae (Acari: Oribatida). I. Kunstidamaeus Miko, 2006 with comments on Damaeus sensu lato. Zootaxa, 1820: 1-26. doi:10.11646/zootaxa.1820.1.1 ![]()
Minor M.A., Babenko A.B., Ermilov S.G., Khaustov A.A., Makarova O.L. 2016a. Effects of cushion plants on high-altitude soil microarthropod communities: cushions increase abundance and diversity of mites (Acari), but not springtails (Collembola). Arct. Antarct. Alp. Res., 48(3): 485-500. doi:10.1657/AAAR0015-064 ![]()
Minor M.A., Ermilov S.G., Philippov D.A., Prokin A.A. 2016b. Relative importance of local habitat complexity and regional factors for assemblages of oribatid mites (Acari: Oribatida) in Sphagnum peat bogs. Exp. Appl. Acarol., 70(3): 275-286. doi:10.1007/s10493-016-0075-9 ![]()
Mourek J., Miko L., Skubała P. 2011. Taxonomy of European Damaeidae (Acari: Oribatida) V. Redescription of Epidamaeus bituberculatus (Kulczynski, 1902). Int. J. Acarol., 37(4): 282-292. doi:10.1080/01647954.2010.517565 ![]()
Niedbała W. 2011. Ptyctimous mites (Acari, Oribatida) of the Palaearctic Region. Systematic part. Warszawa: Natura optima dux Foundation. pp. 472.
Niedbała W. 2014. Supplement to the knowledge of ptyctimous mites (Acari, Oribatida) from Palaearctic Region. Zootaxa, 4057(3): 301-339. doi:10.11646/zootaxa.4057.3.1 ![]()
Norton R.A. 1977. A review of F. Grandjean's system of leg chaetotaxy in the Oribatei and its application to the Damaeidae. Symposium on the Oribatei-Eastern Branch Meeting of the Entomological Society of America. Philadelphia, Pa. (USA). Oct 1975.
Pavlichenko P.G. 1994. Key to ceratozotoid mites (Oribatei, Ceratozetoidea) of Ukraine. Kiev: Schmalhausen Institute of Zoology. pp. 143.
Potapov M.B., Kuznetsova N.A. 2011. Methods of research of microarthropods communities. Moscow: KMK Scientific Press. pp. 88.
Ryabinin N.A. 2009. Peculiarities of oribatid mite distribution in soils of the Far East. Vestnik Far East Branch Russ. Acad. Sci., 3(145): 54-60.
Seniczak S., Plichta W. 1978. Structural dependence of moss mite populations (Acari, Oribatei) on patchiness of vegetation in moss lichen tundra at the north coast of Hornsund, West Spitsbergen). Pedobiologia, 18(8): 145-152.
Seniczak S., Seniczak A., Gwiazdowicz D.J., Coulson S.J. 2014. Community structure of oribatid and gamasid mites (Acari) in moss-grass tundra in Svalbard (Spitsbergen, Norway). Arct. Antarct. Alp. Res., 46(3): 591-599. doi:10.1657/1938-4246-46.3.591 ![]()
Shaldybina E.S. 1972. Some features of the ceratozetoid morphology. Uchenye zapiski Gorky State Pedagogical Inst. Ser. Biol. Sci., 130: 35-66.
Sidorchuk E.A. 2009. New data on the fauna of oribatid mites (Acari, Oribatida) from the Polar Urals. Entmol. Rev., 89: 554-563 doi:10.1134/S0013873809050054 ![]()
Skubała P. 1997. The structure of oribatid mite communities (Acari, Oribatida) on mine dumps and reclaimed land. Zool. Beitr., 38: 59-80.
Stanyukovich K.V. 1973. Vegetation of the USSR mountains. Dushanbe: Donish. pp. 416.
Thomas R.H., MacLean Jr S.F. 1988. Community structure in soil Acari along a latitudinal transect of tundra sites in northern Alaska. Pedobiologia, 31: 113-138.
Tikhonov A.V. 2003. The oribatid mites of zonal and mountain tundra [Diploma Thesis]. Moscow: Lomonosov Moscow State University.
Tiunov A.V., Scheu S. 2000. Microfungal communities in soil, litter and casts of Lumbricus terrestris L. (Lumbricidae): a laboratory experiment. Appl. Soil Ecol., 14(1): 17-26. doi:10.1016/S0929-1393(99)00050-5 ![]()
Tiunov A.V., Scheu S. 2005. Arbuscular mycorrhiza and Collembola interact in affecting community composition of saprotrophic microfungi. Oecologia, 142(4): 636-642. doi:10.1007/s00442-004-1758-1 ![]()
Walter H. 1970. Vegetationszonen und Klima. Verlag Eugen Ulmer, Stuttgart. pp. 382.
Weigmann G. 1991. Oribatid communities in transects from bogs to forests in Berlin indicating the biotope qualities. Mod. Acarol. VOL. 1. P. 359-364.
Weigmann G. 2006. Hornmilben (Oribatida): Acari, Actinochaetida. Keltern: Goecke & Evers. pp. 520.
Woas S. 1986. Beitrag zur Revision der Oppioidea sensu Balogh, 1972 (Acari, Oribatei). Karlsruhe: Landessammlungen für Naturkunde. pp. 204.
Zaitsev A.S. 2013. Oribatid mite communities (Acari: Oribatida) in different habitats of the Polistovsky Nature Reserve (Pskov Region, Russia). Estonian J. Ecol., 62(4): 276-286. doi:10.3176/eco.2013.4.04 ![]()
Zenkova I.V., Melekhina E.N. 2014. The Oribatid mites (Acari: Oribatida) of the Khibiny Mountains. In: Evdokimova G.A., Vandysh O.I. (Eds). Environmental problems of the northern regions and ways of their solution. Apatity: Kola Science Center of the Russian Academy of Sciences. p. 135-140.
Zenkova I.V., Zaitsev A.S., Zalish L.V., Liskovaya A.A. 2011. List of oribatid mites (Acariformes: Oribatida) in tundra and northern taiga soils of the Murmansk region. Transactions Karelian Res. Cent. Russ. Acad. Sci., 1: 54-67.
Zyromska-Rudzka H. 1977. Changes in Oribatid mite community after chemical fertilizer application in a meadow. Ecol. Bull. 25: 133-137.



2019-09-16
Date accepted:
2020-04-08
Date published:
2020-04-15
Edited by:
Baumann, Julia

This work is licensed under a Creative Commons Attribution 4.0 International License
2020 Leonov, Vladislav D. and Rakhleeva, Anna A.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)