



Effects of temperature on a Chinese population of Amblyseius andersoni (Acari: Phytoseiidae) fed with Tetranychus urticae
Li, Yu-Jing1 ; Liu, Qing-Yan2 ; Chang, Jing3 ; Jia, Yong-Hong4 and Meng, Rui-Xia5
1College of Hoticulture and Plant Protection, Inner Mongolia Agricultural University, Hohhot, Inner Mongolia 010019, China.
2College of Hoticulture and Plant Protection, Inner Mongolia Agricultural University, Hohhot, Inner Mongolia 010019, China.
3College of Hoticulture and Plant Protection, Inner Mongolia Agricultural University, Hohhot, Inner Mongolia 010019, China.
4College of Hoticulture and Plant Protection, Inner Mongolia Agricultural University, Hohhot, Inner Mongolia 010019, China.
5✉ College of Hoticulture and Plant Protection, Inner Mongolia Agricultural University, Hohhot, Inner Mongolia 010019, China.
2019 - Volume: 59 Issue: 4 pages: 475-483
https://doi.org/10.24349/acarologia/20194344Original research
Keywords
Abstract
Introduction
The two-spotted spider mite, Tetranychus urticae Koch, is one of the most serious agricultural pests in the world. It is a polyphagous species able to feed on more than 1,100 plant species belonging to more than 250 plant families (Migeon and Dorkeld 2010). This phytophagous mite is widely distributed and is responsible for important yield losses in cultivated crops by damaging leaf tissue and sucking cell contents (Grbic et al. 2011; Gerson and Weintraub 2012; Meck et al. 2013). This pest species develops well under conditions of high temperature and low relative humidity, displaying explosive growth (Hussey and Scopes 1985). In China, T. urticae only occurred sporadically before the 1990's, and it spread rapidly later (Gu et al. 1996, Zhang and Shen 2001) and thus becomes a serious problem requiring a large use of pesticides. One explanation for this increase is that the widespread use of pesticide has eliminated natural enemies especially predatory mites (Choi et al. 2004, Prischmann et al. 2005). The challenge is thus the development of biological control strategies (Bustos et al. 2016).
Species of the family Phytoseiidae are the most common plant inhabiting predatory mites; they play an important role in the natural control of phytophagous mites and insects. Several phytoseiid species are commercially produced for the control of pest mites (McMurtry et al. 2013). Biological control of T. urticae with predatory mites, such as Phytoseiulus persimilis Athias-Henriot, has acquired special significance for a long time (Hussey and Scopes 1985, Skirvin et al. 2002). P. persimilis and Neoseiulus californicus (McGregor) are the subject of ongoing research in China, and have shown potential use in biological control of T. urticae (Xu et al. 2015, Wu et al. 2016). Recently, another phytoseiid mite, Amblyseius andersoni (Chant), was found on wild Chinese wolfberry in the city of Bayan Nur in Inner Mongolia in 2017 ; it is a new species recorded in China (Liu et al. 2019). Amblyseius andersoni a generalist phytoseiid species commonly found in apple orchards in several countries (Croft 1994, Duso and Pasini 2003), especially in Europe (Komlovszky and Jenser 1987; Hegyi et al. 2003, Tixier et al. 2014). A survey conducted in Syria, showed A. andersoni was a dominant species on citrus trees and on wild plants within or around orchards (Barbar, 2013). Unlike other phytoseiid mite (e.g. N. californicus and P. persimilis), adult longevity of A. andersoni is relatively long under experimental conditions (Amano and Chant, 1977). This predatory mite was often observed feeding on alternative foods (McMurtry 1992) and reaching high population levels (McMurtry and Rodriquez 1987). Some authors showed that, it can keep the population of spider mites under the economic threshold levels in fruit orchards (Duso 1992, Duso and Pasini 2003). The ability of A. andersoni to control phytophagous mites depends on several factors, such as the climatic conditions in the area (Croft et al. 1993), the morphological characteristics of plant leaves (Duso and Vettorazzo 1999), inter-specific competition (Duso 1989) and the impact of pesticides (Duso et al. 1992, Pozzebon et al. 2002).
Many studies have demonstrated that temperature significantly affects predatory mite performance (e.g. functional response, mating, and oviposition behaviour), and ultimately fitness (Skirvin and Fenlon 2003, Nguyen and Amano 2009, Jafari et al. 2010, 2012, Xia et al. 2012, Wang et al. 2014). In this paper, we studied the effects of temperature on a population of A. andersoni fed with T. urticae. The life table parameters of A. andersoni were also studied providing useful data for biological control applications.
Materials and methods
Mite species and populations
The T. urticae population used in this study was obtained from the leaves of mulberry in Salaqi, Tumut Right Banner, Baotou City, Inner Mongolia Autonomous Region (40º20'N, 110º19'E) and kept on lima bean plants in an environmental chamber at 25 ± 1 °C, 60 ± 5% RH, and L:D = 16:8. The population of A. andersoni was collected on Chinese wolfberry leaves in Urad Front Banner, Bayan Nur City, Inner Mongolia Autonomous Region (40º43'N, 108º39'E), in September 2017. The population was kept in laboratory on excised bean leaves heavily infested with all stages of T. urticae (25 ± 1 °C). The predator and prey individuals were reared for two generations before use in the experiments.
Experimental setup
Experiments were carried out in the laboratory at 25 ± 1 °C, 60 ± 5% RH, and L:D = 16:8. However, since leaflets on which the experiments were conducted were placed on water-saturated cotton we can suggest that humidity reached higher levels. Excised leaves infested with the different stages of T. urticae were placed dorsal side down on absorbent cotton in a large glass petri dish (12 cm in diameter and 1.5 cm in depth). The absorbent cotton was saturated with distilled water to prevent mite escape. Furthermore, the petri dish was placed on a plastic tray (40 x 35 x 2.5 cm) filled with distilled water.
For the study on developmental times, ovipositing females of A. andersoni were kept on detached leaflets with abundant prey. At 16 °C, eggs did not hatch, so it was impossible start the development experiments at this temperature. Therefore, experiments were carried out at 17, 20, 25, 30 and 35 °C. Eggs were removed every 6 h from the mass rearing and placed singly on detached leaflets (4 cm in diameter) which abundant prey for 30 replicates for each temperature considered. To determine the duration of the immature stages (egg, larva, protonymph and deutonymph), inspections were carried out twice a day under a stereomicroscope until the predators reached adulthood. Also, the sex of the progeny was recorded. The leaflet was renewed once a week. Data from females which died accidentally were excluded from the analyses.
Oviposition and life table parameters
After a new generation of females emerged, the durations of preoviposition, oviposition, postoviposition, the daily fecundity and longevity of each female were recorded daily during her life. Daily age-specific survival (lx ) and fecundity (mx ) rates were calculated for each temperature with the method proposed Birch (1948).
A life table was constructed considering survival and fecundity rates for individuals. Parameters at constant temperatures were calculated as reported by Andrewartha and Birch (1954) as follows.
The net reproductive rate: R0 = ∑lxmx
The intrinsic rate of increase: ∑(e-rm∙xlx mx) = 1
The mean generation time: t = lnR0 / rm
The finite rate of increase: λ = erm
The doubling time: DT = ln2 / rm
where rm is the intrinsic rate of increase, x is female age in days, lx is the fraction of females surviving to age x (age-specific survival rate), and mx is the expected number of daughters produced per female alive at age x (age-specific fecundity rate), obtained by multiplying the number of eggs by the age-specific sex ratio, which is defined as the proportion of females in the progeny (Roy et al. 2002).
Statistical analysis
The influence of five temperatures on developmental time of immature stages, durations of preoviposition, oviposition and postoviposition, longevity and fecundity of A. andersoni were analyzed using one-way analysis of variance (ANOVA). When a significant difference was detected, the means were compared using Duncan's multiple ranges test. Hatchability and survival rates were compared with chi-square test using the software SPSS 19.
The life table parameters of A. andersoni were calculated with Excel 2007 software : intrinsic rate of increase (rm ), the finite rate of increase (λ), the net reproductive rate (R0) and the mean generation time (t).
Results
Development and survival
The developmental time of different immature stages of A. andersoni at five constant temperatures are presented in the Table 1. Temperature had a significant influence on eggs (F4, 93 = 369.386; P < 0.001), larval (F4, 93 = 112.489; P < 0.001), protonymphal (F4, 93 = 518.669; P < 0.001), deutonymphal (F4, 93 = 287.256; P < 0.001), and overall immature periods (F4, 93 = 886.114; P < 0.001) for female progeny. Egg duration decreased from 6.75 days at 17 °C to 0.79 days at 35 °C. The larval stage was the shortest for all temperatures and varied between 2.82 days at 17 °C and 0.58 day at 35 °C. The protonymphal and deutonymphal stages had almost similar durations with the deutonymphal stage slightly longer for all temperatures tested. The total immature developmental duration (egg to adult) decreased when temperature increased from 17 °C (24.71 days) to 35 °C (4.54 days). Similarly, temperature had a significant influence on egg (F4, 26 = 138.230; P < 0.001), larval (F4, 26 = 34.919; P < 0.001), protonymphal (F4, 26 = 49.844; P < 0.001), deutonymphal (F4, 26 = 43.167; P < 0.001) and overall immature periods (F4, 26 = 563.898; P < 0.001) for male progeny. Incubation period decreased when temperature increased from 17 °C (6.30 days) to 35 °C (1.33 days). The longest larval stage was 2.30 days at 17 °C, and the shortest 0.50 days at 30 °C and 35 °C. The total immature developmental duration decreased from 21.70 days at 17 °C to 3.92 days at 35 °C.


Hatchability and survival rates of A. andersoni at five constant temperatures are presented in the Table 2. Amblyseius andersoni successfully developed to adulthood at all temperatures tested when fed with T. urticae. Temperature had a significant influence on hatchability (χ2 = 17.07; df = 4; P = 0.007) and survival rate (egg to adult) (χ2 = 10.86; df = 4; P = 0.028). The shortest hatchability and survival rates (egg to adult) were 73.33% and 63.33% at 17 °C, and the highest were 100% and 93.33% at 30 °C. However, temperature had no significant influence on survival rates in larvae (χ2 = 3.75; df = 4; P = 0.440), protonymph (χ2 = 2.54; df = 4; P = 0.637) and deutonymph (χ2 = 0.09; df = 4; P = 0.999).
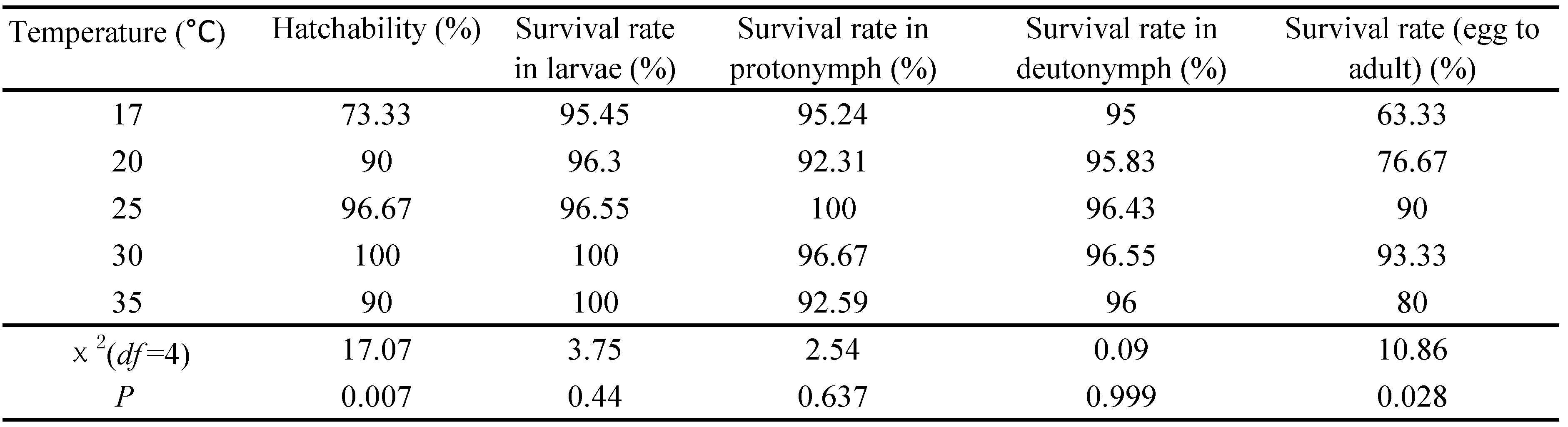
Reproduction and longevity
The reproduction parameters are presented in the Table 3. Significant differences were observed for the preoviposition duration of A. andersoni at the five temperatures tested (F4, 87 = 45.82; P < 0.001) ; it decreased from 27.00 days at 17 °C to 0.97 days at 35 °C, gradually. The oviposition duration decreased from 30.00 days at 20 °C to 16.50 days at 35 °C, and the shortest (9.16 days) was observed at 17 °C (F4, 87 = 19.22; P < 0.001). The total fecundity was the highest at 25 °C (45.68 eggs) and the lowest at 17 °C (9.13 eggs) (F4, 87 = 59.32; P < 0.001). The daily egg production was also affected by temperature and ranged from 0.64 to 1.86 eggs/female/day (F4, 87 = 53.77; P < 0.001). The maximum female-biased sex ratio was 3.64: 1 (female: male) at 25 °C. The female adult longevity (F4, 93 = 31.34; P < 0.001) and male adult longevity (F4, 26 = 6.23; P = 0.002) decreased significantly with temperature increase. The female adult mites lived longer at 17 °C (40.50 days) followed at 20 °C (31.56 days), and the shortest longevity (17.31 days) was observed at 35 °C. Males had almost similar life durations as females. However, at all temperatures tested, the female adult longevity was a little longer.
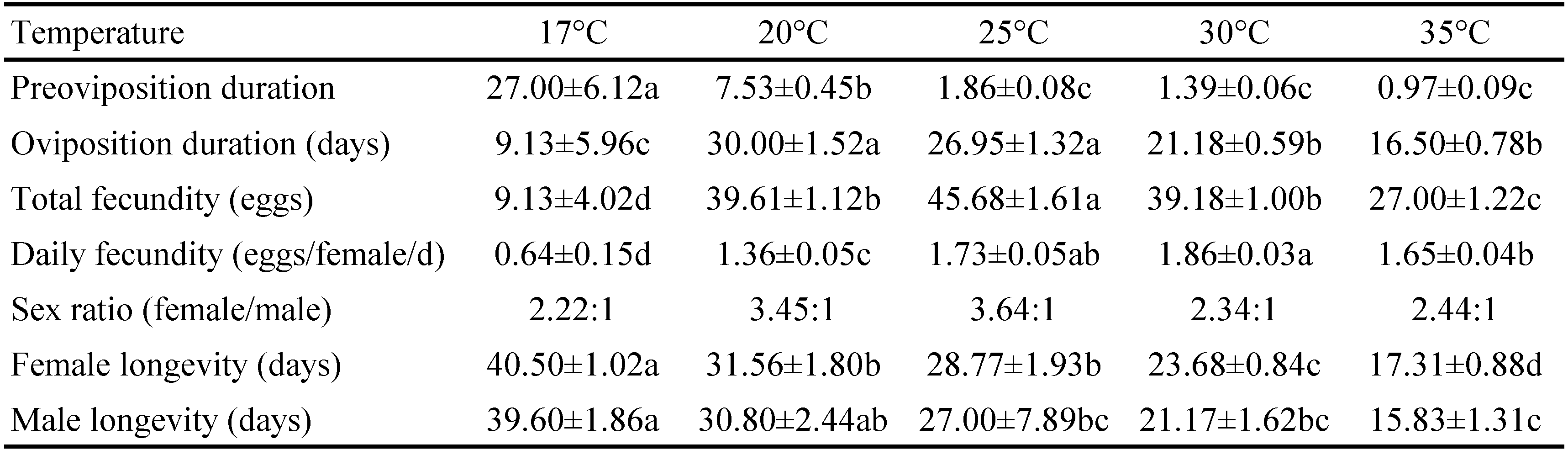
The age-specific survival rate (lx ) and fecundity (mx )
The age-specific survival rate (lx ) and fecundity (mx ) curves of A. andersoni at different temperatures are given in Figure 1. Age-specific survival rate started to drop at an earlier age as the temperature increased from 17 to 35 °C. The first death of an adult female occurred on day 15 at 35 °C, which is earlier than that at the other four temperatures. The curves of age specific fecundity showed that oviposition pattern varied among the temperatures, and the maximum amount of females were recorded on the 53th, 32th, 30th, 15th and 8th days at 17, 20, 25, 30 and 35 °C, respectively; and the females began to oviposit on the 25th, 15th, 9th, 8th, and 6th days at 17, 20, 25, 30 and 35 °C, respectively.
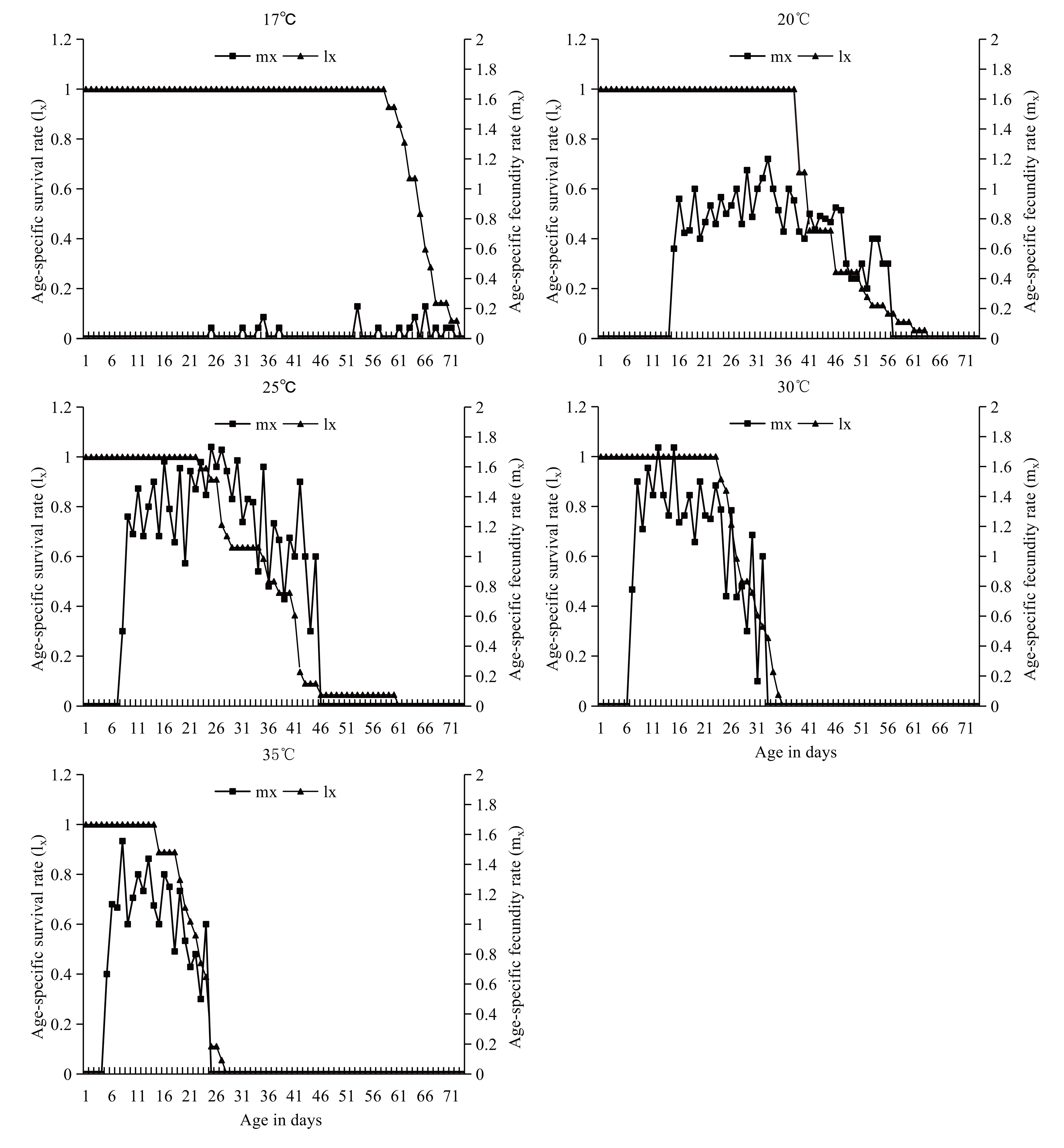
Demographic parameters
The values of life table parameters of A. andersoni at five constant temperatures are shown in the Table 4. Net reproductive rate (R0) was the highest at 25 °C (36.47) and the lowest at 17 °C (1.02). When the temperature increased, the generation time of A. andersoni decreased gradually. The longest mean generation time was reached at 17 °C (48.16), and the shortest at 35 °C (14.68). There was a linear negative correlation between temperature and mean generation period, and the equation was y = -1.6638x + 68.663 (R2 = 0.81, P < 0.05). The intrinsic rate of increase of A. andersoni increased gradually with increasing temperature. The lowest rate was observed at 17 °C (0.0003), and the highest at 35 °C (0.1995). Temperature and the rate of population increase were positively correlated (y = 0.0101x - 0.1247, R2 = 0.80, P < 0.05). When the temperature increased, the doubling time of the population of A. andersoni decreased. The maximum population doubling time was obtained at 17 °C (2197.14), and the shortest at 35 °C (3.47). The lowest finite rate of increase was at 17 °C (1.000), and the highest at 35 °C (1.221). The relationship between temperature and finite rate of increase was linear (y = 0.011x + 0.856, R2 = 0.82, P < 0.05).

Discussion
Our results revealed that temperature significantly affected the developmental time of A. andersoni, and that this predator could successfully develop under a wide range of temperatures from 17 to 35 °C. Total developmental time of A. andersoni recorded by Amano and Chant (1977) fed on T. pacificus at 23 °C (7.43 days) is close to our finding at 25 °C (8.18 days).
The preoviposition periods and female longevity of A. andersoni shortened as the temperature increased, showing a similar trends as reported by Amano and Chant (1977) although the specific values (1.86) obtained in the present study was different from the reported value (2.14). The ovipostion rate of A. andersoni was low at low temperatures, and only a few individuals laid eggs at 17 °C. The reason for this might be that the predator entered reproductive diapauses at low temperatures (Van Houten et al. 1988). Genini et al. (1991) reported that the daily egg production of A. andersoni fed on T. urticae was 1.76 at 20 °C, which was higher than the value (1.36) in the present work. The reason for this may be from different geographic populations of the predator species considered.
The sex ratio of phytoseiids is generally female biased and varies within and between species, but poor prey conditions may reduce the proportion of females (Friese and Gilstrap 1982, Sabelis 1985a,b, Ganjisaffar et al. 2011). In our experiments the sex ratio of A. andersoni was female biased at five constant temperatures, but the maximum sex ratio was found at 25 °C. Sabelis and Nagelkerke (1988) revealed that at low temperatures a pseudo-arrhenotoky process led to production of higher number of sons. Our results suggested that this phenomenon in A. andersoni could occur not only at low temperatures, but also at higher temperatures.
The intrinsic rate of increase (rm ) is a key demographic parameter useful for predicting the population growth potential of an animal under given environmental conditions (Andrewartha and Birch 1954). Our results showed that, like other phytoseiid mites, the rm value of A. andersoni is affected by temperature. At 17 °C, the lower rm value was due to low values of fecundity and survival rate, low production of female offspring in the population, and long life span. At 20 °C, fecundity and survival rate increased, but the rm value was still lower compared to that at the other higher tested temperatures. We concluded that the long life span at 20 °C led to a low value of rm . Genini et al. (1991) found that the rm of A. andersoni fed on T. urticae was 0.151 at 20 °C, which was higher than the value obtained (0.108) in the present work. This difference may be due to differences in geographic populations. At temperatures above 20 °C, it seems that the shorter life span of females is the major reason for increasing rm . Our results showed that the main effect of temperature on rm value of A. andersoni was mostly due to effect on developmental time of the predator. The present study indicated that the maximum rm value of A. andersoni was obtained at 35 °C, which showed this predator seems to be a thermophile species, even though the highest value of survival and fecundity was at 25 °C and 30 °C, respectively.
Finally, although our results indicated that the population of A. andersoni was limited by low temperature, A. andersoni has a high inherent potential for the control of the T. urticae, especially at a temperature range of 30–35 °C. Therefore further experiments on the life table of the local population of T. urticae, under more realistic circumstances, are needed in order to get a better understanding of its efficiency for a successful biological control of T. urticae.
Acknowledgements
The project is supported by the National Key R&D Program of China (2017YFD0201000) and National Natural Science Foundation of China (31560529).
References
Amano H., Chant D.A. 1977. Life history and reproduction of two species of predacious mites, Phytoseiulus persimilis Athias-Henriot and Amblyseius andersoni (Chant) (Acarina: Phytoseiidae). Can. J. Zool., 55: 1987-1983. doi:10.1139/z77-255 ![]()
Andrewartha H.G., Birch L.C. 1954. The distribution and abundance of animals. Quarterly Review of Biology, 121(4): 389-390.
Barbar Z. 2013. Survey of phytoseiid mite species (Acari: Phytoseiidae) in citrus orchards in Lattakia governorate, Syria. Acarologia, 53 (3): 247-261. doi:10.1051/acarologia/20132098 ![]()
Birch L.C. 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol., 17: 15-26. doi:10.2307/1605 ![]()
Bustos A., Rodríguez D., Cure J.R., Cantor F. 2016. A simulation model of the mass rearing of Tetranychus urticae Koch (Acari: Tetranychidae) on beans. Neotropical Entomol., 45(3): 291-299. doi:10.1007/s13744-016-0366-1 ![]()
Choi W.I., Lee S.G., Park H.M., Ahn Y.J. 2004. Toxicity of plant essential oils to Tetranychus urticae (Acari: Tetranychidae) and Phytoseiulus persimilis (Acari: Phytoseiidae). J. Econ. Entomol., 97(2): 553-558. doi:10.1093/jee/97.2.553 ![]()
Croft B.A., Messing R.H., Dunley J.E., Strong W.B. 1993. Effects of humidity on eggs and immatures of Neoseiulus fallacis, Amblyseius andersoni, Metaseiulus occidentalis and Typhlodromus pyri (Phytoseiidae): implications for biological control on apple, caneberry, strawberry and hop. Exp. Appl. Acarol., 17: 451-459. doi:10.1007/BF00120503 ![]()
Duso C. 1989. Role of the predatory mites Amblyseius aberrans (Oud.), Typhlodromus pyri Scheuten and Amblyseius andersoni (Chant) (Acari, Phytoseiidae) in vineyards. I. The effect of single or mixed phytoseiid population releases on spider mite densities (Acari, Tetranychidae). J. Appl. Entomol., 107: 474-492. doi:10.1111/j.1439-0418.1989.tb00283.x ![]()
Duso C. 1992. Biological control of tetranychid mites in peach orchards of northern Italy: Role of Amblyseius finlandicus (Oud.) (Acari: Phytoseiidae). Acta Phytopathologica Et Entomol Hungaric. 27: 211-217.
Duso C., Camporese P. Geest L.P.S.V.D. 1992. Toxicity of a number of pesticides to strains of Typhlodromus pyri and Amblyseius andersoni (Acari: Phytoseiidae). Entomophaga, 37(3): 363-372. doi:10.1007/BF02373110 ![]()
Duso, C., Pasini M. 2003. Distribution of the predatory mite Amblyseius andersoni chant (acari: phytoseiidae) on different apple cultivars. Anz. Schädling., 76(2): 33-40. doi:10.1046/j.1439-0280.2003.03003.x ![]()
Duso C., Vettorazzo E. 1999. Mite population dynamics on different grape varieties with or without phytoseiids released (Acari: Phytoseiidae). Exp. Appl. Acarol., 23: 741-763. doi:10.1023/A:1006297225577 ![]()
Friese D.D., Gilstrap F.E. 1982. Influence of prey availability on reproduction and prey consumption of Phytoseiulus persimilis, Amblyseius californicus and Metaseiulus occidentalis (Acarina: Phytoseiidae). Intern. J. Acarol., 8: 85-89. doi:10.1080/01647958208683283 ![]()
Ganjisaffar F., Fathipour Y., Kamali K. 2011. Temperature-dependent development and life table parameters of Typhlodromus bagdasarjani (phytoseiidae) fed on two-spotted spider mite. Exp. Appl. Acarology, 55 (3): 259-272. doi:10.1007/s10493-011-9467-z ![]()
Genini M., Klay A., Baumgärtner J, Delucchi V., Baillod M. 1991. Comparative studies on the influence of temperature and food on the development of Amblyseius andersoni, Neoseiulus fallacis, Galendromus longipilus and Typhlodromus pyri (acari: phytoseiidae). Entomophaga, 36: 1139-1154. doi:10.1007/BF02374645 ![]()
Gerson U., Weintraub P.G. 2012. Mites (Acari) as a factor in greenhouse management. Ann. Rev. Entomol., 57(1): 229-247. doi:10.1146/annurev-ento-120710-100639 ![]()
Grbic M., T. Van Leeuwen, Clark R.M., Rombauts S., RouzéP., Grbii V., Osborne E.J., Dermauw W., Ngoc P.C.T., Ortego F., Hernández-Crespo P., Diaz I., Martinez M., Navajas M., Sucena É., Magalhães S., Nagy L., Pace R., Djuranovii S., Smagghe G., Iga M., Christiaens O., Veenstra J.A., Ewer J., Villalobos R.M., Hutter J.L., Hudson S.D., Velez M., Yi S.V., Zeng J., Pires-da Silva A., Roch F., Cazaux M., Navarro M., Zhurov V., Acevedo G., Bjelica A., Fawcett J.A., Bonnet E., Martens C., Baele G., Wissler L., Sanchez-Rodriguez A., Tirry L., Blais C., Demeestere K., Henz S.R., Gregory T.R., Mathieu J., Verdon L., Farinelli L., Schmutz J., Lindquist E., Feyereisen R. Van de Peer Y. 2011. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature, 479(7374): 487-492. doi:10.1038/nature10640 ![]()
Gu Y., Zhang Y.C., Zhang Z.F. 1996. Study on the predation of Stethorus punctillum Weise to Tetranychus urticae (Koch). Natural Enemies of Insects, 18(4): 163-166.
Hegyi T., Molnár J., Földes L.Sz. 2003. Ragadozóatkák dominancia-viszonyai Szabolcs-Szatmár-Bereg megye almaültetvényeiben. Kertgazdaság, 35: 5-9.
Hussey N.W., Scopes N. 1985. Biological pest control the glasshouse experience. Ithaca: Cornell University Press, 50-51pp.
Jafari S., Fathipour Y., Faraji F. 2012. The influence of temperature on the functional response and prey consumption of Neoseiulus barkeri (Acari: Phytoseiidae) on Tetranychus urticae (Acari: Tetranychidae). J. Entomol. Soc. Iran, 31(2): 39-52. doi:10.1111/j.1744-7917.2011.01444.x ![]()
Jafari S., Fathipour Y., Faraji F., Bagheri M. 2010. Demographic response to constant temperatures in Neoseiulus barkeri (Phytoseiidae) fed on Tetranychus urticae (Tetranychidae). Syst. Appl. Acarol., 15(2): 83-99. doi:10.11158/saa.15.2.1 ![]()
Komlovszky Sz.I., Jenser G. 1987. The frequent occurrence of the predatory mites Amblyseius finlandicus Oudemans and Phytoseius plumifer Canestrini et Fanzago on fruit trees. Növényvédelem, 23: 193-201.
Liu J. Q., Fang X. D., Liu Q. Y., Meng R. X. 2019. The species of phytoseiid mites on Chinese wolfberry in the city of Bayan Nur in Inner Mongolia. J. Shanxi Agricultural Sci., 47(2): 256-258.
McMurtry J.A. 1992. Dynamics and potential impact of ''generalist" phytoseiids in agroecosystems and possibilities for establishment of exotic species. Exp. Appl. Acarol., 14: 371-382. doi:10.1007/BF01200574 ![]()
McMurtry J.A., De Moraes G.J. Sourasso, N.F. 2013. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol., 18(4), 297-320. doi:10.11158/saa.18.4.1 ![]()
McMurtry J.A., Rodriquez J. 1987. Nutritional ecology of phytoseiid mites. In: Slansky, F., Rodriquez, J. (Eds.), Nutritional Ecology of Insects, Mites and Spiders. John Wiley and Sons, New York, 1016p.
Meck E.D., Kennedy G.G. Walgenbach J.F. 2013. Effect of Tetranychus urticae (Acari: Tetranychidae) on yield, quality, and economics of tomato production. Crop Protection, 52: 84-90. doi:10.1016/j.cropro.2013.05.011 ![]()
Messing R.H., Croft B.A. 1991. Biosystematics of Amblyseius andersoni and A. potentillae (acarina: phytoseiidae): implications for biological control. Exp. Appl. Acarol., 10(3): 267-278. doi:10.1007/BF01198655 ![]()
Migeon A., Dorkeld F., Nouguier E. 2010. Spider mites web: a comprehensive database for the Tetranychidae. Trends in Acarology. Proceedings of the 12th International Congress, Amsterdam (21/VIII/2006-26/VIII/2006): 557-560. doi:10.1007/978-90-481-9837-5\_96 ![]()
Nachman G, Zemek R. 2002. Interactions in a tritrophic acarine predator-prey metapopulation system III: Effects of Tetranychus urticae (Acari: Tetranychidae) on host plant condition. Exp. Appl. Acarol., 25: 27-42.
Nguyen T.T.P., Amano H. 2009. Mating duration and egg production of the predaceous mite Neoseiulus californicus (Acari: Phytoseiidae) vary with temperature. J. Asia-Pacific Entomol., 12(4): 297-299. doi:10.1016/j.aspen.2009.06.003 ![]()
Pozzebon A., Duso C., Pavanetto E. 2002. Effects of some fungicides on phytoseiid mites (Acari, Phytoseiidae) in north-Italian vineyards. Anz. Schadlings., 75(5): 132-136. doi:10.1046/j.1472-8206.2002.02037.x ![]()
Prischmann D.A., James D.G., Wright L.C., Teneyck R.D., Snyder W.E. 2005. Effects of chlorpyrifos and sulfur on spider mites (acari: tetranychidae) and their natural enemies. Biol. Contr., 33(3): 0-334. doi:10.1016/j.biocontrol.2005.03.008 ![]()
Roy M., Brodeur J., Cloutier C. 2002. Relationship between temperature and developmental rate of Stethorus punctillum (Coleoptera: Coccinellidae) and its prey Tetranychus mcdanieli (Acarina: Tetranychidae). Environ. Entomol., 31: 177-187. doi:10.1603/0046-225X-31.1.177 ![]()
Sabelis M.W. 1985a. Development. In: Helle W, Sabelis MW (eds) Spider mites: their biology, natural enemies and control, vol 1B. Elsevier, Amsterdam, The Netherlands, pp 43-45.
Sabelis M.W. 1985b. Sex allocation. In: Helle W, Sabelis MW (eds) Spider mites: their biology, natural enemies and control, vol 1B. Elsevier, Amsterdam, The Netherlands, pp 83-94.
Sabelis M.W., Nagelkerke C.J. 1988. Evaluation of pseudo-arrhenotoky. Exp. Appl. Acarol., 4(3):301-318. doi:10.1007/BF01196192 ![]()
Skirvin D.J., Fenlon J.S. 2003. The effect of temperature on the functional response of Phytoseiulus persimilis (Acari: Phytoseiidae). Exp. Appl. Acarol., 31(1), 37-49. doi:10.1023/B:APPA.0000005107.97373.87 ![]()
Skirvin D.J., Williams M.E.D.C., Fenlon J.S., Sunderland K. 2002. Modelling the effects of plant species on Biocontrol effectiveness in ornamental nursery crops. J. Appl. Ecol., 39(3): 469-480. doi:10.1046/j.1365-2664.2002.00728.x ![]()
Tixier M.-S., Lopes I., Blanc G., Dedieu J.-L., Kreiter S. 2014. Phytoseiidae mites (Acari : Mesostigmata) in French orchards and assessment of their spatial distribution. Acarologia, 54(1): 97-111. doi:10.1051/acarologia/20142114 ![]()
Wang Z.Y., Qin S.Y., Xiao U.F., Liu H. 2014. Effects of temperature on development and reproduction of Euseius nicholsi (Ehara & Lee). Syst. Appl. Acarol., 19(1):44-50. doi:10.11158/saa.19.1.2 ![]()
Wu S.Y., Xie H.C., Li M.Y., Xu X.N., Lei Z.R. 2016. Highly virulent Beauveria bassiana strains against the two-spotted spider mite, Tetranychus urticae, show no pathogenicity against five phytoseiid mite species. Exp. Appl. Acarol., 70(4): 421-435. doi:10.1007/s10493-016-0090-x ![]()
Xia B., Zou Z.W., Li P.X. Lin P. 2012. Effect of temperature on development and reproduction of Neoseiulus barkeri (Acari: Phytoseiidae) fed on Aleuroglyphus ovatus. Exp. Appl. Acarol., 56(1): 33-41. doi:10.1007/s10493-011-9481-1 ![]()
Xu X.N., Lv J.L., Wang E.D. 2015. Predatory mite research in mass rearing and field applications. Chin. J. Biol. Contr., 31: 647-656.
Zhang X.H., Sheng H.M. 2001. The predatory function of Euseius finlandicus (Oudemans) to Tetranychus urticae Koch. J. Gansu Sci., 13(2): 35-37.



2019-01-28
Date accepted:
2019-10-16
Date published:
2019-11-14
Edited by:
Tixier, Marie-Stéphane

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Li, Yu-Jing; Liu, Qing-Yan; Chang, Jing; Jia, Yong-Hong and Meng, Rui-Xia
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)