



A new species of Arrenurus (Acari, Parasitengona, Hydrachnidia) found in the crop of a Yellow-billed Teal Anas flavirostris in Bolivia
Valdecasas, Antonio G.1
1✉ Departamento de Biodiversidad y Biología Evolutiva, Museo Nacional de Ciencias Naturales, CSIC, Madrid, Spain
2019 - Volume: 59 Issue: 2 pages: 253-260
https://doi.org/10.24349/acarologia/20194329ZooBank LSID: 6A5F717F-CE15-40EA-897B-83853C8EBDD4
Original research
Keywords
Abstract
Introduction
Models that aim to explain the distribution of organisms range from absolute dispersion to vicariance, and include those that differentially combine these two extremes (Zink et al. 2000). In some cases, the underlying causal mechanisms of dispersion are unknown, leading to a search of the most parsimonious model to explain a given pattern of organismal occurrence. In other cases, the dispersal mechanisms may be known but the actual distribution of the species is not. A common mechanism of organismal dispersal is via birds (for a recent review on waterfowl dispersal, see Van Leeuwen et al. 2012). This is potentially the case of two specimens of a new water mite species of the genus Arrenurus that were found in the crop of an Anas flavirostris duck in Bolivia. The mites, a male and a female, were in perfect condition; however, it is uncertain if they would have been viable.
Of the nearly 1,000 species of Arrenurus Dugés that have been described worldwide (Gerecke et al. 2016), over 160 are known from South America (Rosso de Ferradas and Fernandez 2005). Of these species, 17 belong to the subgenus Arrenurus. Another 26 species may also belong to this subgenus; however, as their descriptions are based only on female specimens, they cannot be assigned to a subgenus rank (Rosso de Ferradas and Fernandez 2005). The two water mite specimens described herein, a male and a female, represent a new species of Arrenurus (Arrenurus). Moreover, to my knowledge, this study is the first to report the presence of water mites in the crop of a duck.
Materials and methods
The type material, a male and a female, were found in the crop of a Yellow-billed Teal, Anas flavirostris, near Cochabamba (Bolivia). The specimens were fixed in Koenike's fluid (Cook 1974), and the palps, chelicerae and gnathosoma dissected, oriented and permanently mounted on a slide with glycerin jelly (O. Lundblad's formulation provided by T. Gledhill, Freshwater Biological Association, Ambleside, Cumbria, England), together with the main body. The slides were deposited in the invertebrate collection held at the Museo Nacional de Ciencias Naturales, Madrid (MNCN). Abbreviations: P-I, first segment palp; P-II, second segment palp and so on. I-Leg-3, third segment of the first leg; I-Leg-4, fourth segment of the first leg and so on. All measurements are in µm.
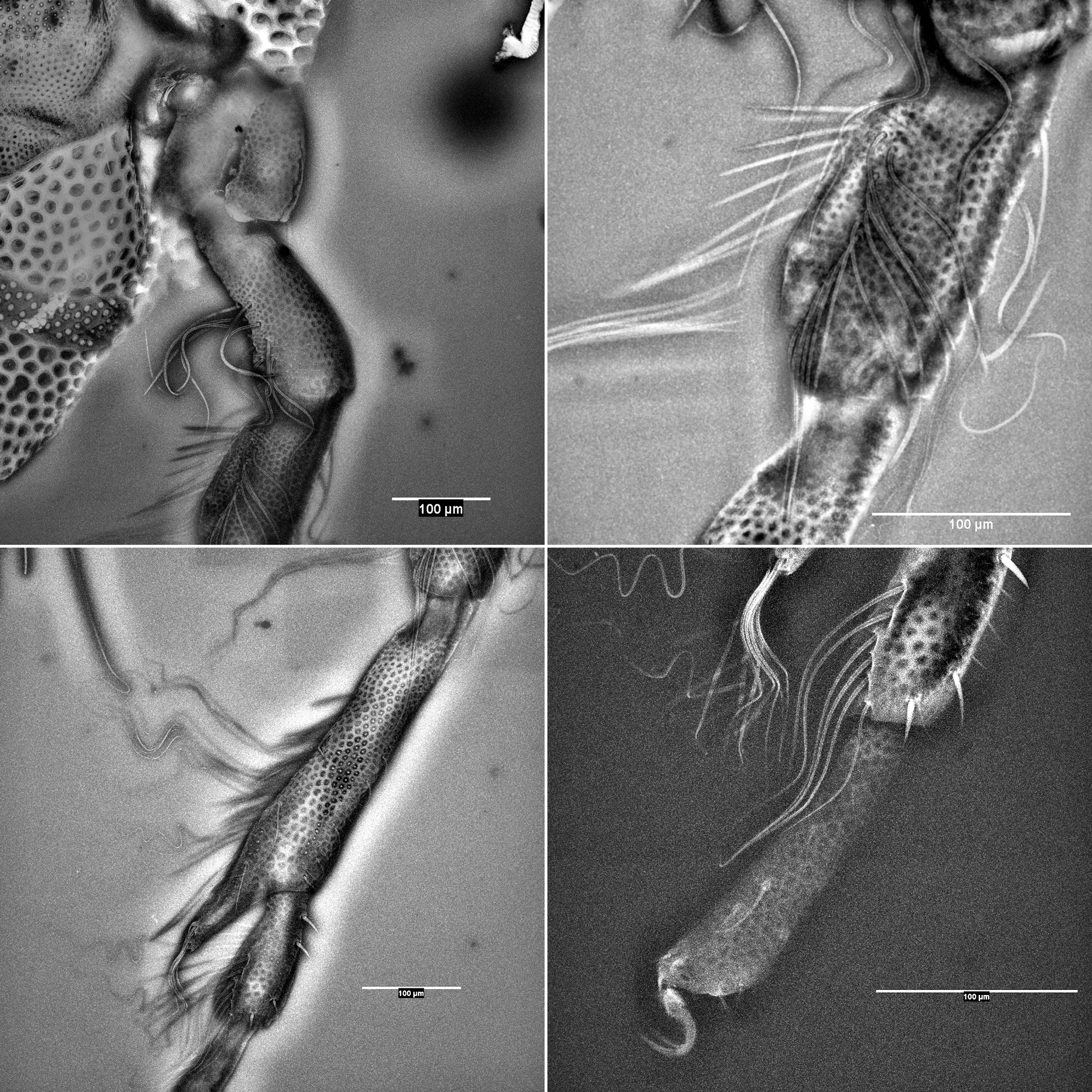
A drawing tube attached to a standard Zeiss compound microscope was used to facilitate drawings. Laser scanning confocal microscopy stacks were acquired with a Leica SPE (for detailed methodology, see Valdecasas and Abad 2010). Image stacks were processed with Fiji/ImageJ and Amira (ver. 5.4.3). The autofluorescence of the mounting medium competed with the natural autofluorescence typically found in arthropod specimens. Therefore, several strategies were used to improve the resolution of the leg setation images: maximum and median projections were both rendered, as was the average of both projections, followed by contrast limited adaptive histogram equalization (CLAHE) and unsharp masking (Malin 1977) plus background subtraction, followed by maximum intensity projection (MIP).
Results
Family: Arrenuridae Thor, 1900
Genus: Arrenurus Dugès, 1834
Subgenus: Arrenurus Dugès, 1834
Type species: Arrenurus viridis Dugès, 1834
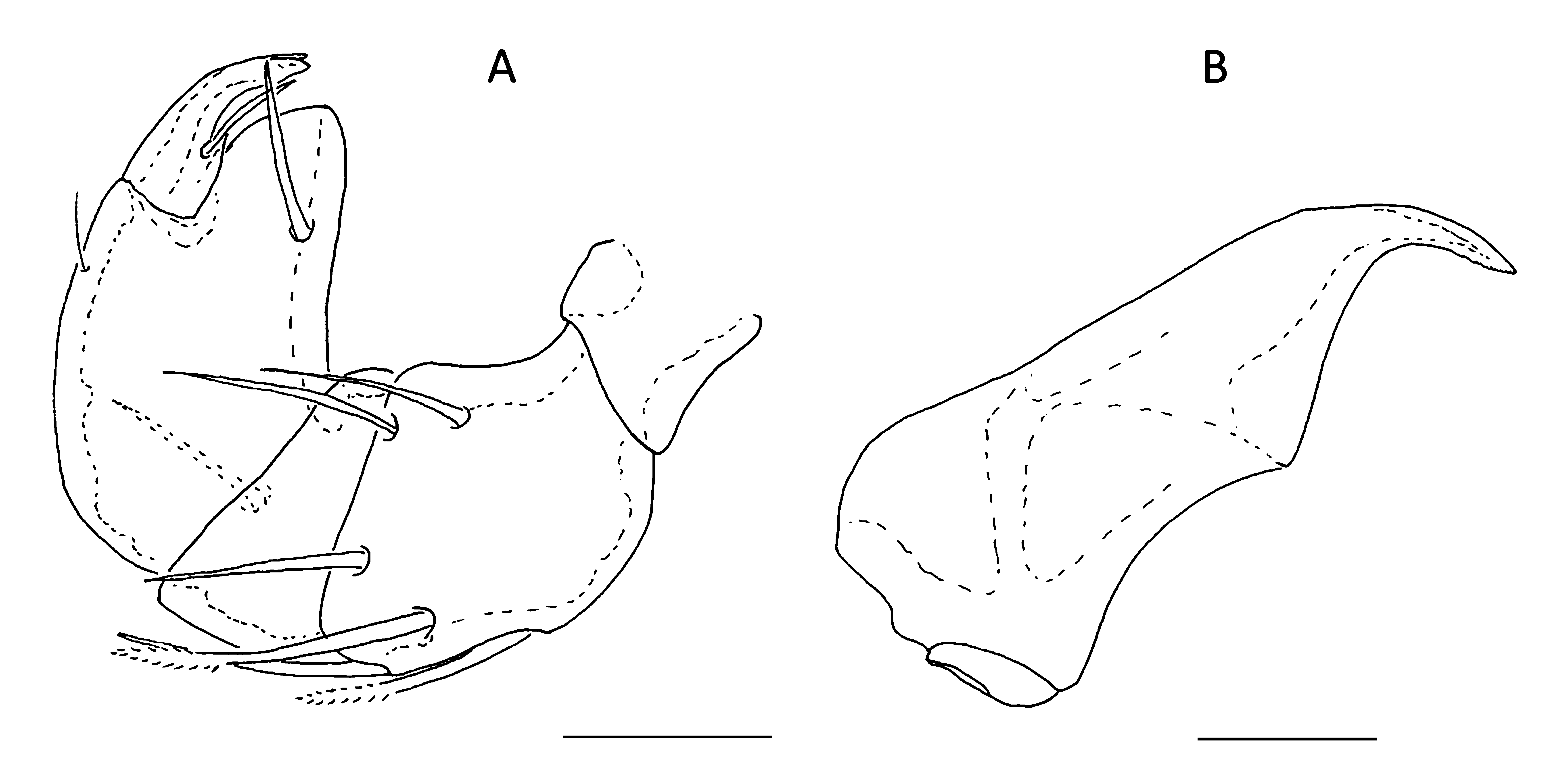
Arrenurus (Arrenurus) caboti n. sp. (Figs 1–6)
ZOOBANK: 3142EB5C-BD53-4BB2-8D6C-C43D70AB8FAD ![]()
Material examined
Holotype male, Cochabamba, Bolivia, found in the crop of a Yellow-billed Teal, Anas flavirostris. 9 January 1983, leg., José Cabot Nieves. Collection number: MNCN 20.02/19390. Paratype, one female, same data as holotype. Collection number: MNCN 20.02/19391.
Description
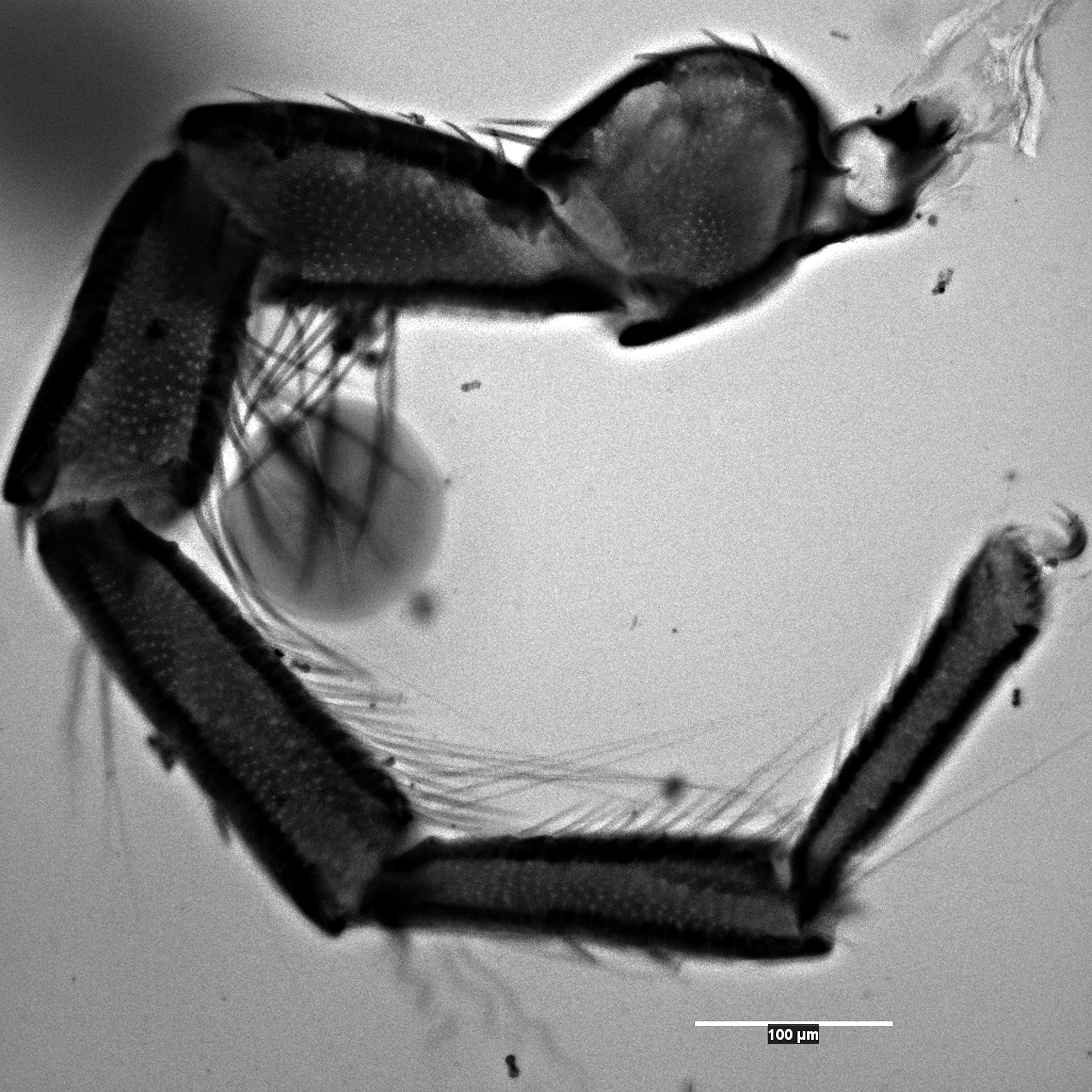
Male — Colour not registered. Body rounded, anterior end convex between the eyes, cauda absent (Fig. 1). Body, excluding petiole, 991 µm long with a maximum width of 966 µm. Cauda indistinct, with a rectangular petiole without an evident notch nor a hyaline membrane. Petiole 91 µm long, with a maximum width at the tip of 69 µm. Dorsal furrow complete, with two medial and two terminal humps. Pygal lobes absent. Gonopore 59 µm long. Acetabular plates extending laterally.

Dorsal length of the palps: P-I, 42; P-II, 87; P-III, 59; P-IV, 95; P-V, 46. P-II with six setae, three of which are pennate. P-III and P-IV with one setae each. Chelicera total length 203 µm (Fig. 2). Gnathosoma 147 µm long and 114 µm wide. Legs have a complex chaetotaxy (see Table 1 and Figs 3 and 4). Dorsal length of leg segments: I-Leg-3: 125; I-Leg-4: 169; I-Leg-5: 159; I-Leg-6: 169; II-Leg-3: 141; II-Leg-4: 188; III-Leg-5: 194; II-Leg-6: 203; III-Leg-4: 234; III-Leg-5: 226; III-Leg-6: 197; IV-Leg-2: 203; IV-Leg-3: 203; IV-Leg-4: 340; IV-Leg-5: 153; IV-Leg-6: 169.
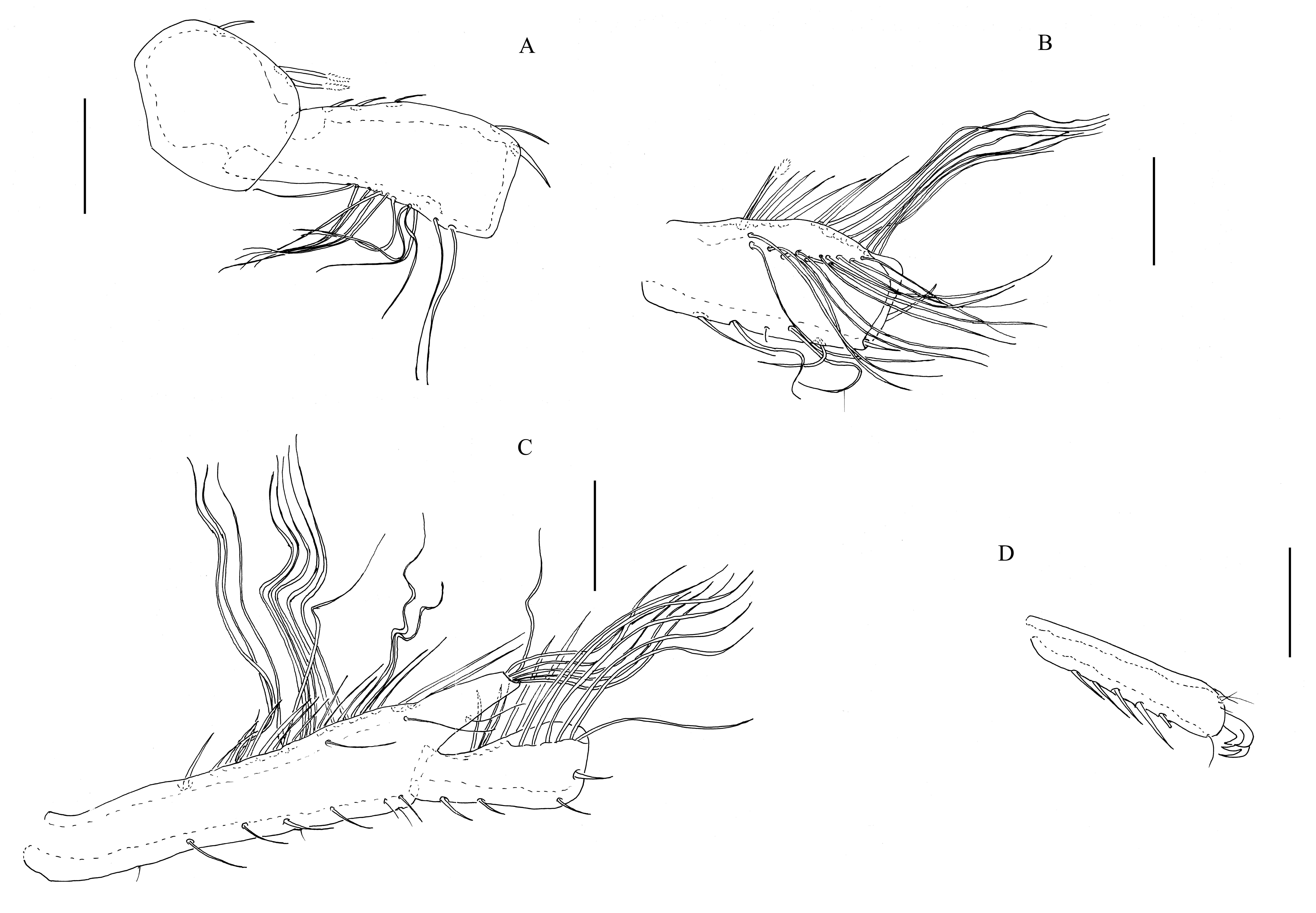


Female — Colour not registered. Body rounded, 1421 µm long with a maximum width of 1290 µm (Fig. 4). Dorsal furrow complete, maximum width 733 µm. Gonopore 172 µm long. Dorsal length of the palps: P-I, 46; P-II, 91; P-III, 70; P-IV, 112; P-V, 57. Palp shape similar to the male; P-II with five setae, and P-III with one setae. Cheliceral shape similar to the male, total length 178 µm. Gnathosoma 200 µm long and 153 µm wide. Dorsal length of the IV-Leg segments: IV-Leg-2: 184; IV-Leg-3: 197; IV-Leg-4: 262; IV-Leg-5: 238; IV-Leg-6: 200. Number of swimming setae on the IV-Leg segments: IV-Leg-3: 11; IV-Leg-4: 9+11; IV-Leg-5: 10 (Table 2 and Fig. 6).

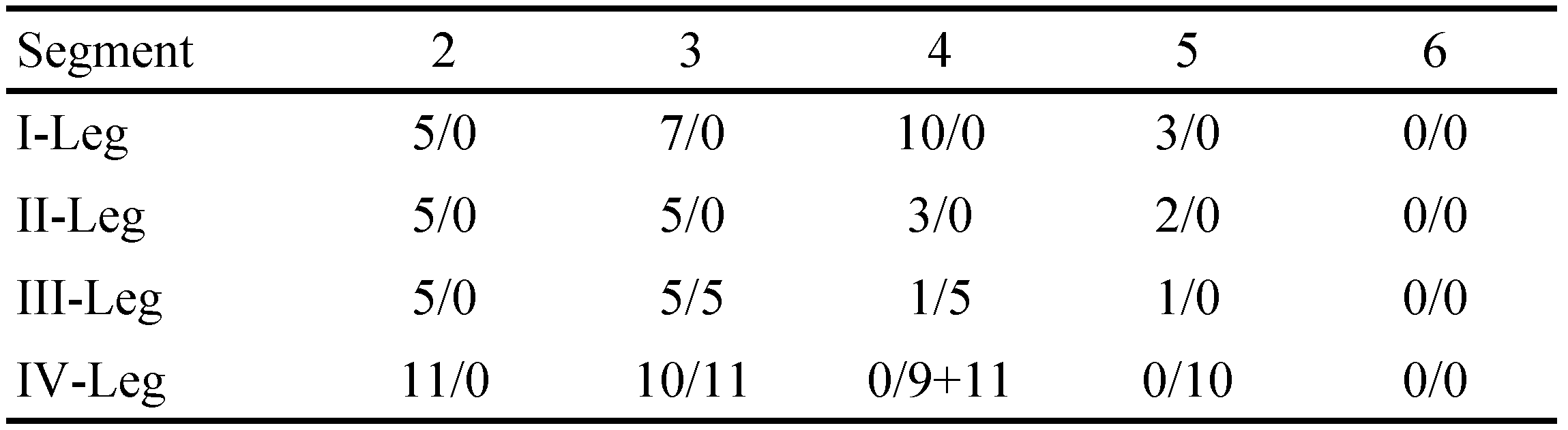
Etymology
The new species is named after José Cabot who gave me the specimens during a stay in Bolivia.
Discussion
Van Leeuwen et al. (2012) provide a review of macroinvertebrate and macrophyte species transported by waterbirds via their faecal droppings, and although crustacean, insect, molluscan and bryozoan species were all recorded, no chelicerates were found. In a more recent study that focuses on the dispersal of wetland plant seeds, Kleyheeg and van Leeuwen (2015) highlight regurgitation as an overlooked dispersal mechanism. Given that this event can happen within a few hours, regurgitated organisms may potentially remain viable. The relevance of this mechanism for the occasional transportation of non-resting stages of invertebrates is unknown. However, as speculative at it may be, De Queiroz (2014) shows that, given sufficient time, almost inconceivable dispersal events can occur.
According to BirdLife International (2019), the Yellow-billed Teal is `resident' in the area where it was found. The bodies of the male and female water mite specimens were complete, and the female had no eggs. Whether the specimens were alive at the time of collection is unknown, as their vital activity was not documented. However, one can speculate that they could have possibly passed through the digestive system of the duck intact due to the hard body composition of the species and, if the female had been gravid and the bird's droppings fell into water, she could have hatched offspring. In another possible scenario, they could have been regurgitated soon after ingestion and been viable as adults. Had the female been gravid, her eggs could have also been viable. Regardless of the potential viability of the two specimens, the present study is the first to report the presence of a water mite species in the crop of a mallard.
Similarities in palp shape and chaetotaxy can be used to identify male and female specimens of the same species within Arrenurus. The two specimens of A. caboti n. sp., besides being found together, share a similar morphology and distribution of palp setae. The new species can be easily distinguished from the 17 species of the subgenus Arrenurus (Arrenurus) that have been registered for South America (Rosso de Ferradas and Fernandez 2005) by, among other characters, the presence of a cauda on the male body of the following species: A. brasiliensis Lundblad, 1941; A. brevipetiolatus Lundblad, 1936; A. erectipetiolatus Viets, 1954; A. flagellatus Lundblad, 1936; A. inflatipalpis Lundblad, 1944; A. lobatus Lundblad, 1941; A. mystrophorus Lundblad 1938; A. oxyurus Ribaga, 1902; A. plaumanni Lundblad, 1936; A. rectipetiolatus Lundblad, 1941; A. ringueleti Rosso de Ferradas, 1980; A. sellatus Viets, 1954; A. subulifer Lundblad, 1938; A. trichophorus Daday, 1905; A. valdiviensis Viets, 1964; A. valencius Marshall, 1919 and A. vicinus Lundblad, 1944.
In their survey of South American Arrenurus species, Rosso de Ferradas and Fernandez (2005) included two other species within the subgenus Arrenurus, A. conspicuus Viets and Böttger, 1986 and A. ripiphorus, Lundblad, 1937, despite their original descriptions being based only on female specimens. The female body shape of A. conspicuus and A. caboti n. sp. are similar; however, the two species differ in the number of setae on P-II, three in A. conspicuus and six in A. caboti n. sp. They also differ in the shape of P-IV, almost rectangular in A. conspicuus but concave-convex in A. caboti n. sp. The new species is easily distinguished from A. ripiphorus, which has an oval-shaped body that is not truncated at the front.
The new species can also be distinguished from the other 24 South American Arrenurus species described only from female specimens by the number and size of setae on P-II and/or the general body shape of the female. Compared to the five setae in A. caboti n. sp., 8 of the 24 species have numerous setae (i.e. brittle brush) on P-II: A. acutiepimeratus Viets, 1954; A. amplipenicillatus Viets, 1954; A. boettgeri Viets, 1968; A. caquetiorum Rosso de Ferradas and Fernandez, 2001; A. dentipalpis Lundblad, 1936; A. hirsutipalpis Walter, 1919; A. mutangulus Daday, 1905 and A. papilliger Viets, 1959. Two of the species, A. tumulosus tumulosus Viets, 1954 and A. tumulosus intercursor Viets, 1954, have 10–12 strong short setae on P-II. The rounded body shape of A. caboti n. sp. clearly differentiates it from the species with an almost square/rectangular body shape (A. acutidentatus Viets, 1959; A. amplipenicillatus Viets, 1954; A. mutangulus Daday, 1905; A. multipapillatus Viets, 1954 and A. uberifer Viets, 1954) and from those with an ovoid body shape including ones that narrow at the anterior end (A. apertus Daday, 1905), the posterior end (A. anisitsi Daday1905 and A. tumulosus tumulosus Viets, 1954) or both ends (A. quadrisetosus Viets, 1954). An almost strictly ovoid female body shape is found in A. boettgeri Viets, 1968; A. caquetiorum Rosso de Ferradas and Fernandez, 2001; A. cometes Lundblad 1937; A. confertus Viets, 1954; A. honoratus Thor, 1911; A. ovum Viets, 1954; A. pennapodus Rosso de Ferradas, 1989; A. promacrus Lundblad, 1936; A. propincuus Daday, 1905 and A. tortus Viets, 1954). Taken all together, the morphological characters described above validate A. caboti n. sp. as a new species of the Arrenurus subgenus.
Acknowledgements
I thank José Cabot for the loan of the of Arrenurus water mite specimens. Melinda Modrell revised the English of several versions of the manuscript. I celebrate the Biodiversity Heritage Library for making old and rare taxonomic literature more accessible.
References
BirdLife International. 2019. Species factsheet: Anas flavirostris. [Internet]. [8 March 2019]. Available from http://www.birdlife.org ![]()
Cook D.R. 1974. Water mite genera and subgenera. Memoirs of the American Entomological Institute, 21, pp. 860.
de Queiroz A. 2014. The Monkey's Voyage: How Improbable Journeys Shaped the History of Life. New York: Basic Books (AZ). pp. 368.
Gerecke R., Gledhill T., Pešić V., Smit H. 2016. Chelicerata: Acari III. In: Gerecke, R., Ed. Süßwasserfauna von Mitteleuropa, 7/2-3. Heidelberg: Springer-Verlag Berlin. pp. 429. doi:10.1007/978-3-8274-2689-5 ![]()
Kleyheeg, E., Van Leeuwen, C.H.A. 2015. Regurgitation by waterfowl: an overlooked mechanism for long-distance dispersal of wetland plant seeds? Aquatic Botany, 127, 1-5. doi:10.1016/j.aquabot.2015.06.009 ![]()
Malin D.F. 1977. Unsharp masking. AAS Photo-Bulletin, 16: 10-13.
Rosso de Ferradas B., Fernandez H.R. 2005. Elenco y Biogeografía de los ácaros acuáticos (Acari, Parasitengona, Hydrachnidia) de Sudamérica. Graellsia, 61: 181-224. doi:10.3989/graellsia.2005.v61.i2.19 ![]()
Valdecasas A.G., Abad A. 2010. Morphological confocal microscopy in arthropods and the enhancement of auto fluorescence after proteinase K extraction. Microscopy and Microanalysis, 17: 109-113. doi:10.1017/S1431927610094213 ![]()
Van Leeuwen C.H.A., Van der Velde G., Van Groenendael J.M., Klaassen M. 2012. Gut travellers: internal dispersal of aquatic organisms by waterfowl. Journal of Biogeography, 39: 2031-2040. doi:10.1111/jbi.12004 ![]()
Zink R.M., Blackwell-Rago R.C., Ronquist F. 2000. The shifting roles of dispersal and vicariance in biogeography. Proceedings of the Royal Society Biological Sciences Series B, 267: 497-503. doi:10.1098/rspb.2000.1028 ![]()



2019-04-01
Date accepted:
2019-06-20
Date published:
2019-07-11
Edited by:
Mąkol, Joanna

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Valdecasas, Antonio G.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)