



Two new species of feather mites of the subfamily Pterodectinae (Analgoidea: Proctophyllodidae) from Indonesia
Constantinescu, Ioana Cristina1 ; Chişamera, Gabriel Bogdan2 ; Petrescu, Angela3 and Adam, Costică4
1✉ “Grigore Antipa” National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest, Romania.
2“Grigore Antipa” National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest, Romania.
3“Grigore Antipa” National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest, Romania.
4“Grigore Antipa” National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest, Romania.
2019 - Volume: 59 Issue: 2 pages: 196-210
https://doi.org/10.24349/acarologia/20194324ZooBank LSID: 3B434AD7-CEDA-46D3-8D1C-C986C3146E36
Original research
Keywords
Abstract
Introduction
The subfamily Pterodectinae is one of the two subfamilies currently recognized within the family Proctophyllodidae, and is divided into two tribes: Rhamphocaulini and Pterodectini. Representatives of the tribe Rhamphocaulini occur exclusively on hummingbirds (Apodiformes: Trochilidae), while the Pterodectini occur mostly on passerines (Passeriformes), in addition to other scattered bird orders. In the host birds' plumage, pterodectines mainly inhabit the primaries and secondaries of the wings, where they are located in corridors on the ventral surface of the vane (Mironov et al. 2012).
According to modern concepts, the genus Montesauria belongs to the Pterodectes generic group, characterized by the posterior or postero-lateral position of setae ps3 in relation to the anal suckers in the male. Along with four other genera it belongs to the Montesauria generic complex, whose representatives have the genital papillae situated at level of the genital arch or posterior to it in the male (Mironov et al. 2012). Based on morphology, the genus Montesauria was divided into nine species groups and its species were revised (Mironov, 2006). Later, an additional species group, macronoi, was proposed (Mironov et al. 2012). This is the species-richest genus of the subfamily Pterodectinae currently including 64 species distributed mostly in the Old World (Till 1954, 1957, Gaud 1957, Gaud and Mouchet 1957, Mironov and Kopij 1996a, 1996b, 1997, Mironov and Fain 2003, Mironov 2006, 2008, Kuroki et al. 2006, Hernandes et al. 2010, Mironov et al. 2010, 2012, Mironov and Tostenkov 2013, Hernandes and OConnor 2017, Constantinescu et al. 2018b). To date, 14 species of Montesauria have been described from Asia: 8 from Vietnam (Gaud and Petitot 1948, Mironov et al. 2012, Mironov and Tostenkov 2013), 2 from Japan (Kuroki et al. 2006), 2 from India (Constantinescu et al. 2018b), 1 from Taiwan (Sugimoto 1941) and 1 from Indonesia (Mironov 2006). Atyeo (in: McClure and Ratanaworabhan 1973), reported 81 supposedly new species of this genus from Southeastern Asia; unfortunately, new species from this material have never been described.
The genus Proterothrix, along with seven other genera, belongs to the Proterothrix generic group, which incorporates putatively early derivative pterodectines with setae ps3 anterior to the adanal suckers in males and seta wa situated anterior to setae ra and la on tarsi I and II in both sexes (Mironov 2009; Mironov and Proctor 2009, Hernandes and Valim 2014, Mironov and OConnor 2017). Out of the 30 species of this genus, twenty nine have been arranged into three species groups: megacaula (3 species), schizothyra (4 species) and wolffi (22 species); one species, P. aculeata (Canestrini, 1899), remains unassigned to any species group (Gaud 1952, 1962, 1968, 1979, Mironov et al. 2008, 2010, 2012, Mironov and Proctor 2009, Mironov and Tolstenkov 2013, Constantinescu et al. 2014, 2017a, 2017b, 2018a). Species of the schizothyra group are restricted to kingfishers (Coraciiformes: Alcedinidae) and those of the wolffi group are known from various passerine families of the infraorders Corvida (Acanthizidae, Dicruridae, Meliphagidae, Monarchidae, Paradisaeidae, Ptilonorhynchidae and Rhipiduridae), and Passerida (Leiothrichidae, Muscicapidae, Paradoxornithidae, Eurylaimidae and Pellorneidae), and the suborder Tyranni (Eurylaimidae), while species of the megacaula group are only known from hosts of the infraorder Passerida (Muscicapidae).
The present paper describes two new feather mite species of pterodectines belonging to the genera Montesauria and Proterothrix, collected from birds in Indonesia (Kalimantan).
Materials and methods
Mites were collected by the authors from museum specimens of birds preserved in ethanol, from the Ornithological Collection of ``Grigore Antipa'' National Museum of Natural History (Bucharest, Romania). The ethanol in which the birds were preserved was filtered through filter paper, then the mite specimens were collected manually with a fine needle placed in vials filled with 96% ethanol. Then mite specimens were cleared in 90% lactic acid for 24 hours, and mounted on microscope slides in Hoyer's medium. Drawings were made using an Olympus CX21 compound microscope, equipped with a camera lucida drawing device. The taxonomy of birds follows Lepage et al. (2019). The body setation of mites follows that of Griffiths et al. (1990) with the modifications by Norton (1998) concerning coxal setae, while the leg setation follows Gaud and Atyeo (1996). Species redescriptions follow the standard style accepted in taxonomic works describing pterodectine mite species (Mironov and Fain 2003, Mironov 2006, Valim and Hernandes 2006, Mironov et al. 2008); we measured a set of standard morphological diagnostic features following Mironov and Proctor (2009). Measurements are given for the holotype (male) and followed by ranges for paratypes. All measurements are in micrometers (μm). The mite specimens studied herein are deposited in the Acari Collection of ``Grigore Antipa'' National Museum of Natural History, Bucharest, Romania (MGAB) and in the Acari Collection of the Department of Zoology of the Universidade Estadual Paulista, Rio Claro, São Paulo, Brazil (DZUnesp-RC).
Taxonomy
Superfamily Analgoidea Trouessart & Mégnin, 1884
Family Proctophyllodidae Trouessart & Mégnin, 1884
Subfamily Pterodectinae Park & Atyeo, 1971
Genus Montesauria Oudemans, 1905
Montesauria indonesiana Constantinescu sp. n.
Figures 1 – 4
ZOOBANK: 6F70104D-14FE-4968-AE23-C08E8B391B45 ![]()
Type material — Holotype male and 16 paratypes (7 male and 9 female), from Aplonis panayensis (Scopoli) (Passeriformes, Sturnidae), INDONESIA, Kalimantan Island, Lhok Tuan, Kutai National Park, 22 May 1991, bird inventory number 15717, no other data.
Type deposition — Holotype male (ANA 801), 6 male (ANA 802–807) and 8 female (ANA 808–815) paratypes in MGAB collection, 1 male and 1 female paratypes in DZUnesp-RC collection.
Etymology — The specific name indonesiana refers to the name of the country (Indonesia) where the mite was collected.

Description — Male. Length of idiosoma 322 (308–312), width 140 (120–140), length of hysterosoma 212 (204–208). Prodorsal shield: entire, anterior margin with lateral margins slightly concave at level of scapular setae, posterior margin with acute median extension, length along midline 110 (104–108), width at posterior margin 118 (100–118), surface with numerous pit-like lacunae. Scapular setae se separated by 65 (58–64). Humeral shields small, separated from epimerites III, not encompassing setae cp. Setae c2 situated dorsally on soft tegument. Subhumeral setae c3 lanceolate, 18 (16–18) × 8 (6–8). Hysteronotal shield: greatest length from anterior margin to lobar apices 200 (196–204), width at anterior margin 122 (102–120), anterior margin slightly convex, surface of this shield with numerous pit-like lacunae. Prodorsal and hysteronotal shields separated by narrow band of soft tegument. Opisthosomal lobes short, roughly trapezoidal, with pair of small extensions at base of setae h2 and with extensions at base of setae h3, terminal cleft V-shaped, 24 (22–24) long. Setae f2 absent. Setae h3 lanceolate, with acute tip, 38 (36–37) long, 12 (10–11) wide, setae h2 150 (140–160) long, 5 (4–5) wide. Supranal concavity circular, well outlined. Setae ps2 66 (54–60) long, thickened in basal part and with filiform apical part, setae ps1 filiform, about 10 (6–8) long, situated on margins of terminal cleft slightly anterior to level of setae h3 (Fig. 1A). Distance between bases of dorsal setae: c2–d2 104 (84–100), d2–e2 96 (84–100), e2–h3 52 (48–52), d1–d2 46 (40–50), e1–e2 22 (22–26), ps2–h1 10 (6–10), h2–h2 40 (42–44), h3–h3 22 (20–22), ps2–ps2 70 (64–66).
Epimerites I fused into a Y, posterior end of sternum connected to medial part of epimerites II (Fig. 1B). Coxal fields I, II without wide sclerotized areas. Rudimentary sclerites rEpIIa present. Coxal fields II and III open. Epimerites IVa present. Genital arch small, 16 (17–20) in length, 26 (24–28) in width, basal sclerite of genital apparatus shaped semicircular, aedeagus straight, sword-shaped, 70 (62–65) long, extending to or beyond anterior margin of anal suckers. Genital shield absent, adanal shields present, small, circular, surrounding base of setae g. Anal suckers 14 (13–14) in diameter, corolla without indentations, surrounded by radially striated area. Opisthoventral shields small, circular, surrounding base of setae ps3 (Fig. 1B). Distance between ventral setae: 3a–4b 30 (28–32), 4b–4a 28 (28–32), 4a–g 44 (42–46), g–ps3 40 (40–41), ps3–ps3 66 (62–64), ps3–h3 38 (34–37).

Legs I noticeably longer and thicker than legs II. Genua and tibiae I, II with narrow dorsal crest, femora I, II with ventro-basal crest. Solenidion σ1of genu I lanceolate, setae cGI and cGII spine-like (Fig. 2A, B). Tarsus IV 20 (20–22) long, with small apical claw-like process, modified setae d, e button-like (Fig. 2D). Length of solenidia: ω1I 11 (12–14), ω1II 6 (6–9), φI 68 (60–68), φII 46 (40–44), φIII 24 (20–24), φIV 25 (20–26).


Female (range for 5 paratypes). Length of idiosoma excluding terminal appendages 452–460, width 139–160, length of hysterosoma 300–312. Prodorsal shield: shape as in male, except for convex posterior margin, 144–152 long, 122–142 wide, posterior part with pit-like lacunae (Fig. 3A). Scapular setae se separated by 68–76. Humeral shields small, not encompassing setae cp. Setae c2 situated on soft tegument. Setae c3 lanceolate, 17–18 × 6–7. Prodorsal and hysteronotal shields touching each other. Anterior and lobar parts of hysteronotal shield well delimited from each other by a transverse groove. Hysteronotal shield: anterior margin slightly convex, posterior margin concave in median part, length 236–248, width at anterior margin 130–148, with pit-like lacunae. Length of lobar region excluding terminal appendages 70–78, width at level of setae h2 83–84, median part of anterior margin slightly convex. Terminal cleft as narrow V, 56–60 long, extended beyond level of setae h2. Lateral margins of opisthosomal lobes posterior to setae h2 membranous. Setae f2 absent, setae h1 on lobar shield, near its anterior margin. Setae h2 spindle-like, 36–38 long, 8–10 wide. Setae h3 filiform, 16–21 in length, about 1/6 of the length of terminal appendages. Setae ps1 situated close to lateral margins of opisthosomal lobes (Fig. 3A). Distance between bases of dorsal setae: c2–d2 82–104, d2–e2 100–116, e2–h2 44–48, h2–h3 54–60, d1–d2 34–47, e1–e2 56–60, h1–h2 14–16, ps1–h2 22–29, h1–h1 60–61, h2–h2 68–72.
Epimerites I fused into a Y, posterior end of sternum with acute extensions directed laterally but not connected with epimerites II (Fig. 3B). Lateral parts of coxal fields I without heavily sclerotized areas, coxal fields II with heavy sclerotized areas. Epimerites II do not reach the level of anterior margin of epigynum. Epimerites IVa absent. Translobar apodemes of opisthosomal lobes present, fused with ventral sclerotization of lobar region and posterolateral margins of anterior hysteronotal shield. Epigynum horseshoe-shaped, greatest width 52–65, apodemes of ovipore connected with coxal apodemes IIIa. Head of spermatheca weakly sclerotized, primary spermaduct with ball-shaped enlargement, secondary spermaducts 2 times longer than distance between enlargement of primary spermaduct and spermatheca (Fig. 4E). Distance between pseudanal setae: ps2–ps2 56–58, ps3–ps3 20–24, ps2–ps3 24–28.
Legs I hypertrophied, tibia and genu I strongly inflated on dorsal side, heavily sclerotized and 2.5–3 times thicker than respective segments of legs II. Tibia I with well-developed dorsal crest, its proximal part rounded with indentations, distal part of genu I with large angular paraxial crest and with narrow antaxial crests. Tibia and genu II with narrow dorsal crest and femur II with ventro-basal crest. Genu IV with rounded dorsal crest. Solenidion σ1 of genu I lanceolate, setae cGI and cGII spine-like (Fig. 4A, B). Length of solenidia: ω1I 16 (14–16), ω1II 8 (6–10), φI 72 (66–72), φII 44 (40–48), φIII 24 (22–24), φIV 7 (7–8).
Differential diagnosis. The new species, Montesauria indonesiana sp. n., belongs to the pachypa species group. This new species is most similar to M. mainati (Trouessart, 1885), which was originally described from Common hill myna Gracula religiosa Linnaeus, 1758 (Passeriformes, Sturnidae) in Indonesia (Trouessart 1885) and later redescribed (Mironov 2006). Both species have similar ornamentation of the dorsal shields and setae cGI and cGII are spiniform in male and female. In males of both species, setae h3 are lanceolate, setae ps2 are thickened in the basal part and filiform in apical part, and epimerites IVa are present. In females, the terminal cleft is extended beyond the level of setae h2. M. indonesiana sp. n. clearly differs from M. mainati by the following character states in males (corresponding character states of M. mainati are in parentheses): the prodorsal and hysteronotal shields are separated by a narrow band of soft tegument (vs. both shields almost touching), the terminal cleft has a triangular anterior end (vs. round), the rudimentary sclerites rEpIIa are present (vs. absent), the adanal shields are present (vs. absent), the opisthoventral shields are small, circular, surrounding the base of setae ps3 (vs. narrow, with short extension bearing setae ps3). In females of the new species, the anterior and lobar parts of hysteronotal shields are well delimited from each other by a transverse groove (vs. poorly delimited by a pair of thin striae on the sides of the opisthosoma), the dorsal crest of genu IV has a round angle (vs. acute angle), the dorsal crest of tibia I has its proximal margin rounded, with indentations (vs. sharp and without indentations), the dorsal crest of genu IV has a round angle (vs. acute angle).
Genus Proterothrix Gaud, 1968
Proterothrix murariui Constantinescu sp. n.
Figures 5–8
ZOOBANK: BB2C9D83-9308-433C-A422-5606060C5283 ![]()
Type material — Holotype male and 19 paratypes (9 males and 10 females), from Meiglyptes tristis (Horsfield) (Piciformes, Picidae), INDONESIA, Kalimantan Island, Lhok Tuan, Kutai National Park, 22 May 1991, bird inventory number 15755, no other data.
Type deposition. Holotype male (ANA 1453), 8 male (ANA 1454–1461) and 9 female (ANA 1462–1470) paratypes in MGAB collection, 1 male and 1 female paratypes in DZUnesp-RC collection.
Etymology — This species is named in honour of Dr. Dumitru Murariu, mammologist, correspondent member of the Romanian Academy, who led the expedition organized by ``Grigore Antipa'' National Museum of Natural History in the islands of the Indonesian archipelago in 1991, and collected himself some of the bird specimens on which the new species of feather mite was identified.

Description — Male (holotype, range for 4 paratypes). Length of idiosoma 244 (244–248), width 94 (106–124), length of hysterosoma 162 (164–166). Prodorsal shield entire, with lateral longitudinal bands of sclerotized tegument in anterior third, anterolateral extensions short and with acute tips, lateral margins with a short incision anterior to bases of setae se, posterior margin with wide blunt-angular extension, posterior angles round, length 78 (78–88), width 76 (72–76), surface without ornamentation (Fig. 5A). Scapular setae se separated by 44 (40–42). Scapular shields well developed. Humeral shields well developed, separated from outer sclerotized areas of epimerites III. Setae cp spiculiform, situated ventrally, setae c2 filiform, situated dorsally on humeral shields. Subhumeral setae c3 lanceolate, 18 (18–19) × 6 (4–6). Hysteronotal shield with anterior margin concave and wavy lateral margins, anterior angles rounded, distance from anterior margin to bases of setae h3 145 (146–152), greatest width in anterior part 72 (68–74), surface without ornamentation. Opisthosomal lobes short and narrow, with two almost trapezoidal membranous extensions at the base of setae h3 and h2. Setae h3 leaf-like, lanceolate with acute apex, 60 (60–70) × 16 (13–16). Setae h2 situated slightly anterior to setae h3, 120 (120–140) × 5 (5–6). Terminal cleft narrowly ovate, 20 (20–22) in length. Supranal concavity clearly outlined, triangular. Setae f2 slightly anterior to bases of setae ps2. Setae h1 near lateral margins of opisthosoma. Setae ps1 filiform, length 10 (6–8), situated near lateral margins of opisthosomal lobes, anterior to bases of setae h3. Setae c1 absent. Dorsal measurements: c2–d2 50 (48–56), d2–e2 51 (52–56), e2–h3 46 (46–56), d1–d2 20 (18–22), e1–e2 26 (22–24), h1–ps2 14 (14–20), h2–h2 43 (40–44), h3–h3 20 (20–22), ps2–ps2 56 (54–60).
Epimerites I fused into a Y, with a short sternum, posterior end of sternum with acute extensions directed laterally but not connected with epimerites II. Epimerites II long, with acute extensions on inner margins. Coxal field I almost closed, coxal field II and III open, coxal fields IV with narrow sclerotized areas at bases of trochanters IV. Epimerites IVa present, well developed (Fig. 5B). Pregenital sclerite (epiandrum) present, large. Genital arch 24 (24–25) long, 24 (20–24) wide, aedeagus band-shaped thickened in anterior half and narrowed in posterior half, 39 (40–42) long, extending to level of setae g. Genital papillae situated anterior to genital arch. A pair of adanal shields present, represented by slim plates, bearing setae ps3. Anal suckers 14 (12–14) in diameter, corolla indented, with 9 small teeth. Measurements of ventral side: 3a–4b 20 (20–22), 4b–4a 22 (20–22), 4a–g 34 (33–36), g–ps3 23 (22–24), ps3–ps3 8 (7–8), ps3–h3 44 (43–44).

Legs I longer than legs II, femora II with ventral crest (Fig. 6A, B). Seta e of tarsus I filiform. Setae mGI and mGII spiculiform, setae d of tarsi II, III much shorter than corresponding setae f. Tarsus IV 22 (20–22) long, setae w situated on small protuberance of tarsus, setae d, e button-like, seta d bigger in diameter than e, situated in basal and apical parts of segment, respectively (Fig. 6D). Length of solenidia: ω1I 10 (10–12), ω1II 7 (6–8), φI 46 (44–48), φII 36 (36–41), φIII 16 (16–18), φIV 26 (24–28).
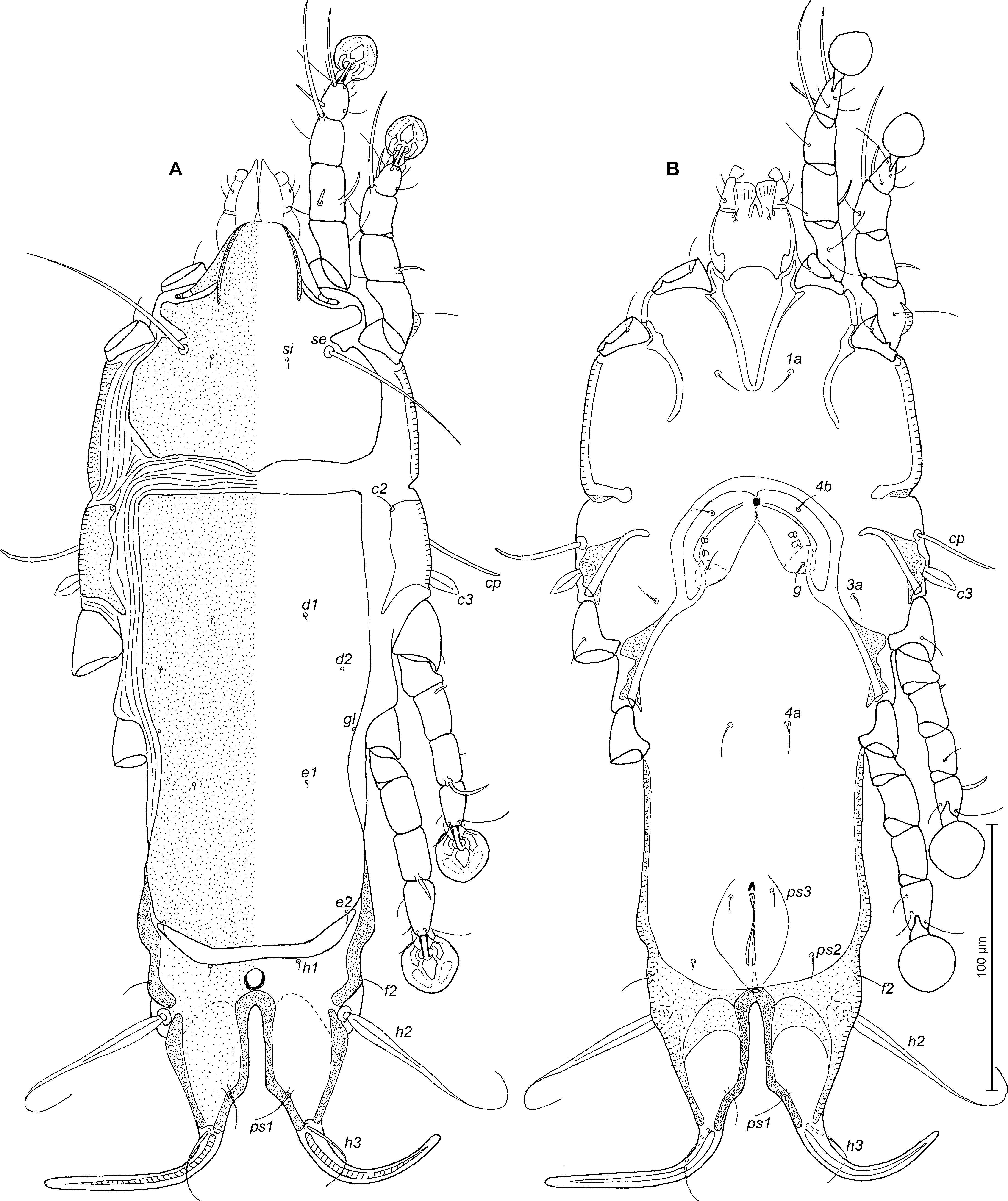
Female (Figs. 7A, B; 8A–E; range for 5 paratypes): Length of idiosoma 344–360, width 126–134, and length of hysterosoma 248–262. Prodorsal shield entire, with lateral longitudinal bands of sclerotized tegument in anterior third, anterolateral extensions with rounded tips, lateral margins with short incisions anterior to bases of setae se, posterior margin with wide blunt-angular extension, length 90–94, width 96–100, surface without ornamentation (Fig. 7A). Scapular setae se separated by 54–57. Scapular shields narrow. Humeral shields well developed, separated from outer sclerotized areas of epimerites III. Setae cp spiculiform, situated ventrally, setae c2 filiform, situated dorsally on humeral shields. Subhumeral setae c3 lanceolate, 17–20 × 6–7. Anterior hysteronotal shield roughly rectangular, anterior margin straight, posterior margin convex, greatest length 160–176, greatest width in anterior part 90–94, surface without ornamentation. Length of lobar region 70–80, width at level of setae h2 76–82. Terminal cleft narrow parallel-sided in anterior 2/3 and large and with oblique margins in posterior 1/3, 48–54 in length. Setae ps1 closer to inner margins than outer margins of opisthosomal lobes. Supranal concavity well developed, ovoid. Setae h1 on lobar shield, near anterior margin of this shield, surface of lobar shield without ornamentation. Setae h2 spindle-shaped, with terminal filament, 72–84 × 6–9. Setae h3 24–36 long, about 1/2 from the length of terminal appendages. Dorsal measurements: c2–d2 62–66, d2–e2 86–92, e2–h2 40–43, h2–h3 44–50, d1–d2 25–28, e1–e2 37–48, h1–h2 24–28, h2–ps1 38–42, h1–h1 36–38, h2–h2 60–70.

Epimerites I fused as a V. Coxal fields I, II without sclerotized areas, epimerites IVa absent (Fig. 7B). Translobar apodemes of opisthosomal lobes present, fused to each other anterior to terminal cleft. Epigynum horseshoe-shaped, greatest width 60–64. Secondary spermaducts short, about 10 long (Fig. 8E). Distance between pseudanal setae: ps2–ps2 46–54, ps3–ps3 15–20, ps2–ps3 28–34.
Legs I slightly longer than legs II, femora II with wide ventral crest, setae mGI and mGII spiculiform. Length of solenidia: ω1I 10–13, ω1II 6–10, φI 50–51, φII 40–44, φIII 14–19, φIV 6–8 (Fig. 8A–D).
Remarks — Proterothrix murariui Constantinescu, sp. n. is the first species of the genus discovered on a host of the family Picidae. This species has unique combination of character states and does not resemble any of the species of Proterothrix described so far. It cannot be included in any of the species groups defined so far within the genus. Males of Proterothrix murariui have three morphological character states within are unique in Proterothrix: a pregenital sclerite (epiandrum) present; the aeadegus is band-shaped, thickened in the anterior half and narrowed in the posterior half; and the terminal lobes have two pairs of trapezoidal, membranous extensions. In both sexes, dorsal setae c1 are absent, and setae cp are spiculiform. Females have the terminal cleft narrow, parallel-sided in the anterior 2/3, and large with oblique margins in the posterior 1/3. The presence of epiandrum in males is a rare occurrence among mites of the subfamily Pterodectinae, encountered only in some species of the genus Anisodiscus (Hernandes and OConnor 2013). The absence of dorsal setae c1 is a character also found to some species of the Proterothrix megacaula species group. It is too premature to define a new group of species within the genus Proterothrix, based only on this species described here, but maybe the discovery of other species of the genus from woodpeckers, sharing common morphological characters, will make this possible in the future.
Acknowledgements
We are very grateful to the research team who collected bird specimens from some islands of the Indonesian archipelago in the expedition organized by ``Grigore Antipa'' National Museum of Natural History in 1991: Dr. Dumitru Murariu, the chief of expedition, Dr. Modest Guțu, Dr. Alexandru Marinescu and Dr. Corneliu Pârvu.
References
Constantinescu I.C., Chişamera G.B., Adam C. 2018a. Redescription of six feather mite species of the genus Proterothrix Gaud, 1968 (Analgoidea: Proctophyllodidae: Pterodectinae) from the "Édouard Louis Trouessart" Collection. Zootaxa, 4486: 451-479. doi:10.11646/zootaxa.4486.4.3 ![]()
Constantinescu I.C., Chişamera G.B., Mukhim D.K.B., Adam C. 2014. Three new species of feather mite of the genus Proterothrix Gaud, 1968 (Analgoidea: Proctophyllodidae: Pterodectinae) from passerines in Meghalaya, North East India. Syst. Parasitol., 89: 45-58. doi:10.1007/s11230-014-9508-1 ![]()
Constantinescu I.C., Cobzaru I., Geamana N.A., Mukhim D.K.B., Adam C. 2017a. Two new species of feather mites (Acarina: Psoroptidia) from the blue-throated blue flycatcher, Cyornis rubeculoides (Passeriformes: Muscicapidae). J. Nat. Hist., 51: 277-297. doi:10.1080/00222933.2017.1280194 ![]()
Constantinescu I.C., Popa O.P., Popa L.O., Cobzaru I., Mukhim D.K.B., Adam C. 2017b. A new feather mite species of the genus Proterothrix Gaud, 1968 (Acarina, Proctophyllodidae) from the Large Niltava, Niltava grandis (Passeriformes, Muscicapidae) - an integrative description. ZooKeys, 661: 1-14. doi:10.3897/zookeys.661.11793 ![]()
Constantinescu I.C., Popa O.P., Popa L.O., Cobzaru I., Mukhim D.K.B., Adam C. 2018b. Two new feather mite species of the genus Montesauria Oudemans (Analgoidea: Proctophyllodidae) from thrushes (Passeriformes: Turdidae) in the Indian Subcontinent. Acarologia, 58: 881-896. doi:10.24349/acarologia/20184294 ![]()
Gaud J. 1952. Sarcoptides plumicoles des oiseaux de Madagascar. Mém. LʼInst. Scien. Madagascar, 7: 81-107.
Gaud J. 1957. Acariens plumicoles (Analgesoidea) parasites des oiseaux du Maroc. Bull. Soc. Sci. Nat. Phys. Maroc, 37: 105-136.
Gaud J. 1962. Sarcoptiformes plumicoles (Analgesoidea) parasites dʼoiseaux de IʼIle Rennell. The Nat. Hist. Rennell Isl., Brit.Solomon Is., 4: 31-51.
Gaud J. 1968. Sarcoptiformes plumicoles (Analgoidea) parasites dʼoiseaux de IʼIle Rennell. The Nat. Hist. Rennell Isl., Brit.Solomon Is., 5: 121-151.
Gaud J. 1979. Sarcoptiformes plumicoles des oiseaux Coraciiformes dʼAfrique. II. Parasites des Alcedinidae. Rev. Zool. Afr., 93: 245-266.
Gaud J., Atyeo W.T. 1996. Feather mites of the world (Acarina, Astigmata): The supraspecific taxa. Ann. Mus. roy. Afr. centr., 277:1-193 (Part 1, text), 1-436 (Part 2, illustrations).
Gaud J., Mouchet J. 1957. Acariens plumicoles (Analgesoidea) des oiseaux du Cameroun. I. Proctophyllodidae. Ann. Parasitol. Hum. Comp., 32: 491-546.
Gaud J., Petitot M.L. 1948. Sarcoptides plumicoles des oiseaux d'Indochine. Ann. Parasit. Hum. Comp., 23: 337-347. doi:10.1051/parasite/1948235337 ![]()
Griffiths D.A., Atyeo W.T., Norton R.A., Lynch C.A. 1990. The idiosomal chaetotaxy of astigmatid mites. J. Zool., 220: 1-32. doi:10.1111/j.1469-7998.1990.tb04291.x ![]()
Hernandes F.A., OConnor B.M. 2013. A review of the genus Anisodiscus Gaud & Mouchet (Acari, Proctophyllodidae) from Madagascar sunbirds (Passeriformes: Nectariniidae), with observations on male genital morphology. Internat. J. Acarol., 35: 423-434. doi:10.1080/01647954.2013.813583 ![]()
Hernandes F.A., OConnor B.M. 2017. Out of Africa: the mite community (Arachnida: Acariformes) of the common waxbill, Estrilda astrild (Linnaeus, 1758) (Passeriformes: Estrildidae) in Brazil. Parasite. Vector., 10: 299. doi:10.1186/s13071-017-2230-5 ![]()
Hernandes F.A., Valim M.P. 2014. On the identity of two species of Proctophyllodidae (Acari: Astigmata: Analgoidea) described by Herbert F. Berla in Brazil, with a description of Lamellodectes gen. nov. and a new species. Zootaxa, 3794: 179-200. doi:10.11646/zootaxa.3794.1.8 ![]()
Hernandes F.A., Valim M.P., Mironov S.V. 2010. On the identity of Pterodectes ralliculae Atyeo and Gaud, 1977 (Astigmata: Proctophyllodidae). J. Nat. Hist., 44: 369-377. doi:10.1080/00222930903383594 ![]()
Kuroki T., Nagahori M., Mironov S.V. 2006. Two New Feather Mite Species of the Genus Montesauria (Astigmata: Proctophyllodidae) from Thrushes of the Genus Zoothera (Passeriformes: Turdidae) in Japan. J. Acarol. Soc. Jpn., 15: 55-68. doi:10.2300/acari.15.55 ![]()
Lepage D. Avibase - Bird Checklists of the World [Internet]. [15 January 2019]. Available from: https://avibase.bsc-eoc.org/checklist.jsp?region=ID&list=clements ![]()
McClure H.E., Ratanaworabhan N. 1973. Some ectoparasites of the birds of Asia, Applied Scientific Research, Jintana Printing Ldt, Bangkok. pp. 219.
Mironov S.V. 2006. Feather mites of the genus Montesauria Oudemans (Astigmata: Proctophyllodidae) associated with starlings (Passeriformes: Sturnidae) in the Indo-Malayan region, with notes on systematic of the genus. Acarina, 14: 21-40.
Mironov S.V. 2008. Three new species of the feather mite subfamily Pterodectinae (Acari: Astigmata: Proctophyllodidae) from passerines (Aves: Passeriformes) in Central Africa. Ann. Zool., 58: 403-418. doi:10.3161/000345408X326744 ![]()
Mironov S.V. 2009. Phylogeny of feather mites of the subfamily Pterodectinae (Astigmata: Proctophyllodidae) and their host associations with passerines (Aves: Passeriformes). Proc. Zool. Inst. Russ. Acad. Sci., 313: 97-118.
Mironov S.V., Fain A. 2003. New species of feather mite subfamily Pterodectinae (Astigmata: Proctophyllodidae) from African passerines (Aves: Passeriformes). Bull. Annls. Soc. Roy. Belge. Ent., 139: 75-91.
Mironov S.V., Kopij G. 1996a. New feather mite species (Acarina: Analgoidea) from some starlings (Passeriformes: Sturnidae) of South Africa. J. Afr. Zool., 110: 257-269.
Mironov S.V., Kopij G. 1996b. Three new species of the feather mite family Proctophyllodidae (Acarina: Analgoidea) from some south African passerine birds (Aves: Passeriformes). Acarina, 4: 27-33.
Mironov S.V., Kopij G. 1997. New feather mite species of the subfamily Pterodectinae (Astigmata: Proctophyllodidae) from some passerines (Aves: Passeriformes) of South Africa. J. Afr. Zool., 111: 449-463.
Mironov S.V., OConnor B.M. 2017. A new feather mite of the genus Neodectes Park and Atyeo 1971 (Acari: Proctophyllodidae) from New Zealand wrens (Passeriformes: Acanthisittidae). Acta Parasitol., 62: 171-177. doi:10.1515/ap-2017-0020 ![]()
Mironov S.V., Proctor H.C. 2009. Feather mites of the genus Proterothrix Gaud (Astigmata: Proctophyllodidae) from parrotbills (Passeriformes: Paradoxornithidae) in China. J. Parasitol., 95: 1093-1107. doi:10.1645/GE-1961.1 ![]()
Mironov S.V., Tolstenkov O.O. 2013. Three new feather mites of the subfamily Pterodectinae (Acari: Proctophyllodidae) from passerines (Aves: Passeriformes) in Vietnam. Proc. Zool. Inst. Russ. Acad.Sci., 317: 11-29.
Mironov S.V., Diao W., Zhang Y., Zhang C., Yan Z. 2008. A new feather mite species of the genus Proterothrix Gaud (Astigmata, Proctophyllodidae) from Ficedula zanthopygia (Hay) (Passeriformes: Muscicapidae) in China. Acarina, 16: 31-38.
Mironov S.V., Literak I., Čapek M., Koubek P. 2010. New species of the feather mite subfamily Pterodectinae (Astigmata, Proctophyllodidae) from passerines in Senegal. Acta Parasitol., 55: 399-413. doi:10.2478/s11686-010-0051-1 ![]()
Mironov S.V., Literak I., Hung M.N., Čapek M. 2012. New feather mites of the subfamily Pterodectinae (Acari: Proctophyllodidae) from passerines and woodpeckers (Aves: Passeriformes and Piciformes) in Vietnam. Zootaxa, 3440: 1-49. doi:10.11646/zootaxa.3440.1.1 ![]()
Norton A.R. 1998. Morphological evidence for the evolutionary origin of Astigmata (Acari: Acariformes). Exp. Appl. Acarol., 22: 559-594. doi:10.1023/A:1006135509248 ![]()
Sugimoto M. 1941. Studies on the Formosan mites (Fourth Report) (On the feather mites, Analgesidae Canestrini, 1892, Part II). Sylvia, 5: 129-149.
Till W.M. 1954. Five new feather mites of the genus Pterodectes (Acarina: Analgesoidea). Moçambique, 79: 85-100.
Till W.M. 1957. Two new Pterodectes species from passeriform birds (Acarina: Proctophyllodidae). J. Entomol. Soc. South. Afr., 20: 450-453.
Trouessart E.L. 1885. Note sur la classification des Analgésiens et diagnoses d'espèces et de genres nouveaux. Bull. Soc. Etud. Scient. Angers, 14: 46-89.
Valim M.P., Hernandes F.A. 2006. Redescription of four species of the feather mite genus Pterodectes Robin, 1877 (Acari: Proctophyllodidae: Pterodectinae) described by Herbert F. Berla. Acarina, 14: 41-55.



2019-01-23
Date accepted:
2019-05-03
Date published:
2019-05-20
Edited by:
Auger, Philippe

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Constantinescu, Ioana Cristina; Chişamera, Gabriel Bogdan; Petrescu, Angela and Adam, Costică
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)