



Predation and oviposition potential of Brazilian populations of the predatory mite Amblyseius tamatavensis (Acari: Phytoseiidae) on eggs of Bemisia tabaci (Insecta: Hemiptera)
Massaro, Marcela1 and Moraes, Gilberto J. de2
1✉ Departamento de Entomologia e Acarologia, Escola Superior de Agricultura 'Luiz de Queiroz', Universidade de São Paulo, Piracicaba, Brazil.
2Departamento de Entomologia e Acarologia, Escola Superior de Agricultura 'Luiz de Queiroz', Universidade de São Paulo, Piracicaba, Brazil.
2019 - Volume: 59 Issue: 1 pages: 120-128
https://doi.org/10.24349/acarologia/20194314Original research
Keywords
Abstract
Introduction
The whitefly Bemisia tabaci (Gennadius) is a pest of great economic importance attacking a variety of crops, as cotton, beans, soybeans, ornamental plants and vegetables (Brown et al . 1995; de Barro et al. 2011; Lourenção et al. 2015) and transmitting about 128 plant viruses (Hogenhout et al. 2008). The control of this pest/ vector is mainly performed using pesticides. However, due to the high reproductive potential, wide host range (allowing easy migration between crops) and numerous cases of pesticide resistance, this method is not effective enough. Moreover, there is a growing concern about maintaining or improving the quality of agricultural products and environment, searching for less harmful methods for controlling pests, including biological control. In this context, some predatory mites of the family Phytoseiidae have been used for the biological control of B. tabaci (Calvo et al. 2015).
Biological variability is expected to occur among populations of a same species, especially for the most widely distributed ones. Some authors report the impact of intraspecific variability of Phytoseiidae species on biological control efficiency (i.e. Furtado et al. 2007; Ferrero et al. 2010; Domingos et al. 2012). Furthermore, abundant information can be found on intraspecific variability in regard to resistance to pesticides (Thistlewood et al. 1995; Sato et al. 2000; Hardman et al. 2000; Salman et al. 2015), diapause (Hoy 1984; Field and Hoy 1985; Morewood and Gilkeson 1991; van Houten et al. 1995 b), or tolerance to extreme temperatures (Voroshilov 1979) and low humidity (Walzer et al. 2007; Gómez-Moya et al. 2018).
Amblyseius tamatavensis Blommers is a phytoseiid mites that seems to fit type III-b (generalist predators living on glabrous leaves) group defined by McMurtry et al. (2013). Cavalcante et al. (2017) reported this species as a promising natural enemy of B. tabaci. Experimental releases of this predator on caged plants in a screenhouse caused the reduction of the density of B. tabaci on pepper plants by up to 60-80%. It can be rather easily produced in large numbers in laboratory conditions (Massaro et al., 2018) when fed with astigmatine mites, which could facilitate the eventual mass production for augmentative biological control. This species is reported from over 20 countries around the world (Africa, Asia, America and Ocenia). Given this wide world distribution, the origin of this species is difficult to assess. However, as far as we know, it seems that it has not been introduced to Brazil on purpose, where it is reported in 12 states from the northern to southeastern regions of the country (Demite et al. 2018). The hypotheses herein tested is that populations other than that studied by Cavalcante et al. (2017) could better perform for controlling B. tabaci.
Many biological characteristics may affect the performance of potential biological control agents. Their relative importance is expected to vary according to the prevailing abiotic and biotic environmental conditions of the region where the agent is expected to be used. Thus, finding ``the best'' predator population to control a pest species seems to be a complex task. However, for augmentative biological control under controlled conditions, as in greenhouses, the task is facilitated, by pre-setting limits to many of the variables (as the climatic factors, plant characteristics, etc). In this framework, the characteristics that have been considered in the selection of the most effective predadory phytoseiids are predation rates and oviposition potential (van Houten et al. 1995 a).
The present work aims at comparing populations of A. tamatavensis from different regions of Brazil in relation to their predation and oviposition potential when fed with eggs of B. tabaci biotype B. An evaluation of the relation between the average size of the mites of the distinct populations and each of those factors was conducted. This study is the first step of a long process for development of a population better adapted for the control of that insect.
Material and Methods
The populations considered
Populations
Substrate
1.Olho d’Água das Flores
(Alagoas)
Ipomoea pes-caprae
(Convolvulaceae)
2.Jataí (Goiás)
Several plants
3.Nova Crixás (Goiás)
Several plants
4.Bom Repouso (Minas
Gerais)
Several plants
5.Ituiutaba (Minas Gerais)
Several plants
6.Senador Amaral (Minas
Gerais)
Several plants
7.Campinas (São Paulo)
Psidium guajava (Myrtaceae)
8.Cananéia (São Paulo)
Persea americana (Lauraceae)
9.Mogi Guaçu (São Paulo)
Citrus sp. (Rutaceae)
10.Piracicaba, ESALQ (São
Paulo)
Gossypium sp. (Malvaceae)
11.Piracicaba, Areão (São
Paulo)
Rosa sp. (Rosaceae)
12.Saltinho (São Paulo)
Cocos nucifera (Arecaceae)
13.Santa Maria da Serra
(São Paulo)
Several plants
14. Laboratory
Capsicum annuum (Solanaceae)

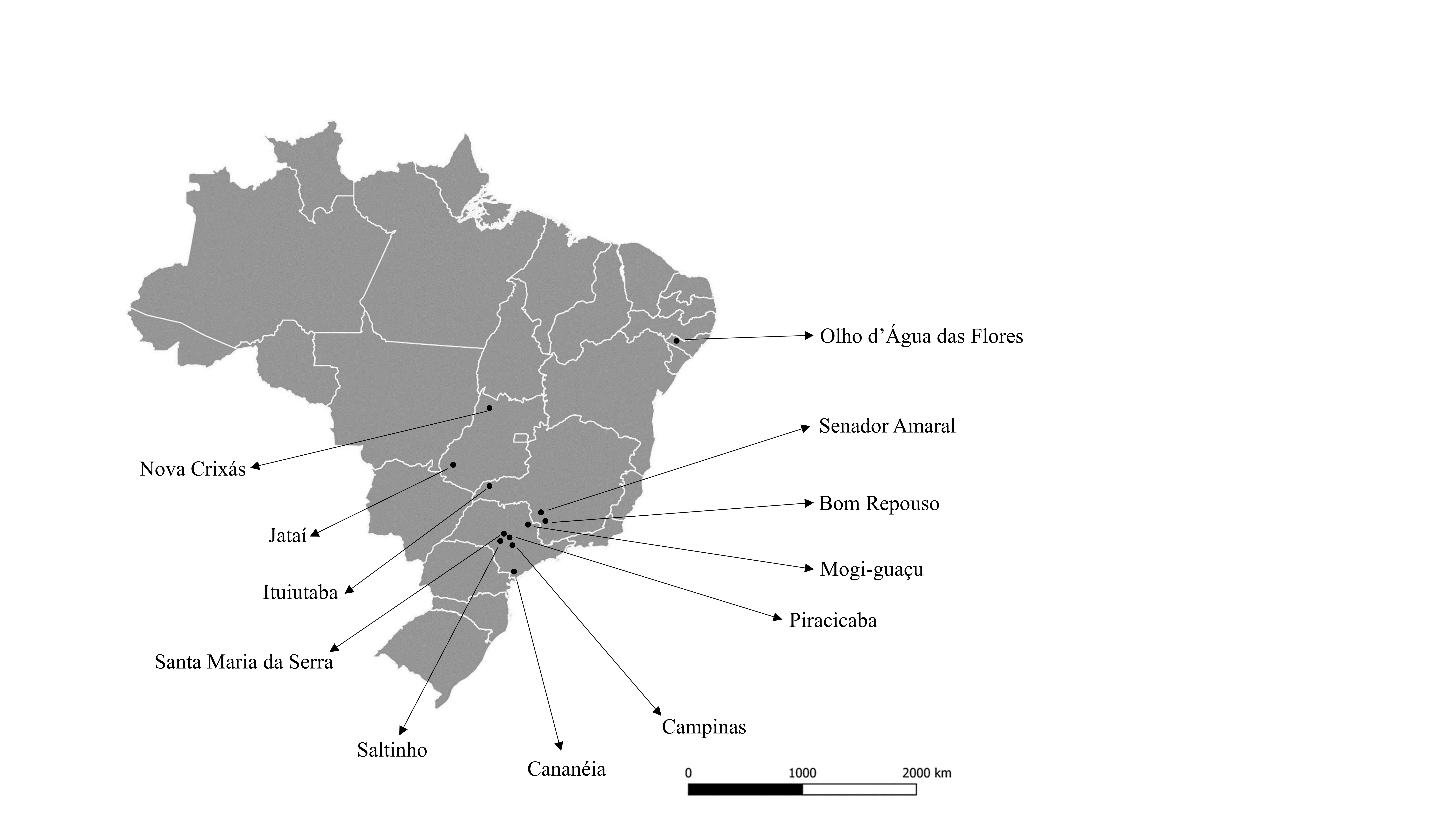
Aerial parts of glabrous leaf plants (preferred by A. tamatavensis) belonging to different species were collected from May 2015 to May 2016 in Brazilian regions with different eco-climatic characteristics. Mites identified as Amblyseius (with long Z5 setae and shiny dorsal shield) were collected using a stereomicroscope and then transferred to rearing units similar to those described by McMurtry and Scriven (1965). They were fed with pollen of Typha domingensis (Persoon) Steud. and different stages of Thyreophagus cracentiseta Barbosa, Oconnor and Moraes (Cavalcante et al. 2015; Massaro et al. 2016). About 15 days after the establishment of each colony, 5-10 adult females were mounted in Hoyer's medium to identify the species under a phase contrast microscope (Leica, DMLB). In total, 14 colonies were established (Table 1, Figure 1). One of these is of uncertain origin and had been maintained in the laboratory for about 5 years (population 14). The straight line distance between the most extreme sites of collection (Olho d'Água das Flores and Cananeia, sites 1 and 8 respectively) is 2,560 km.
Morphological characterization
To reconfirm the identity of the mites, 20 females of each population were mounted on slides using Hoyer's medium, at the beginning of the biological observations, described in the next section. According to Tixier (2012), for a precision of 15%, the measurement of 10 phytoseiid specimens would be required. Forty morphological parameters of adult females were measured. These were: dorsal shield length, dorsal shield width, length of the ventrianal shield, anterior and posterior width of the ventrianal shield, length of the dorsal idiosomal setae (j1, j3, j4, j5, J2, J5, z2, z4, z5, Z1, Z4, Z5, s4, S2, S4, S5, r3, R1); length of ventral idiosomal setae st1, st2, st3, st4, st5, JV1, JV2, JV4, JV5, ZV1, ZV2 and ZV3; distances between the bases of st1-st3, st2-st2, st5-st5; length of leg IV macrosetae; and length of calix of spermatheca. Dorsal shield setae terminology followed Lindquist and Evans (1965), as adapted by Rowell et al. (1978). Ventral shield terminology followed Chant and Yoshida-Shaul (1991). The measurements were made using a phase contrast microscope (Leica, DMLB) containing a graduated eyepiece. As the data satisfied the normality (Shapiro-Wilk test) and homoscedasticity (Levene test) assumptions, an analysis of the variance (ANOVA) followed by a mean comparison test (Tukey´s test, p < 0.05) was carried out to compare the means of each morphometric character of the 14 populations. Statistical analyzes were performed using R program (R Development Core Team 2013).

Predation and oviposition rates
These experiments were conducted in a rearing chamber at 30 ± 1 °C during the day and 23 ± 1 °C at night, 75 ± 10% RH and 12-h daily photoperiod. Forty mated females were randomly collected from each colony and individualized in experimental units. Each unit consisted of an acrylic mini Petri dish (1.5 x 3.0 cm) with the basis covered by a layer of a gypsum and activated charcoal mixture (9v: 1v) (Abbatiello 1965). In each unit, 15 eggs of B. tabaci (up to 24 h of age) were added, all of which (attacked by the predator during the experiment or not) replaced daily. To ensure that the eggs were intact when offered to the predators, they were not detached from the leaves; instead, the leaf substrate next to each egg was cut, and then egg and adjacent leaf substrate were transferred to the experimental unit. The experimental units were kept moist with daily addition of distilled water and sealed with plastic film (Magipac®) to prevent mites from escaping.
The study was conducted during 11 days, counting daily the number of eggs of B. tabaci killed and the number of eggs laid by the predator in each unit. The first day of evaluation was not considered in the analysis, due to the possible interference with the previous feeding. As oviposition and predation data did not satisfy the normality (Shapiro-Wilk test) and homoscedasticity (Levene test) assumptions, the non-parametric Kruskal-Wallis test was used to test the statistical difference between treatments ( P < 0.05), followed by Dunn´s test. Statistical analyzes were performed using R program (R Development Core Team 2013).
Finally, linear correlation analyses (Pearson's test) were performed to evaluate possible relationships between predation and oviposition, and mite size (estimated by dorsal shield length). All the analyses were performed using R program (R Development Core Team 2013).
Results
Morphological characterization
Significant differences between populations were observed for few morphological parameters (Table 2). The populations of Olhos d'Água das Flores and the laboratory population had larger dorsal and ventrianal shield dimensions as well as larger distance between the setae st4. The means of all characters of the different populations were very similar to each other. As differences among populations were minor, no attempt was made at evaluating the different levels of similarities among them.
Parameter1
Olho d’Água das Flores
Jataí
Nova-Crixás
Bom Repouso
Ituiutaba
Senador Amaral
Campinas
Cananéia
Mogi-Guaçu
Piracicaba (Areão)
Piracicaba (ESALQ)
Saltinho
Santa Maria da Serra
Lab.
Original descrip.
DSL
341±6 a
336±3 b
328±4 c
337±4 b
336±3 b
335 ± 7 b
333±3 b
338±5 b
337±5 b
333±2 c
336±4 b
324±6 c
325±2 c
342±5 a
340
DSW
231±4 a
226±4 b
223±3 bc
226±2 b
227±4 b
225±3 b
219±2 c
220±1 c
224±1 b
226±3 b
226±3 b
224±3 b
226 ± b
233±2 a
250
j1
32±0 a
31±0 a
32±0 a
32±0 a
32±0 a
32±1 a
30±3 a
31±1 a
33±0 a
32±1 a
31±2 a
31±1 a
31 ± 0 a
32±0 a
32
j3
51±1 b
54±2 a
50±4 b
52±3 b
54±1 a
55±2 a
52±1 b
51±5 b
55±2 a
52±1 b
55±1 a
52±1 b
51 ± 1 b
55±1 a
54
j4
5±0 a
5±0 a
5±0 a
5±0 a
5±0 a
6±0 a
5±0 a
5±1 a
5±0 a
5±0 a
5±0 a
5±0 a
5 ± 0 a
5±0 a
4
j5
4±0 a
3±0 a
4±0 a
4±0 a
4±0 a
4±0 a
3±0 a
3±0 a
4±0 a
4±0 a
4±0 a
4±0 a
4±0 a
4±0 a
3
J2
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
5±0 a
5±0 a
6±0 a
5±1 a
6±0 a
6±0 a
6±0 a
6 ± 0 a
6±0 a
5
J5
8±0 a
7±1 a
7±1 a
7±0 a
7±1 a
7±0 a
7±1 a
8±0 a
7±1 a
8±0 a
7±1 a
7±0 a
7 ± 1 a
7±1 a
6
z2
6±0 a
6±1 a
6±0 a
6±0 a
6±0 a
6±0 a
6±1 a
6±0 a
6±0 a
7±0 a
7±0 a
6±1 a
7 ± 0 a
6±0 a
6
z4
7±0 a
7±0 a
6±0 a
7±0 a
6±1 a
7±0 a
6±1 a
6±0 a
7±0 a
7±0 a
7±0 a
6±0 a
6 ± 0 a
7±0 a
6
z5
4±0 a
4±1 a
4±0 a
4±0 a
4±0 a
4±0 a
4±0 a
3±0 a
4±0 a
4±0 a
4±0 a
4±0 a
4 ± 0 a
4±0 a
3
Z1
6±0 a
6±1 a
7±0 a
6±0 a
6±0 a
6±0 a
6±1 a
6±0 a
6±0 a
7±0 a
6±0 a
7±0 a
6 ± 0 a
7±0 a
5
Z4
105±2 a
107±3 a
105±2 a
105±2 a
105±2 a
107±3 a
104±1 a
106±2 a
105±3 a
105±3 a
104±2 a
104±2 a
106 ±1 a
106±1 a
115
Z5
229±2 a
226±5 a
225±2 a
227±2 a
228±3 a
227±4 a
225±3 a
228±4 a
228±3 a
228±1 a
228±2 a
229±2 a
227±1a
229±2 a
250
s4
86±3 a
88±2 a
87±3 a
87±3 a
88±2 a
89±2 a
86±2 a
86±2 a
89±3 a
88±2 a
88±3 a
86±2 a
88±1 a
87±3 a
88
S2
7±0 a
6±1 a
7±0 a
6±0 a
7±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
7±0 a
7±0 a
6 ± 0 a
6±0 a
5
S4
6±1 a
6±0 a
6±0 a
6±0 a
7±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6 ± 0 a
6±0 a
6
S5
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6±0 a
6 ± 0 a
6±0 a
6
r3
14±1 a
15±1 a
13±1 a
14±1 a
14±1 a
14±1 a
13±2 a
14±1 a
15±1 a
15±1 a
15±1 a
14±1 a
14 ± 0 a
15±1 a
15
R1
7±0 a
7±1 a
7±0 a
7±0 a
7±0 a
7±0 a
7±0 a
7±1 a
7±1 a
7±1 a
7±0 a
7±0 a
7 ± 0 a
7±1 a
5
VASL
117±2 a
110±4 b
110±5 b
114±1 ab
110±2 b
108±4 b
111±3 b
112±3 b
113±2 b
113±2 b
108±3 b
109±3 b
111 ± 1 b
119±3 a
120
VASWant
99±3 a
97±4 a
94±3 b
95±3 b
95±2 b
93±2 b
91±4 b
98±4 a
97±3 a
95±2 b
97±2 a
91±3 b
92 ± 1 b
100±4 a
100
VASWpost
62±3 a
60±2 a
64±3 a
63±2 a
62±2 a
63±2 a
62±1 a
63±2 a
64±3 a
63±2 a
64±2 a
62±3 a
62± 2 a
62±4 a
-
St1-St2
32±0 a
32±0 a
32±1 a
33±1 a
33±0 a
32±0 a
32±1 a
32±0 a
34±0 a
32±1 a
32±1 a
32±1 a
33 ± 0 a
33±0 a
-
St2-St2
68±2 a
69±0 a
68±2 a
68±1 a
68±0 a
68±1 a
67±1 a
69±1 a
69±1 a
67±1 a
68±0 a
68±0 a
68 ± 1 a
70±0 a
-
St3-St3
76±2 a
77±2 a
76±2 a
78±3 a
76±2 a
78±1 a
75±1 a
77±2 a
77±2 a
76±2 a
76±2 a
76±1 a
77 ± 1 a
78±2 a
-
St1-St3
59±1 a
60±2 a
61±1 a
60±1 a
60±1 a
60±1 a
59±1 a
60±1 a
61±1 a
60±1 a
59±1 a
59±1 a
61 ± 0 a
62±1 a
-
St2-St3
27±1 a
27±1 a
28±1 a
27±1 a
28±1 a
28±0 a
27±1 a
28±1 a
28±1 a
28±1 a
28±1 a
27±0 a
28 ± 0 a
29±1 a
-
St4-St4
94±3 a
86±2 b
81±3 c
86±2 b
84±1 b
88±8 ab
81±3 c
84±2 b
84±3 b
86±3 b
86±2 b
81±3 c
85 ± 1 b
92±3 a
-
St5-St5
74±2 a
74±1 a
74±2 a
75±1 a
74±2 a
74±2 a
73±1 a
75±1 a
76±2 a
76±2 a
74±1 a
74±2 a
75 ± 2 a
76±2 a
-
Jv1
15±1 a
16±0 a
16±1 a
17±1 a
16±1 a
17±1 a
16±1 a
15±1 a
17±1 a
16±1 a
17±0 a
16±1 a
17 ± 0 a
17±1 a
-
Jv2
15±1 a
16±1 a
15±1 a
15±1 a
15±0 a
15±1 a
16±0 a
15±1 a
15±2 a
15±1 a
15±0 a
15±0 a
15 ± 0 a
16±1 a
-
Jv4
8±0 a
7±0 a
7±1 a
8±0 a
8±0 a
8±0 a
8±0 a
8±1 a
8±1 a
8±0 a
8±0 a
8±0 a
8 ± 0 a
7±0 a
-
Jv5
78±3
73±2
77±4
78±3
79±3
81±4
74±1
80±3
83±4
81±3
82±2
80±3
81 ± 2 a
82±3
84
Zv1
14±1 a
16±0 a
14±1 a
14±1 a
15±0 a
15±0 a
14±1 a
14±1 a
14±1 a
14±1 a
15±0 a
15±1 a
15 ± 0 a
14±0 a
-
Zv2
12±1 a
12±0 a
12±1 a
12±1 a
12±1 a
12±1 a
11±1 a
12±1 a
12±0 a
12±0 a
13±0 a
12±1 a
12 ± 0 a
13±1 a
-
Zv3
9±0 a
9±1 a
9±0 a
9±0 a
9±0 a
9±0 a
9±0 a
9±1 a
9±1 a
9±0 a
10±1 a
9±0 a
9 ± 0 a
10±0 a
-
SgeIV
98±2 a
97±3 a
97±3 a
98±3 a
97±2 a
99±2 a
98±2 a
100±2 a
100±4 a
100±1 a
98±2 a
98±3 a
99 ± 2 a
100±3 a
120
StiIV
66±4 a
65±3 a
66±2 a
66±3 a
67±4 a
64±2 a
65±1 a
66±4 a
65±4 a
64±3 a
66±3 a
64±3 a
65 ± 2 a
64±4 a
75
StIV
61±4 a
63±2 a
62±3 a
63±1 a
64±2 a
63±2 a
62±3 a
62±2 a
64±1 a
64±3 a
64±2 a
62±2 a
63 ± 0 a
65±2 a
73
Calyx
20±1 a
19±1 a
19±1 a
20±1 a
19±1 a
18±2 a
19±1 a
20±1 a
19±1 a
19±1 a
19±0 a
20±1 a
19±1 a
21±1 a
16
1 DSL: dorsal shield length
along the midline; DSW: dorsal shield width at the widest level of the
podonotal region; VASL: length of the ventrianal shield along the midline;
VSWant and VSWpost: anterior (at level of ZV2) and posterior (at level of anal opening) width of the
‘ventrianal shield; SgeIV, StiIV and StIV: lengths of macrosetae of
genu, tibia and tarsus of leg IV; calyx: length of calyx of spermatheca.
In a same
line, means followed by the same letters are not statistically different
(Tukey´s test; P
> 0.05); (-) Not given.
Predation and oviposition
Significant differences between populations were observed for both mean daily predation and oviposition rates (ratios maximum/minimum = 1.58 and 1.71, respectively) (Table 3). Daily predation was the highest for the population of Olho d'Água das Flores (7.9 eggs/ female), followed by the laboratory population (7.0 eggs/ female). The daily oviposition of the Olho d'Água das Flores (1.2 eggs/ female) population was significantly higher than for the other populations, but not for that obtained for the laboratory population (1.1 eggs/ female).
A positive and significant correlation was observed between the oviposition rates and the predation rates (y = 0.18x - 0.23, where y = number of eggs of A. tamatavensis produced daily, x = number of eggs of B. tabaci consumed daily by predator; P = 0.0004, R2 = 0.663). No significant correlations between dorsal shield length and predation or oviposition rates were observed. However, populations with the highest predation and oviposition rates (Table 3) were also those with the highest dorsal shield length (Table 2).
Collection sites
Predation
Oviposition
Olho d’água das
Flores
7.9 ± 0.7 a
1.2 ± 0.2 a
Laboratório
7.0 ± 0.4 b
1.1 ± 0.3 ab
Jataí
6.5 ± 0.4 c
0.8 ± 0.1 c
Saltinho
6.5 ± 0.3 c
1.0 ± 0.1 b
Mogi-Guaçu
6.5 ± 0.3 c
0.8 ± 0.0 c
Piracicaba - Fazenda
Areão
6.5 ± 0.2 c
1.0 ± 0.2 b
Senador Amaral
6.5 ± 0.1 c
0.9 ± 0.3 bc
Bom Repouso
6.3 ± 0.3 cd
0.9 ± 0.2 bc
Ituiutaba
6.3 ± 0.2 cd
0.8 ± 0.1 c
Campinas
6.3 ± 0.2 cd
1.0 ± 0.1 b
Cananéia
6.3 ± 0.1 cd
0.9 ± 0.2 bc
Santa Maria da
Serra
6.1 ± 0.4 d
0.8 ± 0.1 c
Piracicaba - ESALQ
6.0 ± 0.5 d
0.9 ± 0.1 bc
Nova Crixás
5.0 ± 0.3 e
0.7 ± 0.2 c
For each column,
rates followed by the same letter are not statistically different
(Kruskal-Wallis ANOVA; Dunn´s test).
Discussion
Despite some small morphological differences between populations, the populations were considered belong to the same species. The measurements of the 14 populations are compatible to those reported in the original description (Blommers 1974) of A. tamatavensis. The differences herein observed are compatible with the expected intraspecific variation of a continuous structure, suggested by Tixier (2012) to be ca. 20% around the average for the phytoseiids. Significant differences in size of some morphological structures have been reported in the literature for distinct populations of a same phytoseiid species, e.g. Sourassou et al. (2011) for Neoseiulus paspalivorus De Leon, Sourassou et al. (2012) for Neoseiulus baraki Athias-Henriot, Lopes et al. (2018) for Euseius concordis (Chant) and Tixier et al. (2003) for Kampimodromus aberrans (Oudemans). In a short-term laboratory study, Lopes et al. (2018) observed marked morphological differences for colonies of E. concordis maintained under different diet or temperature conditions. Differences in temperature, diet, host plant or other factors could explain the larger size observed for the A. tamatavensis population of the Olho d'Água das Flores, the most isolated population among those evaluated in this study. The same factors could also explain the differences between morphological features reported in the original description of A. tamatavensis and those of the populations considered herein.
The values obtained in the present study for predation and oviposition are similar to those reported by Cavalcante et al. (2015) in a study conducted with A. tamatavensis under approximately the same environmental conditions. In that study, daily mean consumption was 8.0 eggs and oviposition was 1.4 eggs each day.
The significant relationship between predation and oviposition seems logical. It is indeed expected that individuals that fed more would also be more prolific. It could also be expected that larger mites could prey more and probably oviposit more (Sabelis 1981). The absence of significant correlation between size and rate of predation, or between size and rate of oviposition (as observed in the present study) seems to contradict that assumption. Yet, it is noteworthy to point out that the only populations that stood out in relation to size (as indicated by dimensions of dorsal shield) in the study were from Olho d'Água das Flores and the laboratory population, and those had the highest predation and oviposition rates; the size of the other populations were not statistically different. However, the difference in size between those two and other populations was small, and size might not be a reliable trait to assess rates of predation or oviposition. However, if size is really determinant, populations of the type locality of the species (Madagascar, with dorsal shield 340 µm long) or from Africa (with dorsal shield on the average 354 µm long; Zannou et al. 2007) would be expected to have similar or better performance than the best Brazilian populations. From the available re-descriptions of this species (cited by Demite et al. 2018), specimens of the A. tamatavensis from other countries (for which measurements were provided) are not distinctly larger than specimens collected in this study.
Regardless of the cause of the highest predation or oviposition rates and focusing on the search for the most promising A. tamatavensis population for the control of B. tabaci, it seems that priority should be given to the population collected in Olho d'Água das Flores or the laboratory populations. It seems that each of them could have sufficient variability for the selection of a new, improved population (with higher potential for predation (mainly) or oviposition) when fed with B. tabaci. In this study, daily ranges of predation and oviposition rates were 5.6–12.0 and 1.1–1.5 for Olho d'Água das Flores population, and 6.2–8.2 and 0.8–1.3 for laboratory population.
Genetic improvement implies the change of genetic characteristics of a species by humans to suit their needs (Hoy 1985). Thus, this process could involve directed and purposive genetic changes to improve the efficacy of natural enemies for pest control (Hoy 1990), given that artificial selection can accelerate events that rarely occur in the field, producing results that would hardly occur naturally.
Selection processes made by humans over time have allowed the production of plant varieties and animal breeds much more adequate to meet consumption needs (Garland and Rose 2009). Studies of this type have been little explored by researchers in the use of predatory mites (Lommen et al. 2017), and the results of this study indicate the great potential of applying to these organisms similar selection processes adopted for many years for plants and animals.
Acknowledgements
To Geovanny Barroso, for collecting the specimens from Olho d'Água das Flores. To Michele Ennes, for collecting the specimens from Mogi-guaçu.
References
Abbatiello M.J. 1965. A culture chamber for rearing soil mites. Turtox News, 43 7: 162–164.
Blommers L. 1974. Species of the genus Amblyseius Berlese, 1914, from Tamatave, east Madagascar (Acarina: Phytoseiidae). Bull. Zool. Mus. Univ. Amst. 3:143–155.
Brown J.K., Frohlich D.R., Rosell R.C. 1995. The sweetpotato or silverleaf whiteflies: biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol., 40: 511–534. doi:10.1146/annurev.en.40.010195.002455 ![]()
De Barro, P.J., Liu, S.S., Boykin, L.M., Dinsdale, A.B. 2011. Bemisia tabaci: a statement of species status. Annu. Rev. Entomol., 56: 1-19. doi:10.1146/annurev-ento-112408-085504 ![]()
Cavalcante A.C.C., Mandro M.E.A., Paes E.R., Moraes G. J. 2017. Amblyseius tamatavensis Blommers (Acari: Phytoseiidae) a candidate for biological controlo of Bemisia tabaci (Gennadius) biotype B (Hemiptera: Aleurodidae) in Brazil. Intern. J. Acarol., 43: 10-15. doi:10.1080/01647954.2016.1225816 ![]()
Cavalcante A.C.C., Santos V.L.V., Rossi L.C., Moraes G.J. 2015. Potential of five brazilian populations of Phytoseiidae (Acari) for the biological controlo of Bemisia tabaci (Insecta: Hemiptera). J. Econ. Entomol. 108: 29-33. doi:10.1093/jee/tou003 ![]()
Calvo F.J., Knapp M., van Houten Y.M., Hoogerbrugge H., Belda J. E. 2015. Amblyseius swirskii: What made this predatory mite such a successful biocontrol agent? Exp. Appl. Acarol. 65: 419-433. doi:10.1007/s10493-014-9873-0 ![]()
Chant D. A., Yoshida-Shaul E. 1991. Adult ventral setal patterns in the family Phytoseiidae (Acari: Gamasina). Int. J. Acarol. 17: 187-199. doi:10.1080/01647959108683906 ![]()
Demite P.R., Moraes G.J. de, McMurtry J.A., Denmark H.A., Castilho R. C. 2018. Phytoseiidae Database. Available from: www.lea.esalq.usp.br/phytoseiidae (accessed 22/XII/2018).
Domingos C.A., Oliveira L.O., Morais E.G.F., Navia D., Moraes G. J., Gondim Jr. M.G.C. 2012. Comparison of two populations of the pantropical predator Amblyseius largoensis (Acari: Phytoseiidae) for biological control of Raoiella indica (Acari: Tenuipalpidae). Exp. Appl. Acarol. 60: 83-93. doi:10.1007/s10493-012-9625-y ![]()
Ferrero M., Gigot C., Tixier M.S., van Houten Y.M., Kreiter S. 2010 Egg hatching response to a range of air humidities for six species of predatory mites. Entomol. Exp. Appl., 135 (3), 237-244. doi:10.1111/j.1570-7458.2010.00992.x ![]()
Field R.P., Hoy M.A. 1985. Diapause behavior of genetically improved strains of the spider mite predator Metaseiulus occidentalis (Acarina: Phytoseiidae). Entomol. Exp. Appl., 38: 113-120. doi:10.1111/j.1570-7458.1985.tb03507.x ![]()
Furtado I.P., Moraes G.J., Kreiter S., Tixier M.-S., Knapp M. 2007. Potential of a brazilian population of the predatory mite Phytoseiulus longipes as a biological control agent of Tetranychus evansi (Acari: Phytoseiidae, Tetranychidae). Biol. Control, 42: 139-147. doi:10.1016/j.biocontrol.2007.04.016 ![]()
Garland T, Jr., Rose MR, 2009. Experimental Evolution: Concepts, Methods, and Applications of Selection Experiments. University of California Press, Berkeley, California.
Gómez-Moya C.A., Gondim Jr. M.G.C., Moraes G.J. de, Morais E.G.F. de. 2018. Effect of relative humidity on the biology of the predatory mite Amblyseius largoensis (Acari: Phytoseiidae). Int. J. Acarol., 44(7): 279-287. doi:10.1080/01647954.2018.1530300 ![]()
Hardman J.M., Moreau D.L., Snyder M., Gaul S.O., Bent E.D. 2000. Performance of a pyrethroid-resistant strain of the predator mite Typhlodromus pyri (Acari: Phytoseiidae) under different insecticide regimes. J. Econ. Entomol. 93: 590-604. doi:10.1603/0022-0493-93.3.590 ![]()
Hogenhout S.A., Ammar E.D., Whitfield A.E., Redinbaugh M.G. 2008. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 46: 327-359. doi:10.1146/annurev.phyto.022508.092135 ![]()
Hoy M.A. 1984. Genetic improvement of a biological control agent: Multiple pesticide resistances and non-diapause in Metaseiulus occidentalis (Nesbitt) (Phytoseiidae). In: D.A. Griffiths (ed.), Acarology, 2: 673-679.
Hoy M.A. 1985. Recent advances in genetics and genetic improvement of the Phytoseiidae. Annu. Rev. Entomol. 30: 345–370. doi:10.1146/annurev.en.30.010185.002021 ![]()
Hoy M.A. 1990. Genetic improvement of arthropod natural enemies: becoming a conventional tactic? New Directions in Biological Control: Alternatives for Suppressing Agricultural Pests and Diseases (ed. by RR Baker & PE Dunn), 405–417.
Lindquist E., Evans G. 1965. Taxonomic Concepts in the Ascidae, with a Modified Setal Nomenclature for the Idiosoma of the Gamasina (Acarina: Mesostigmata). Mem. Entomol. Soc. Can., 97(S47), 5-66. doi:10.4039/entm9747fv ![]()
Lommen S.T.E., Jong P.W., Pannebakker B.A. It is time to bridge the gap between exploring and exploiting: prospects for utilizing intraspecific genetic variation to optimize arthropods for augmentative pest control – a review. Entomol. Exp. Appl. 162: 108-123. doi:10.1111/eea.12510 ![]()
Lopes P.C., Kanno R.H., Sourassou N.F., Moraes G.J. 2018. Effect of temperature and diet on the morphology of Euseius concordis (Acari: Phytoseiidae). Syst. Appl. Acarol., 23: 1322-1332. doi:10.11158/saa.23.7.9 ![]()
Lourenção A.L., Sakate R.K., Valle G.E. 2015. Bemisia tabaci biótipo B. In: Vilella, E.; Zucchi, R.A. (Ed.). Pragas introduzidas no Brasil: insetos e ácaros. Piracicaba: FEALQ, 2015.
Massaro M., Martin J.P.I., Moraes G.J. 2016. Factitious food for mass production of predaceous phytoseiid mites (Acari: Phytoseiidae) commonly found in Brazil. Exp. Appl. Acarol., 70: 411. doi:10.1007/s10493-016-0087-5 ![]()
Massaro M., Montrazi M., Melo J.W.S., Moraes G.J. 2018. Production of Amblyseius tamatavensis with Thyreophagus crasentiseta (Acari: Phytoseiidae, Acaridae). International Journal of Pest Management (in press).
McMurtry J.A., Moraes G.J., Famah Sourassou N. 2013. Revision of the lifestyle of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol, 8: 297-320. doi:10.11158/saa.18.4.1 ![]()
McMurtry JA, Scriven GT. 1965. Insectary production of phytoseiid mites. J. Econ. Entomol. 58: 282–284. doi:10.1093/jee/58.2.282 ![]()
Morewood W.D., Gilkeson L.A. 1991. Diapause induction in the thrips predator Amblyseius cucumeris (Acarina: Phytoseiidae) under greenhouse conditions. Entomophaga, 36:253-263. doi:10.1007/BF02374561 ![]()
R Development Core Team. 2013. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.r-project.org/ ![]()
Rowell H.J., Chant D.A., Hansell R.I.C. 1978. The determination of setal homologies and setal patterns on the dorsal shield on the family Phytoseiidae (Acari: Mesostigmata). Can. Entomol., 110: 859-876. doi:10.4039/Ent110859-8 ![]()
Sabelis M.W. 1981. Biological control of two spotted spider mites using phytoseiid predators. Part I: Modelling the predator-prey interaction at the individual level. Wageningen: Centre for Agricultural Publishing and Documentation.
Salman S.Y., Aydinli F., Ay R. 2015. Selection for resistance: Cross-resistance, inheritance, synergists and biochemical mechanisms of resistance to acequinocyl in Phytoseiulus persimilis A.H. (Acari: Phytoseiidae). Crop Prot., 67:109-115. doi:10.1016/j.cropro.2014.10.001 ![]()
Sato M.E., Miyata T., Kawai A., Nakano O. 2000. Selection for resistance and susceptibility to methidathion and cross resistance in Amblyseius womersleyi Schicha (Acari: Phytoseiidae). Appl. Entomol. Zool., 53: 393-399. doi:10.1303/aez.2000.393 ![]()
Sourassou N. F., Hanna R., Zannou I., de Moraes G., Negloh K., Sabelis M.W. 2011. Morphological variation and reproductive incompatibility of three coconut-mite-associated populations of predatory mites identified as Neoseiulus paspalivorus (Acari: Phytoseiidae). Exp. Appl. Acarol., 53: 323-338. doi:10.1007/s10493-010-9413-5 ![]()
Sourassou F. N, Hanna R., Zannou I., Breeuwer J.A.J., de Moraes G.J., Sabelis M.W. 2012. Morphological, molecular and cross-breeding analysis of geographic populations of coconut-mite-associated predatory mites identified as Neoseiulus baraki: evidence for cryptic species? Exp Appl Acarol 57:15–36. doi:10.1007/s10493-012-9534-0 ![]()
Thistlewood H.M.A., Pree D.J., Crawford L.A. 1995. Selection and genetic analysis of permethrin resistance in Amblyseius fallacis (Garman) (Acari: Phytoseiidae) from Ontario apple orchards. Exp. Appl. Acarol., 19:707-721. doi:10.1007/BF00052082 ![]()
Tixier M.-S., Kreiter S., Cheval B., Auger P. 2003. Morphometric variation between populations of Kampimodromus aberrans (Oudemans) (Acari: Phytoseiidae): implication for the taxonomy of the genus. Invertebr. Syst., 17(2), 349–358. doi:10.1071/IS02004 ![]()
van Houten Y.M., van Rijn P.C.J., Tanigoshi L.K., van Stratum P., Bruin J. 1995a. Preselection of predatory mites to improve year-round biological control of western flower thrips in greenhouse crops. Entomol. Exp. Appl., 74: 225–234. doi:10.1111/j.1570-7458.1995.tb01895.x ![]()
Tixier M.-S. 2012. Statistical approaches to assess intraspecific variations of morphological continuous characters: the case study of the family Phytoseiidae (Acari: Mesostigmata). Cladistics, 28: 489–502. doi:10.1111/j.1096-0031.2012.00394.x ![]()
van Houten Y.M., van Startum P., Bruin J., Veerman A. 1995b. Selection for non-diapause in Amblyseius cucumeris and Amblyseius barkeri and exploration of the effectiveness of selected strains for thrips control. Entomol. Exp. Appl., 77: 289-295. doi:10.1111/j.1570-7458.1995.tb02326.x ![]()
Voroshilov, H.V. 1979. Heat-resistant lines of the mite Phytoseiulus persimilis A-H. Genetika, 15: 70-76.
Walzer A., Castagnoli M., Simoni S., Liguori M., Palevsky E., Schausberger P. 2007. Intraspecific variation in humidity susceptibility of the predatory mite Neoseiulus californicus: survival, development and reproduction. Biol. Control. 41:42–52. doi:10.1016/j.biocontrol.2006.11.012 ![]()
Zannou I.D., Moraes G.J. de, Ueckermann E.A., Oliveira A.R., Yaninek J.S., Hanna R. 2007. Phytoseiid mites of the subtribe Amblyseiina (Acari: Phytoseiidae: Amblyseiini) from sub-Saharan Africa. Zootaxa, 1550, 1–47. doi:10.11646/zootaxa.1550.1.1 ![]()



2018-12-27
Date accepted:
2019-02-01
Date published:
2019-02-12
Edited by:
Tixier, Marie-Stéphane

This work is licensed under a Creative Commons Attribution 4.0 International License
2019 Massaro, Marcela and Moraes, Gilberto J. de
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)