



Morphological observations of fluorescence in different tick species (Family: Ixodidae)
Shade, Dakota A.  1
; Oller, Anna R.
2
and Wilson, Stephen W.3
1
; Oller, Anna R.
2
and Wilson, Stephen W.3
1✉ Department of Biological and Clinical Sciences, University of Central Missouri, WCM 306, Warrensburg, MO 64093, USA & Independent Researcher, Kansas City, MO, USA.
2Department of Biological and Clinical Sciences, University of Central Missouri, WCM 306, Warrensburg, MO 64093, USA.
3Department of Biological and Clinical Sciences, University of Central Missouri, WCM 306, Warrensburg, MO 64093, USA.
2018 - Volume: 58 Issue: 4 pages: 989-994
https://doi.org/10.24349/acarologia/20184305Corrected version 16 April 2024
Keywords
Abstract
Ticks are important ectoparasites to the medical and veterinary world due to their capability of transmitting pathogenic organisms (Ghosh et al., 2007). Fluorescence is a phenomenon that can be observed in numerous biological systems and has been previously found in ticks (Dillinger and Kesel, 2002). It occurs due to the excitation of an electron into a higher energy orbital. The return of the electron to the ground state results in the emission of a photon of lower energy (Lakowicz, 2006). The phenomenon can be due to different compounds, microscopic structures and a mixture of both. Little knowledge exists as to the overall function, distribution, and evolution of fluorescence in biological systems (Lagorio et al., 2015). Fluorescence has been found in multiple arthropods including crustaceans (Giurginca et al., 2015), insects (Phillips, 1959; Vukusic and Hooper, 2005; Jiang et al., 2012), and arachnids (Volschenk, 2005; Gaffin et al., 2012; Andrews et al., 2007). Andrews et al. (2007) showed that every spider they examined had several fluorescent structures. As the presence of external fluorescence varied among spiders, they concluded it must have an adaptive role in some species (Andrews et al., 2007). Jumping spiders that preyed on orb weaving spiders were shown to approach UV reflective webs more often than non-UV reflective webs, suggesting UV may possibly play roles in communication and interactions between arthropods (Li and Lim, 2005). Dillinger and Kesel (2002) were the first to observe fluorescence in Ixodes ricinus Linnaeus (Acari: Ixodidae) ticks. They described fluorescence in the alloscutum, leg joints, between the idiosoma and capitulum, setae, and various structures of the mouthparts (Dillinger and Kesel, 2002). The presence and role of fluorescence in other ticks is unknown. The scope of this study is thus to perform morphological observations of fluorescence in different tick species and discuss its potential adaptive significance.
Six individuals of each of six species of unfed adults (three males and three females) were purchased from the Oklahoma State University tick lab: Amblyomma americanum (Linnaeus), A. maculatum Koch, Dermacentor variabilis (Say), D. andersoni (Stiles), Ixodes scapularis Say, and Rhipicephalus sanguineus Latreille (Acari: Ixodidae). Ticks were sacrificed and stored in 70% ethanol 24 hours prior to imaging. Images were captured using an Olympus SZX7 stereo microscope (Olympus Corporation, Tokyo, Japan) with an attached Zeiss axiocam 105 camera (Carl Zeiss, Oberkochem, Germany). The excitation sources were a NightSea SFA-UV (360-380 nm) with a 412 nm longpass filter and NightSea SFA-Royal Blue (RB) (440-460 nm) with a 500 nm longpass filter (NightSea, Lexington Massachusetts). ZEN imaging software was used for imaging and ImageJ for image enhancements.
Under UV excitation (360-380 nm), the fluorescence appeared blue in structures. Under RB excitation (440-460 nm), the fluorescence was green/yellow in color. All species observed in this study showed similar fluorescence with few exceptions (Figures 1-3).


Table 1 shows the positive and negative results for fluorescence in both sexes. The following structures showed fluorescence in all species and sexes: anal groove, genital pore, spiricals, setae, ventral idiosoma, hypostome, palpal articles, cheliceral sheath, leg segments, arthrodial membrane, and the apotele. All species with eyes and ornamentation present showed fluorescence in those structures (Figure 1). Fovae and lamellae were intentionally observed only in A. americanum females due to limited access to a fluorescent microscope, but these structures are presumed to be fluorescent in females of the other species. Portions of the basis capitulum are not fluorescent in I. scapularis contrary to the other species.

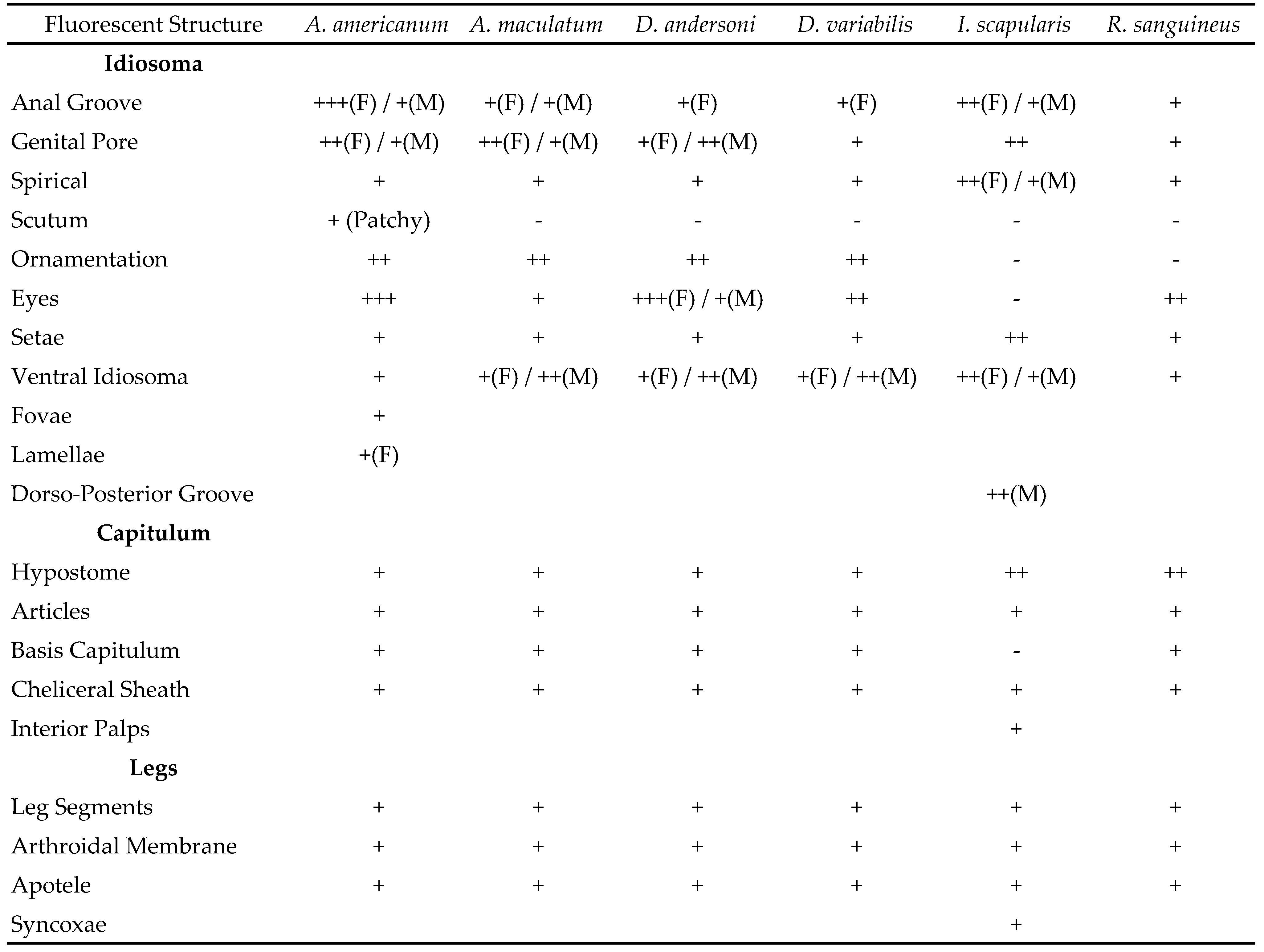
Fluorescence appears to be widespread in the six species studied and is likely the case for all species of ixodid ticks. Much of the observed fluorescence appears to be related to flexible cuticle and may be a by-product of the compounds used to make this layer. Dillinger and Kesel (2002) hypothesized fluorescence found in I. ricinus was due to resilin, but the study did not test for the protein. Andersen and Roepstorff (2005) concluded the alloscutum of I. ricinus contains 8% chitin, meaning most proteins had a Rebers-Riddiford consensus and are bound to other proteins by cross-links with other compounds. The components of resilin, dityrosine and trityrosine, were found to be present, however, the amount was much less compared to insect resilin. 3-monochlorotyrosine and 3,5-dichlorotyrosine were isolated in the distended alloscutum, but were concluded to play a role in cross-linking or were products of oxidative stress (Andersen and Roepstorff, 2005). The nature of these compounds and their role in fluorescence is unknown.
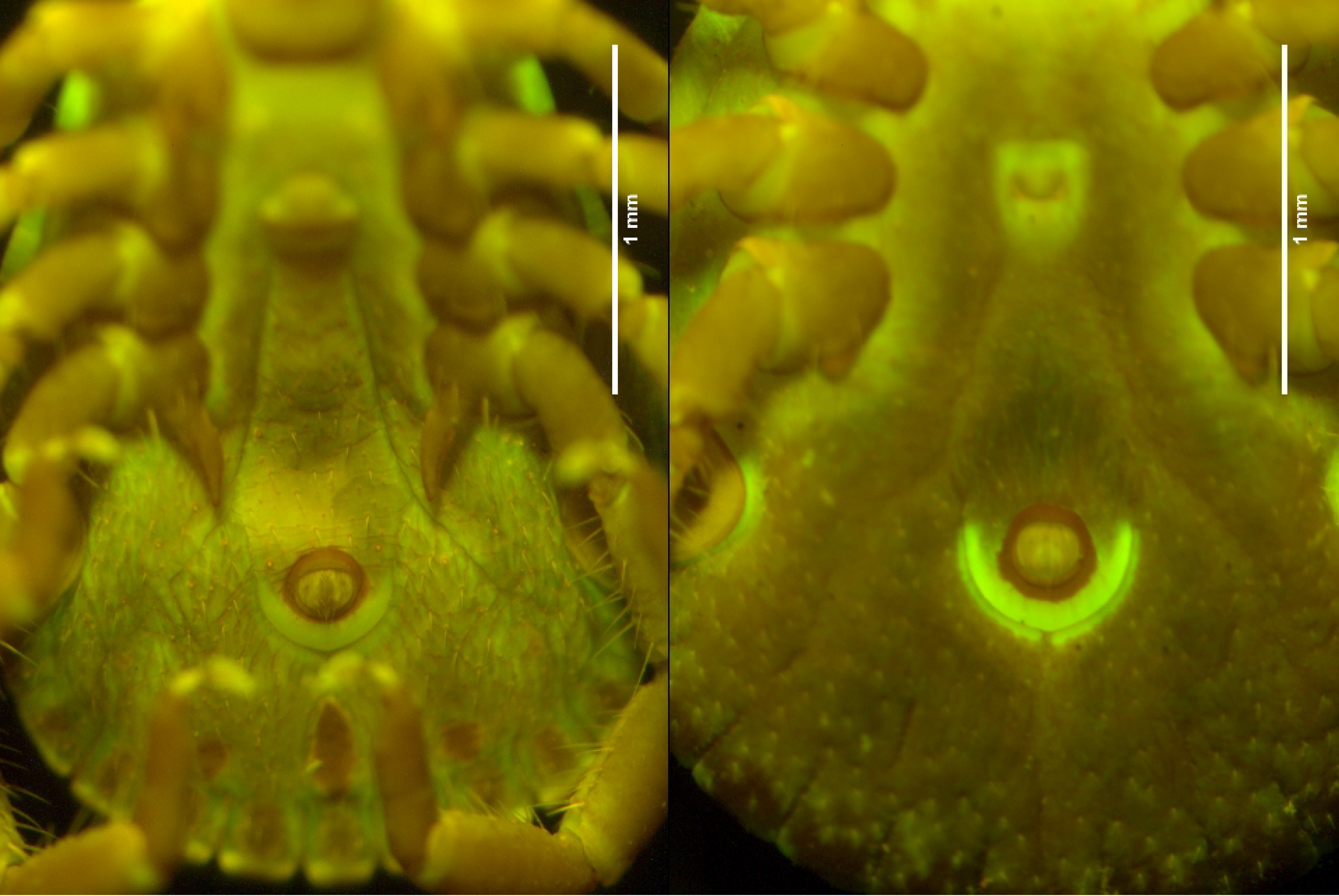
The anal groove of females in almost all species observed appears to have brighter fluorescence compared to their male counterparts (Figure 2). The fluorescence of the female anal groove could be due to pre-emptive preparation for expansion while feeding. As the tick feeds, the alloscutum extends to a considerable size, but the anal groove does not expand in size during this process. In contrast, ixodid males feed very little in the adult stage and do not increase significantly in size, suggesting a link between feeding morphology and fluorescence.

I found that I. scapularis has fluorescent syncoxae on the underside of the coxa of legs 1 and 2 (Figure 3). The purpose of these structures are unknown, but they do not expand during engorgement. Fluorescence in such structures may thus be by-products of shared compounds used in flexible cuticle e.g., leg segments. In other observed fluorescent structures that do not appear to be involved in expansion, the molecular make-up may be different. For example, tick ornamentation does not have a structural purpose, but also shows fluorescence that differs in comparison to other fluorescent structures. Interestingly, most ornate tick species have eyes. Ornamentation fluorescence may therefore serve one or more functions in ticks. Indeed, Kaltenrieder et al. (1989) showed that the spectral acuity of tick eyes has features of both spider eyes and insect ocelli. The use of visual cues may aid in aggregation and mate choice, ensuring viable copulations occur on or off the host. Ornamentation could also potentially serve a function in disruptive/cryptic coloration. Crypsis is a trait that reduces an animal’s risk of being detected (Stevens and Merilaita, 2009). Ticks may utilize fluorescent ornamentation as a form of coloration to break up their overall shape. Indeed, the ornamentation fluorescence contrasts well with the dark non-fluorescent portions of the tick and this contrast can be deceptive to a visual predator. Crypsis/disruptive coloration could potentially benefit those tick species that quest in more open habitat compared to those that quest in leaf litter and/or dense wooded areas. All the species examined here tend to search for their hosts in open habitats. Intraspecific and interspecific behavioral studies will now be needed to ascertain whether this is a viable avenue to explore further.


This study showed that ticks exhibit wide range fluorescence like other related arthropods. New fluorescent structures have been identified and the number is likely to increase as more tick species are viewed under UV light. Fluorescence may play a role in intraspecific communication in tick species or act in crypsis to avoid detection by potential predators. Chemical and behavior studies on ticks is now called for to better understand the true significance of fluorescence in these arthropods. The future potential for exploring fluorescence in ticks appears to be very fruitful, as it may uncover more unknown biological characteristics of ticks.
I wish to thank my daughter Molly for all her help in the lab. “Don’t touch that!”
Andersen, S. O., Roepstorff, P., 2005. The extensible alloscutal cuticle of the tick, Ixodes ricinus. Insect Biochemistry and Molecular Biology. 35(10): 1181-1188. doi:10.1016/j.ibmb.2005.05.009 ![]()
Andrews, K., Reed, S. M., Masta, S. E., 2007. Spiders fluoresce variably across many taxa. Biology Letters., 3(3): 265-267. doi:10.1098/rsbl.2007.0016 ![]()
Dillinger, S. C. G., Kesel, A. B., 2002. Changes in the structure of the cuticle of Ixodes ricinus L. 1758 (Acari, Ixodidae) during feeding. Arthropod Structure and Development. 31(2): 95-101. doi:10.1016/S1467-8039(02)00042-7 ![]()
Gaffin, D. D., Bumm, L. A., Taylor, M. S., Popokina, N. V., Mann, S., 2012. Scorpion fluorescence and reaction to light. Animal Behaviour. 83(2): 429-436. doi:10.1016/j.anbehav.2011.11.014 ![]()
Ghosh, S., Azhahianambi, P., Yadav, M. P., 2007. Upcoming and future strategies of tick Control: a review. Journal of Vector Borne Diseases. 44(2): 79-89.
Giurginca, A., Šustr, V., Tajovský, K., Giurginca, M., Matei, I., 2015. Spectroscopic parameters of the cuticle and ethanol extracts of the fluorescent cave isopod Mesoniscus graniger (Isopoda, Oniscidea). ZooKeys. (515): doi:10.3897/zookeys.515.9395 ![]()
Jiang, L., Dong, B., Liu, X., Liu, F., Zi, J., 2012. Structural origin of sexual dichromatic coloration and luster in the beetle Goliathus cacicus. Chinese Science Bulletin. 57(24): 3211-3217. doi:10.1007/s11434-012-5343-4 ![]()
Kaltenrieder, M., Labhart, T., Hess, E., 1989. Spectral sensitivity, absolute threshold, and visual field of two tick species, Hyalomma dromedarii and Amblyomma variegatum. Journal of Comparative Physiology A. 165(2): 155-164. doi:10.1007/BF00619190 ![]()
Lagorio, M. G., Cordon, G. B., Iriel, A., 2015. Reviewing the relevance of fluorescence in Biological systems. Photochemical and Photobiological Sciences. 14(9): 1538-1559. doi:10.1039/C5PP00122F ![]()
Lakowicz, J. R. 2006. Principles of fluorescence spectroscopy, 3rd Edn. Springer. New York, NY. pp. 63-69. doi:10.1007/978-0-387-46312-4_3 ![]()
Li, D., Lim, M. L., 2005 Ultraviolet cues affect the foraging behaviour of jumping spiders. Animal Behaviour. 70(4): 771-776. doi:10.1016/j.anbehav.2004.12.021 ![]()
Phillips, L. S., 1959. Fluorescence in the colors of certain Lepidoptera observed under ultraviolet light. Journal of the Lepidopterists' Society. 13(2): 73-77.
Stevens, M., Merilaita, S., 2009. Animal camouflage: current issues and new perspectives.Philosophical Transactions of the Royal Society B: Biological Sciences. 364(1516): 423-427. doi:10.1098/rstb.2008.0217 ![]()
Volschenk, E. S., 2005. A new technique for examining surface morphosculpture of scorpions. Journal of Arachnology. 33(3): 820-825. doi:10.1636/S03-047.1 ![]()
Vukusic, P., Hooper, I., 2005. Directionally controlled fluorescence emission in butterflies. Science. 310(5751): 1151. doi:10.1126/science.1116612 ![]()
Dakota Shade
a. Department of Biological and Clinical Sciences, University of Central Missouri, WCM 306, Warrensburg, MO 64093, USA.
b. Independent Researcher, Kansas City, MO, USA
The online, pdf and HAL repository versions have been changed on April 16, 2024 and includes the corrections.



2018-01-22
Date accepted:
2018-11-21
Date published:
2018-11-30
Edited by:
McCoy, Karen

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Shade, Dakota A.; Oller, Anna R. and Wilson, Stephen W.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)