



Two new feather mite species of the genus Montesauria Oudemans (Analgoidea: Proctophyllodidae) from thrushes (Passeriformes: Turdidae) in the Indian Subcontinent
Constantinescu, Ioana Cristina1 ; Popa, Oana Paula2 ; Popa, Luis Ovidiu3 ; Cobzaru, Ioana4 ; Mukhim, D. Khlur B.5 and Adam, Costică6
1✉ “Grigore Antipa” National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest 1, Romania.
2“Grigore Antipa” National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest 1, Romania.
3“Grigore Antipa” National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest 1, Romania.
4Ecology, Taxonomy and Nature Conservation Department, Institute of Biology, Romanian Academy, Splaiul Independenței no. 296, 060031 Bucharest, Romania.
5Department of Zoology, Lady Keane College, 793001 Shillong, Meghalaya, India.
6“Grigore Antipa” National Museum of Natural History, Sos. Kiseleff no.1, 011341 Bucharest 1, Romania.
2018 - Volume: 58 Issue: 4 pages: 881-896
https://doi.org/10.24349/acarologia/20184294ZooBank LSID: F5A394F4-E894-4675-9166-CA0C3C5D67A3
Keywords
Abstract
The feather mite genus Montesauria Oudemans, 1905 (Analgoidea: Proctophyllodidae) is the species-richest genus of the subfamily Pterodectinae and currently includes 62 species distributed mostly in the Old World (Till 1954, 1957, Gaud 1957, Gaud and Mouchet 1957, Mironov and Kopij 1996a, 1996b, 1997, Mironov and Fain 2003, Mironov 2006, 2008, Kuroki et al. 2006, Hernandes et al. 2010, Mironov et al. 2010, 2012, Mironov and Tostenkov 2013, Hernandes and OConnor 2017). To date, 12 species of Montesauria have been described from Asia: 8 from Vietnam (Gaud and Petitot 1948, Mironov et al. 2012, Mironov and Tostenkov 2013), 2 from Japan (Kuroki et al. 2006), 1 from Taiwan (Sugimoto 1941) and 1 from Indonesia (Trouessart 1885). Atyeo (in: McClure and Ratanaworabhan 1973), reported 81 supposedly new species of this genus from Southeastern Asia; unfortunately, new species from this material have never been described. Consequently, this genus seems to be widespread on passerines of the Old World. Mironov (2006) performed a rigorous systematic study and based on clearly differentiated morphological characters divided the genus into nine species groups: cylindrica, dolichodectina, emberizae, heterocaula, jesionowskii, listroprocta, merulae, pachypa and papillo. Later, he proposed one additional group, macronoi (Mironov et al. 2012).
The present paper gives descriptions of two new Montesauria species from the merulae species group found on thrushes (Turdidae) in India.
The material used in the present paper was collected in the neighborhood of Shnongrim (Meghalaya, India) in January 2014. The birds were captured using mist-nets, identified and visually checked for the presence of mites and after collecting of mites released back to the wild. Mite specimens were collected manually with a needle and placed in tubes with 96% ethanol. Then, in laboratory conditions, mite specimens were cleared in 90% lactic acid for 24 hours and mounted on microscope slides in Hoyer’s medium. Drawings were made using an Olympus CX21 microscope with a camera lucida drawing device. The bird specimens were identified according to Rasmussen and Anderton (2012) and Grimmett et al. (2011), and the taxonomy of birds follows Clements et al. (2017). The body setation of mites follows that of Griffiths et al. (1990) with the modifications by Norton (1998) concerning coxal setae, while the setation of legs follows Gaud and Atyeo (1996). The description of the new species of Montesauria is given according to the current format used for species of pterodectine mites (Mironov and Fain 2003, Mironov 2006, Valim and Hernandes 2006, Mironov et al. 2008, 2010, 2012, Mironov and Tolstenikov 2013, Hernandes and OConnor 2017) and the measuring techniques of particular structures used were described by Mironov and Proctor (2009).
We give the full set of measurements for a holotype (male) and a range of measurements for corresponding paratypes. All measurements are in micrometers (μm). The holotype and paratypes of both new species are deposited in the Acarological Collection of the “Grigore Antipa” National Museum of Natural History, Bucharest, Romania (MGAB). Inventory numbers are given in brackets for all type specimens.
Six paratype specimens of Montesauria hernandesi sp. n. (three males ANA1442 C1, ANA1443 C2, ANA1444 C3, and three females ANA1445 C4, ANA 1446 C5, ANA1447 C6) and three paratype specimens of M. caerulea sp. n. (two males ANA1436 B1, ANA1438 B3, and one female ANA1441 B5) were used to isolate DNA using DNAeasy Tissue Kit (Qiagen). All specimens used for molecular analyses were mounted and kept as reference vouchers for morphological examination. The specimens preserved in ethanol 96% were transferred in 180μl ATL Buffer with 20 μl of Proteinase K and incubated overnight at 56 °C on a shaking thermoblock. After 24 h, 5 μL of Proteinase K were added and incubation was continued until 72h. For the rest of the protocol we followed the manufacturer specifications and the modification suggested by Dabert et al. (2008).
We used as DNA barcode region a 663bp fragment near the 5' terminus of the COI gene, amplified by PCR with the degenerate primers bcdF05 (5' – TTTTCTACHAAYCATAAAGATATTGC – 3') and bcdR04 (5`– TATAAACYTCDGGATGNCCAAAAAA – 3'), according to Dabert et al. (2008). The PCRgenotyping reaction was performed in a 50 μL total volume containing DNA template, 1X Green GoTaq® Flexi Buffer, 2.5 mM MgCl2, each dNTP at 0.1 mM, each primer at 0.5 μM (the primers were M13 tailed) and 1.5 units of GoTaq® DNA polymerase (5U/μL) (Promega, Madison, USA). The PCR products were isolated from samples containing visible bands and sent for sequencing to Macrogen (Seoul, Korea).
Sequence electrophoregrams were edited and assembled with CodonCode Aligner version 3.7.1. Pairwise distances between sequences were computed with MEGA version 7 (Kumar et al. 2016) using K2P distance model (Kimura 1980). DnaSP v5 was used to obtain data about the genetic polymorphism in the studied specimens (Librado and Rozas 2009).
Family Proctophyllodidae Trouessart & Mégnin, 1884
Subfamily Pterodectinae Park & Atyeo, 1971
Genus Montesauria Oudemans, 1905
ZOOBANK: 9689B8C4-F5BC-418D-BDC8-A39B46050FDF ![]()
Type material — Holotype male and 25 paratypes (12 males and 13 females), from Turdus dissimilis Blyth, 1847 (Passeriformes: Turdidae), India, Meghalaya, Jaintia Hills, Shnongrim village, 25°21'12.36"N, 92°31'3.06"E, 25 Jan. 2014, 1151 m, subtropical forest, collector D. K. B. Mukhim.
Type deposition — Holotype male (ANA1412), 12 males (ANA1408 –1411, ANA1413–1417, ANA1442 C1, ANA1443 C2, ANA1444 C3) and 13 female (ANA1418–1427, ANA1445 C4, ANA1446 C5, ANA1447 C6) paratypes in MGAB collection.
Etymology — The species is named after Dr. Fabio Akashi Hernandes (Universidade Estadual Paulista, Rio Claro, São Paulo, Brazil), a prominent expert on feather mites.
Male (holotype, range for 4 paratypes). Length of idiosoma 296 (292–296), width 106 (90–114), length of hysterosoma 204 (200–208).
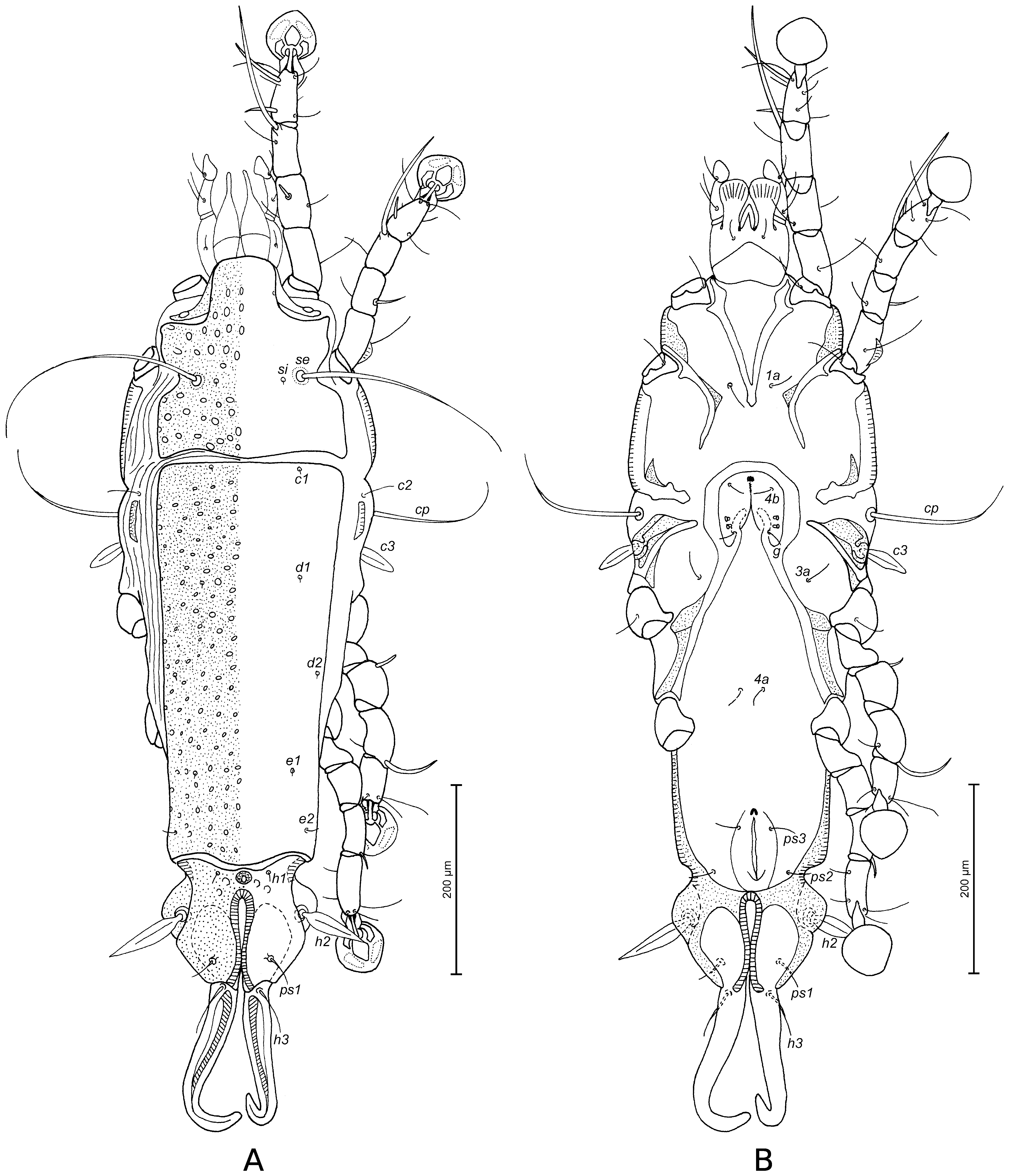
Prodorsal shield: entire, lateral margins slightly concave at level of scapular setae, posterior margin with acute median extension, length along midline 90 (88–96), width at posterior margin 80 (82–90), surface with round and ovate lacunae. Setae ve absent. Scapular setae se separated by 43 (42–46). Humeral shields poorly developed, represented by narrow dorsal sclerites, separated from epimerites III, not encompassing setae cp. Setae c2 situated dorsally on soft tegument. Subhumeral setae c3 lanceolate, 23 (19–24) × 8 (6–7). Hysteronotal shield: greatest length from anterior margin to lobar apices 190 (180–190), width at anterior margin 80 (82–90), anterior margin almost straight, surface of this shield with round and ovate lacunae. Opisthosomal lobes well developed, short and almost straight, posterior margin of lobes with small notch, terminal cleft V-shaped, 21 (18–20) in length. Setae f2 absent, setae h2 and h3 slightly distant from margin, h3 spindle-shaped, with acute apices, 40 (38–42) in length, 4 (4–5) in width, h2 144 (126–150) in length, 5 (4–5) in width. Supranal concavity circular, well outlined. Setae h1 situated at level of anterior margin of supranal concavity. Setae ps1 situated in anterior half of opisthosomal lobes, approximately equidistant from outer and inner margins of these lobes, 6 (5–6) in length. Setae ps2 70 (70–80) in length (Fig. 1A). Distance between bases of dorsal setae: c2–d2 80 (72–81), d2–e2 70 (74–80), e2–h3 36 (34–40), d1–d2 45 (29–42), e1–e2 24 (23–34), h1–ps2 24 (22–27), h2–h2 40 (38–40), h3–h3 23 (20–24), ps2–ps2 54 (48–54).


Epimerites I fused posteriorly into a V, fused part with a pair of rounded transverse extensions almost touching medial part of epimerites II (Fig. 1B). Coxal fields I, II without wide sclerotized areas. Rudimentary sclerites rEpIIa present. Coxal fields I, II and III open. Epimerites IVa present, long, exceeding level of setae 4a bases. Genital arch with wing-like lateral extensions, 20 (17–22) in length, 24 (20–25) in width including extensions, basal sclerite of genital apparatus shaped as inverted trapezium, aedeagus 55 (54–58) long, extending to level of anterior margin of anal suckers. Genital papillae situated at midlevel of genital apparatus, touching at their bases. Genital shield absent. Anal suckers 9 (8–9) in diameter, corolla without indentations, surrounded by radially striated area. Opisthoventral shields well-developed, roughly triangular, situated posterior to the level of anal suckers, setae ps3 situated on these shields. Anterior end of terminal cleft surrounded by small sclerotized plate (Fig. 1B). Distance between ventral setae: 3a–4b 24 (18–28), 4b–4a 32 (26–33), 4a–g 34 (34–42), g–ps3 40 (36–42), ps3–ps3 32 (32–34), ps3–h3 21 (22–25).
Legs I slightly longer than legs II, femora II with ventral crest. Solenidion σ1 of genu I spiculiform, situated in proximal half of the segment (Fig. 2). Setae mGII thickened basally, with filiform apex. Setae d of tarsi II, III shorter than corresponding setae f. Genu IV with ovate ventral apophysis. Tarsus IV with bidentate apex, 20 (20–22) in length, setae d and e ovate and small, diameter of seta e twice as short as that in seta d (Fig. 2D). Length of solenidia: ω1I 10 (8–10), ω1II 10 (8–10), σ1I 4 (4–9), σIII 6 (5–7), φIII 20 (20–25), φIV 24 (27–30).


Female (range for 5 paratypes). Length of idiosoma excluding terminal appendages 396–408, width 122–150, length of hysterosoma 284–292.
Prodorsal shield: shape and ornamentation as in male, 86–110 long, 98–112 wide (Fig. 3A). Setae ve absent. Scapular setae se separated by 52–56. Humeral shields poorly developed, represented by narrow dorsal sclerites. Setae c2 situated on soft tegument. Setae c3 lanceolate, 24–26 × 6–7. Anterior and lobar parts of hysteronotal shield separated by narrow transverse band of soft tegument. Hysteronotal shield: anterior margin straight, posterior margin convex in median part, length 208–220, width at anterior margin 92–112, with ovate lacunae. Length of lobar region excluding terminal appendages 60–70, width at level of setae h2 76–82, median part of anterior margin slightly convex, ornamentation with few ovate lacunae in anterior part. Terminal cleft narrow, with lateral margins almost parallel and touching at level of setae ps1, 48–54 long. Supranal concavity circular, small, strongly outlined. Setae f2 absent, setae h1 on lobar shield, near its anterior margin. Setae h2 spindle-like, with acute apices, 48–52 long, 8–10 wide. Setae h3 filiform, 26–30 in length, about 1/3 of the length of terminal appendages. Setae ps1 situated dorsally on lobes. Distance between bases of dorsal setae: c2–d2 98–108, d2–e2 84–90, e2–h2 40–46, h2–h3 40–42, d1–d2 47–58, e1–e2 27–36, h1–h2 22–30, ps1–h2 27–30, h1–h1 22–28, h2–h2 56–62.

Epimerites I fused into a Y, with short sternum (Fig. 3B). Lateral parts of coxal fields I, II without heavily sclerotized areas. Epimerites II do not reach the level of anterior margin of epigynum. Epimerites IVa absent. Translobar apodemes of opisthosomal lobes present, fused to each other anterior to terminal cleft. Epigynum horseshoe-shaped, greatest width 50–60, apodemes of ovipore connected with coxal apodemes IIIa. Head of spermatheca weakly sclerotized, with longitudinal striation, secondary spermaducts longer than distance between large bulb-shaped enlargement of primary spermaduct and small bulge near head of spermatheca. Bulb-shaped enlargement of primary spermaduct with small tubercles (Fig. 4E). Distance between pseudanal setae: ps2–ps2 37–40, ps3–ps3 18–20, ps2–ps3 19–22.
Legs I slightly longer than legs II, femur II with ventral crest; other segments of legs I, II without processes. Solenidion σ1 of genu I spiculiform, situated in distal half of the segment (Fig. 4A). Setae mGII thickened basally, with filiform apex. Setae d of tarsi II–IV much shorter than corresponding setae f. Length of solenidia: ω1I 8–12, ω1II 7–10, σ1I 6–11, σIII 4–6, φIII 22–26, φIV 5–9.


Differential diagnosis — The new species, Montesauria hernandesi sp. n., belongs to merulae species group and is most similar to M. sibirica Kuroki, Nagahori and Mironov, 2006 described from the Siberian thrush Geokichla sibirica davisoni (Hume, 1877) (Passeriformes, Turdidae) (Kuroki et al. 2006). In males of both species, the posterior margin of opisthosomal lobes is notched, the opisthoventral shields have a triangular form, and the inner angles of these shields are slightly posterior to the level of anal suckers, the anterior end of terminal cleft is surrounded by small sclerotized plate, setae h1 are situated at the level of anterior margin of supranal concavity, the genua IV have an ovate ventral apophysis, setae d and e of tarsus IV are ovate, the diameter of seta e is twice as short as that in seta d; in females, the primary spermaduct has a bulb-shaped enlargement, and setae h3 are about 1/3 the length of terminal appendages. Montesauria hernandesi sp. n. clearly differs from M. sibirica by the following features (corresponding character states of M. sibirica in parentheses): in both sexes, the ornamentation of prodorsal and hysteronotal shields is represented by ovate lacunae (vs. small pit-like lacunae), in males, tarsus IV has a bidentate apex (vs. with apical claw-like process); in females, the lobar shield bears a few ovate lacunae in the anterior part (vs. without ornamentation), epimerites I are fused into a Y (vs. fused into a V), the secondary spermaducts are longer than the distance between the large bulb-shaped enlargement of primary spermaduct and the small bulge near the head of spermatheca (vs. shorter than this distance).
ZOOBANK: 26BD4193-539F-4C7E-A27B-A7972AB8AD43 ![]()
Type material — Holotype male and 22 paratypes (11 males and 11 females), from Myophonus caeruleus (Scopoli, 1786) (Passeriformes: Turdidae), India, Meghalaya, Jaintia Hills, Shnongrim village, 25°21'12.36"N, 92°31'3.06"E, 22 Jan. 2014, 1151 m, subtropical forest, collector D. K. B. Mukhim.
Type deposition — Holotype male (ANA 1395), 11 males (ANA1388–1394, ANA1396–1397, ANA1436 B1, ANA1438 B3) and 11 females (ANA1398–1407, ANA1441 B5) paratypes in MGAB collection.
Etymology — The species epithet is taken from the species name of the type host and is an adjective.
Male (holotype, range for 4 paratypes). Length of idiosoma 316 (312–332), width 116 (114–124), length of hysterosoma 228 (224–232).
Prodorsal shield: entire, lateral margins slightly concave at level of scapular setae, posterior margin concave, length along midline 88 (84–94), width at posterior margin 94 (90–93), surface with ovate lacunae. Setae ve absent. Scapular setae se separated by 44 (44–48). Humeral shields poorly developed, represented by narrow dorsal sclerites, separated from epimerites III, not encompassing setae cp. Setae c2 situated dorsally on soft tegument. Subhumeral setae c3 lanceolate, 22 (22–23) × 6 (6–8). Prodorsal and hysteronotal shields almost touching in median part, separated by very narrow transverse band of soft tegument. Hysteronotal shield: greatest length from anterior margin to lobar apices 220 (208–232), width at anterior margin 96 (86–92), anterior margin slightly convex, surface of this shield with big circular lacunae in anterior half and small ovate lacunae in posterior half. Opisthosomal lobes well developed, short and with apices slightly narrowed, distal margins with small notch, terminal cleft V-shaped, 22 (20–22) in length. Setae f2 absent, setae h2 and h3 slightly distant from margin, h3 spindle-shaped, 66 (62–72) in length, 4 (4–5) in width, h2 164 (160–180) in length, 6 (4–6) in width. Supranal concavity circular, well outlined. Setae h1 situated anterior to the level of supranal concavity. Setae ps1 situated in anterior half of opisthosomal lobes, approximately equidistant from outer and inner margins of these lobes, 7 (5–8) in length. Setae ps2 84 (70–90) in length (Fig. 5A). Distance between bases of dorsal setae: c2–d2 80 (78–80), d2–e2 100 (100–110), e2–h3 32 (26–34), d1–d2 44 (34–42), e1–e2 40 (46–52), h1–ps2 38 (22–36), h2–h2 38 (30–38), h3–h3 22 (20–22), ps2–ps2 50 (46–50).


Epimerites I fused posteriorly into a V, fused part with transverse extensions connected to medial part of epimerites II (Fig. 5B). Coxal fields I, II without wide sclerotized areas. Rudimentary sclerites rEpIIa absent. Coxal fields I, II and III open. Epimerites IVa present, long, exceeding bases of setae 4a. Genital arch with wing-like extensions, 22 (18–21) in length, 30 (26–28) in width including extensions, basal sclerite of genital apparatus shaped as inverted trapezium, aedeagus 60 (56–60) long, reaching anterior margin of anal opening. Genital papillae situated at midlevel of genital apparatus, connected by their bases. Genital shield absent. Anal suckers 11 (10–12) in diameter, corolla without indentations, surrounded by striated area. Opisthoventral shields well-developed, trapezoidal in shape, situated at level of anal suckers, setae ps3 situated on these shields. Anterior end of terminal cleft surrounded by small sclerotized plate (Fig. 5B). Distance between ventral setae: 3a–4b 26 (24–26), 4b–4a 34 (32–40), 4a–g 34 (34–36), g–ps3 42 (42–46), ps3–ps3 40 (38–40), ps3–h3 32 (30–34).
Legs I slightly longer than legs II, femora I, II with ventral crest. Solenidion σ1 of genu I spiculiform, situated in proximal half of the segment (Fig. 6A). Setae mGII thickened basally, with filiform apex. Setae d of tarsi II, III shorter than corresponding setae f. Genu IV with semi-rounded ventral apophysis. Tarsus IV with bidentate apex, 24 (20–24) in length, setae d and e button-like. (Fig. 6D). Length of solenidia: ω1I 12 (8–12), ω1II 8 (6–8), σ1I 9 (6–10), σIII 7 (6–7), φIII 22 (22–24), φIV 30 (26–31).


Female (range for 5 paratypes). Length of idiosoma (excluding terminal appendages) 412–440, width 126–150, length of hysterosoma 308–336.
Prodorsal shield: shaped as in male, 94–104 long, 94–118 wide, with ovate lacunae in anterior half and few circular and ovate lacunae in central part of posterior half (Fig. 7A). Setae ve absent. Scapular setae se separated by 50–58. Humeral shields present, poorly developed as narrow dorsal sclerites. Setae c2 situated on soft tegument. Setae c3 lanceolate, 24–25 × 7–8. Anterior and lobar parts of hysteronotal shield separated by narrow transverse band of soft tegument. Hysteronotal shield: anterior margin straight, posterior margin convex in median part, length 236–248, width at anterior margin 100–120, surface of this shield with big and almost round lacunae in anterior half and small ovate lacunae in posterior half. Length of lobar region excluding terminal appendages 72–80, width at level of setae h2 72–80, median part of anterior margin slightly convex, anterior part few ovate lacunae. Terminal cleft narrow, with lateral margins almost touching in posterior half, 48–54 long. Supranal concavity circular, small, strongly outlined. Setae f2 absent, setae h1 on lobar shield near its anterior margin. Setae h2 spindle-like, with acute apices, 40–50 long, 8–9 wide. Setae h3 filiform, 30–46 in length, about 1/2 of the length of terminal appendages. Setae ps1 situated dorsally on lobes. Distance between bases of dorsal setae: c2–d2 102–120, d2–e2 88–98, e2–h2 50–54, h2–h3 36–40, d1–d2 55–64, e1–e2 30–40, h1–h2 28–34, ps1–h2 28–34, h1–h1 18–22, h2–h2 54–60.


Epimerites I fused into a Y, with short sternum (Fig. 7B). Lateral parts of coxal fields I, II without heavily sclerotized areas. Epimerites II do not reach the level of anterior margin of epigynum. Epimerites IVa absent. Translobar apodemes of opisthosomal lobes present, fused to each other anterior to terminal cleft. Epigynum horseshoe-shaped, greatest width 54–66, apodemes of ovipore connected with coxal apodemes IIIa. Head of spermatheca weakly sclerotized, with longitudinal striation, secondary spermaducts shorter than distance between large bulb-shaped enlargement of primary spermaduct and small bulge near head of spermatheca. Bulb-shaped enlargement of primary spermaduct with small tubercles (Fig. 8E). Distance between pseudanal setae: ps2–ps2 34–40, ps3–ps3 16–22, ps2–ps3 24–29.
Femur II with ventral crest; other segments of legs I, II without processes. Solenidion σ1 of genu I spiculiform, situated in distal half of the segment (Fig. 8A). Setae mGII thickened basally, with filiform apex. Setae d of tarsi II–IV much shorter than corresponding setae f. Length of solenidia: ω1I 10–16, ω1II 8–10, σ1I 6–11, σIII 6–8, φIII 22–30, φIV 6–10.


Differential diagnosis — Montesauria caerulea sp. n. belongs to merulae species group and is most similar to M. aurea Kuroki, Nagahori and Mironov, 2006 described from White's thrush Zoothera aurea toratugumi (Momiyama, 1940) (Passeriformes, Turdidae) in Japan (Kuroki et al. 2006). Males of the both species have a similar shape of opisthosomal lobes, which are short, with apices slightly narrowed, and with notched distal margins, the aedeagus does not reach the level of anterior margin of anal suckers, the genua IV have semi-round ventral apophysis, the setae h1 are situated anterior to the level of the supranal concavity; females have setae h3 about 1/2 the length of terminal appendages. Montesauria caerulea sp. n. clearly differs from M. aurea by the following features (corresponding character states of M. aurea in parentheses): in both sexes, the ornamentation of hysteronotal shield is represented by big and almost circular lacunae in the anterior half and small ovate lacunae in the posterior half (vs. small pit-like lacunae), in males, epimerites I and II are connected (vs. disconnected), the basal sclerite of the genital apparatus is shaped like an inverted trapezium (vs. crescent-shaped), the opisthoventral shields are almost trapezoidal in shape and located at the level of anal suckers (vs. almost triangular and located slightly anterior to the level of anal suckers), tarsus IV has bidentate apex (vs. apical claw-like process). In females of the new species, the humeral shields are present (vs. humeral shields absent), the epimerites I are fused into a Y (vs. fused into a V), the posterior margin of the hysteronotal shield is convex in median part (vs. concave), the secondary spermaducts are shorter than the distance between the large bulb-shaped enlargement of the primary spermaduct and the small bulge near the head of spermatheca (vs. longer than that distance).
GenBank accession numbers for molecular voucher ANA1442 C1 male MH746221, ANA1443 C2 male MH746222, ANA1444 C3 male MH746223, ANA1445 C4 female MH746224, ANA1446 C5 female MH746225, ANA1447 C6 female MH746226.
In the resulted alignment of 482-pb fragment of the mitochondrial cytochrome C oxidase subunit I (COI) gene, we identified 5 polymorphic sites (three with singleton variable sites and two parsimony informative ones). Three haplotypes were identified: H1 (ANA1442 C1 male, ANA1446 C5 female), H2 (ANA1443 C2 male), H3 (ANA1444 C3 male, ANA1445 C4 female, ANA1447 C6 female). Intraspecific genetic distances between the analyzed specimens using K2P model is 0.5%. Also, all observed nucleotide substitutions were synonymous and did not have differences in the amino acid sequence.
GenBank accession numbers for molecular voucher ANA1436 B1 male MH746227, ANA1438 B3 male MH746228, ANA1441 B5 female MH746229.
In the resulted alignment of 482-pb fragment of the mitochondrial cytochrome C oxidase subunit I (COI) gene, we identified 4 singleton variable sites. Three haplotypes were identified: H1 (ANA1436 B1 male), H2 (ANA1438 B3 male) and H3 (ANA1441 B5 female). Intraspecific genetic distances between the analyzed specimens using K2P model is 0,6%. All observed nucleotide substitutions were synonymous and did not have differences in amino acid sequence.
The reported intraspecific genetic distances for the two species of Montesauria described in the present study are comparable with genetic distances found in other related mites like Proterothrix chachulae (0.8%) (Constantinescu et al. 2017) also belonging to the subfamily Pterodectinae and distributed the Oriental Region.
We are grateful to the Additional Principal Chief Conservator of Forests, Wildlife & Chief Wildlife Warden from Shillong (Meghalaya, India) for the permission to catch birds (permission No. FWC.G/173/Pt.). Ioana Cobzaru was supported by the Institute of Biology Bucharest of Romanian Academy (project no. RO1567–IBB04/2018).
Clements J.F., Schulenberg T.S., Iliff M.J., Roberson D., Fredericks T.A., Sullivan B.L., Wood C.L. 2016. The eBird — Clements checklist of birds of the world [Internet]. Version 2017. Available from: rlbibhttp://www.birds.cornell.edu/clementschecklist/download/
Constantinescu I.C., Popa O.P., Popa L.O., Cobzaru I., Mukhim D.K.B., Adam C. 2017. A new feather mite species of the genus Proterothrix Gaud, 1968 (Acarina, Proctophyllodidae) from the Large Niltava, Niltava grandis (Passeriformes, Muscicapidae) – an integrative description. ZooKeys, 661: 1-14. doi:10.3897/zookeys.661.11793 doi:10.3897/zookeys.661.11793 ![]()
Dabert J., Ehrnsberger R., Dabert M. 2008. Glaucalges tytonis sp. n. (Analgoidea, Xolalgidae) from the barn owl Tyto alba (Strigiformes, Tytonidae): compiling morphology with DNA barcode data for taxon descriptions in mites (Acari). Zootaxa, 1719: 41-52. doi:10.5281/zenodo.181058
Gaud J., Atyeo W.T. 1996. Feather mites of the world (Acarina, Astigmata): the supraspecific taxa. Ann. Mus. Roy. Afr.centr., 277: 1-191 (Part 1, text), 1-436 (Part2, illustrations).
Gaud J. 1957. Acariens plumicoles (Analgesoidea) parasites des oiseaux du Maroc. Bull. Soc. Sci. Nat. Phys. Maroc, 37: 105-136.
Gaud J., Mouchet J. 1957. Acariens plumicoles (Analgesoidea) des oiseaux du Cameroun. I. Proctophyllodidae. Ann. Parasitol. Hum. Comp., 32: 491-546.
Gaud J., Petitot M.L. 1948. Sarcoptides plumicoles des oiseaux d'Indochine. Ann. Parasit. Hum. Comp., 23 : 337-347. doi:10.1051/parasite/1948235337 ![]()
Griffiths D.A., Atyeo W.T., Norton R.A., Lynch C.A. 1990. The idiosomal chaetotaxy of astigmatid mites. Jour. Zool, 220: 1-32. doi:10.1111/j.1469-7998.1990.tb04291.x doi:10.1111/j.1469-7998.1990.tb04291.x ![]()
Grimmett R., Inskipp C., Inskipp T. 2011. Helm Field Guides: Birds of the Indian Subcontinent. Christopher Helm, London, 528 pp.
Hernandes F.A., OConnor B.M. 2017. Out of Africa: the mite community (Arachnida: Acariformes) of the common waxbill, Estrilda astrild (Linnaeus, 1758) (Passeriformes: Estrildidae) in Brazil. Parasite. Vector., 10: 299. doi:10.1186/s13071-017-2230-5 doi:10.1186/s13071-017-2230-5 ![]()
Hernades F.A., Valim M.P., Mironov S.V. 2010. On the identity of Pterodectes ralliculae Atyeo and Gaud, 1977 (Astigmata: Proctophyllodidae). J. Nat. Hist., 44: 369-377. doi: 10.1080/00222930903383594 doi:10.1080/00222930903383594 ![]()
Kimura M. 1980. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol., 16: 111-120. doi:10.1007/BF01731581 ![]()
Kumar S., Stecher G., Tamura K. 2016. MEGA7: Molecular evolutionary genetics enalysis version 7.0 for bigger datasets. Molecular Biology and Evolution Jul;33 (7):1870-4. doi: 10.1093/molbev/msw054. Epub 2016 Mar 22. doi:10.1093/molbev/msw054 ![]()
Kuroki T., Nagahori M., Mironov S.V. 2006. Two new feather mite species of the genus Montesauria (Astigmata: Proctophyllodidae) from Thrushes of the genus Zoothera (Passeriformes: Turdidae) in Japan. J. Acarol. Soc. Jpn., 15: 55-68. doi:10.2300/acari.15.55 ![]()
Librado P., Rozas J. 2009. DnaSP v5 - A software for a comprehensive analysis of DNA polymorphism data. Bioinformatics, 25: 1451-1452. doi:10.1093/bioinformatics/btp187 doi:10.1093/bioinformatics/btp187 ![]()
McClure H.E., Ratanaworabhan N. 1973. Some ectoparasites of the birds of Asia, Applied Scientific Research, Jintana Printing Ldt, Bangkok, 219 pp.
Mironov S.V. 2006. Feather mites of the genus Montesauria Oudemans (Astigmata: Proctophyllodidae) associated with starlings (Passeriformes: Sturnidae) in the Indo- Malayan region, with notes on systematic of the genus. Acarina, 14: 21-40.
Mironov S.V. 2008. Three new species of the feather mite subfamily Pterodectinae (Acari: Astigmata: Proctophyllodidae) from passerines (Aves: Passeriformes) in Central Africa. Ann. Zool., 58: 403-418. doi:10.3161/000345408X326744 doi:10.3161/000345408X326744 ![]()
Mironov S.V., Fain A. 2003. New species of feather mite subfamily Pterodectinae (Astigmata: Proctophyllodidae) from African passerines (Aves: Passeriformes). Bull. Annls. Soc. Roy. Belge. Ent., 139: 75-91.
Mironov S.V., Kopij G. 1996a. New feather mite species (Acarina: Analgoidea) from some starlings (Passeriformes: Sturnidae) of South Africa. J. Afr. Zool., 110: 257-269.
Mironov S.V., Kopij G. 1996b. Three new species of the feather mite family Proctophyllodidae (Acarina: Analgoidea) from some south African passerine birds (Aves: Passeriformes). Acarina, 4: 27-33.
Mironov S.V., Kopij G. 1997. New feather mite species of the subfamily Pterodectinae (Astigmata: Proctophyllodidae) from some passerines (Aves: Passeriformes) of South Africa. J. Afr. Zool., 111: 449-463.
Mironov S.V., Proctor H.C. 2009. Feather mites of the genus Proterothrix Gaud (Astigmata: Proctophyllodidae) from parrotbills (Passeriformes: Paradoxornithidae) in China. J. Parasitol., 95:1093-1107. doi:10.1645/GE-1961.1 doi:10.1645/GE-1961.1 ![]()
Mironov S.V., Tolstenkov O.O. 2013. Three new feather mites of the subfamily Pterodectinae (Acari: Proctophyllodidae) from passerines (Aves: Passeriformes) in Vietnam. Proc. Zool. Inst. Russ. Acad.Sci., 317: 11-29.
Mironov S.V., Literak I., Čapek M. 2008. New feather mites of the subfamily Pterodectinae (Acari: Astigmata: Proctophyllodidae) from passerines (Aves: Passeriformes) in Mato Grosso do Sul, Brazil. Zootaxa, 1947: 1-38. doi:10.11646/zootaxa.4297.1.1 doi:10.11646/zootaxa.4297.1.1 ![]()
Mironov S.V., Literak I., Čapek M., Koubek P. 2010. New species of the feather mite subfamily Pterodectinae (Astigmata, Proctophyllodidae) from passerines in Senegal. Acta Parasitol., 55: 399-413. doi:10.2478/s11686-010-0051-1 doi:10.2478/s11686-010-0051-1 ![]()
Mironov S.V., Literak I., Hung M.N., Čapek M. 2012. New feather mites of the subfamily Pterodectinae (Acari: Proctophyllodidae) from passerines and woodpeckers (Aves: Passeriformes and Piciformes) in Vietnam. Zootaxa, 3440: 1-49.
Norton A.R. 1998. Morphological evidence for the evolutionary origin of Astigmata (Acari: Acariformes). Exp. Appl. Acarol., 22: 559-594. doi:10.1023/A:1006135509248 doi:10.1023/A:1006135509248 ![]()
Rasmussen P.C., Anderton J.C. 2012. Birds of South Asia. The Ripley Guide. Volumes 1 and 2. Second Edition. National Museum of Natural History – Smithsonian Institution, Michigan State University and Lynx Edicions, Washington, D.C., Michigan and Barcelona, vol. 1: 684 pp., vol. 2: 378 pp.
Sugimoto M. 1941. Studies on the Formosan mites (Fourth Report) (On the feather mites, Analgesidae Canestrini, 1892, Part II). Sylvia, 5: 129-149.
Till W.M. 1954. Five new feather mites of the genus Pterodectes (Acarina: Analgesoidea). Moçambique, 79: 85-100.
Till W.M. 1957. Two new Pterodectes species from passeriform birds (Acarina: Proctophyllodidae). J. Entomol. Soc. South. Afr., 20: 450-453.
Valim M.P., Hernandes F.A. 2006. Redescription of four species of the feather mite genus Pterodectes Robin, 1877 (Acari: Proctophyllodidae: Pterodectinae) described by Herbert F. Berla. Acarina, 14: 41-55.



2018-07-24
Date accepted:
2018-10-22
Date published:
2018-10-30
Edited by:
Auger, Philippe

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Constantinescu, Ioana Cristina; Popa, Oana Paula; Popa, Luis Ovidiu; Cobzaru, Ioana; Mukhim, D. Khlur B. and Adam, Costică
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)