



First quantitative data on the ectoparasitic mites of Sceloporus torquatus (Squamata) from the Ecological Reserve of Pedregal de San Angel in Central Mexico
Guzmán-Cornejo, Carmen1 ; García-Prieto, Luis2 and Zúñiga-Vega, J. Jaime3
1✉ Laboratorio de Acarología, Departamento de Biología Comparada, Facultad de Ciencias, Universidad Nacional Autónoma de México, Mexico City, Mexico.
2Colección Nacional de Helmintos, Instituto de Biología, Universidad Nacional Autónoma de México, Mexico City, Mexico.
3Laboratorio de Ecología Evolutiva y Demografía Animal, Departamento de Ecología y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Autónoma de México, Mexico City, Mexico.
2018 - Volume: 58 Issue: 4 pages: 868-874
https://doi.org/10.24349/acarologia/20184292Keywords
Abstract
The Reserva Ecológica del Pedregal de San Ángel (REPSA) is a natural area of 237.3 ha protected by the Universidad Nacional Autónoma de México (UNAM), located in their central campus at Mexico City; it has a vegetation of xerophytic scrub in low areas, as well as lake landscapes and wetlands located in a buffer zone known as the Cantera Oriente (Peralta-Higuera and Prado-Molina 2009). Its specific richness of amphibians and reptiles is high – about 30 species, including turtles, lizards and snakes. In the particular case of the lizards, six species of the genera Barisia and Sceloporus have been reported, the latter represented by Sceloporus aeneus Wiegmann, 1828, Sceloporus anahuacus Lara-Góngora, 1983, Sceloporus grammicus Wiegmann, 1828, Sceloporus palaciosi Lara-Góngora, 1983, and Sceloporus torquatus Wiegmann, 1828.
The spiny collared lizard (S. torquatus) is a species endemic to Mexico, distributed in the north and center of the country (Méndez de la Cruz et al. 2009). In the REPSA it is distributed from 2240 to 2580 m.a.s.l., being abundant in moderately disturbed areas and on the border of the Reserve (Méndez de la Cruz et al. 2009).
Known mite fauna associated with this lizard species includes four species of Trombidiformes (Acariformes): three Pterygosomatidae, Geckobiella texana (Banks, 1904), Geckobiella pelaezi (Cunliffe, 1949), Geckobiella trombidiformis (Berlese, 1920) and one Trombiculidae, Eutrombicula alfreddugesi (Oudemans, 1910) (Paredes-León et al., 2008). Specifically in the REPSA, previous record for this lizard includes three mite species: G. texana, G. pelaezi and E. alfreddugesi (Paredes-León et al. 2008; Montiel-Parra et al. 2009). However, no study analyzing the population parameters of this acari-lizard association has been conducted. Objective of our work was to determine main parameters of mite infestation on S. torquatus in the REPSA during rainy season of 2016.
The 24 S. torquatus analyzed were collected during May and October of 2016 (rainy season) in the botanical garden of the Instituto de Biología at UNAM, located in the buffer zone of REPSA (19.31826N, 99.19431W) (Peralta-Higuera and Prado-Molina, 2009). Lizards were collected using a slipknot tied to a fishing rod, under collection permit SGPA/DGVS/11338/15, issued to JJZV by the Secretaría de Medio Ambiente y Recursos Naturales, Mexico. They were released after their examination. Mites were removed directly from the body surface of the host using brushes and fine-tipped tweezers, and a census of mites on each host was made. In each collection date, we fixed and conserved all mites obtained from an individual host in a vial with 80% ethanol. Specimens were mounted in Hoyer’s medium and identified under compound microscope (Leica ICC50HD), using specialized taxonomic keys (Kohls et al. 1965; Hoffmann 1990; Paredes-León et al. 2012). Reference material of each mite species was deposited in the Colección del Laboratorio de Acarología “Anita Hoffmann”, Facultad de Ciencias, UNAM, Mexico City, with the catalog numbers LAFC-A 132-142. Data from both collection dates was pooled. For each mite species found in S. torquatus, the following ecological parameters were calculated: prevalence, mean abundance, and mean intensity (according to Bush et al. 1997). We conducted statistical comparisons of these three ecological parameters among mite species by building and comparing confidence intervals through a bootstrap resampling procedure (1000 replicates) (Manly 2007). These intervals had a confidence level of 84%, which, according to MacGregor-Fors and Payton (2013), is equivalent to an analysis of variance with a significance level of 0.05.
A total of 1251 mites belonging to three families and four species (Argasidae: Ornithodoros talaje (Guèrin-Méneville, 1849); Pterygosomatidae: G. pelaezi and G. texana; Trombiculidae: E. alfreddugesi) were collected on the 24 studied lizards (Table 1; Figure 1).
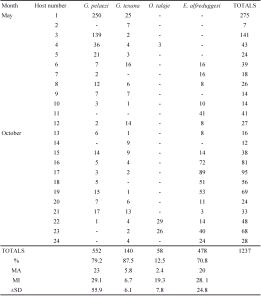
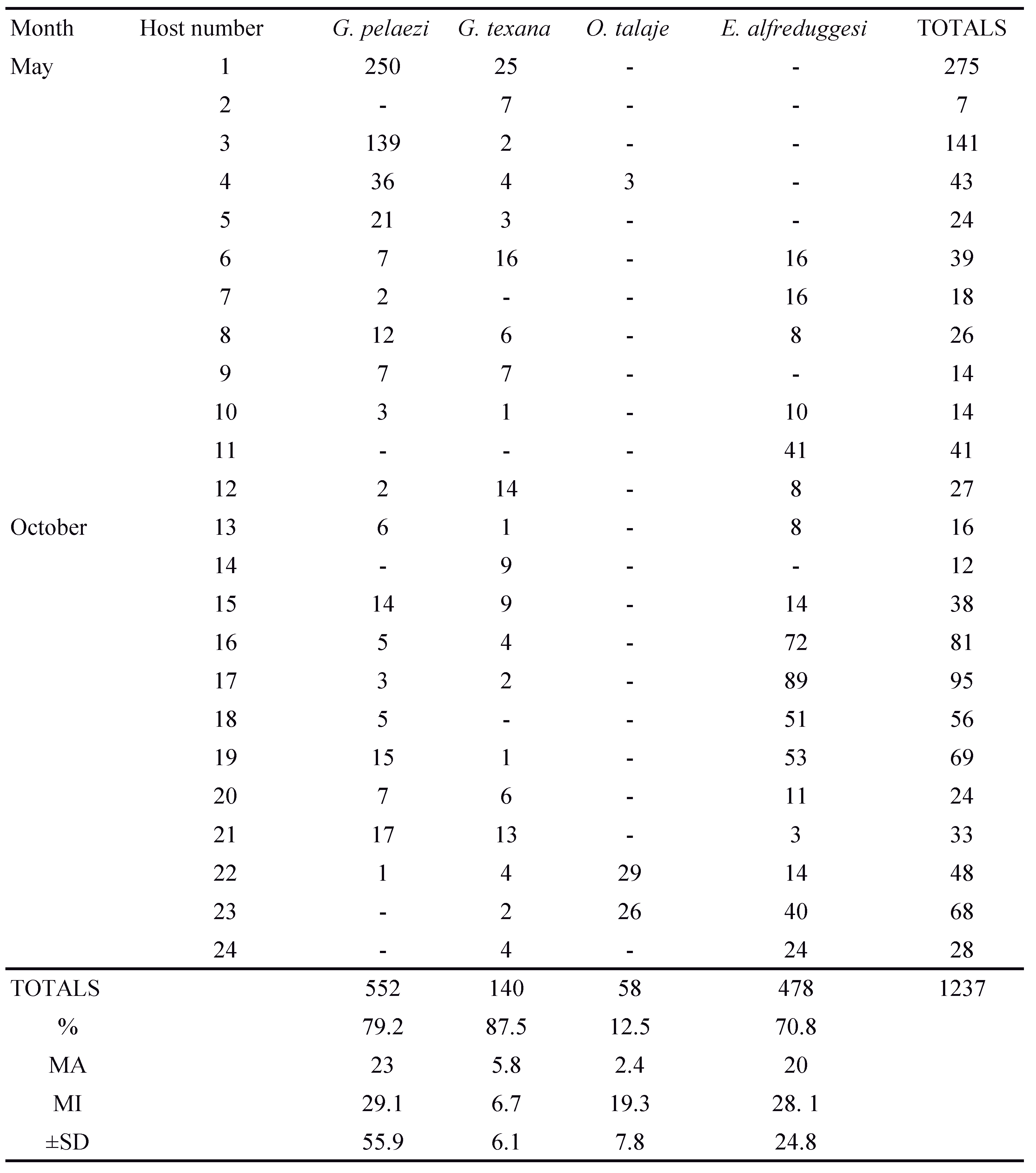


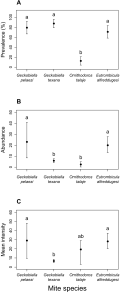
We detected significant differences among mite species in their prevalence, mean abundance, and mean intensity (Figure 2A, B and C). The prevalence of G. pelaezi, G. texana, and E. alfreddugesi was relatively high (>70% for these three mites) and statistically different from the prevalence of O. talaje (<20%, Figure 2A). We found the latter mite species on only 12.5% of the lizards. The highest abundances corresponded to G. pelaezi and E. alfreddugesi while the abundances of G. texana and O. talaje were significantly lower (Figure 2B). Finally, the lowest mean intensity corresponded to G. texana, and was significantly lower than the mean intensity of G. pelaezi and E. alfreddugesi (Figure 2C).
With the exception of O. talaje, species that constitute the acarofauna associated with S. torquatus had been previously registered in the same host and locality (Paredes-León et al. 2008; Montiel-Parra et al. 2009). The species of the family Pterygosomatidae that we collected are permanent parasites and specialists of lizards of the family Phrynosomatidae (Paredes-León et al. 2012). In contrast, the species of Trombiculidae is a protelean generalist mite that has been recorded parasitizing reptiles, birds and mammals (Hoffmann and López-Campos 2000; Rocha et al. 2008). The taxonomic determination of our material as O. talaje is based on the re-description of larvae reared from adults collected in Guatemala by Kohls et al. (1965); however, Venzal et al. (2008) pointed out that larvae collected by them near the type locality (Salamá, Baja Verapaz, Guatemala) differ morphologically from the ones collected in Mexico and southeastern United States. For this reason, in this study we keep the original name of the species until we have strong morphological and/or molecular evidence that suggests its inclusion into another.
The addition of O. talaje in the acarological record of S. torquatus increases to five the number of species associated with this host, four of them being collected in the very locality of our study (i.e., Pedregal de San Ángel). With this, the spiny collared lizard is the species of the genus Sceloporus with the highest richness of associated mites in Mexico, because in the 24 remaining species of the genus that have been studied from this perspective in the country, the richness value goes from one to four species, but collected in different localities (Paredes-León et al. 2008).
According to Cooley and Kohls (1944), rodents of the genera Dipodomys, Neotoma, and Rattus are frequent hosts of O. talaje; however this species has been recorded in a wide variety of hosts such as birds, serpents, domestic mammals, and even humans (Cooley and Kohls 1944). The first record of O. talaje parasitizing a reptile was made by Dunn (1933), who reported this mite on a rainbow boa Epicrates cenchria in a suburb of Panama City. In Mexican reptiles, this species has been previously recorded in three lizard species of the genus Phyllodactylus from Oaxaca (Paredes-León et al. 2008). The presence of this tick species in only three hosts in the REPSA could be explained by the occasional contact of the lizards with the nests of their primary host, i.e., rodents (Hoffmann and López-Campos 2000).
The acarofauna of S. torquatus is composed by two groups of species: permanent parasites (G. texana and G. pelaezi) and temporal parasites (E. alfreddugesi and O. talaje). The permanent infestation of pterygosomatid mites on the lizards is probably the main factor that determines the significant differences found in the prevalence levels of our sample between the pterygosomatid mites and O. talaje. On the contrary, the high observed prevalence of E. alfreddugesi, which is a temporal parasite of these lizards, may be explained by the fact that we conducted our sampling during the rainy season. In temperate zones, temperature seems to act as the main determining factor whereas the amount of precipitation is a more important cause in tropical regions (Sasa 1961). Thus, environmental humidity seems to favor the abundance of E. alfreddugesi, which is consistent with the general pattern observed in Trombiculidae. In this family, population numbers increase toward the higher levels of environmental humidity (Michener 1946). Particularly for E. alfreddugesi, Clopton and Gold (1993) demonstrated that larval population densities are greatest in areas of high relative humidity, moderate temperature, and low incident sunlight.
In addition, during the rainy season, females of S. torquatus undergo vitellogenesis, allocating energy and fat reserves to the production of oocytes (Feria-Ortiz et al. 2001). On the other hand, males of this species also carry out reproductive activities during rains, such as the establishment and defense of territories (Feria-Ortiz et al. 2001). These activities are costly and can negatively affect certain components of the lizards' immune system (Belliure et al. 2004), which apparently makes them particularly susceptible to the infestation by E. alfreddugesi during the wet season.
Interestingly, even though G. texana was found in almost 90% of the lizards that we examined, its abundance and mean intensity were significantly lower compared to those of G. pelaezi. These differences are difficult to explain based on the punctual sample that we made. However, we can hypothesize that factors such as host immune response, seasonal variations in the population levels of both mite species, host preference and even interspecific competition could be elements determining such differences.
The poor knowledge of mites associated with reptiles of Mexico (51 species associated with 92 reptile species according to Paredes-León et al. 2008) highlights the importance of continuing the inventory of this taxonomic group, considering the richness and endemism of reptiles in this country (864 species, see Flores-Villela and García-Vázquez (2014)). In parallel with the above, the knowledge of biological and ecological aspects of these arthropods must be increased, in order to achieve an integral understanding of this parasite-host relationship.
We thank Víctor Argaez, Israel Solano-Zavaleta and Abril Villagrán-Vázquez for their help in the collection of hosts and ectoparasites; Laura Del Castillo-Martínez for assisting in the mounting process of mites. Ana Isabel Bieler Antolín for the edition of the photomicrographs. Financial support for this study was obtained from the Dirección General de Asuntos del Personal Académico-Universidad Nacional Autónoma de México through the Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT, project IN203516).
Belliure J., Smith L., Sorci G. 2004. Effect of testosterone on T cell mediated immunity in two species of Mediterranean lacertid lizards. J. Exp. Zool. Part A: Ecological Genetics and Physiology. 301: 411¬-418. https://doi.org/10.1002/jez.a.20068 doi:10.1002/jez.a.20068 ![]()
Bush A.-O., Lafferty K.-D., Lotz J.-M., Shostak A.-W. 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol., 83: 575-583. http://www.jstor.org/stable/3284227
Cooley G.-M., Kohls R.-A. 1944. The Argasidae of North America, Central America and Cuba. Amer. Midl. Naturalist. Monogr., 1:1-152.
Clopton R.-E., Gold R.-E. 1993. Distribution and seasonal and diurnal activity patterns of Eutrombicula alfreddugesi (Acari: Trombiculidae) in a forest edge ecosystem. J. Med. Entomol., 30: 47-53. doi:10.1093/jmedent/30.1.47 ![]()
Dunn H.-L. 1933. Observations on the host selection of Ornithodoros talaje Guern., in Panama. Am. J. Trop. Med., 13: 475-483. doi:10.4269/ajtmh.1933.s1-13.475 ![]()
Feria-Ortiz M., Nieto-Montes de Oca A., Salgado-Ugarte I.-H. 2001. Diet and reproductive biology of the viviparous lizard Sceloporus torquatus torquatus (Squamata: Phrynosomatidae). J. Herpetol., 35: 104-112. http://www.jstor.org/stable/1566029 doi:10.2307/1566029 ![]()
Flores-Villela O., García-Vázquez U.-O. 2014. Biodiversidad de reptiles en México. Rev. Mex. Biodiv., 85: 467-475. https://doi.org/10.7550/rmb.43236 doi:10.7550/rmb.43236 ![]()
Hoffmann A. 1990. Los trombicúlidos de México (Acarida: Trombiculidae). Publicaciones Especiales del Instituto de Biología 2. Ciudad de México: Universidad Nacional Autónoma de México. pp. 275.
Hoffmann A., López-Campos G. 2000. Biodiversidad de los ácaros en México. Ciudad de México: Universidad Nacional Autónoma de México and Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. pp. 230.
Kohls G.-M., Sonenshine D.-E., Clifford C.-M. 1965. The systematics of the subfamily Ornithodorinae (Acarina: Argasidae). II. Identification of the larvae of theWestern Hemisphere and description of three new species. Ann. Entomol. Soc. Am., 58: 331-364. doi:10.1093/aesa/58.3.331 ![]()
MacGregor-Fors I., Payton M.-E. 2013. Contrasting diversity values: statistical inferences based on overlapping confidence intervals. PLoS ONE 8: e56794. https://doi.org/10.1371/journal.pone.0056794
Manly B.-F.-J. 2007. Randomization, bootstrap and Monte Carlo methods in biology. Boca Raton: Chapman and Hall/CRC. pp. 455.
Méndez de la Cruz F., Díaz de la Vega P.-A.-H., Jiménez A.-V.-H. 2009. Herpetofauna. In: Biodiversidad del Ecosistema del Pedregal del Pedregal de San Ángel. Lot A., Cano-Santana Z. (Eds). Ciudad de México: Universidad Nacional Autónoma de México. p. 243-260.
Michener C.-D. 1946. Observations on the habits and life history of a chigger mite, Eutrombicula batatas (Acarina: Trombiculinae). Ann. Entomol. Soc. Am., 39: 101-118. doi:10.1093/aesa/39.1.101 ![]()
Montiel-Parra G., Paredes-León R., Guzmán-Cornejo C., Hortelano-Moncada Y., Pérez T.-M. 2009. Ácaros asociados a vertebrados. In: Biodiversidad del Ecosistema del Pedregal del Pedregal de San Ángel. Lot A. Cano-Santana Z. (Eds). Ciudad de México: Universidad Nacional Autónoma de México. p. 385-393.
Paredes-León R., García-Prieto L., Guzmán-Cornejo C., León-Règagnon V., Pérez T.-M. 2008. Metazoan parasites of Mexican amphibians and reptiles. Zootaxa. 1904: 1-166.
Paredes-León R., Klompen H., Pérez T.-M. 2012. Systematic revision of the genera Geckobiella Hirst, 1917 and Histiella Berlese, 1920 (Acari: Prostigmata: Pterigosomatidae) with description of a new genus for American species parasites on geckos formely placed in Histiella. Zootaxa. 3510: 1-40.
Peralta-Higuera A., Prado-Molina J. 2009. Los límites y la cartografía. In: Biodiversidad del Ecosistema del Pedregal de San Ángel. Lot A., Cano-Santana Z. (Eds). Ciudad de México: Universidad Nacional Autónoma de México. p. 27-42. doi:10.1016/j.rmb.2015.04.024 ![]()
Rocha C.F.-D., Cunha-Barros M., Menezes V.-A., Fontes A.-F., Vrcibradic D., Van S.-M. 2008. Patterns of infestation by the trombiculid mite Eutrombicula alfreddugesi in four sympatric lizard species (genus Tropidurus) in northeastern Brazil. Parasite, 15: 131-136. doi:10.1051/parasite/2008152131 ![]()
Sasa M. 1961. Biology of chiggers. Annu. Rev. Entomol., 6:221-244. doi:10.1146/annurev.en.06.010161.001253 ![]()
Venzal J.-M., Estrada-Peña A., Mangold A.-J., González-Acuña D., Guglielmone A.-A. 2008. The Ornithodoros (Alectorobius) talaje species group (Acari: Ixodida: Argasidae): description of Ornithodoros (Alectorobius) rioplatensis n. sp. from southern South America. J. Med. Entomol. 45: 832-40. doi:10.1603/0022-2585(2008)45[832:TOATSG]2.0.CO;2 ![]()



2018-04-27
Date accepted:
2018-09-28
Date published:
2018-10-12
Edited by:
Sidorchuk, Ekaterina

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Guzmán-Cornejo, Carmen; García-Prieto, Luis and Zúñiga-Vega, J. Jaime
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)