



Soil mesostigmatid mites as a potential tool for bioindication concerning ecological status of forest
Călugăr, Adina1
1✉ Institute of Biological Research Iași, branch of N.I.R.D.B.S., Bucharest, Romania.
2Lascăr Catargi Street no. 47, 700107, Iași, Romania.
2018 - Volume: 58 Issue: Suppl pages: 18-24
https://doi.org/10.24349/acarologia/20184285Keywords
Abstract
The waters of the Danube River, after a course of over 2,860 km, flow into the Black Sea, forming one of the most beautiful deltas in Europe or maybe in the whole world. Before flows into the Black Sea, it splits into three branches – Chilia, Sulina, and Sfântu Gheorghe; along these many habitat types are formed: aquatic, lakes covered with flooded reed beds, flooded islets, flooded reeds and willows, riverine forest of willows and poplars, sandy and muddy beaches, wet meadows, arid meadows, salted meadows, sandy and rocky areas, steep banks and forests on high banks. All these make the Danube Delta a natural genetic bank with an incalculable value for global natural heritage. Although it is the newest territory of Europe, Danube Delta is a key centre for biodiversity in Europe, owing to the great abundance of plants, animals and microorganisms in the entire river basin. Unfortunately, there has been a tendency to over-exploit some of these natural resources lately. Pollution, damming, industries, agriculture, livestock, tourism and urban settlements have disrupted the fragile equilibrium of the deltaic biome. To avoid an irreparable disaster, the DDBR has been listed as a wetland of international importance under the Ramsar Convention since September 1990, and since December 1990 it has been included as a strictly protected area in the World Heritage List under the World Heritage Convention.
This study is part of a wider investigation dedicated to the edaphic mesofauna on the whole, in order to evaluate the functional state and the intensity of human pressure exerted on forest ecosystems of Danube Delta Reserve Biosphere. Soil microarthropods with special remarks referring to Oribatida and Trombidiformes were investigated in the same sites as in the current study (Călugăr and Ivan 2016). The aim of the present investigation was to analyse the fauna composition of edaphic Mesostigmata in order to characterize the conservation status of the chosen forests. Mesostigmata mites are considered as bioindicators for several reasons: their functional role in the soil ecosystems as predators, high species richness of their communities, their relative high abundances and their robustness towards sampling and extraction methods (Koehler 1999). Predators are more active and respond more sensitively over shorter periods of time to environmental influences in soil ecosystems than other soil arthropods (Karg and Freier 1995, Koehler 1999).
In previous studies on Acari, in the forest habitats from Danube Delta, 140 species, 95 genera and 52 families have been identified. Among the species reported here numerous are new for the Romanian fauna and some faunal elements are present only or predominantly in the Danube Delta and Dobrogea’s territory. From Mesostigmata, the faunistic material amounted to a total of 60 species, 29 of them were identified as belonging to Uropodina from different types of delta ecosystems while the rest belonging to Gamasina was found only from forest habitats. (Feider and Huţu 1972, Huţu 1993, Huţu and Călugăr 1994, Ivan et al. 2006, Călugăr 2008).
The study was carried out in two different types of forest ecosystems: natural forests (Letea -Hăşmacul Mare, Chilia Veche, Plauru I) and forest plantations (Plauru II) placed on the DDBR territory (Table 1).


Series of five samples with a surface of 100 cm2, with a depth of 5cm each, consisting in soil and litter (litter and fermentation layer – olf and humification layer – ah) were taken in July 2015 from each stand. Berlese funnels were used to extract the edaphic mesofauna and then Mesostigmata mites were separated from the rest of the collected fauna by means of a stereomicroscope. The identification of the adults and some preadults was performed at species level. The analysis of the Mesostigmata communities structure was based on the analytical ecological indices (average abundance of each species expressed individuals/m2 – ā, frequency – C%, relative density – D.r.% (Lions 1975) and synthetic indices (the index of ecological significance – W%. The frequecy (C) is expressed in the classes: I (accidental, C under 5%), II (accessory, C between 25.1–50%), III (constant, C between 50.1–70%), IV (euconstant, C over 75% ). The relative density (D.r.) is expressed in the classes: I (subrecedent, D.r. under 1%), II (recedent, D.r. between 1.1–2%), III (subdominant, D.r. between 2.1 – 5%), IV (dominant, D.r. between 5.1–10%), and V (eudominant, D.r. over 10%). The index of ecological significance (W) is also expressed in the classes: I, II (accompanying species, W under 1%), III (influential species, W between 1.1–5%), V, IV (edifying species, W over 5%) (Dziuba 1968, after Stan 1995).
The global characterization of the communities also used the following parameters: global average abundance (Ā), expressed as individuals/m2, standard deviation and Pearson coefficient of variation (cv%), number of species (S), and the average number of species/sample (S'), expressing the richness in species and their horizontal distribution. For the vertical distribution analysis, the percentage of individuals identified in the litter and fermentation layer compared to the one identified in depth layer was calculated. The adults/preadults ratio (A/P – expressed as an absolute value) providing information on the demographic structure of the studied communities was also calculated.
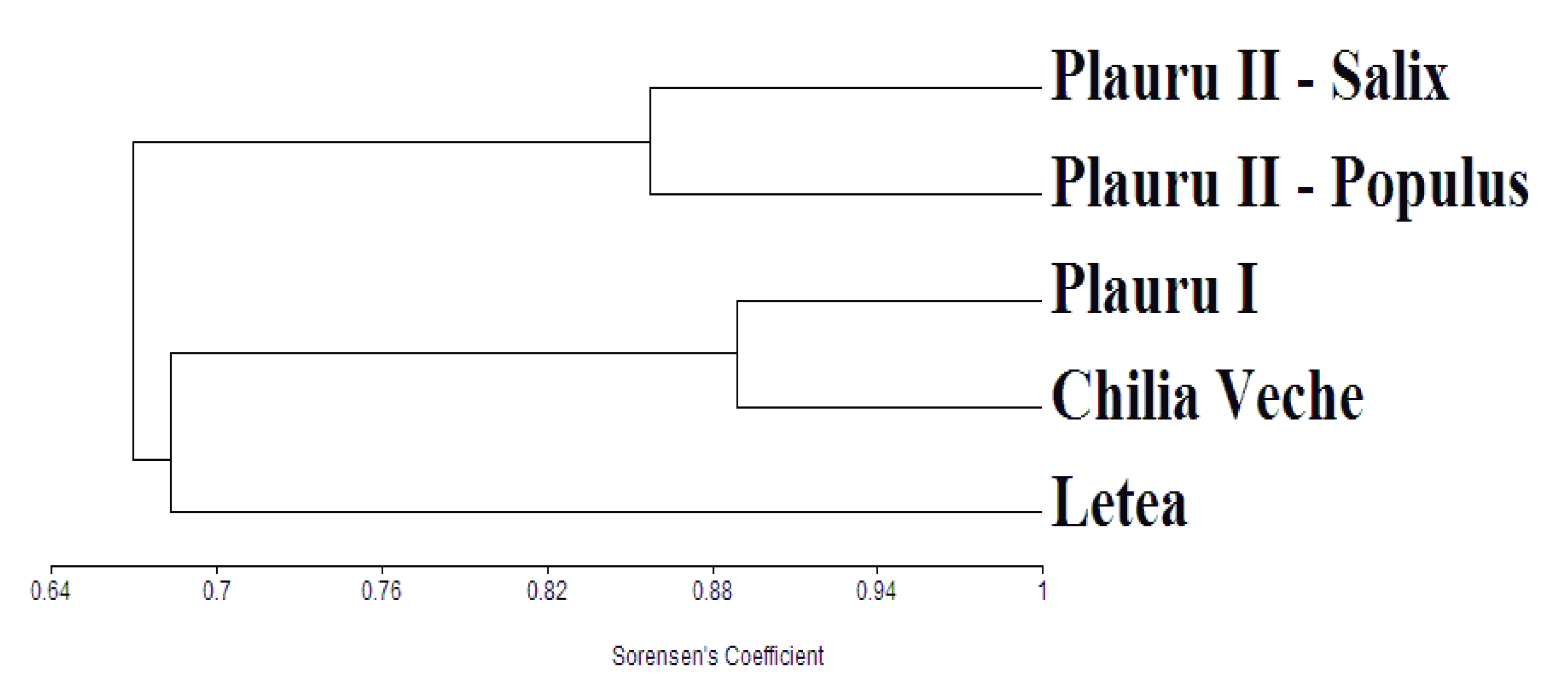
The resemblance degree of Mesostigmata communities from the 5 ecosystems was tested by Cluster analysis (UPGMA method) using Sorensen’s coefficient of similarity. All statistical calculations for this research were done in MVSP 3.2 software.
In the present study we identified mites belonging to the three cohorts of the Mesostigmata: Antennophorina, Uropodina and Gamasina (Krantz and Walter 2009).
Antennophorina included only one family – Celaenopsidae, represented by a single species (Celaenopsis badius), present only in the forest soil from Chilia Veche. About 5.75% of all encountered Mesostigmata were Uropodina. Within Gamasina, 31 species belonging to 21 genera and 10 families were identified. Among these the best represented families were Ascidae with 7 genera and 9 species, followed by Hypoaspididae with 2 genera and 6 species and Parasitidae respectively Phytoseiidae with 3 genera and 3 species each.
The average density ranged between 820 (Salix plantation) and 2280 individuals/m2 (Plauru I) and the number of species varied between 4 (Salix plantation) and 16 species (Letea forest) (Table 2). Analysis of spatial distribution of the species in all studied cases revealed big differences between S and S’ which points to an uneven distribution of the species. The highest value of coefficient of variation was recorded at Plauru II in the Salix alba plantation and the smallest at Plauru I. The vertical distribution indicates a mainly populating of the humification layer (ah), with 73-100% of the total number of individuals. In all studied sites the migration in the depth is a consequence of the dryness which characterized the sampling period (Table 2).


Only three species are common to the majority of the investigated sites: Hypoaspis nolli, Rhodacarellus silesiacus and Ololaelaps placentula; H. nolli is an edifying species for the mesostigmatid mites communities in all studied sites. R. silesiacus and O. placentula are absent only in the plantation of Salix. The first one is an accompanying species at Letea forest and an edifying one in the rest of the sites. O. placentula is influential at Plauru I and it is accompanying in the rest. Some species present in only one stand were highlighted: 11 species at Letea, 2 at Chilia Veche, 7 at Plauru I, 4 at Plauru II in Populus x canadensis plantation and only one at Plauru II in Salix alba plantation. Among the investigated sites the most equilibrated community was that of Plauru II with almost 50% accompanying species, 16.7% influential species and 33.4% edifying species (Table 2). The demographic structure of the communities was analysed using numerical ratio adults/preadults, with values between 1.9–8.4 (Table 2).
The results of a previous research targeted on the edaphic mesofauna from the same forest sites indicated that Mesostigmata mites were ranging between 2.7% and 10.02% of the total Acari. They hold either second place after Oribatida (Letea and Plauru II Salix plantation), or the third one after Oribatida and Trombidiformes (in the rest of the sites) (Călugăr and Ivan 2016). Studies conducted in other wetland forests showed that oribatid mites prevail within the mites fauna and gamasids are the next ones, both in forests and plantations (Călugăr 2006). Results of the researches regarding the Mesostigmata mites from forest ecosystems in DDBR carried out in 2006 pointed out the human influence on the biodiversity of these mites. The most diverse fauna was identified in the natural forest soils and the human influence in the anthropized forests and plantations was evident by the reduction of the number of species. Almost 70% of the identified species was found in the natural forests, 16% in the anthropized forests from the embanked enclosure and about 48% in the forest plantations; about 42% were identified exclusively in natural forests and 23% in plantations, a percentage of 23% being common to natural forests and plantations (Ivan et al. 2006).
The results of the present investigation showed that the highest number of species and individuals was also obtained in the soil of the natural forests and the lowest ones in Salix alba plantation. The global average abundance, as well as the number of species have ample variations from one stand to another, both in the case of natural forests and plantations, mainly driven by bio-edaphic conditions specific to each forest stand. In the case of oribatid mites it was shown that horizontal distribution of the individuals illustrated by Pearson variation coefficient of abundance is more uniform in plantations than in natural forests where litter is more diverse and spread mosaic – like (Călugăr and Ivan 2016). In Mesostigmata such a demarcation was not established. Both in natural forests and plantations, the Pearson variation coefficient of abundance had high values with one exception – natural forest from Plauru I. So, only here the distribution of the Mesostigmata individuals was uniform while in the rest of the ecosystems, both forests and plantations, the distribution was an aggregate type. In the case of mesostigmatid mites the horizontal distribution pattern is probably linked to the distribution of the groups of edaphic fauna which constitute their preferred prey.
Cluster analysis indicates that natural ecosystems from Plauru and Chilia have common characteristics in terms of faunistic spectrum of Mesostigmata mites, as well as the two plantations from Plauru, which also have a high degree of similarity. In a lower proportion Letea forest have common features with the other two natural forests. Also, we noticed a low degree of similarity between natural forests and plantations, as it can be observed in the figure 1 with two distinctive clusters (sites 1, 2 and 3 versus 4 and 5).

Among the natural forests, Letea displayed the highest number of identified species, which was actually expected since this is a strictly protected area. This forest that is considered the most northern subtropical forest in the world is the oldest natural reserve in Romania, being declared a nature reserve since 1938. This was the nucleus of the Danube Delta Biosphere Reserve which today is a World Heritage Site. However, in the last period this forest, although it is fenced, is affected by some impact factors especially by the approximately 2000 horses abandoned by local people, which are deemed to be a threat to the flora of the forest, including to some plants on the IUCN Red List of Threatened Species. Compared with the other natural forest – Plauru I – where the highest value of density was found, at Letea the abundances of the Mesostigmata mites is about 1.4 times lower. Several authors (Chachaj and Seniczak 2005, Battigelli et al. 2003) remarked in their studies developed in pastures that grazing leads to lower densities of some groups of mites, mostly Oribatida and Prostigmata, but only at high rates of grazing and especially of cattle. At Letea forest the horses’ influence on the edaphic fauna by grazing is possible, but the decreased density of mesostigmatid mites compared with Plauru I is probably due to the particular pedological conditions, this forest being developed on Letea Sand Bank and growing in narrow stripes between the dunes. It is noteworthy that the uropodid mites which are considered particularly sensitive and thus good bioindicators of environmental changes (Athias-Binche1981, Karg 1989, Gulvik 2007) were present only in natural forests, the most abundant being in Letea Reserve.
In the context of the present study three new species for the Romanian fauna from the genus Cheiroseius were identified: C. nepalensis (Evans and Hyat 1960), C. borealis (Berlese 1904) and C. mutilus (Berlese 1916). This genus seems to be perfectly adapted to delta's habitats, as literature data recorded that Cheiroseius species are abundant in moist soils and in wet forest litter, especially along lake and stream margins. Many species appear to be subaquatic and able to tolerate flooding (Krantz and Walter 2009, Karg 1993). Moreover, the presence of many rare, less cited species like Celaenopsis badius of the Antennophorina cohort and Epicriopsis palustris, Arctoseius insularis, Paragarmania dendriticus, Cheiroseius viduus, Eviphis ostrinus, Antennoseius sp. was noted for Gamasina. Among them, the first three ones were identified before from DDBR: from Letea bank (A. insularis, C. badius) and from Letea – Hăşmacul Mare forest (E. palustris); in addition, another 5 species were found previously in DDBR: Hypoaspis nolli, Ololaelaps placentula, H. aculeifer, Veigaia nemorensis, H. vacua (Table 2) (Ivan et al. 2006, Călugăr 2006). In terms of environmental preferences it is interesting to note that over 40% of the species identified in the studied sites are hygrophillic or meso-hygrophillic elements which prove the bioindicator potential of these soil mites. Beside the hygrophillic E. palustris, A. insularis, C. viduus and hygromesophilic E. ostrinus, some ubiquitous species were identified too (V. nemorensis, H. aculeifer, H. vacua) (Karg 1993, Salmane 1999).
The species O. placentula and R. silesiacus common for four sites have the same requirements to the humidity factor (Table 2, 3). After Salmane (1999), both are hygromesophillic species and after Karg (1993), R. silesiacus is hygromesophilic and O. placentula euryhygrophilic, but certainly both are well adapted to the peculiar conditions of the Danube Delta. Hypoaspis nolli – the edifying species in all investigated sites – is characteristic to wet meadows and considered to be flood resistant (Salmane 1999, Griegel 2008). Its development in all investigated sites is presumably owing to similar biopedoclimatic conditions or to trophic resources which coincide (Table 2, 3).


All these results are encouraging and further research that will be made in the frame of the BIODIVERS Research Programme will include investigations into the dynamics and some other forests as well as other types of ecosystems; thus the obtained results will constitute an important database on the diversity and density of soil Mesostigmata in the DDBR.
This study is part of a wider investigation dedicated to the edaphic mesofauna on the whole and conducted within the BIODIVERS Research Programme, funded by the Romanian Ministry of Education and Scientific Research.
Athias-Binche F. 1981. Écologie des Uropodides edaphiques (Arachnides: Parasitiformes) de trois écosystèmes forestiers. 1. Introduction, matériel, biologie. Vie et milieu, 31: 137-147.
Battigelli J., McIntyre G. S., Broersma K., Krzic M. 2003. Impact of cattle grazing on prostigmatid mite densities in grassland soils of southern interior British Columbia. Can. J. Soil Sci., 83(5): 533-535. doi:10.4141/S02-064 ![]()
Călugăr A. 2006. Qualitative and quantitative data concerning the edaphic mesofauna in the forest ecosystems from the middle section of the Prut. Sc. Ann. of DDI, 12: 7-12.
Călugăr A. 2008. New and rare species of mites (Acari: Gamasina) from some forest ecosystems of the Danube Delta Biosphere Reserve. In: Bertrand N., Kreiter K., McCoy K. D. (Eds). Integrative Acarology; Montpellier: Euraac. p. 167-175.
Călugăr A., Ivan O. 2016. Soil microarthropods and their bioindicator value regarding the bio-edaphic conditions in forest ecosystems of Danube Delta. St. Univ. "V. Goldiș", Șt. Vieții, 26 (2): 215-219.
Chachaj B., Seniczak S. 2005. The influence of sheep, cattle and horse grazing on soil mites (Acari) of Lowland Meadows. Folia biol. (Kraków), 53 (Suppl.): 127-132. doi:10.3409/173491605775789362 ![]()
Griegel A. 2008. Effects of the summer flood 1997 on the collembolan and gamasid fauna in a Lower Oder Valley floodplain, Peckiana, 5: 105-114.
Gulvik M. 2007. Mites (Acari) as indicators of soil biodiversity and land use monitoring: a review. Polish J. Ecol., 55 (3): 415-440.
Ivan, O., Călugăr, A., Vasiliu, N. 2006. A survey of the edaphic mites fauna (Acari: Oribatida, Gamasina) from the main types of forest ecosystems in the Danube Delta Biosphere Reserve. Sc. Annals of D. D. I., 12: 45-54.
Karg W. 1989. Acari (Acarina), Milben Unterordnung Parasitiformes (Anactinochaeta) Uropodina Kramer, Schildkrötenmilben.. Jena: Gustav Fischer Verlag. pp 203.
Karg, W. 1993. Acari (Acarina), Milben. Parasitiformes (Anactinochaeta), Cohors Gamasina Leach, Raubmilben. Tierwelt Deutschlands, 59.pp 523.
Krantz G. W., Walter D. E. 2009. A Manual of Acarology. Lubbock, Texas:Texas Tech University Press. pp. 807.
Koehler H. H. 1999. Predatory mites (Gamasina, Mesostigmata). Agriculture, Ecosystems and Environment, 74: 395–410. doi:10.1016/S0167-8809(99)00045-6 ![]()
Lions J. C. 1989. Application du concept de la diversité spécifique à la dynamique de trois population d'oribates (acariens) de la fôret de la Saint Baume (Var). Ecol. Mediterranéenne, 165-192.
Salmane I. 1999. Soil free-living predatory Gamasina mites (Acari, Mesostigmata) from the coastal meadows of Riga Gulf, Latvia. Latvijas Entomologs, 37: 104-114.
Stan Gh. 1995. Statistical methods with application in entomological researches (VII) (in Romanian). Bull. inf. Soc. lepid. rom., 6 (1-2): 67-96.



2016-11-03
Date accepted:
2018-02-19
Date published:
2018-09-28
Edited by:
Roy, Lise

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Călugăr, Adina
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)