



Distribution of microarthropods across altitude and aspect in the sub-Antarctic: climate change implications for an isolated oceanic island
Hugo-Coetzee, Elizabeth A.1 and Le Roux, Peter C.2
1✉ National Museum, P.O. Box 266; Department of Zoology and Entomology, University of the Free State, Bloemfontein, 9301, University of Stellenbosch, Stellenbosch, 7602, South Africa.
2Department of Plant and Soil Sciences, University of Pretoria, Pretoria, 0083, South Africa.
2018 - Volume: 58 Issue: Suppl pages: 43-60
https://doi.org/10.24349/acarologia/20184278Keywords
Abstract
Current climate change is unequivocally altering species distributions (Walther et al. 2002; Root et al. 2003; Parmesan 2006), with many species' ranges shifting poleward and upward in elevation (Fielding et al. 1999; Parmesan et al. 1999; Parmesan and Yohe 2003; Walther et al. 2002; Chen et al. 2011). Changes in species ranges will likely lead to local extinctions (e.g. loss of lowland populations) and, because species tend to respond individualistically to changing climatic conditions, to the alteration of community structure (Root et al. 2003; Thomas et al. 2004; Parmesan 2006; Le Roux and McGeoch 2008c; Chen et al. 2011). Knowledge of the current regional and local distributions of species, as well as continued monitoring of species distributions, is therefore critically important for understanding and predicting the impacts of climate change on individual species and entire communities (Heaney 2001; Peterson et al. 2002; Walther et al. 2002; Sanders et al. 2003; Thomas et al. 2004).
One approach to predicting species future distributions is to examine the spatial distribution of species across an environmental gradient, such as, for example, the temperature gradient found across altitude (Parmesan et al. 2005). Gradients can be seen as natural experiments (Smith et al. 2002; Brehm and Fiedler 2003), offering an analogue for future climates (Hodkinson and Bird 1998; Fielding et al. 1999; Hodkinson and Wookey 1999). For example, the current occurrence pattern of a species at cold, high altitudes is expected to change with increasing temperature to resemble the species' distribution currently observed at warmer, lower altitudes. Abiotic factors change rapidly from high to low altitudes (e.g. temperature, humidity and wind speed), so that species responses to these factors can be assessed across a relatively small geographic area (Peterson et al. 1997). However, altitudinal gradients may not always be direct analogues of changing climates, since other factors, such as habitat type, local wind patterns and topography may vary independently of the climatic variable(s) of interest (Parmesan et al. 2005).
Another approach to understand the likely impacts of climate change is manipulative experiments, in which environmental conditions can be controlled to isolate a single or few variables of interest (Callaghan and Jonasson 1995; Hodkinson and Wookey 1999; Dunne et al. 2004; Parmesan et al. 2005). However, short-term experiments may not reveal realistic species responses to climate change as biota can have delayed responses to experimental treatments or even different initial responses to short-term manipulations compared with responses observed when the manipulation is sustained over longer periods (Chapin et al. 1995; Dunne et al. 2004; Epstein et al. 2004). As a result, the integration of experiments and gradient studies may provide the most robust approach to predicting the impact of climate change on species distributions (Hodkinson and Wookey 1999; Dunne et al. 2004; Parmesan et al. 2005).
Sub-Antarctic islands are ideal for gradient and experimental studies to examine the likely consequences of climate change on species assemblages due to the occurrence of steep abiotic gradients and the islands’ relative isolation from other anthropogenic influences (see Vitousek 2002). Only a few gradient studies (Davies and Melbourne 1999, Tweedie 2000 – Macquarie Island; le Roux and McGeoch 2008b, Chown et al. 2013 – Marion Island) and experimental studies (Le Roux et al. 2005, McGeoch et al. 2006 – Marion Island; Bokhorst et al. 2007 – Falkland Islands) have been conducted. These islands rise from sea level to high altitudes in the span of only a few kilometres (Huntley 1972), presenting steep temperature and moisture gradients over relatively small distances. Furthermore, ecosystems on sub-Antarctic islands are remote, relatively closed systems with a harsh environment and fairly low species richness (Smith and Steenkamp 1990; Chown and Froneman 2008). One such sub-Antarctic island is Marion Island (Prince Edward Island group), situated halfway between Africa and Antarctica (Chown and Froneman 2008).
High latitude areas have experienced, and are expected to continue to experience, substantial climate changes (IPCC 2001). On Marion Island large changes in climate have been observed over the past 50 years, with a 1.5 ºC increase in mean annual air temperature, a 1.4 ºC increase in sea temperature, an 800+ mm decrease in mean annual precipitation, and higher mean wind speed (Smith 2002; Melice et al. 2003; Lutjeharms and Ansorge 2008; Le Roux and McGeoch 2008a). The number and size of rainfall events also declined, resulting in longer dry spells (Le Roux and McGeoch 2008a) which might have even stronger impacts on organisms than the decline in mean annual rainfall (Weltzin et al. 2003). Indeed, there is already evidence of considerable impacts of climate change on the distribution of biota on and around Marion Island. For example, the island's vascular flora has expanded to higher altitudes, (Le Roux and McGeoch 2008c), and surface nesting seabird abundance has declined due to lower prey availability in the ocean (Crawford and Cooper 2003; Crawford et al. 2003; Pakhomov et al. 2004; Ryan and Bester 2008).
Microarthropods (mites and springtails) are, together with other macro-invertebrates, the main detritivores on sub-Antarctic islands and are essential for nutrient cycling and primary production in terrestrial ecosystems (Burger 1985; Smith and Steenkamp 1990; Chown et al. 2002). These microarthropods may, though, be sensitive to changes in climate. Indeed, a climate change experiment investigating the impacts of declining rainfall and warming temperatures on microarthropods inhabiting a keystone plant species (Azorella selago) found large changes in abundance and biomass after only one year (McGeoch et al. 2006). Experiments in the Arctic and Antarctic show similar results, possibly indicating that moisture, at least for some species, is a more important determinant of species distributions in the region than temperature (Strathdee and Bale 1998; Sinclair et al. 2003; Leinaas and Sømme 1984; Leinaas and Fjellberg 1985; Convey et al. 2002; Convey et al. 2003). Additionally, the Antarctic experiments revealed a general decrease in microarthropod (especially springtail) abundance with warming (Convey 2001; Convey et al. 2002; Hayward et al. 2003). Therefore, a warmer and drier climate is likely to alter microarthropod community structure (Bergström and Chown 1999; McGeoch et al. 2006), and a better understanding of current spatial patterns of microarthropods and the possible future distributions as a consequence of climate change are therefore critical.
Therefore, in this study we examine the distribution of microarthropods from the cushion plant, Azorella selago (Apiaceae) along a temperature gradient (i.e. across altitude) and between the drier, eastern and wetter, western aspects of Marion Island. We then compare the results from this study to observations from related climate change experiments to predict the likely impact of climate change on species distributions on Marion Island (following Dunne et al. 2004). Since species might not only respond to environmental factors, but also to specific habitat variables, such as cushion size, the relationships between species richness, abundance and community structure and A. selago plant characteristics were also examined. In this study we investigated 1) the relationships between microarthropod species richness, abundance and community structure and altitude and aspect of the island as well as plant characteristics, and 2) how individual species respond to altitude and aspect.
Marion Island (46º54’S; 37º45’E) lies about 1770 km south of Africa, with a surface area of 290 km², which rises to an altitude of 1230 m a.s.l. (Chown and Froneman 2008). The climate is hyperoceanic, with a mean annual temperature of 6.4 ºC, mean annual rainfall of approximately 1900 mm (as measured at the meteorological station on the north-eastern coast of Marion Island), a high degree of cloudiness and strong westerly winds (Schulze 1971; Smith 2002; Le Roux and McGeoch 2008a). Mean air, soil and within-plant temperatures decrease with an increase in altitude (Blake 1996; Nyakatya and McGeoch 2008; Table 1), with an air temperature lapse rate of approximately 4.0 ºC per 1000 m proposed for Marion Island (Klok and Chown 2003; supported by soil temperature measurements: Nyakatya and McGeoch 2008, Table 1). Wind speed is highest at high altitudes and rainfall peaks at mid-altitudes (Blake 1996). Freeze-thaw events are more frequent at mid-altitudes, but at high altitudes these events occur for longer periods with deeper soil freezing (Boelhouwers et al. 2003; Le Roux and McGeoch 2008a). Additionally, the western aspect of the island is colder than the eastern aspect, at least in summer (Table 1), and receives more rain and more consistent wind due to its exposure to the dominant westerly winds (Schulze 1971; Gremmen 1981; Le Roux and McGeoch 2008a).


The plant Azorella selago (Apiaceae) occurs from sea level to about 650 m a.s.l. (highest record 840 m) on Marion Island (Phiri et al. 2015). It exhibits a cushion growth form, with a compact, prostrate hemispherical form, varying in shape, size and stem density (Hugo et al. 2004; Phiri et al. 2015). Azorella selago plants are thought to present a more favourable habitat in terms of microclimate and nutrients for microarthropods (Acari—mites and Collembola—springtails) than the fellfield soils in which it grows as microarthropod densities are higher in plants than in the surrounding epilithic biotope (Barendse and Chown 2001). Azorella selago also provides a favourable habitat for some vascular plants (le Roux and McGeoch 2008b; 2010; le Roux et al. 2013). The grass Agrostis magellanica (Lam.) Vahl. is particularly favourably impacted by A. selago, with the species exhibiting higher abundance, cover, biomass and reproductive effort when rooted in A. selago than when growing in the adjacent soil. Agrostis magellanica is the vascular plant that most commonly grows on A. selago from about 100 to 500 m a.s.l. (Gremmen 1981).
Twelve plots consisting of 36 Azorella selago plants (maximum diameters > 0.15 m) each were selected at three altitudes in four different areas of Marion Island (see Nyakatya and McGeoch 2008 and Appendix 1 for location details). All plots were located within fellfield vegetation (i.e. dominated by A. selago, with a total vascular plant cover < 50 %). Plants within plots may be considered independent as neither plant or microarthropod community variables were found to be significantly nor consistently autocorrelated at this scale (Hugo 2006). The low altitude plots were located between 150 and 250 m a.s.l, middle altitude plots between 350 and 450 m a.s.l and high altitude plots between 500 and 600 m a.s.l.. Sets of plots (high, middle and low altitude in each area) comprised transects. Two transects were on the eastern aspect (Tafelberg and Stony Ridge) and two were situated on the western aspect (Mixed Pickle and Swartkops) of the island. Each A. selago plant was digitally photographed from above prior to sampling. Image analyses (software: SigmaScan Pro 5.0, Illinois, USA) were conducted to determine cushion size (surface area; m²), percentage of dead tissue (brown or black stem or leaf areas on the plant surface) and percentage of the plant surface covered by the grass A. magellanica.
Microarthropods were sampled between November 2002 and February 2003, coinciding with peak abundances observed during summer (Barendse and Chown 2001). Sampling was alternated between east and west transects, finishing one transect (high, middle, low) at a time. Sampling was conducted using an O’Connor split corer (inside diameter of core = 70 mm, height = 70 mm, surface area = 0.0039 m²). Core samples were taken from all 36 plants in each plot (except in the Mixed Pickle high altitude plot where one plant was too small to sample), giving a total of 431 cores. The microarthropods were extracted from the cores using a MacFadyen high-gradient extraction protocol of two days at 25 ºC and two days at 30 ºC (Southwood 1978; Hugo et al. 2004). Microarthropod specimens were identified to the lowest possible taxonomic level (usually species), counted and stored in 99 % ethanol at the University of Stellenbosch. Species richness, abundance and community structure analyses, as well as numbers presented in the tables were based on individuals per core sample.
Generalized linear models (McCullagh and Nelder 1989) were used to determine the relationship between microarthropod species richness and abundance (separate models for each of these dependent variables) in the Azorella selago plants and explanatory variables. Explanatory variables included were the aspect of the island (east, west), altitude (low, middle, high), plant surface area, plant height, and percentage of dead stems. In addition, separate models for species richness and abundance with aspect, altitude, surface area, plant height, percentage dead and percentage grass cover were constructed for the middle and low altitude plots because of the scarcity of grass at high altitude plots. A Poisson error distribution was assumed for species richness and abundance, a logarithmic link function was implemented, and deviance used as a measure of goodness of fit (where necessary, overdispersion was accounted for during the analysis, Dobson 2002). Weighted marginal means and their 95 % confidence intervals were used to establish differences between means (non-overlapping confidence limits were interpreted as weighted means significantly different at P < 0.05, Sokal and Rohlf 1995).
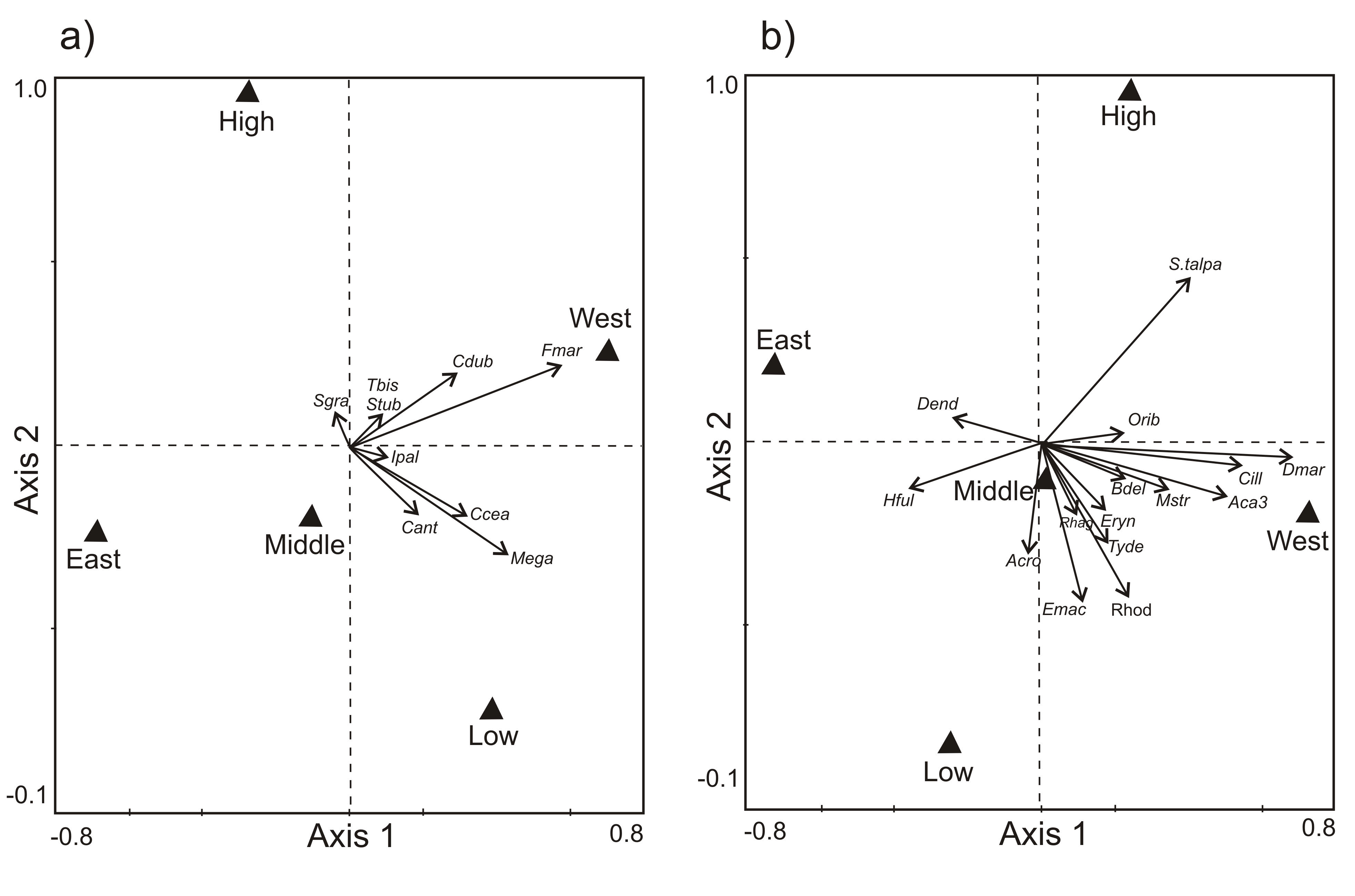
Redundancy Analysis was conducted to examine differences in microarthropod community structure between aspect and altitudes, and the responses of individual species to these environmental gradients (CANOCO v4.5; Ter Braak and Šmilauer 2002; Lepš and Šmilauer 2003). Redundancy Analysis (RDA) was chosen after examining the gradient lengths of species in the environmental space following Detrended Correspondence Analysis (DCA) (Jongman et al. 1995). RDA analysis was performed separately on log-transformed springtail and mite species data (with post-transformation of species scores and species centering) and displayed in ordination biplots. Springtails and mites were examined separately because of their different responses to environmental variables (McGeoch et al. 2006). Also, the eigenvalues for the first two axes were used to denote how much of the total variance in the species data was explained by the environmental variables (Lepš and Šmilauer 2003). Mite species that had > 5 % of their variation explained by the first ordination axis were included, therefore, excluding several species including the dominant Eupodes minutus.
A total of 31 (22 mite and 9 springtail) species and 115'112 individuals were collected across the 12 sample plots (Appendix 3). The Mixed Pickle middle altitude plot had the most species (30 species), and the low altitude plot along this same transect the highest mean abundance per sample (349 individuals/sample) (Table 1). The lowest species richness was found at the Tafelberg middle altitude plot (22 species) and the lowest mean abundance at Stony Ridge middle plot (172 individuals/sample). The mite species Eupodes minutus constituted 40% of all individuals sampled and 76% of all mite individuals (80% of mites on the eastern and 73% on the western aspect). The most abundant springtail species was Folsomotoma marionensis, which comprised 35% of all springtail individuals (22% of springtails on the eastern and 41% on the western aspect).
Island aspect and/or altitude contributed significantly to explaining mite and springtail species richness and abundance in all models (Table 2, Appendix 2). In contrast, plant characteristics were only significantly related to species richness and abundances in a few models (Table 2, Appendix 2). Mite and springtail species richness, and springtail abundance were significantly higher on the western aspect of the island and significantly lower at high altitudes (Figure 1). Microarthropod abundance declined monotonically with altitude, although not significantly (Figure 1, Table 1). The effect of altitude on species richness differed between the eastern and western aspects (i.e. there was a significant interaction between island aspect and altitude for mite and springtail species richness; Table 2, Appendix 2), with mite and springtail species richness peaking at mid altitude on the west. In contrast, on the eastern aspect the mid-altitude plots had the lowest species richness (Figure 1). Plant characteristics were not consistently related to microarthropod richness and abundance (Table 2, Appendix 2). Cushion area and height were significantly related to microarthropod abundance and richness in some analyses, but the impact of larger plants was not consistently positive or negative. Similarly, the cover of A. magellanica growing on A. selago was significantly related to the abundance of springtails and mites, but with contrasting effects on the two species groups (Appendix 2).

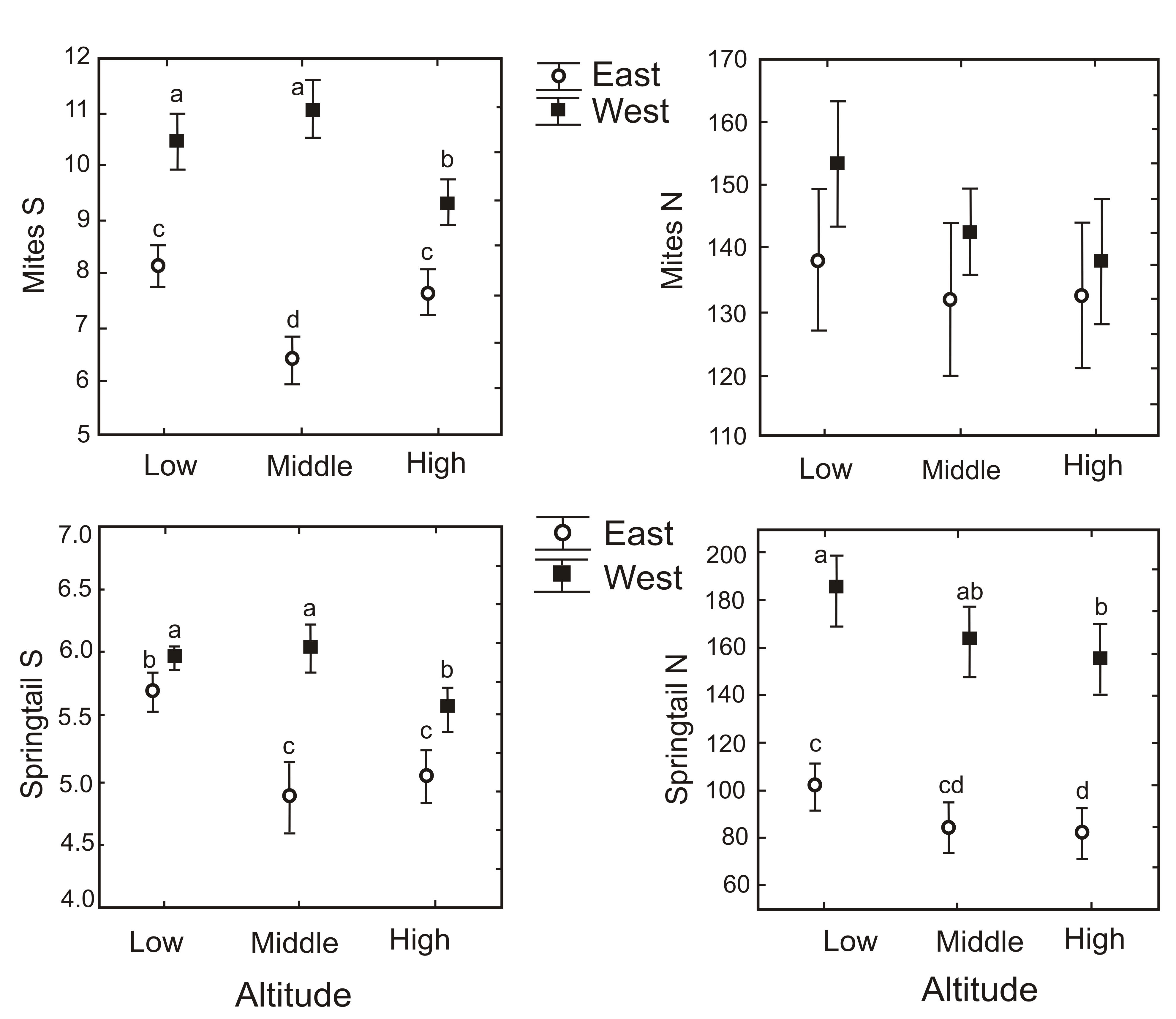


Community structure of springtails (Figure 2a) and mites (Figure 2b) also corresponded strongly to altitude and aspect of the island, with the first axis separating the eastern and western aspect and the second axis distinguishing the high altitude from the middle and low altitude. Eight of the nine springtail species were more abundant in the western plots, while Mucrosomia caeca and Megalothorax minimus were also more abundant at low altitudes (Figure 2a). Only two mite species (Halozetes fulvus, Dendrolaelaps sp.) were more abundant in communities in the eastern plots, and one species (Schwiebea talpa) was more abundant in high altitude plots (Figure 2b).

Microarthropod species richness, abundance and community structure were strongly related to aspect and altitude, with generally higher richness and abundance on the western aspect and at low and middle altitudes. In contrast, plant characteristics did not contribute strongly to explain species richness and abundance, showing inconsistent patterns across taxa and sites. Therefore, at the island-wide scale abiotic, rather than biotic, factors appeared to be more important determinants of patterns in microarthropod distribution and community.
Temperature and moisture availability are likely the most important abiotic drivers of microarthropod abundance and distribution in this system. At high latitudes these two variables can be strongly related to each other (Hodkinson et al. 1999; Weltzin et al. 2003), since for example, wet habitats are more thermally buffered than drier habitats. Temperature and moisture have also both changed considerably on Marion Island over the last half century, and are likely to continue to change rapidly in the sub-Antarctic over the coming decades.
Since the western aspect of Marion Island receives more rainfall and has a higher cloud cover than the eastern aspect (Schulze 1971; Gremmen 1981; Nyakatya and McGeoch 2008), a moisture gradient is thought to exist across the island, presumably resulting in higher and/or more consistent moisture content in plants on the west. This difference in moisture, together with the more favourable, buffered temperature inside plants (Nyakatya and McGeoch 2008), are likely to be important determinants of microarthropod distribution and abundance patterns at the island-wide scale. Apart from the direct effects of moisture on microarthropods, moisture also controls rates of organic matter decomposition in fellfield environments (Smith et al. 1993), with higher decomposition rates potentially favouring microarthropods by increasing food resources.
Moisture is also likely to be important in determining the distribution of desiccation-susceptible species (Block and Harrison 1995; Strathdee and Bale 1998; Sinclair et al. 2003; Leinaas and Sømme 1984; Leinaas and Fjellberg 1985; Convey et al. 2002; Convey et al. 2003; Sømme 2012). Most springtail species are not desiccation resistant (Hopkin 1997; Hodkinson and Wookey 1999; Convey et al. 2003; Sømme 2012), and this may explain why springtail abundance was higher on the western aspect. The absence of a strong difference in mite abundance between aspects of the island may, in contrast, be due to the higher desiccation tolerance of mites in general when compared to springtails (Coulson et al. 1996; Hodkinson et al. 1999; Convey et al. 2003).
In agreement with the general pattern for species richness and abundance to decline with altitude (Rahbek 1995; Gaston and Spicer 2004), mite and springtail abundance and richness tended to be lower at higher elevations. There are a number of potential explanations for this decline, which are not always independent of each other (e.g., lower plant productivity and habitat availability; Heaney 2001). For example, soil microarthropod abundances decreased with altitude in Tibet due to increasing environmental harshness and decreasing resource availability (Jing et al. 2005), while lower arthropod abundance and species richness at high altitudes in the Andes was linked to lower plant productivity (Janzen et al. 1976). On sub-Antarctic Macquarie Island arthropod richness also declined with altitude, as a result of lower temperature, higher wind exposure, or a combination of the two variables (Davies and Melbourne 1999). Finally, leaf litter invertebrate abundances decreased with altitude in western Panama due to low decomposition rates at high altitudes (Olson 1994).
All of these mechanisms may play a role in driving the altitudinal patterns in microarthropod abundance and richness on Marion Island. In particular, temperature (and the associated impacts on decomposition), wind speed and soil freezing depth and frequency differ strongly between low and high altitude plots (Table 1; Blake 1996; Boelhouwers et al. 2003). As most high latitude invertebrates are likely adapted to withstand cold temperatures (Hodkinson and Wookey 1999; Callaghan et al. 2004; Chown and Nicolson 2004) and other climatic stresses, other non-climatic factors (e.g. lower resource availability) may also contribute strongly to lower abundances of microarthropods at high altitudes (Lawton et al. 1987; Heaney 2001; Jing et al. 2005).
Dunne et al. (2004) argued that a combination of results from gradient and experimental climate change studies is useful to understand and predict the impact of climate change on species distributions (also see Hodkinson and Wookey 1999; Shaver et al. 2000). Therefore, by comparing the results from this study to those from a climate manipulation experiment on Marion Island (McGeoch et al. 2006), as well as to other field experiments, it may be possible to make more accurate species-specific predictions of the responses of microarthropods on Marion Island to climate change.
These predictions are based on the assumption that microarthropods respond predominantly to the direct effect of changes in temperature and moisture. However, some species responses are likely to depend on both biotic and abiotic factors (and interactions between them), generating complex patterns that are not clearly linked to changes in temperature or moisture (Convey et al. 2002). Nonetheless, some predictions can be made for the species showing consistent responses in both this gradient study and the climate manipulation experiment.
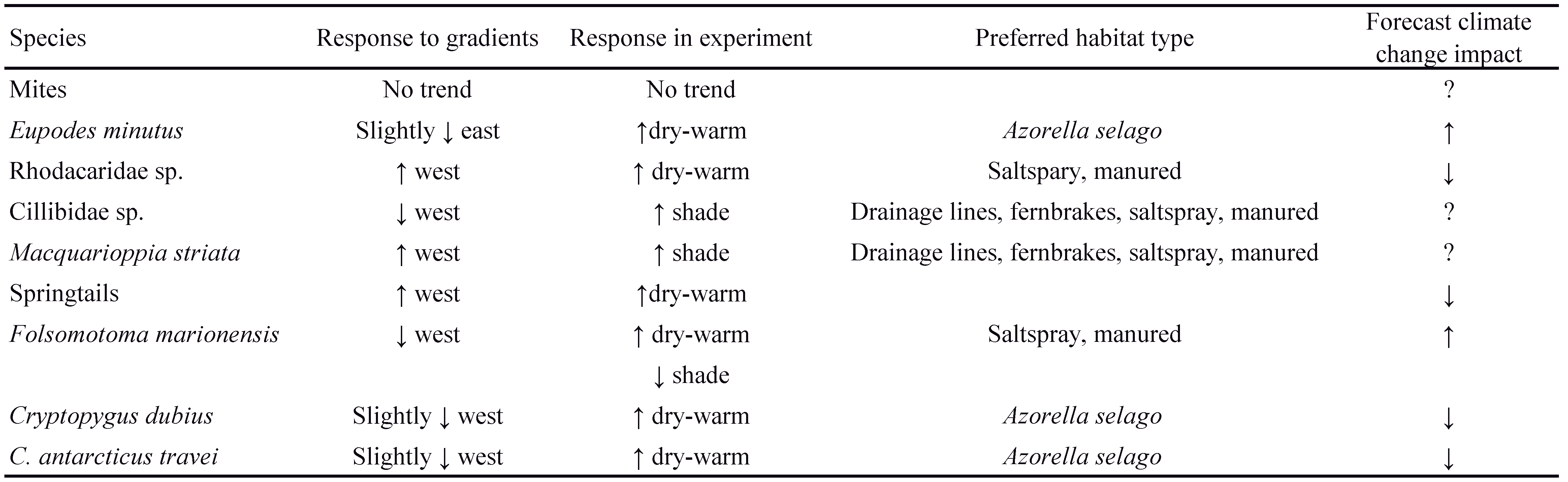
Continued changes in precipitation are forecast for the sub-Antarctic (le Roux 2008), which may have important impacts on the abundance, richness and composition of microarthropod communities. The contrasting result of springtail, but not mite, abundance differing between aspects of the island mirrors the results from the climate manipulation experiment, where springtail abundance was significantly lower under the dry-warm treatments, while mite abundances was unaffected (McGeoch et al. 2006) (see Table 3 for a summary). Therefore, both the gradient study and the experiment highlight the importance of moisture for microarthropods (in agreement with e.g., Convey et al. 2002).

Warming experiments at high latitudes have shown that the response of microarthropod abundance to higher temperatures is variable. For example, Arctic springtail abundance declined significantly under warming, while oribatid mites were unaffected (Coulson et al. 1996; Webb et al. 1998; see also Convey et al. 2002). In contrast, springtail and mite abundances increased under warming in Antarctica (Kennedy 1994). Across most studies species showed idiosyncratic responses to treatments (Kennedy 1994; Webb et al. 1998; Convey et al. 2002). Indeed, in the climate manipulation experiment on Marion Island the mite and springtail species that responded significantly to treatments exhibited different responses (McGeoch et al. 2006). It is thus clear, that generalizations regarding the impact of warmer temperatures on microarthropod abundance and distributions should be made with caution. Nonetheless, all the studies clearly demonstrate that springtails are generally responsive to changes in climate.
Due to idiosyncratic species responses, and the resulting inability to generalize predictions across the two microarthropod groups, the response of mites and springtails to climate change may be strongly driven by the response of individual species, as observed in both the gradient study and the experiment. Indeed, the response of the mite, E. minutus, can strongly drive community responses to changing conditions as this species was the most abundant species in both the experiment and in this study. The species was significantly more abundant under the dry-warm treatment, but its abundance did not differ significantly between island aspects or across altitude. However, it was slightly more dominant on the drier eastern aspect, and therefore, together with the experimental results, it is likely that E. minutus prefers drier habitats. Therefore, E. minutus abundance might increase, and has possibly already increased, with drying and warming on Marion Island (Table 3).
Rhodacaridae sp. abundances were significantly lower under experimental drying and were higher on the wetter, western aspect of the island. Rhodacaridae sp. thus seems to prefer habitats with higher humidity, in agreement with the high abundance of Rhodacaridae sp. observed in wet coastal vegetation on the island (Barendse et al. 2002). Thus, Rhodacaridae sp. abundance is likely to decline with climate warming and drying on Marion Island. In contrast, Cillibidae sp. and Macquarioppia striata were more abundant on the western aspect of the island but also in the shade treatment of the experiment (characterized by higher humidity and a lower temperature range than the controls, Le Roux et al. 2005). These results are consistent as the greater cloud cover on the western aspect of the island is associated with buffered temperature, matching conditions under the shading treatment of the experiment (Schulze 1971; Nyakatya and Mcgeoch 2008). Furthermore, since abundances of these two species are highest in drainage lines, fern brakes, saltspray and manured habitats (Barendse et al. 2002), Cillibidae sp. and M. striata seem to prefer moist, shaded habitats, with buffered temperatures. Although cloud cover on Marion Island has slightly decreased over the past 50 years (Smith 2002), the cover and distribution of several vascular plant species are increasing, possibly resulting in greater shading of Azorella selago plants (Gremmen and Smith 1999; Chown et al. 2011). Therefore, predicting the responses of these species to changing conditions will require an understanding of changes in variables other than temperature and moisture.
The springtail Tullbergia bisetosa was least abundant under the dry-warm treatment and most abundant at high altitudes and on the eastern aspect. Temperature, rather than moisture, is therefore likely to be the dominant determinant of T. bisetosa’s distribution, suggesting that its abundance will decrease under further warming. Folsomotoma marionensis abundance was lower under the dry-warm treatment, but simultaneously also higher under the shade treatment. In this study, F. marionensis abundance was higher on the wetter and cloudier western aspect. Furthermore, F. marionensis was found to be most abundant in saltspray and manured areas (Gabriel et al. 2001) and thus possibly prefers moist, shaded habitats with buffered temperatures. Therefore, similar to Cilibidae sp. and M. striata, to predict the response of this species other variables than temperature and moisture should be taken into consideration. Finally, Cryptopygus dubius and C. antarcticus travei both had lower abundances under the dry-warm treatment, while their abundances were slightly higher on the western aspect. Additionally, both species are more abundant in A. selago plants compared to other plant species on Marion Island (Gabriel et al. 2001), possibly reflecting the thermal buffering effect of A. selago. Therefore, C. dubius and C. antarcticus travei seem to prefer habitats with lower, buffered temperatures, and may therefore decrease in abundance with warming (Table 3).
Assuming direct responses of species to abiotic factors, future warming and drying is likely to have a negative effect on the abundance of Rhodacaridae sp., T. bisetosa and F. marionensis, while E. minutus is predicted to increase with ongoing drying. Although such predictions cannot be made for all species, it is clear that the microarthropod communities of Marion Island will change considerably under further climate change.
The microarthropod community occurring within Azorella selago plants appears to be strongly influenced by differences in moisture and temperature. While species generally responded idiosyncratically to variation in these abiotic factors, microarthropods, especially springtails, are likely to be negatively affected by continued warming and drying on Marion Island. Although the indirect effects of climate change on microarthropods are still uncertain (e.g. impacts via changes in their host plant species), it is clear that changes in climatic conditions may dramatically affect microarthropod community structure.
The National Research Foundation’s South African National Antarctic Programme funded this research, while the Department of Environmental Affairs (South Africa) provided logistic support. We thank Prof. M.A. McGeoch (Monash University, Australia) for her substantial contribution to this research.
Barendse J., Chown S.L. 2001. Abundance and seasonality of mid-altitude fellfield arthropods from Marion Island. Polar Biol., 24: 73-82. doi:10.1007/s003000000172 ![]()
Barendse J., Mercer R.D., Marshall D.J., Chown S.L. 2002. Habitat specificity of mites on sub-Antarctic Marion Island. Environ Entomol., 31: 612-625. doi:10.1603/0046-225X-31.4.612 ![]()
Bergström D., Chown S.L. 1999. Life at the front: history, ecology and change on southern ocean islands. Trends Ecol. Evol., 14: 472-477. doi:10.1016/S0169-5347(99)01688-2 ![]()
Blake B.J. 1996. Microclimate and prediction of photosynthesis at Marion Island. M.Sc Thesis, Department of Botany and Genetics, University of the Free State, Bloemfontein.
Block W., Harrison P.M. 1995. Collembolan water relations and environmental change in the maritime Antarctic. Global Change Biol. 1: 347-359. doi:10.1111/j.1365-2486.1995.tb00033.x ![]()
Boelhouwers J., Holness S., Sumner P. 2003. The maritime sub-Antarctic: a distinct periglacial environment. Geomorph., 52: 39-55.
Bokhorst S., Huiskes A., Convey P., Aerts R. 2007. The effect of environmental change on vascular plant and cryptogam communities from the Falkland Islands and the Maritime Antarctic. BMC Ecol., 7(1): 15. doi:10.1186/1472-6785-7-15 ![]()
Brehm G., Fiedler K. 2003. Faunal composition of geometrid moths changes with altitude in an Andean montane rain forest. J. Biogeogr., 30: 431-440. doi:10.1046/j.1365-2699.2003.00832.x ![]()
Burger A.E. 1985. Terrestrial food webs in the sub-Antarctic: island effects. In: Siegfried W., Condy P., Laws R. (Eds). Antarctic nutrient cycles and food webs. Berlin: Springer. p. 582-591. doi:10.1007/978-3-642-82275-9_79 ![]()
Callaghan T.V., Bjorn L.O., Chernov Y., Chapin T., Christensen T.R., Huntley B., Ims R.A., Johansson M., Jolly D., Johansson S., Matveyeva N., Panikov N., Oechel W., Shaver G., Elster J., Henttonen H., Laine K., Taulavuori K., Taulavuori E., Zockler C. 2004. Biodiversity, distributions and adaptations of Arctic species in the context of environmental change. Ambio, 33: 404-417.
Callaghan T.V., Jonasson S. 1995. Implications for changes in Arctic plant biodiversity from environmental manipulation experiments. In: Chapin III F.S., Körner C. (Eds). Arctic and Alpine biodiversity: patterns, causes and ecosystem consequences. Berlin: Springer Verlag. p. 151-166. doi:10.1007/978-3-642-78966-3_11 ![]()
Chapin III F.S., Shaver G.R., Giblin A.E., Nadelhoffer K.J., Laundre J.A. 1995. Responses of Arctic tundra to experimental and observed changes in climate. Ecology, 76: 694-711.
Chen I.C., Hill J.K., Ohlemüller R., Roy D.B., Thomas C.D. 2011. Rapid range shifts of species associated with high levels of climate warming. Science, 333: 1024-1026.
Chown S.L., Froneman P.W. (Eds). 2008 The Prince Edward Islands: land-sea interactions in a changing ecosystem. Stellenbosch, South Africa: African Sun Press. pp. 450.
Chown S.L., le Roux P.C., Ramaswiela T., Kalwij J.M., Shaw J.D., McGeoch M.A. 2013. Climate change and elevational diversity capacity: do weedy species take up the slack?. Biol. Lett., 9: 20120806. doi:10.1098/rsbl.2012.0806 ![]()
Chown S.L., McGeoch M.A., Marshall D.J. 2002. Diversity and conservation of invertebrates on the sub-Antarctic Prince Edward Islands. Afr. Entomol., 10: 67-82.
Chown S.L., Nicolson S.W. 2004. Insect Physiological Ecology: mechanisms and patterns. New York: Oxford University Press. pp. 254. doi:10.1093/acprof:oso/9780198515494.001.0001 ![]()
Chown S.L., Terauds A., Huntley B.J., le Roux P.C., Ramaswiela T., Shaw J.D., McGeoch M.A. 2011. South Africa's southern sentinel: Terrestrial environmental change at sub-Antarctic Marion Island. In: Zietsman L. (ed.). Observations on Environmental Change in South Africa, Stellenbosch: Sun Media. p. 139-146.
Convey P. 2001. Terrestrial ecosystem responses to climate changes in the Antarctic. In: Walther G.-R., Burga C.A., Edwards P.J. (Eds). "Fingerprints" of climate change: adapted behaviour and shifting species ranges. New York: Kluwer Academic /Plenum Publishers. p 17-42. doi:10.1007/978-1-4419-8692-4_2 ![]()
Convey P., Block W., Peat H.J. 2003. Soil arthropods as indicators of water stress in Antarctic terrestrial habitats?. Global Change Biol., 9: 1718-1730. doi:10.1046/j.1365-2486.2003.00691.x ![]()
Convey P., Pugh P.J.A., Jackson C., Murray A.W., Ruhland C., TXiong F.S., Day T.A. 2002. Response of Antarctic terrestrial microarthropods to long-term climate manipulations. Ecology, 83: 3130-3140.
Coulson S.J., Hodkinson I.D., Webb N.R., Block W., Bale J.S., Strathdee A.T., Worland M.R., Wooley C. 1996. Effects of experimental temperature elevation on high-arctic soil microarthropod populations. Polar Biol., 16: 147-153. doi:10.1007/BF02390435 ![]()
Crawford R.J.M., Cooper J. 2003. Conserving surface-nesting seabirds at the Prince Edward Islands: the roles of research, monitoring and legislation. Afr. J. Mar. Sci., 25: 415-426. doi:10.2989/18142320309504031 ![]()
Crawford R.J.M., Cooper J., Dyer B.M., Greyling M.D., Klages N.T.W., Ryan P.G., Petersen S.L., Underhill L.G., Upfold L., Wilkinson W., De Villiers M.S., Du Plesis S., Du Toit M., Leshoro T.M., Makhado A.B., Mason M.S., Merkle D., Tshingana D., Ward V.L., Whittington P.A. 2003. Populations of surface-nesting seabirds at Marion Island, 1994/95. 2002/03. S. Afr. J. Mar. Sci., 25: 427-440. doi:10.2989/18142320309504032 ![]()
Davies K.F., Melbourne B.A. 1999. Statistical models of invertebrate distribution on Macquarie Island: a tool to assess climate change and local human impacts. Polar Biol., 21: 240-250. doi:10.1007/s003000050359 ![]()
Dobson A.J. 2002. An introduction to generalized linear models. Boca Raton: Chapman and Hall/CRC Texts in Statistical Science. pp. 240.
Dunne J.A., Saleska S.R., Fischer M.L., Harte J. 2004. Integrating experimental and gradient methods in ecological climate change research. Ecology, 85: 904-916.
Epstein H.E., Calef M.P., Walker M.D., Chapin III F.S., Starfield A.M. 2004. Detecting changes in arctic tundra plant communities in response to warming over decadal time scales. Global Change Biol., 10: 1325-1334. doi:10.1111/j.1529-8817.2003.00810.x ![]()
Fielding C.A., Whittaker J.B., Butterfield J.E.L., Coulson J.C. 1999. Predicting responses to climate change: the effect of altitude and latitude on the phenology of the Spittlebug Neophilaenus lineatus. Funct. Ecol., 13 (Suppl. 1): 65-73. doi:10.1046/j.1365-2435.1999.00009.x ![]()
Gabriel A.G.A., Chown S.L., Barendse J., Marshall D.J., Mercer R.D., Pugh P.J.A., Smith V.R. 2001. Biological invasions on Southern Ocean islands: the Collembola of Marion Island as a test of generalities. Ecography, 24: 421-430.
Gaston K.J., Spicer J. 2004. Biodiversity: an introduction. Massachusetts, USA, Malden: Blackwell Publishing Ltd., pp. 191.
Gremmen NJM. (1981) The vegetation of the Subantarctic islands Marion and Prince Edward: Geobotany. The Netherlands, The Hague: Dr. W. Junk. pp. 145.
Gremmen N.J.M., Smith V.R. 1999. New records of alien vascular plants from Marion and Prince Edward Islands, sub-Antarctic. Polar Biol., 21: 401-409. doi:10.1007/s003000050380 ![]()
Hayward S.A.L., Worland M.R., Convey P., Bale J.S. 2003. Temperature preference of the mite Alaskozetes antarcticus, and the collembolan, Cryptopygus antarcticus from the maritime Antarctic. Physiol. Entomol., 28: 114-121. doi:10.1046/j.1365-3032.2003.00327.x ![]()
Heaney L.R. 2001. Small mammal diversity along elevational gradients in the Phillippines: an assessment of patterns and hypotheses. Global Ecol. Biogeogr., 10: 15-39. doi:10.1046/j.1466-822x.2001.00227.x ![]()
Hodkinson I.D., Bird J. 1998. Host-specific insect herbivores as sensors of climate change in Arctic and Alpine environments. Arct. Alp. Res., 30: 78-83. doi:10.2307/1551747 ![]()
Hodkinson I.D., Webb N.R., Bale J.S., Block W. 1999. Hydrology, water availability and tundra ecosystem function in a changing climate: the need for a closer integration of ideas?. Global Change Biol., 5: 359-369. doi:10.1046/j.1365-2486.1999.00229.x ![]()
Hodkinson I.D., Wookey P.A. 1999. Functional ecology of soil organisms in tundra ecosystems: towards the future. Appl. Soil. Ecol., 11: 111-126. doi:10.1016/S0929-1393(98)00142-5 ![]()
Hopkin S.P. 1997. Biology of the Springtails (Insecta: Collembola). Oxford: Oxford University Press. pp. 340.
Hugo E.A. 2006. Spatial patterns in the microarthropod community associated with Azorella selago (Apiaceae) on the sub-Antarctic Prince Edward Islands [PhD Thesis]. University of Stellenbosch: South Africa. pp.162.
Hugo E.A., McGeoch M.A., Marshall D.J., Chown S.L. 2004. Fine scale variation in microarthropod communities inhabiting the keystone species Azorella selago on Marion Island. Polar Biol., 27: 466-473. doi:10.1007/s00300-004-0614-4 ![]()
Huntley B. 1972. Aerial standing crop of Marion Island plant communities. S. Afr. J. Bot., 38: 115-119.
IPCC 2001. Intergovernmental Panel on Climate Change Working Group I, Climate Change 2001: The Scientific Basis. Cambridge University Press, Cambridge.
Janion-Scheepers C., Deharveng L., Bedos A., Chown S.L. 2015. Updated list of Collembola species currently recorded from South Africa. ZooKeys, 503: 55-88.
Janzen D.H., Ataroff M., Farinas M., Reyes S., Rincon N., Soler A., Soriano P., Vera M. 1976. Changes in the arthropod community along an elevational transect in the Venezuelen Andes. Biotropica, 8: 193-203.
Jing S., Solhøy T., Wang H.F., Vollan T.I., Xu R.M. 2005. Differences in soil arthropod communities along a high altitude gradient at Shergyla Mountain, Tibet, China. Arc. Antarct. Alp. Res., 37: 261-266. doi:10.1657/1523-0430(2005)037[0261:DISACA]2.0.CO;2 ![]()
Jongman R.H.G., Ter Braak C.J.F., Tongeren O.F.R. 1995. Data analysis in community and landscape ecology. The Netherlands: Cambridge University Press. pp. 299. doi:10.1017/CBO9780511525575 ![]()
Kennedy A.D. 1994. Simulated climate change: a field manipulation study of polar microarthropod community response to global warming. Ecography, 17: 131-140.
Klok C.J., Chown S.L. 2003. Resistance to temperature extremes in sub-Antarctic weevils: interspecific variation, population differentiation and acclimation. Biol. J. Linn. Soc., 78: 401-414. doi:10.1046/j.1095-8312.2003.00154.x ![]()
Lawton J.H., MacGarvin M., Heads P.A. 1987. Effects of altitude on the abundance and species richness of insect herbivores on bracken. J. Anim. Ecol., 56: 147-60. doi:10.2307/4805 ![]()
Leinaas H.P., Fjellberg A. 1985. Habitat structure and life history strategies of two partly sympatric and closely related lichen feeding collembolan species. Oikos, 44: 448-458.
Leinaas H.P., Sømme L. 1984. Adaptations in Xenylla maritima and Anurophorus laricis (Collembola) to lichen habitats on alpine rocks. Oikos, 43: 197-206.
le Roux P.C. 2008. Climate and climate change. In: Chown S.L., Froneman P.W. (Eds). The Prince Edward Islands: land-sea interactions in a changing ecosystem. Stellenbosch, South Africa: African Sun Press, p. 39-64. doi:10.18820/9781928357063/03 ![]()
le Roux P.C., McGeoch M.A. 2008a. Changes in climate extremes, variability and signature on sub-Antarctic Marion Island. Clim. Change, 86: 309-329. doi:10.1007/s10584-007-9259-y ![]()
le Roux P.C., McGeoch M.A. 2008b. Spatial variation in plant interactions across a severity gradient in the sub-Antarctic. Oecol., 155: 831-844.
le Roux P.C., McGeoch M.A. 2008c. Rapid range expansion and community reorganization in response to warming. Global Change Biol., 14: 2950-2962 doi:10.1111/j.1365-2486.2008.01687.x ![]()
le Roux P.C., McGeoch, M.A. 2010. Interaction intensity and importance along two stress gradients: adding shape to the stress-gradient hypothesis. Oecol., 162: 733-745. doi:10.1007/s00442-009-1484-9 ![]()
le Roux P.C., McGeoch M.A., Nyakatya M.J., Chown S.L. 2005. Effects of a short-term climate change experiment on a sub-Antarctic keystone plant species. Global Change Biol., 11: 1628-1639. doi:10.1111/j.1365-2486.2005.001022.x ![]()
le Roux P.C., Shaw J.D., Chown S.L. 2013. Ontogenetic shifts in plant interactions vary with environmental severity and affect population structure. New Phytol., 200: 241-250. doi:10.1111/nph.12349 ![]()
Lepš J., Šmilauer P. 2003. Multivariate analysis of ecological data using CANOCO. Cambridge: Cambridge University Press. pp. 254. doi:10.1017/CBO9780511615146.020 ![]()
Lutjeharms J.R.E., Ansorge I.J. 2008. Oceanographic setting of the Prince Edward Islands. In: Chown S.L., Froneman P.W. (Eds). The Prince Edward Islands: land-sea interactions in a changing ecosystem. Stellenbosch, South Africa: African Sun Press. p. 17-38. doi:10.18820/9781928357063/02 ![]()
McCullagh P., Nelder J.A. 1989. Generalized Linear Models. Florida: Chapman and Hall/CRC. pp. 510. doi:10.1007/978-1-4899-3242-6 ![]()
McGeoch M.A., Le Roux P.C., Hugo E.A., Chown S.L. 2006. Species and community responses to short-term climate manipulation: Microarthropods in the sub-Antarctic. Aust. Ecol., 31: 719-731. doi:10.1111/j.1442-9993.2006.01614.x ![]()
Melice J.L., Lutjeharms J.R.E., Rouault M., Ansorge I.J. 2003. Sea-surface temperatures at the sub-Antarctic islands Marion and Gough during the past 50 years. S. Afr. J. Sci., 99: 363-366.
Nyakatya M.J., McGeoch M.A. 2008. Temperature variation across Marion Island associated with a keystone plant species (Azorella selago Hook. (Apiaceae)). Polar Biol., 31: 139-151. doi:10.1007/s00300-007-0341-8 ![]()
Olson D.M. 1994. The distribution of leaf litter invertebrates along a Neotropical altitudinal gradient. J. Trop. Ecol., 10: 129-150. doi:10.1017/S0266467400007793 ![]()
Pakhomov E.A., McClelland J.W., Bernard K., Kaehler S., Montoya J.P. 2004. Spatial and temporal shifts in stable isotope values of the bottom-dwelling shrimp Nauticaris marionis at the sub-Antarctic archipelago. Mar. Biol., 144: 317-325. doi:10.1007/s00227-003-1196-3 ![]()
Parmesan C. 2006. Ecological and evolutionary responses to recent climate change. Ann. Rev. Ecol. Evol. Syst., 37: 637-669. doi:10.1146/annurev.ecolsys.37.091305.110100 ![]()
Parmesan C., Gaines S., Gonzalez L., Kaufman D.M., Kingsolver J., Peterson A.T., Sagarin R. 2005. Empirical perspectives on species borders: from traditional biogeography to global change. Oikos, 108: 58-75. doi:10.1111/j.0030-1299.2005.13150.x ![]()
Parmesan C., Ryrholm N., Stefanescu C., Hill J.K., Thomas C.D., Descimon H., Huntley B., Kaila L., Kullberg J., Tammaru T., Tennent W.J., Thomas J.A., Warren M. 1999. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature, 399: 579-583.
Parmesan C., Yohe G. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature, 421: 37-42.
Peterson A.T., Ortega-Huerta M.A., Bartley J., Sanchez-Cordero V., Soberon J., Buddemeier R.H., Stockwell D.R.B. 2002. Future projections for Mexican faunas under global climate change scenarios. Nature, 416: 626-628.
Peterson D.L., Schreiner E.G., Buckingham N.M. 1997. Gradients, vegetation and climate: spatial and temporal dynamics in the Olympic Mountains, U.S.A.. Global Ecol Biogeogr. Lett., 6: 7-17. doi:10.2307/2997523 ![]()
Phiri E.E., McGeoch M.A., Chown S.L. 2015. The abundance structure of Azorella selago Hook. f. on sub-Antarctic Marion Island: testing the peak and tail hypothesis. Polar Biol., 38: 1881-1890. doi:10.1007/s00300-015-1749-1 ![]()
Rahbek C. 1995. The elevational gradient of species richness: a uniform pattern?. Ecography, 18: 200-205. doi:10.1111/j.1600-0587.1995.tb00341.x ![]()
Root T.L., Price J.T., Hall K.R., Schneider S.H., Rosenzweig C., Pounds J.A. 2003. Fingerprints of global warming on wild plants and animals. Nature, 421: 57-60
Ryan P.G., Bester M.N. 2008. Pelagic predators. In: Chown S.L., Froneman P.W. (Eds). The Prince Edward Islands: land-sea interactions in a changing ecosystem. Stellenbosch, South Africa: African Sun Press. p. 121-164. doi:10.18820/9781928357063/06 ![]()
Sanders N.J., Moss J., Wagner D. 2003. Patterns of ant species richness along elevational gradients in an arid ecosystem. Global Ecol. Biogeogr., 12: 93-102. doi:10.1046/j.1466-822X.2003.00324.x ![]()
Schulze B.R. 1971. The climate of Marion Island. In: Van Zinderen Bakker E., Winterbottom J., Dyer R. (Eds). Marion and Prince Edward Islands. Report on the South African biological and geological expedition 1965-1966. Cape Town, South Africa: A. A. Balkema. p. 16-31.
Shaver G.R., Canadell J., Chapin F.S., Gurevitch J., Harte J., Henry G., Ineson P., Jonasson S., Melillo J., Pitelka L., Rustad L. 2000. Global warming and terrestrial ecosystems: A conceptual framework for analysis. Bioscience, 50: 871-882. doi:10.1641/0006-3568(2000)050[0871:GWATEA]2.0.CO;2 ![]()
Sinclair B.J., Klok C.J., Terblanche J.S., Chown S.L. 2003. Diurnal variation in supercooling points of three species of Collembola from Cape Hallett, Antarctica. J. Insect Physiol., 49: 1049-1061. doi:10.1016/j.jinsphys.2003.08.002 ![]()
Smith J.L., Halvorson J.J., Bolton Jr H. 2002. Soil properties and microbial activity across a 500 m elevation gradient in a semi-arid environment. Soil Biol. Biochem., 34: 1749-1757. doi:10.1016/S0038-0717(02)00162-1 ![]()
Smith V.R. 2002. Climate change in the sub-Antarctic: An illustration from Marion Island. Clim. Change, 52: 345-357. doi:10.1023/A:1013718617277 ![]()
Smith V.R., Steenkamp M. 1990. Climatic changes and its ecological implications at a subantarctic island. Oecol., 85: 14-24.
Smith V.R., Steenkamp M., French D.D. 1993. Soil decomposition potential in relation to environmental factors on Marion Island (Sub-Antarctic). Soil Biol. Biochem., 25: 1619-1633. doi:10.1016/0038-0717(93)90018-7 ![]()
Sokal R.R., Rohlf J. 1995. Biometry: the principles and practice of statistics in biological research. New York: Freeman and Company. pp. 880
Sømme L. 2012. Invertebrates in hot and cold arid environments. Berlin: Springer Science and Business Media. pp. 275.
Southwood T.R.E. 1978. Ecological methods with particular reference to the study of insect populations. London: Chapman and Hall. pp. 524.
Strathdee A.T., Bale J.S. 1998. Life on the edge: insect ecology in Arctic environments. Ann. Rev. Entomol., 43: 85-106. doi:10.1146/annurev.ento.43.1.85 ![]()
Ter Braak C.J.F., Šmilauer P. 2002. CANOCO reference manual and CanoDraw for Windows User's guide: Software for Canonical Community Ordination (version 4.5). Ithaca, NY, USA: Microcomputer Power. pp. 500.
Tweedie C.E. 2000. Climate change and the autecology of six plant species along an altitudinal gradient on sub-Antarctic Macquarie Island. [Ph.D. thesis]. University of Queensland: Brisbane. pp. 334.
Thomas C.D., Cameron A., Green R.E., Bakkenes M., Beaumont L.J., Collingham Y.C., Erasmus B.F.N., Ferreira de Siqueira M., Grainger A., Hannah L., Hughes L., Huntley B., Van Jaarsveld A.S., Midgley G.F., Miles L., Ortega-Huerta M.A., Peterson A.T., Phillips O.L., Williams S.E. 2004. Extinction risk from climate change. Nature, 427: 145-148.
Vitousek P.M. 2002. Oceanic islands as model systems for ecological studies. J. Biogeogr., 29: 573-582. doi:10.1046/j.1365-2699.2002.00707.x ![]()
Walther G., Post E.. Convey P., Menzel A., Parmesan C., Beebee T.J.C., Fromentin J., Hoegh-Guldberg O., Bairlein F. 2002. Ecological responses to recent climate change. Nature, 416: 389-395.
Webb N.R., Coulson S.J., Hodkinson I.D., Block W., Bale J.S., Strathdee A.T. 1998. The effects of experimental temperature elevation on populations of cryptostigmatic mites in high Arctic soils. Pedobiologia, 42: 298-308.
Weltzin J.F., Loik M.E., Schwinning S., Williams D.G., Fay P.A., Haddad B.M., Harte J., Huxman T.E., Knapp A.K., Lin G., Pockman W.T., Shaw R., Small E.E., Smith M.D., Smith S.D., Tissue D.T., Zak J.C. 2003. Assessing the response of terrestrial ecosystems to potential changes in precipitation. Bioscience, 53: 941-952.

Appendix 1. Marion Island showing the positions of the four transects and sampling plots at low, middle and high altitudes. The locations of the climate change experiment (Skua Ridge) and the meteorological base station are also indicated (from Nyakatya and McGeoch 2008).
Appendix 1

Appendix 2. Generalized linear models for mite and springtail species richness (S) and abundance (N). Analyses including grass cover were conducted for only the middle and low altitude plots (8 plots, df = 280) due to the scarcity of grass at high altitude plots. Letters in bold indicate significant values.
Appendix 2

Appedix 3. Total and relative abundance of Acari and Collembola species sampled in four different transects, with abbreviations used in graphs. * = introduced species (Chown and Froneman 2008).
Note: 1Collembola names used in previous papers: Cryptopygus caecus (currently Mucrosomia caeca), Isotomurus cf. palustris (identified as Isotomurus maculatus), Megalothorax sp. (indentified as Megalothorax minimus) (see Janion-Scheepers et al. 2015).
Appendix 3



2016-10-17
Date accepted:
2018-05-24
Date published:
2018-09-28
Edited by:
Sidorchuk, Ekaterina

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Hugo-Coetzee, Elizabeth A. and Le Roux, Peter C.
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)