



Biological control of spider mites in North-Italian vineyards using pesticide resistant predatory mites
Lorenzon, Mauro1 ; Pozzebon, Alberto2 and Duso, Carlo3
1Department of Agronomy, Food, Natural Resources, Animals and Environment – University of Padova, viale dell'Università, 16 35020 Legnaro (PD), Italy.
2✉ Department of Agronomy, Food, Natural Resources, Animals and Environment – University of Padova, viale dell'Università, 16 35020 Legnaro (PD), Italy.
3Department of Agronomy, Food, Natural Resources, Animals and Environment – University of Padova, viale dell'Università, 16 35020 Legnaro (PD), Italy.
2018 - Volume: 58 Issue: Suppl pages: 98-118
https://doi.org/10.24349/acarologia/20184277Keywords
Abstract
The possibility to use phytoseiid mites to control grape mites in European vineyards has been suggested first in the 1970s (Ivancich-Gambaro, 1973) and then been demonstrated to be effective in the 1980s (Girolami, 1981; Baillod et al. 1982; Schruft, 1985). Several factors can affect the success of predatory mites as biological control agents, e.g. pesticide application, cultivar features, presence of alternative food, intraguild competition, activity of predatory insects (Duso, 1992; Duso and Pasqualetto, 1993; English-Loeb et al. 2002; Duso et al. 2012). In Integrated Pest Management (IPM) programs these factors and predatory mite species attributes should be considered to maximize their performance. Mechanisms of intraguild competition may enhance or disrupt biological control (e.g. Croft and McRae, 1993; Rosenheim et al.1995; Seelmann et al. 2007).
Studies conducted in vineyards in North-eastern Italy have shown that predatory mite populations sometimes disappear or get reduced to low levels because of pesticide use leading to spider mites outbreaks (Ivancich Gambaro, 1973; Girolami, 1981; Duso et al. 1983). In other situations, vineyards can be inhabited by predatory mite populations that can tolerate pesticides but disappear when food is scarce or climatic conditions are unsuitable (Duso, 1989; Duso et al. 1991). Certain predatory mites such as Kampimodromus aberrans (Oudemans) and Typhlodromus pyri Scheuten have shown a high persistence in vineyards when prey is scarce (Ivancich Gambaro, 1973; Baillod et al. 1982; Engel and Ohnesorge, 1994). Experimental releases of K. aberrans and T. pyri proved to be successful in terms of spider mite control while those of Amblyseius andersoni (Chant) were ineffective (Duso et al. 1983; Girolami, 1987; Duso, 1989; Duso et al. 1991; Girolami et al. 1992; Duso and Pasqualetto, 1993). Moreover, K. aberrans showed a high competitiveness towards T. pyri, A. andersoni and Phytoseius finitimus Ribaga (Duso and Vettorazzo, 1999).
In the last decades outbreaks of Eotetranychus carpini (Oudemans) have been reported frequently in North-eastern Italy. Pesticides misuse, in particular that of ethylene-bis-dithiocarbamates (EBDCs) and organophosphates (OPs) has been claimed to be the key factor in this phenomenon. EBDCs are used to control Grape downy mildew Plamopara viticola while OPs are employed to control leafhoppers (e.g., Scaphoideus titanus Ball.) and berry moths (e.g., Lobesia botrana Den. & Schiff.). Moreover, native predatory mite populations (A. andersoni or Ph. finitimus) appear to be ineffective in keeping E. carpini densities under economic threshold levels. In two farms where spider mites infestations were previously observed, releases of K. aberrans and T. pyri were planned to compare their effectiveness in controlling E. carpini. At the same time we evaluated the effects of competition between released and native predatory mite species. In contrast with similar experiments carried out in the past (e.g. Duso et al. 1991; Duso and Pasqualetto, 1993; Duso and Vettorazzo, 1999) predatory mites used for releases were known to be resistant to EBDCs and OPs, two pesticide categories frequently involved in the disruption of biological control in vineyards (Ivancich Gambaro, 1973; Duso et al. 1983; Girolami, 1981).
Experiments were carried out in vineyards in two farms (A and B) located in the Veneto region, North-eastern Italy.
Farm A is located at Spresiano (45°47’49” N; 12°15’05” E; 64 m a.s.l.), Treviso province. Four vineyards of the cultivar Glera, planted in the same year but with different canopy management and pruning systems (i.e. Pergola, Sylvoz, G.D.C. and Free cordon) were selected. Spider mite (P. ulmi and E. carpini) infestations were frequently observed in the seasons preceding the experiments. Prior to releases 40 two-years old branches were collected during winter and moved to the laboratory. Here the branches were analyzed under a stereomicroscope using a forceps to remove the bark on the internodes and for bud dissection. Overwintering predatory mites were counted and identified. Their identity was confirmed by mounting adult mites on slides in Hoyer’s medium, and by identified them using a phase contrast microscope and morphological keys (e.g., Tixier et al. 2012). the predatory mites A. andersoni and Paraseiulus talbii (Athias-Henriot) were found on these branches (respectively 1.51 and 0.12 mean overwintering females per internode and a bud).
Farm B is located at Meolo (45°38’06’’ N; 12°27’19” E; 0 m a.s.l.), Venice district. A single vineyard of the Verduzzo trevigiano cultivar (Sylvoz pruning system) was considered. High population densities of E. carpini were recorded in the year preceding experiment. Samples collected during winter showed the presence of moderate densities of Ph. finitimus and A. andersoni (2.78 and 0.43 overwintered females per internode).
The K. aberrans and T. pyri strains released were collected from two farms located in the Veneto region, North-eastern Italy. Kampimodromus aberrans was collected from a farm located in the Verona province (Monteforte d’Alpone) where it was the dominant phytoseiid species. In previous studies the release of this strain in vineyards proved to be successful (Facchin, 1996). The T. pyri strain was collected from a farm located in the Treviso province (San Pietro di Feletto) where it was completely dominant among phytoseiid species. Kampimodromus aberrans and T. pyri strains proved to be resistant to OPs and EBDCs (Posenato, 1994; Moret, 2006; Pozzebon et al. 2010; Tirello et al. 2012).
In January, before releases, 40 two-year old branches were collected from each farm and brought to the laboratory. Here the branches were analyzed as described above, and the identity and density of overwintered phytoseiids were recorded. Densities of 7.21 K. aberrans females per internode and of 11.34 T. pyri females per internode were calculated. These figures were considered in order to assess the number of predatory mites to be released per bud.
Release procedures are described in Duso (1989) and Duso et al. (1991). The number of overwintered females released on each vine was calculated considering the mean number of buds per vine.
In each vineyard of farm A, six release or non-release treatments randomly replicated four times were compared. Treatments were settled according to addition series design (Jolliffe, 2000) that allows us to test the effect of predator diversity and density without confounding between the two factors (Table 1). In particular, the effect of predatory mite diversity was tested by keeping constant the total density in mixed and single species releases, and the effect of predatory mites density was tested by compared two levels of release density (0.5 and 1 predatory mites per bud; Table 1). A control without predatory mite releases was also included (Table 1). Each replicate comprised three continuous vines, separated from the subsequent plot by 10-30 vines, depending on vineyard size. Releases were carried out only in February of 2009 and observations were conducted over the vegetative seasons of 2009 and 2010. The fungicides EBDCs (mixed with dimethomorph), copper hydroxide, wettable sulphur were used to control Grape downy mildew and Grape powdery mildew. EBDCs were used four times in April – May. Regarding insecticides, chlorpyriphos was applied in July of 2009 and thiamethoxam in July of 2010.


In farm B six release or non-release treatments randomly replicated were compared using a similar experimental design to farm B (Table 1). Here the release density was increased compared to Farm A (2 or 4 predatory mites per bud; Table 1) with the aim of obtaining a prompt control of spider mites. Six replicates per treatment were established and each replicate comprised two subsequent vines separated by about ten vines from the subsequent plot. Releases were carried out in February 2010 and observations were conducted from April to September of the same year. EBDC fungicides, copper and wettable sulphur were used as fungicides. No insecticides or acaricides were used.
Seasonal mite abundance was monitored by taking leaf samples every two weeks. Samplings started approximately two months after releases of predatory mites. At each sampling date a total of 48-64 leaves per treatment (Table 1) were collected (eight leaves per replicate) on mid shoots. Leaves were transferred to the laboratory and immediately observed by using a dissecting microscope in order to assess the identity and density of mites. Predatory mites identity was confirmed as described above.
In Farm A, different canopy managements were applied in different vineyards, with potential effect on mite’s population dynamics (e.g., Prischmann et al. 2006). For this reason we assessed the structure of the canopy of each vineyard by applying the “point quadrat” method (Wilson, 1963, Bertamini et al. 1994). Following this method we used a square plastic panel measuring 1 m2 in size with pre-drilled holes at 10 cm intervals. The panel is placed on the vegetation. A pointed metal pole approximately 1 m length was inserted in the holes in the panel and used to penetrate the canopy. The number of contacts of the metal pole with leaves and bunches were recorded. The following parameters were calculated: (1) percentage of empty spaces in the canopy, obtained by calculating the ratio between the number of gaps detected and the total number of penetrations; (2) number of leaf layers, i.e. the number of leaf contacts divided by the number of penetrations; (3) percentage of leaves inside canopy, by dividing the number of leaves (except the first and last of each penetrations) by the total number of leaves detected.
Data on predatory mites populations were analyzed using a Restricted Maximum Likelihood Repeated Measures model with the Proc MIXED of SAS® ver. 9.4. Treatments, time and their interactions were considered as fixed effect and were evaluated with F test (α = 0.05). The Kenward-Roger method was used for degrees of freedom estimation (Littell et al. 1996). We tested the effects of experimental factors on mite populations using contrasts with F test (α = 0.05). The parameters of canopy structure were analyzed using one-way ANOVA and means were separated by Tukey test (α = 0.05).
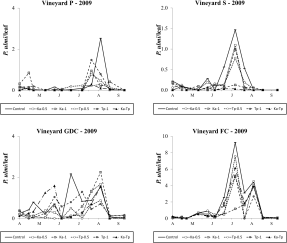
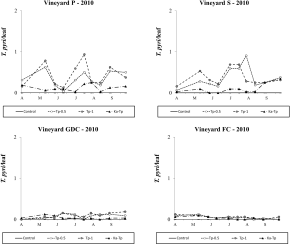
In 2009, P. ulmi populations reached relatively low densities in vineyards trained with Pergola (P), Sylvoz (S) and G.D.C. (GDC) systems; peaks of 2.53, 1.47 and 2.17 motile forms per leaf were observed in plots without predatory mites release. European red spider mite populations reached higher levels (peak of 9.25 motile forms per leaf in the control plots) in the vineyard trained with free cordon system (FC) (Figure 1). Regarding P. ulmi abundance in vineyard P, the effect of treatment was significant (F 5, 76.5=2.50; P=0.038). K. aberrans releases significantly affected P. ulmi densities (F 1, 76.5=7.82; P=0.006) but this result was obtained with the highest release densities (F 1, 76.5=8.43; P=0.005; Figure 1). In contrast, T. pyri releases did not affect significantly P. ulmi abundance (F 1, 76.5=0.86; P=0.357; Figure 1). Therefore, K. aberrans releases were more effective than T. pyri releases in controlling P. ulmi (F 1, 76.5=5.24; P=0.025). Panonychus ulmi densities were not significantly reduced in Ka-Tp treatment compared to the control (F 1, 76.5=0.05; P=0.820). The effect of density in predatory mite releases was not significant (F 1, 76.5=0.09; P=0.768). In single releases, at the same release density, K. aberrans releases proved to be more effective than mixed releases (Ka-Tp) in reducing P. ulmi numbers (F 1, 76.5=7.16; P=0.009; Figure 1). The effect of treatment was not significant in vineyard S (F 5, 49.6=2.03; P=0.091; Figure 1), nor in vineyard GDC (F 5, 56.9=1.51; P=0.202; Figure 1). Nevertheless the contrasts’ analysis showed a significant effect of K. aberrans releases (independently on the release density) on spider mite densities (p < 0.05) in both vineyards. In vineyard FC the effect of treatment was significant (F 5, 60.2=2.90; P=0.021; Figure 1). Kampimodromus aberrans and T. pyri releases significantly reduced P. ulmi numbers (F 1, 60.2=6.42; P=0.014; F 1, 60.2=4.71; P=0.034, respectively; Figure 4). There were no differences between predatory mite species in terms of spider mite abundance (F 1, 60.2=0.20; P=0.659) nor between their density levels (F 1, 60.2=0.28; P=0.599). Panonychus ulmi densities were significantly reduced in Ka-Tp treatment compared to the control (F 1, 60.2=12.56; P<0.001). Considering the same release density, there were no differences between single or mixed releases (F 1, 60.2=2.44; P=0.123; F 1, 60.2=1.16; P=0.285; for K. aberrans and T. pyri respectively; Figure 1). During 2010 P. ulmi reached negligible densities in the four vineyards.

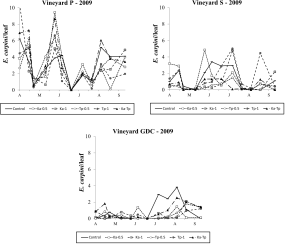
During the 2009 season, E. carpini populations fluctuated at relatively high (i.e., more than 1 motile form per leaf as seasonal average; Figure 2) densities in P and S vineyards, moderate densities in GDC (i.e., 0.5 – 1 motile forms per leaf as seasonal average; Figure 2), and negligible levels in FC vineyard (i.e., less than 0.01 motile form per leaf as seasonal average; data not shown). The effect of treatment was not significant in vineyard P (F 2, 52=1.16; P=0.339; Figure 2) nor in vineyard S (F 5, 36.6=0.92; P=0.482; Figure 2). In contrast, it was significant in GDC vineyard (F 5, 58=2.68; P=0.030; Figure 2) where T. pyri releases significantly reduced E. carpini numbers, independently on release density (F 1, 58=9.51; P=0.003), in contrast with K. aberrans releases (F 1, 58=2.28; P=0.136). Only the lowest release densities of K. aberrans affected E. carpini densities (F 1, 58=4.24; P=0.044). There were no differences in the abundance of spider mites between release plots of the two predatory mites (F 1, 58=3.72; P=0.087). Eotetranychus carpini densities were significantly reduced in Ka-Tp treatment compared to the control (F 1, 58=4.79; P=0.033; Figure 2). The remaining comparisons were not significant.
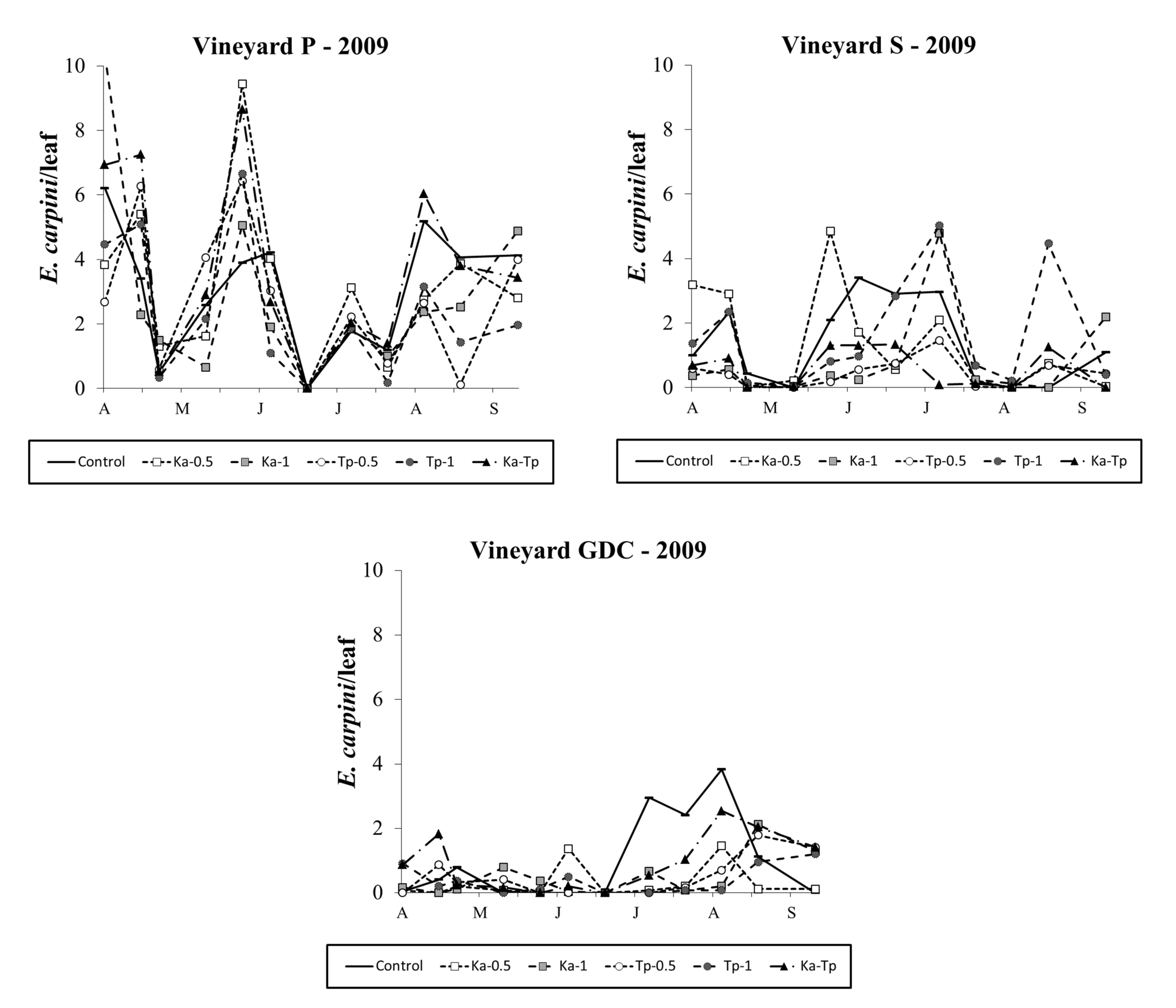
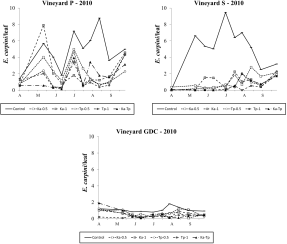
In 2010 E. carpini populations reached relatively high densities in vineyards P and S but low levels in vineyard GDC (Figure 3). The occurrence of this mite pest was still negligible in vineyard FC (data not shown). In the first vineyard differences among treatments were significant (F 5, 53.3=15.45; P<0.001). Lower E. carpini densities were detected in release treatments than in control plots (F 1, 55.3=50.53; P<0.001; F 1, 55.3=70.92; P<0.001, respectively for K. aberrans and T. pyri treatments; Figure 3). No differences were found among released species (F 1, 55.3=2.59; P=0.113) nor the densities of release (F 1, 55.3=1.75; P=0.191). Spider mite levels were significantly reduced in Ka-Tp treatment compared to the control (F 1, 55.3=32.33; P<0.001; Figure 3). In vineyard S, the effect of treatment was significant (F 5, 54.3=16.66; P<0.001; Figure 3) as spider mite populations were lower in release treatments than in the control (F 1, 54.3=8.20; P<0.001; F 1, 54.3=72.88; P<0.001, respectively for K. aberrans and T. pyri treatments; Figure 3). There were no differences between K. aberrans and T. pyri release treatments (F 1, 54.3=1.24; P=0.271) nor between the densities of release (F 1, 54.3=3.26; P=0.077). In Ka-Tp plots there were less spider mites compared to the control (F 1, 54.3=36.96; P< 0.001; Figure 3). Additional differences among treatments emerged in GDC vineyard (F 5, 39.5=2.59; P=0.04; Figure 3) where released predatory mites significantly reduced E. carpini densities compared to the control (F 1, 39.5=8.01; P=0.007; F 1, 39.5=8.86; P=0.005, respectively for K. aberrans and T. pyri treatments; Figure 3). There were no differences between K. aberrans and T. pyri release treatments (F 1, 39.5=0.03; P=0.859), nor between the densities of release (F 1, 39.5=1.12; P=0.296). Spider mite numbers were lower in Ka-Tp treatment than in the control (F 1, 39.5=5.37; P=0.026; Figure 3).

In 2009, native predatory mites (A. andersoni) reached moderate to low densities in the four vineyards. In vineyard P, the effect of treatment was significant (F 5, 63.4=5.02; P<0.001; Figure 4). Compared to the control, A. andersoni numbers were significantly lower in T. pyri release plots (F 1, 63.4=13.65; P<0.001) but not in K. aberrans plots (F 1, 63.4=1.41; P=0.24). Amblyseius andersoni densities were lower in T. pyri than in K. aberrans release plots (F 1, 63.4=9.44; P=0.003) and in Ka-Tp plots compared to the control (F 1, 63.4=8.29; P=0.005). The effect of release density was not significant (F 1, 63.4=0.344; P=0.559; Figure 4).
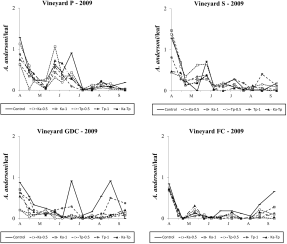
In vineyard S there were no differences among treatments (F 5, 53.5=2.09; P=0.081; Figure 4). However, the contrasts’ analysis showed that releases affected A. andersoni numbers (F 1, 53.5=4.09; P=0.048), in particular K. aberrans releases (F 1, 53.5=4.06; P=0.049; Figure 4).
In contrast, differences among treatments were significant in vineyard GDC ( F 5, 50.7=11.97; P<0.001; Figure 4). Amblyseius andersoni densities were reduced in K. aberrans as well as T. pyri treatments (F 1, 50.7=41.89; P<0.001; F 1, 50.7=31.76; P<0.001, respectively) included Ka-Tp (F 1, 50.7=37.26; P<0.001). There were no differences in the effect of the two released species (F 1, 50.7=1.05; P=0.31). The effect of release density was significant (F 1, 50.7=8.46; P=0.005; Figure 4) but there were less A. andersoni in plots with the lowest release density.
The effect of treatment was significant also in vineyard FC (F 5, 63.2=4.56; P=0.001; Figure 4) where A. andersoni densities were significantly reduced by released predators (F 1, 63.2=18.66; P<0.001). This effect was significant in K. aberrans as well as in T. pyri plots (F 1, 63.2=13.14; P<0.01; F 1, 63.2=16.64; P<0.001, respectively) included Ka-Tp plots (F 1, 63.2=11.45; P<0.01). There were no differences between T. pyri and K. aberrans treatments (F 1, 63.2=0.31; P=0.580). The effect of release density was not significant (F 1, 63.2=0.00; P=0.961; Figure 4).
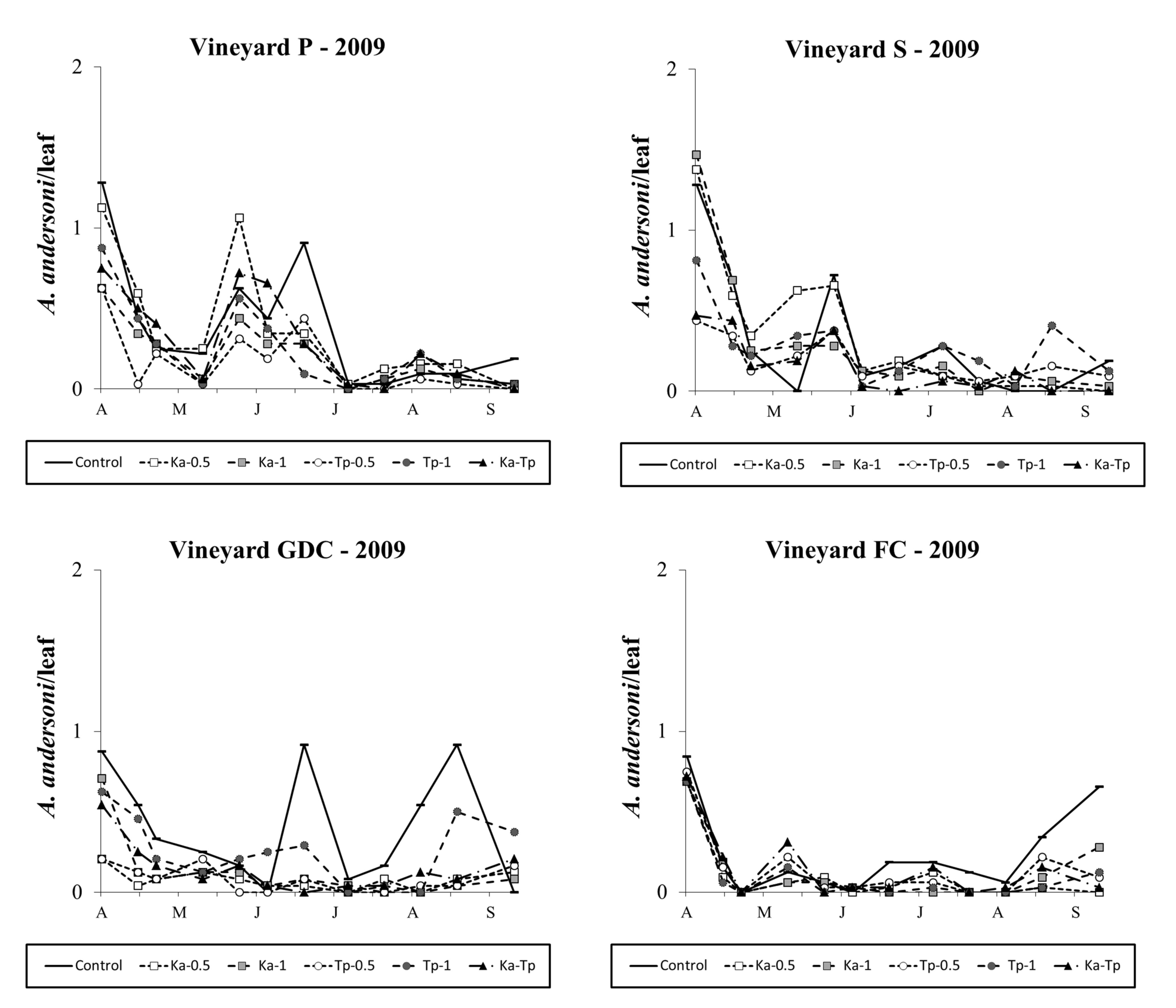
In 2010, A. andersoni populations appeared to be less abundant than in 2009. The effect of treatment was significant in all vineyards (vineyard P: F 5, 37.5=11.82; P<0.001; vineyard S: F 5, 35.4=8.96; P<0.001; vineyard GDC: F 5, 49.9=5.75; P<0.001; vineyard FC: F 5, 61.9=5.61; P<0.001; Figure 5). Amblyseius andersoni densities were reduced in K. aberrans (vineyard P: F 1, 37.5=45.31; P<0.001; vineyard S: F 1, 35.4=39.73; P<0.001; vineyard GDC: F 1, 49.9=21.28; P<0.001; vineyard FC: F 1, 61.9=22.37; P<0.001) as well as in T. pyri treatments (vineyard P: F 1, 37.5=50.23; P<0.001; vineyard S: F 1, 35.4=32.89; P<0.001; vineyard GDC: F 1, 49.9=13.95; P<0.001; vineyard FC: F 1, 61.9=13.14; P<0.001). There were no differences between the released species (vineyard P: F 1, 37.5=0.190; P=0.665; vineyard S: F 1, 35.4=0.48; P=0.491; vineyard GDC: F 1, 49.9=1.29; P=0.626; vineyard FC: F 1, 61.9=2.27; P=0.137) or the density of release (vineyard P: F 1, 37.5=0.01; P=0.912; vineyard S: F 1, 35.4=0.04; P=0.834; vineyard GDC: F 1, 49.9=0.09; P=0.766; vineyard FC: F 1, 61.9=0.74; P=0.392; Figure 5).
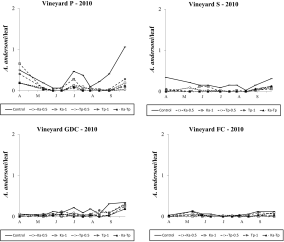

Kampimodromus aberrans populations successfully established after releases, even if predatory mite numbers did not reach high levels. In 2009, the effect of treatment was significant in all vineyards (vineyard P: F 3,46=18.85; P<0.001; vineyard S: F 3, 35.8=22.69; P<0.001; vineyard GDC: F 3, 37.5=33.89; P<0.001; vineyard FC: F 3, 38.8=18.70; P<0.001; Figure 6) and there were more K. aberrans in the respective release plots than in the control (vineyard P: F 1,46=47.62; P<0.001; vineyard S: F 1, 35.8=66.93; P<0.001; vineyard GDC: F 1, 37.5=86.55; P<0.001; vineyard FC: F 1, 38.8=35.72; P<0.001). In Ka-Tp treatments the presence of T. pyri affected K. aberrans densities in vineyards GDC and FC: (respectively: F 1, 37.5=4.32; P=0.044; F 1, 38.8=12.91; P<0.001; Figure 6) but not in vineyards P and S (respectively: F 1,46=2.02; P=0.162; F 1, 35.8=0.01; P=0.988; Figure 6).
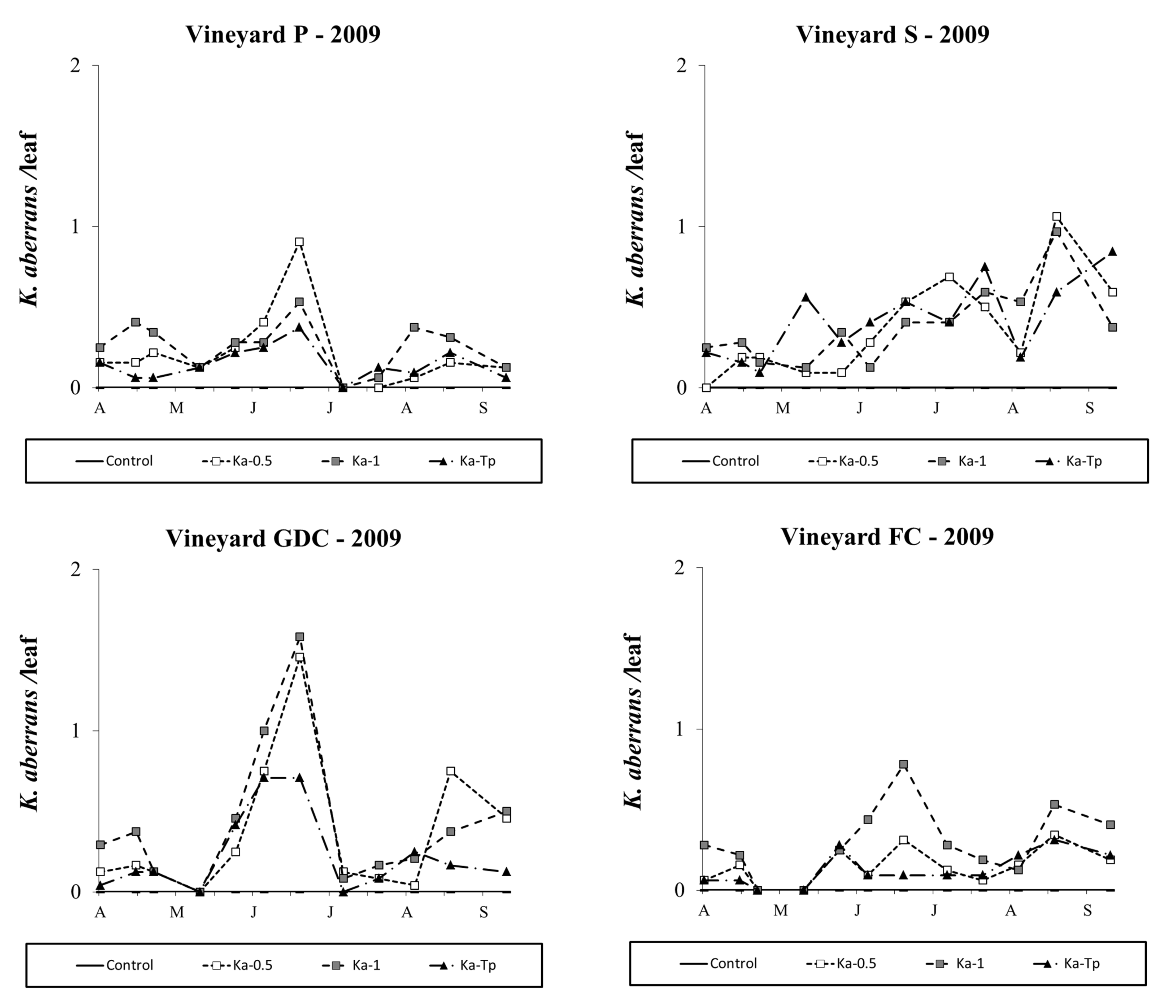
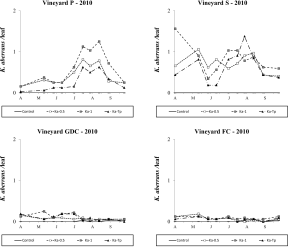
One year later, K. aberrans populations were more abundant in P and S vineyards than in GDC and FC ones. The effect of treatment was significant in all vineyards (vineyard P: F 3,31.2=6.05; P=0.002; vineyard S: F 3, 26.4=3.15; P=0.042; vineyard GDC: F 3, 33.6=27.58; P<0.001; vineyard FC: F 3, 36.7=21.2; P<0.001; Figure 7) and there were more K. aberrans in the respective release plots than in the control (vineyard P: F 1,31.2=14.87; P<0.001; vineyard S: F 1, 26.4=8.77; P=0.006; vineyard GDC: F 1, 33.6=80.12; P<0.001; vineyard FC: F 1, 36.7=47.14; P<0.001). In Ka-Tp treatments the presence of T. pyri did not affect K. aberrans densities (F 1,31.2=2.29; P=0.140; F 1, 26.4=0.68; P=0.417; F 1, 33.6=0.25; P=0.618; F 1, 36.7=3.15; P<0.084; P, S, GDC and FC respectively; Figure 7).

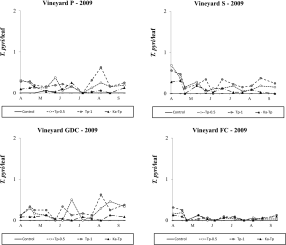
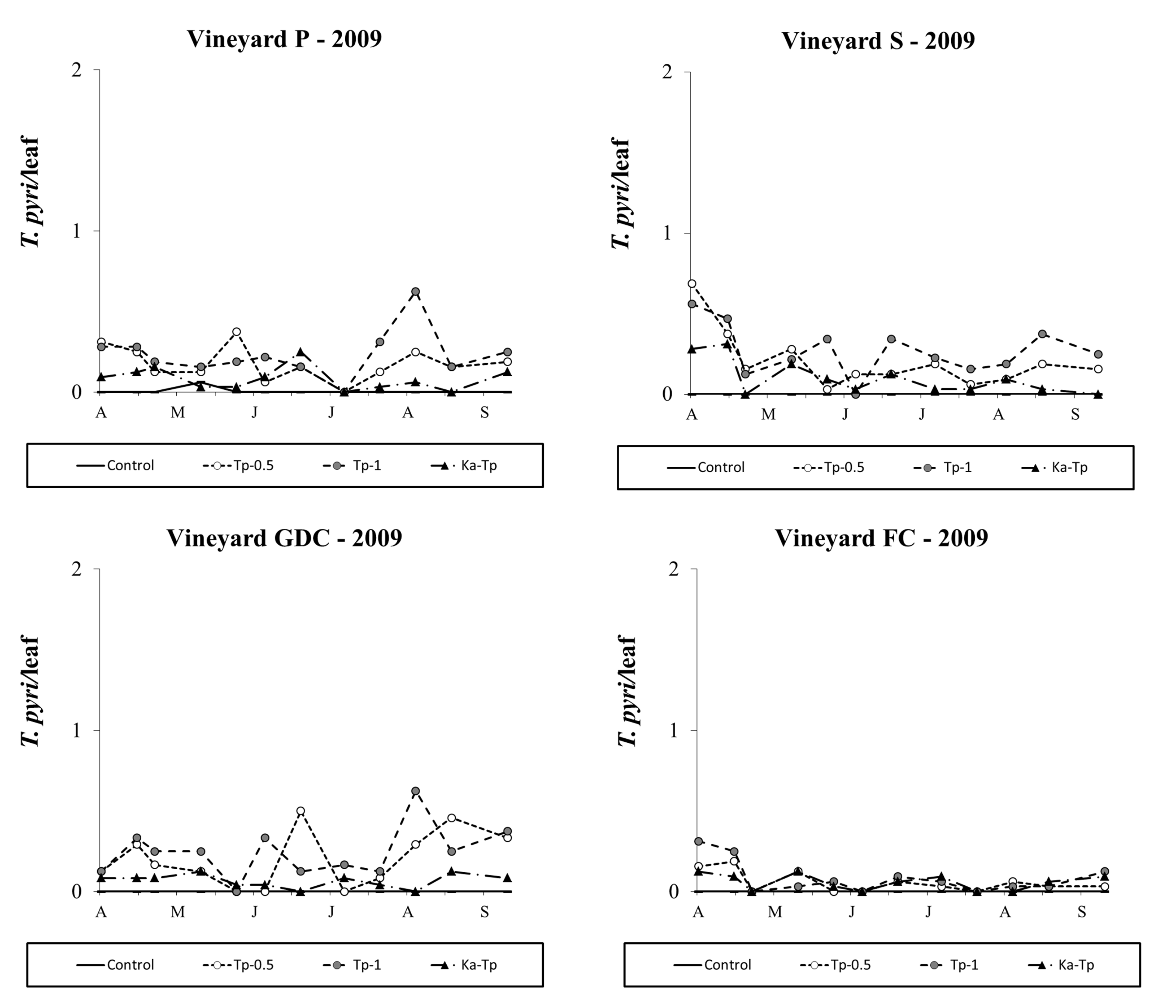
Typhlodromus pyri releases appeared to be successful but its densities were always lower than 1 motile form per leaf in 2009. The effect of treatment was significant in all vineyards (vineyard P: F 3,48.5=22.99; P<0.001; vineyard S: F 3, 25=3.96; P=0.019; vineyard GDC: F 3, 33.6=18.59; P<0.001; vineyard FC: F 3, 30.4=6.78; P=0.001; Figure 8) and there were more T. pyri in the respective release plots than in the control (vineyard P: F 1,48.5=43.79; P<0.001; vineyard S: F 1, 25=4.51; P=0.044; vineyard GDC: F 1, 33.6=33.49; P<0.001; vineyard FC: F 1, 38.8=18.18; P=0.001). In vineyards P and GDC the presence of K. aberrans affected T. pyri densities where the two species were released in the same plots (F 1, 48.5=10.11; P=0.003; F 1, 33.6=12.14; P=0.001; Figure 8) but this phenomenon did not occur in vineyards S (F 1, 25=2.52; P=0.125; Figure 8) and FC (F 1, 38.8=0.01; P=0.988; Figure 8).
In 2010, the effect of treatment confirmed to be significant in vineyards P (F 3, 45.7=29.57; P<0.001; Figure 9), S (F 3, 31=21.57; P<0.001; Figure 9), and GDC (F 3, 30=6.72; P=0.001; Figure 9) but not in vineyard FC ( F 3, 15.7=21.2; P=0.425; Figure 9). Typhlodromus pyri densities were higher in the respective release plots than in the control plots of vineyards P (F 1, 45.7=59.61; P<0.001), S (F 1, 31=36.47; P<0.001) and GDC (F 1, 30=13.57.12; P=0.001); however this effect was not significant in vineyard FC (F 1, 15.7=2.42; P=0.140) where predatory mites reached low population densities. In Ka-Tp treatments the presence of K. aberrans reduced that of T. pyri in vineyards P (F 1, 45.7=20.12; P<0.001; Figure 9) and S (F 1, 31=19.32; P<0.001; Figure 9) in contrast with vineyards GDC (F 1, 30=2.91; P=0.09; Figure 9) and FC (F 1, 15.7=0.18; P=0.675; Figure 9).

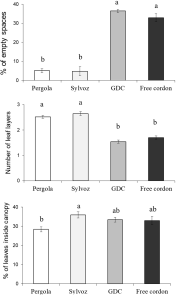
The analysis of canopy structure showed significant differences among vineyards in certain parameters. Vineyards P and S had a lower incidence of empty spaces than vineyards GDC and FC (F 3, 164 = 98.78; P<0.001; Figure 10). The number of leaf layers was higher in canopies of vineyards P and S than in those of GDC and FC (F 3, 164 = 76.28; P<0.001; Figure 10). The proportion of leaves inside the canopy was higher in vineyard P than in vineyard S, while vineyards GDC and FC gave intermediate results (F 3, 164 = 3.72; P=0.013; Figure 10).

Here Panonychus ulmi was not detected. The effect of treatment was significant (F 5, 59.6=11.01; P<0.001), and there were less spider mites in predatory mite release plots than in the control ones (F 1, 59.6=48.85; P<0.001). The effects of K. aberrans or T. pyri releases were both significant ( F 1, 59.6=27.36; P<0.001; F 1, 59.6=49.09; P<0.001, respectively), and the latter were more effective (F 1, 59.6=4.73; P=0.034). Spider mites were reduced significantly even in Ka-Tp plots compared to control (F 1, 59.6=34.50; P<0.001). The effect of density in predatory mite releases was not significant (F 1, 59.6=0.69; P=0.41; Figure 11). All other comparisons were not significant.
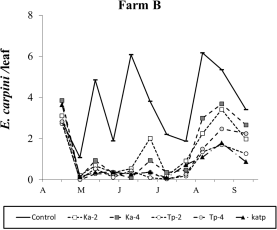

Among native predatory mites the most frequent were Ph. finitimus and A. andersoni. Their populations fluctuated at low to moderate densities (i.e., < 0.5 motile forms per leaf) over the season (Figure 12). Regarding Ph. finitimus the effect of treatment was not significant (F 5, 56.5=1.39; P=0.243). In the contrasts’ analysis Ph. finitimus was less abundant in release plots than in the control (F 1, 56.5=4.42; P=0.04). This result was due to the effect of T. pyri releases (F 1, 56.5=5.06; P=0.028; Figure 12).
eginfigure[t] % figure12 entering ncludegraphics[width=17.5cm]Figure12.pdf aptionSeasonal abundance of predatory mites observed during 2010 (months are indicated in x-axis) on different treatments in Farm B. ndfigure
Regarding A. andersoni, the effect of treatment was significant (F 5, 96.4=3.13; P=0.012; Figure 12). Amblyseius andersoni numbers were significantly reduced in release plots (F 1, 96.4=11.90; P<0.001) and the effects of K. aberrans or T. pyri releases were both significant (F 1, 96.4=5.33; P=0.023; F 1, 96.4=13.18; P<0.001, respectively). There were no differences between K. aberrans or T. pyri treatments (F 1, 96.4=2.63; P=0.108). Significant differences were recorded between Ka-Tp and control plots (F 1, 96.4=9.44; P=0.003). The effect of release density was not significant (F 1, 96.4=0.52; P=0.473; Figure 12).
Kampimodromus aberrans releases were successful but predatory mite populations reached relatively low levels. The effect of treatment was significant (F 3, 45.9=19.61; P<0.001; Figure 12) and there were more K. aberrans in the respective release plots than in the control (F 1, 45.9=45.30; P<0.001). In Ka-Tp treatments the presence of T. pyri did not affect K. aberrans densities S (F 1, 45.9=1.01; P=0.312). Higher K. aberrans numbers were recorded in plots receiving the highest release densities (F 1, 45.9=6.56; P=0.014; Figure 12).
Typhlodromus pyri releases were also successful but also in these cases predatory mites did not exceed densities of 1 motile form per leaf. The effect of treatment was significant (F 3, 47.4=20.16; P<0.001; Figure 12) and T. pyri densities were higher in the respective release plots than in the control plots (F 1, 47.4=46.66; P<0.001). The presence of K. aberrans did not reduce that of T. pyri in Ka-Tp treatments (F 1, 47.4=2.98; P=0.09). The effect of release density was not significant (F 1, 47.4=3.95; P=0.053; Figure 12).
Mite seasonal abundance in farm A showed different patterns in the selected vineyards. The most interesting differences concerned spider mite populations in 2009: E. carpini was dominant in vineyards P, S and GDC, while P. ulmi in vineyard FC. Factors affecting the negligible occurrence of E. carpini in vineyard FC compared to vineyards trained with other systems should be investigated. The canopy of free cordon vineyard appeared to be less vigorous (lower number of leaf layers) and irregular (lower percentage of empty spaces) compared with Pergola and Sylvoz systems. Similar patterns were found in vineyard GDC where E. carpini fluctuated at lower density than in the more vigorous P and S vineyards. Little is known on the ecology of E. carpini and the effect of canopy management on this spider mite needs further investigations. On the other hand P. ulmi was more abundant in vineyard FC than elsewhere. One could suggest that P. ulmi populations were allowed to increase in this vineyard because of the relatively low densities of E. carpini in 2009. High E. carpini densities in early season corresponded to relatively low P. ulmi populations in summer and vice versa (Duso and Pasqualetto, 1993). This topic also requires additional investigations.
In 2010 P. ulmi reached negligible densities but E. carpini persisted longer in three out four vineyards suggesting that this species is more difficult to be controlled by natural enemies. The low populations encountered in GDC vineyard confirm trends seen one year before. In both farms highest densities of spider mites were reached where only native predatory mites (i.e., A. andersoni and Ph. finitimus) were present. Spider mite population levels were, in some cases, close to or above the action thresholds (i.e., 5 and 10 motile forms per leaf for E. carpini and P. ulmi respectively; see Duso et al. 2012). Natural control of P. ulmi by A. andersoni has been associated with successful results in some studies (e.g., Camporese and Duso, 1996) but not in others (e.g., Duso, 1989). The impact of predatory insects seems to be more significant on P. ulmi than on E. carpini populations (Duso and Pasqualetto, 1993). Results of the present study confirm that problems with E. carpini persist despite the occurrence of A. andersoni (Duso and Vettorazzo, 1999) and Ph. finitimus (Duso, unpubl. data). The grape cultivars involved in our trials (Glera at Spresiano, Verduzzo trevigiano at Meolo) are characterized by pubescent leaf surfaces whereas A. andersoni shows a preference for glabrous leaf surfaces (Camporese and Duso, 1996). This could explain its weak response to spider mite populations. The A. andersoni population increase in late summer observed in some treatments, was probably due to the occurrence of Grape Downy Mildew that represents an alternative food for this species and can sustain population increase and persistence on plants in absence of prey (Duso et al. 2003; Pozzebon and Duso, 2008; Pozzebon et al. 2009, 2010). This phenomenon had a limited importance for spider mite control. The impact of Ph. finitimus on E. carpini populations in farm B appeared to be not significant despite the preference of this predatory mite for pubescent grape leaves (Duso and Vettorazzo, 1999). Probably, pesticides affected the performance by A. andersoni and Ph. finitimus in both farms. EBDC fungicides were used more frequently in farm B than in farm A but OPs were also applied in the latter. The use of chlorpyriphos, wettable sulphur and other fungicides (cyprodinil/fludioxonil) in July of 2009 was associated with A. andersoni population decline in farm A. The repeated use of mancozeb in farm B could be involved in similar problems. The impact of wettable sulfur on the survival and fecundity of predatory mites has been demonstrated in several experiments (Bernard et al. 2010). In the same experiments cyprodinil/fludioxonil and mancozeb were highly toxic to predatory mites.
Native predatory mite colonization patterns appeared also different in the four vineyards of farm A. In vineyards P and S, A. andersoni appeared to be more abundant than in the remaining vineyards. Differences may be due to higher E. carpini populations in these vineyards but also to their canopy features.
The releases of K. aberrans and T. pyri obtained successful results with some variation among farms and experimental years. In 2009 (farm A), P. ulmi densities were reduced in vineyard P (K. aberrans releases) and especially in vineyard FC (K. aberrans and T. pyri releases) where spider mites occurred at relatively high levels. In the same year and farm the impact of released predatory mites on E. carpini densities was less clear. Significant effects were obtained in vineyard GDC where E. carpini was not abundant. The weak response by K. aberrans and T. pyri to E. carpini contrasts with previous results obtained in the same area (e.g. Duso and Pasqualetto, 1993; Duso and Vettorazzo, 1999) and may be caused by the use of some pesticides. In fact in vineyards P, GDC and FC, K. aberrans and T. pyri populations declined in July after the use of the above mentioned pesticides. Resistance to OP insecticides and EBDC fungicides has been reported for T. pyri and K. aberrans (Van den Baan et al. 1985; Auger et al. 2004; Bonafos et al. 2007; Tirello et al. 2012; Cassanelli et al. 2015). Predatory mite strains used for releases are known to have low susceptibility to mancozeb and chlorpyriphos. It is likely that the early use of mancozeb as well as that of chlorpyriphos in summer had an impact on their populations due to possible sub-lethal effects induced by these products (Tirello et al. 2013; Duso et al. 2014; Pozzebon et al. 2011, 2014). At the same time, little is known about the effects of wettable sulphur and especially of cyprodinil/fludioxonil on these strains of K. aberrans and T. pyri. Laboratory studies will be conducted to investigate this topic.
Generally, increasing release densities had no significant effects on biological control in terms of spider mites abundance. However, in farm B where higher release densities were used, spider mites were controlled in the same season of the predator releases while a in farm A, spider mites reached the lowest densities in the second season after predator release. Nevertheless, since grapevines can tolerate a certain level of spider mite densities without serious losses (Girolami, 1981, 1987), strategies based on several release points within vineyards, with relatively low release densities should be preferred to those based on few points and higher densities. This could help a faster vineyard colonization by the released predatory mites.
Intraguild competition showed some clear trends. Results suggest that competition effects among predatory mites were not symmetric. In both farms and most cases A. andersoni densities were significantly reduced in K. aberrans and T. pyri release plots. The latter result contrasts with conclusions taken from laboratory studies on interspecific predation between A. andersoni and T. pyri (e.g., Croft and Croft, 1996; Croft et al. 1996). In these trials competition favored A. andersoni and this outcome was explained to be associated with its larger body size. Other factors, such as prey density, climatic conditions, leaf architecture or pesticide use can be involved in these interactions (Zhang and Croft, 1995; Croft and Croft, 1996; Croft et al. 1996; Pozzebon et al. 2015a,b). Our results confirm trends reported in a previous paper where the role of leaf morphology in affecting interspecific competition was emphasized (Duso and Vettorazzo, 1999). Differential effects of pesticides (particularly EBDCs and OPs) on predatory mites likely favored released predatory mites populations in contrast with A. andersoni. Moreover, recent studies showed that A. andersoni females have a less favorable conversion rate of food into eggs compared to the other predatory mites involved here, and this can explain the results obtained here (Lorenzon et al. 2015; Ahmad et al. 2015).
In farm B Ph. finitimus populations occurred at higher densities in early spring, decreased in summer but re-established moderate levels in late season. Trials with Ph. finitimus suggest that this species suffered less from interspecific competition than A. andersoni. Long term studies are required to understand the outcome of interspecific interactions involving this species (Duso and Vettorazzo, 1999). The role of EBDCs in affecting competition should be investigated more in depth.
The competition between K. aberrans and T. pyri in farm A gave interesting results. In 2009, the presence of T. pyri in mixed release plots (Ka-Tp) affected K. aberrans densities in two out of four vineyards (GDC and FC). On the other hand, the presence of K. aberrans also affected T. pyri numbers in two out of four vineyards (P and GDC). In 2010 the presence of K. aberrans reduced that of T. pyri in vineyards P and S but not in vineyards GDC and FC where predatory mites reached low population densities. In contrast the occurrence of T. pyri had no effects on that of K. aberrans.
Results also suggest that interspecific interactions are prey mediated: K. aberrans outcompeted T. pyri where E. carpini was abundant (vineyards P in 2009 and P and S in 2010), while T. pyri outcompeted K. aberrans where P. ulmi was more abundant (vineyard FC in 2009). GDC vineyard represents an interesting case study because densities of both spider mite species were reduced. In GDC both spider mites were found at moderate levels. This aspect can be related to intraguild predation between predatory mites. Theoretical studies predict that coexistence and thus persistence of intraguild predators are likely at intermediate level of common prey availability and when the intraguild prey is a superior competitor for the shared resource than the intraguild predator (Polis et al. 1989, Holt and Polis, 1997, Diehl and Feissel, 2001, Mylius et al. 2001). In this case we did not observe the extinction of one of the two predators, but we observed fluctuations in equilibrium level. According to life-history traits as a measure of prey conversion into offspring, T. pyri appeared to be a superior competitor on both prey (Lorenzon et al. 2012). However, interspecific interaction results confirm the capability of K. aberrans to compete with heterospecific predatory mites reported in other studies (Duso, 1989; Duso and Camporese, 1991; Duso and Pasqualetto, 1993; Duso and Vettorazzo, 1999). In the latter K. aberrans displaced T. pyri after 2-4 years from their releases, in particular when spider mite densities declined. The persistence of spider mites in the second year of the present study (see farm A) could have slowed down this process. Body size and feeding specialization are considered important factors in interspecific competition (Croft and Croft, 1993, 1996; Croft et al. 1996). Studies on interspecific predation between T. pyri and K. aberrans did not suggest a definite trend (Schausberger, 1997, 1999; Lorenzon et al. 2012, 2015; Ahmad et al. 2015). Factors affecting the success of K. aberrans over T. pyri in north-east Italian vineyards and orchards remain partly unexplained (Duso et al. 2009). Kampimodromus aberrans could be more effective than T. pyri to find shelters to escape to competitors. Indeed the exploitation of food and spatial resources can play an interactive role in shaping the outcome of intraguild predation among predatory mites (Pozzebon et al. 2015; Calabuig et al. 2018). This topic should be further explored in the future to clarify the competition between T. pyri and K. aberrans.
We can conclude that releases of predatory mites resistant to pesticides are successful spider mites control tactics that could be included in IPM strategies in vineyards. Released predatory mites were able to outcompete native species improving spider mites control.
We thank Dr. Michele Borgo (formerly Director of the Istituto Sperimentale per la Viticoltura, Conegliano) for the cooperation. This work has been partially supported by province of Treviso and by project BIRD167802/16 to A.P.
Ahmad S., Pozzebon A., Duso C. 2015. Predation on heterospecific larvae by adult females of Kampimodromus aberrans, Amblyseius andersoni, extslTyphlodromus pyri
and Phytoseius finitimus (Acari: Phytoseiidae). Exp. Appl. Acarol., 67: 1-20. doi:10.1007/s10493-015-9940-1Auger P., Bonafos R., Kreiter S. 2004. Mancozeb resistance patterns among Kampimodromus aberrans and Typhlodromus pyri strains from French vineyards. Can. Entomol. 136:663-673. doi:10.4039/n03-115 ![]()
Baillod M., Schmid A., Guignard E., Antonin P.H., Caccia R. 1982. Lutte biologique contre l'acarien rouge en viticulture. II. Equilibres naturals, dynamiques de population et experiences de làchers de typhlodromes. Revue Suisse Vitic. Arboric. Hortic., 14, 6: 345–352.
Bernard M.B., Cole P., Kobelt A., Horne P.A., Altmann J., Wratten S.D., Yen A.L. 2010. Reducing the Impact of Pesticides on Biological Control in Australian Vineyards: Pesticide Mortality and Fecundity Effects on an Indicator Species, the Predatory Mite Euseius victoriensis (Acari: Phytoseiidae). J. Econom. Ent., 103: 2061–2071. doi:10.1603/EC09357 ![]()
Bertamini M., Tardaguila J., Iacono F. 1994. Valutazione dell'equilibrio vegeto-produttivo e microclimatico del vigneto per l'ottimizzazione delle tecniche colturali a verde: aspetti teorici e pratici. Bollettino ISMA, 2: 24–40.
Bonafos R., Serrano E., Auger P., Kreiter S. 2007. Resistance to deltamethrin, lambda-cyhalothrin and chlorpyriphos-ethyl in some populations of Typhlodromus pyri Scheuten and Amblyseius andersoni (Chant) (Acari: Phytoseiidae) from vineyards in the south-west of France. Crop. Prot. 26:169-172 doi:10.1016/j.cropro.2006.10.001 ![]()
Calabuig A., Pekas T., Wäckers F. L. 2018. The quality of nonprey food affects cannibalism, intraguild predation, and hyperpredation in two species of phytoseiid mites. J Econ. Entomol., 111 (1): 72–77, doi:10.1093/jee/tox301 ![]()
Camporese P., Duso C. 1996. Different colonization patterns of phytophagous and predatory mites (Acari: Tetranychidae, Phytoseiidae) on three grape varieties: a case study. Exp. Appl. Acarol., 20: 1–22.
Cassanelli S., Ahmad S., Duso C., Tirello P., Pozzebon A. 2015. A single nucleotide polymorphism in the acetylcholinesterase gene of the predatory mite Kampimodromus aberrans (Acari: Phytoseiidae) is associated with chlorpyrifos resistance. Biol. Control, 90: 75-82. doi:10.1016/j.biocontrol.2015.05.015 ![]()
Croft B.A., Croft M.B. 1996. Intra- and interspecific predation among adult female phytoseiid mites (Acari: Phytoseiidae): effects on survival and reproduction. Environ. Entomol., 25: 853–858. doi:10.1093/ee/25.4.853 ![]()
Croft B.A., Croft M.B. 1993. Larval survival and feeding by immature Metaseiulus occidentalis, Neoseiulus fallacis, Amblyseius andersoni and Typhlodromus pyri on life stage groups of Tetranychus urticae Koch and phytoseiid larvae. Exp. Appl. Acarol., 17: 685–693. doi:10.1007/BF00058508 ![]()
Croft B.A., Kim S.S., Kim D.I. 1996. Intra- and interspecific predation on four life stage groups by the adult females of Metaseiulus occidentalis, Typhlodromus pyri, Neoseiulus fallacis and Amblyseius andersoni (Acari: Phytoseiidae). Exp. Appl. Acarol., 20: 435–444. doi:10.1007/BF00053307 ![]()
Croft B.A., McRae I.V.1993. Biological control of apple mites. Impact of Zetzellia mali (Acari, Stigmaeidae) on Typhlodromus pyri and Metaseiulus occidentalis (Acari, Phytoseiidae). Environ. Entomol. 22: 865–887. doi:10.1093/ee/22.4.865 ![]()
Diehl S., Feissel M. 2001. Intraguild prey suffer from enrichment of their resources: a microcosm experiment with ciliates. Ecology, 82(11): 2977-2983.
Duso C. 1989. Role of the predatory mites Amblyseius aberrans (Oud.), Typhlodromus pyri Scheuten and Amblyseius andersoni (Chant) (Acari, Phytoseiidae) in vineyards. I. The effects of single or mixed phytoseiid population releases on spider mite densities (Acari, Tetranychidae). J. Appl. Entomol., 107: 474–492. doi:10.1111/j.1439-0418.1989.tb00283.x ![]()
Duso C. 1992. Role of predatory mites Amblyseius aberrans (Oudemans), Typhlodromus pyri Scheuten and Amblyseius andersoni (Chant) (Acari: Phytoseiidae) in vineyards. III. Influence of variety characteristics on the success of A. aberrans and T. pyri. J. Appl. Entomol., 114: 455–462. doi:10.1111/j.1439-0418.1992.tb01151.x ![]()
Duso C., Ahmad S., Tirello P., Pozzebon A., Klaric V., Baldessari M., Malagnini V., Angeli G. 2014. The impact of insecticides applied in apple orchards on the predatory mite Kampimodromus aberrans (Acari Phytoseiidae). Exp. Appl. Acarol., 62: 391-414. doi:10.1007/s10493-013-9741-3 ![]()
Duso C., Camporese P. 1991. Developmental times and oviposition rates of predatory mites Typhlodromus pyri and Amblyseius andersoni (Acari, Phytoseiidae) reared on different foods. Exp. Appl. Acarol., 13: 117–128. doi:10.1007/BF01193662 ![]()
Duso C., Fanti M., Pozzebon A., Angeli G. 2009. Is the predatory mite Kampimodromus aberrans a candidate for the control of phytophagous mites in European apple orchards? BioControl, 54: 369–382. doi:10.1007/s10526-008-9177-6 ![]()
Duso C., Girolami V., Borgo M., Egger E. 1983. Influenza di anticrittogamici diversi sulla sopravvivenza di predatori Fitoseidi introdotti su vite. Redia, 66: 469–483.
Duso C., Pasqualetto C. 1993. Factors affecting the potential of phytoseiid mites (Acari: Phytoseiidae) as biocontrol agents in North-Italian vineyards. Exp. Appl. Acarol., 17: 241–258. doi:10.1007/BF02337274 ![]()
Duso C., Pasqualetto C., Camporese P. 1991. Role of the predatory mites Amblyseius aberrans (Oud.), Typhlodromus pyri Scheuten and Amblyseius andersoni (Chant) (Acari, Phytoseiidae) in vineyards. II. Minimum releases of A. aberrans and T. pyri to control spider mite populations (Acari, Tetranychidae). J. Appl. Entomol., 112: 298–308. doi:10.1111/j.1439-0418.1991.tb01059.x ![]()
Duso C., Pozzebon A., Capuzzo C., Bisol P.M., Otto S. 2003. Grape downy mildew spread and mite seasonal abundance in vineyards: Evidence for the predatory mites Amblyseius andersoni and Typhlodromus pyri. Biol. Control, 27: 229–241. doi:10.1016/S1049-9644(03)00016-1 ![]()
Duso C., Pozzebon A., Kreiter S., Tixier M.S., Candolfi M.P., 2012. Management of phytophagous mites in European vineyards. In: Bostanian, N.J., Vincent, C., Isaacs, R. (Eds) Arthropod management in vineyards: pests, approaches, and future directions. New York: Springer. p. 191–217. doi:10.1007/978-94-007-4032-7_9 ![]()
Duso C., Vettorazzo E. 1999. Mite popultion dynamics on different grape varieties with or without phytoseiids released (Acari: Phytoseiidae). Exp. Appl. Acarol., 23: 741–763. doi:10.1023/A:1006297225577 ![]()
Engel R., Ohnesorge B. 1994. Die Rolle von Ersatznahrung und Mikroklima im System Typhlodromus pyri Scheuten (Acari, Phytoseidae). Panonychus ulmi Koch (Acari, Tetranychidae) auf Weinreben I. Freilandversuche. J. Appl. Entomol., 118: 224 –238. doi:10.1111/j.1439-0418.1994.tb00798.x ![]()
English-Loeb G., Norton A., Walker M. 2002. Behavioral and population consequences of acarodomatia in grapes on phytoseiid mites (Acari: Mesostigmata) and implications for plant breeding. Ent. Exp. Appl., 104: 307–319. doi:10.1046/j.1570-7458.2002.01017.x ![]()
Facchin P. 1996. Lotta biologica agli acari fitofagi della vite attraverso l'introduzione degli acari predatori Kampimodromus aberrans (Oudemans), Phytoseius finitimus Ribaga, Typhlodromus pyri Sheuten (Acari: Phytoseiidae). Degree Thesis. University of Padua.
Girolami V. 1981. Danni, soglie di intervento, controllo degli acari della vite. La difesa integrata della vite, 3-4. 1981, Latina: 111–143.
Girolami V., Picotti P., Coiutti C. 1992. Ruolo determinante del fitoseide Amblyseius aberrans (Oud.) nel controllo degli acari fitofagi. Inf.tore agr., XLVIII: 65–69.
Holt R. D., Polis G. A. 1997. A theoretical framework for intraguild predation. American Naturalist, 149(4):745-764. doi:10.1086/286018 ![]()
Ivancich Gambaro P. 1973. Il ruolo del Typhlodromus aberrans Oud. (Acarina Phytoseiidae) nel controllo biologico degli acari fitofagi nel Veronese. Boll. Zool. Agr. Bachic., 11:151–165.
Jolliffe P.A. 2000. The replacement series. J. Ecol., 88: 371-385. doi:10.1046/j.1365-2745.2000.00470.x ![]()
Littell R. C., Henry P. R., Ammerman C. B. 1998. Statistical analysis of repeated measures data using SAS procedures. J. Ani. Sci., 76(4): 1216-1231. doi:10.2527/1998.7641216x ![]()
Lorenzon M., Pozzebon A., Duso C. 2015. Feeding habits of overwintered predatory mites inhabiting European vineyards. BioControl, 60: 605–615.
Moret R. 2006. Effetti principali e collaterali di insetticidi destinati al controllo di "Empoasca vitis". Degree Thesis. University of Padua.
Mylius S.D., Klumpers K., de Roos A.M., Persson L. 2001- Impact of intraguild predation and stage structure on simple communities along a productivity gradient. American Naturalist 158(3):259-276. doi:10.1086/321321 ![]()
Polis G.A., Myers C.A., Holt R.D. 1989. The ecology and evolution of intraguild predation: potential competitors that eat each other. Annu Rev Ecol Syst 20: 297-330. doi:10.1146/annurev.es.20.110189.001501 ![]()
Posenato G. 1994. Popolazioni di Amblyseius aberrans (Oud.) resistenti ad esteri fosforici e ditiocarbammati. Inf.tore agr., 50: 41–43.
Pozzebon A., Ahmad S., Tirello P., Lorenzon M., Duso C. 2014. Does pollen availability mitigate the impact of pesticides on generalist predatory mites?. BioControl, 59, 585–59. doi:10.1007/s10526-014-9598-3 ![]()
Pozzebon A., Borgo M., Duso C. 2010. The effects of fungicides on non-target mites can be mediated by plant pathogen. Chemosphere, 79: 8–17.
Pozzebon A., Duso C. 2008. Grape downy mildew extslPlasmopara viticola
, an alternative food for generalist predatory mites occurring in vineyards. Biol.Control, 45: 441–449. doi:10.1016/j.biocontrol.2008.02.001Pozzebon A., Duso C., Tirello P., Bermudez Ortiz P. 2011. Toxicity of thiamethoxam to Tetranychus urticae Koch and Phytoseiulus persimilis Athias-Henriot (Acari Tetranychidae, Phytoseiidae) through different routes of exposure. Pest Manag. Sci., 67: 352–359. doi:10.1002/ps.2072 ![]()
Pozzebon A., Loeb G.M., Duso C. 2009. Grape powdery mildew as a food source for generalist predatory mites occurring in vineyards: effects on life-history traits. Ann. Appl. Biol., 155: 81-89. doi:10.1111/j.1744-7348.2009.00323.x ![]()
Pozzebon A., Loeb G.M., Duso C. 2015. Role of supplemental foods and habitat structural complexity in persistence and coexistence of generalist predatory mites. Sci. Rep., 5: 14997-14997. doi:10.1038/srep14997 ![]()
Pozzebon A., Tirello T., Moret R., Pederiva M., Duso C. 2015. A fundamental step in IPM on grapevine: Evaluating the side effects of pesticides on predatory mites. Insects, 6: 847-857. doi:10.3390/insects6040847 ![]()
Prischmann D. A., James D. G., Wright L. C., Snyder W. E. 2006. Effects of generalist phytoseiid mites and grapevine canopy structure on spider mite (Acari: Tetranychidae) biocontrol – Env. Entomol., 35(1): 56-67.Rosenheim J.A., Kaya H.K., Ehler L.E., Marois J.J., Jaffee B.A. 1995. Intraguild predation among biological control agents: theory and evidence. Biol. Control, 5: 303–335.
Schausberger P. 1997. Inter- and intraspecific predation on immatures by adult females in Euseius finlandicus, Typhlodromus pyri and Kampimodromus aberrans (Acari, Phytoseiidae). Exp. Appl. Acarol., 21: 131–150. doi:10.1023/A:1018478418010 ![]()
Schausberger P. 1999. Predation preference of Typhlodromus pyri and Kampimodromus aberrans (Acari, Phytoseiidae) when offered con- and heterospecific immature life stages. Exp. Appl. Acarol., 23: 389–398. doi:10.1023/A:1006172320540 ![]()
Schruft G. 1985. Grape. In: Helle W., Sabelis M.W. (Eds.) Spider mites 1B. Their biology, natural enemies and control. Amsterdam: Elsevier. p. 359–365.
Seelmann L., Auer A., Hoffmann D., Schausberger P. 2007. Leaf pubescence mediates intraguild predation between predatory mites. Oikos, 116: 807–817.
Tirello P., Pozzebon A., Duso C. 2013. The effect of insecticides on the non-target predatory mite Kampimodromus aberrans: laboratory studies. Chemosphere, 93: 1139-1144. doi:10.1016/j.chemosphere.2013.06.046 ![]()
Tirello P., Pozzebon A., Duso, C. 2012. Resistance to chlorpyriphos in the predatory mite Kampimodromus aberrans. Exp. Appl. Acarol., 56: 1-8. doi:10.1007/s10493-011-9486-9 ![]()
Tixier M. S., Baldassar A., Duso C., Kreiter S. 2013. Phytoseiidae in European grape (Vitis vinifera L.): bio-ecological aspects and keys to species (Acari: Mesostigmata). Zootaxa, 3721(2): 101-142. doi:10.11646/zootaxa.3721.2.1 ![]()
Van de Baan H.E., Kuijpers L.A.M., Overmeer W.P.J., Oppenoorth F.J. 1985. Organophosphorus and carbamate resistance in the predacious mite Typhlodromus pyri due to insensitive acetylcholinesterase. Exp. Appl. Acarol. 1 (1):3-10. doi:10.1007/BF01262194 ![]()
Wilson J.W. 1963. Estimation of foliage denseness and foliage angle by inclined "point quadrat". Aust. J. Bot., 11, 1: 95–105. doi:10.1071/BT9630095 ![]()
Zang Z.Q., Croft B.A. 1995. Intraspecific competition in immature Amblyseius fallacies, Amblyseius andersoni, Typhlodromus occidentalis and Typhlodromus pyri (Acari: Phytoseiidae). Exp. Appl. Acarol., 19: 65–77. doi:10.1007/BF00052547 ![]()



2016-11-27
Date accepted:
2018-05-24
Date published:
2018-09-28
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Lorenzon, Mauro; Pozzebon, Alberto and Duso, Carlo
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)