



Effects of Aculus schlechtendali (Acari: Eriophyidae) population densities on Golden Delicious apple production
Simoni, Sauro1 ; Angeli, Gino2 ; Baldessari, Mario3 and Duso, Carlo4
1✉ CREA-DC Council for Agricultural Research and Economics - Research centre for Plant Protection and Certification, via Lanciola 12/a, 50125, Florence (Firenze), Italy.
2FEM , Via E. Mach 1, 38010 - San Michele all’Adige (Trento), Italy.
3FEM , Via E. Mach 1, 38010 - San Michele all’Adige (Trento), Italy.
4Department DAFNAE, University of Padova, viale dell’Università 16, 32050 Legnaro (Padova), Italy.
2018 - Volume: 58 Issue: Suppl pages: 134-144
https://doi.org/10.24349/acarologia/20184276Keywords
Abstract
The apple rust mite (ARM) Aculus schlechtendali Nalepa (Acari Eriophyidae) occurs in the main world apple growing areas (e.g., Jeppson et al. 1975, Easterbrook 1996, Easterbrook and Palmer 1996, Li and Cai 1996) but it has been considered an important pest of apple especially in Europe (Ciampolini et al. 1976, Easterbrook 1991, Duso et al., 2010). ARM feeds on flowers, leaves and fruits producing various symptoms. Seriously damaged apple leaves show rusty under surfaces, roll up longitudinally and fall prematurely (Herbert 1974, Kozłowski 1980). ARM feeding on developing fruitlets induces russet on fruits while fruit discoloration can be observed on certain cultivars. The intensity of damage can be affected by ARM population size, cultivar and plant age (Herbert 1974, Ciampolini et al. 1976, Easterbrook and Fuller 1986).
The variability of ARM effects on fruit production has been matter of discussion but the knowledge of effects of ARM on plant physiological processes is limited. According to Kozłowski (1980) and Solhoy et al. (1991), a decrease in photosynthesis was observed when apple leaves were infested in summer but implications for apple yields were not clear.
The most comprehensive studies on this topic were probably those carried out by Spieser et al. (1998, 1999). They evaluated the effects of ARM populations on gas exchange, leaf colour, and leaf tissue features on young apple plants of Jonagold and Golden Delicious cultivars. Plants were infested with ARM and then treated with pesticides or left untreated to obtain different mite densities, i.e. 4,000-5,000 mites/leaf in mid-June on untreated plants while very low populations on treated plants. Infested leaves became intense brown, and consequently net CO2 exchange and transpiration rates were reduced. Detailed observations showed several puncture wounds on epidermal cells, the partial opening of cells stomata and the lost in turgor of guard cells. Damage on leaf epidermis caused by ARM can imply additional negative effects on leaf mesophyll. A number of epidermal cells and spongy parenchyma cells were desiccated. According to Spieser et al. (1998, 1999) the effects of ARM were more severe on Jonagold than on Golden Delicious. These observations confirmed those conducted by Kozłowski and Zielinska (1997) on other apple cultivars. In other studies (Easterbrook and Palmer 1996) the impact of ARM populations on photosynthesis proved to be less severe; this discrepancy can be explained by the different size in populations considered in these studies. Ioriatti et al. (1996) did not obtain significant differences in terms of net photosynthesis and transpiration rates comparing the effects of two different ARM population levels on insecticide-treated and untreated apple (cv. Red Top) plots.
The biology, life-history and ethology of ARM have been extensively investigated (e.g. Herbert 1974, Ciampolini et al. 1976, Easterbrook 1979, 1984, Kozłowski 1980). However, more detailed information on the relationship between ARM abundance and yield or fruit quality is required. Among several factors which can influence fruit yield, fruit size, and fruit quality, e.g. variety susceptibility (Danelski et al. 2015), the phenological phase involved during ARM infestation, population size and climate conditions can be mentioned. It should be crucial to establish, for each cultivar, the ARM population level that can be tolerated without significant losses in production and quality.
Observations were conducted in an experimental orchard to identify trends in ARM abundance over the growing season and evaluate the impact of different ARM population densities on yield and quality parameters of Golden Delicious, the most important apple variety cultivated in Europe. We manipulated pesticide applications to generate different ARM population sizes in definite plots. We investigated possible correlations between ARM cumulative densities and yield external (i.e. fruit size, fruit weight, ground colour hue, russet on fruits) and internal quality parameters of fruits (i.e. soluble solids, malic acidity and related indices). Since mite damage may affect yield efficiency and return bloom, these parameters were also assessed. Preliminary results were reported by Angeli et al. (2007). In addition, laboratory tests were performed to assess the potential for ARM population increase at temperatures similar to those registered in field trials to better understanding the ecology of ARM and the potential impact of its populations on fruit production.
The seasonal abundance of ARM and the mite density-damage relationships were investigated in an apple orchard located in the Non Valley (Pavillo, 530 m asl, 46°22′0″ N, 11°2′0″ E), Trento province, North-eastern Italy. This orchard comprises Golden Delicious plants (2500 plants per hectare), five years old grafted on Golden clone B seedlings, and trained with the Spindle bush system. This orchard is irrigated over-canopy and fertilized annually with 40 kg N, 36 kg P2O5 and 60 kg K2O.
Observations were conducted on four plots (50 trees/plots) treated with fungicides (e.g. dithianon) having negligible to low effects on insects and mites, from April to September in three subsequent seasons (Y1, Y2 and Y3). Preliminary leaf samples showed considerable variation in mite densities even along the same shoot. Therefore, we selected as sample units the 4th-5th leaves from the shoot apex to reduce variability. A total of 80 leaves (20 leaves per plots) were removed and analysed for investigating ARM seasonal abundance. Since the latter can be affected by predators, additional observations were conducted on natural antagonists of rust mites mainly predatory mites. In preliminary samples the occurrence of Phytoseiidae was commonly reported. Therefore their populations were also monitored by removing 25 leaves per plot every month, from May to August of seasons Y1, Y2 and Y3. Leaves were transferred to the laboratory and analysed under a dissecting microscope. Predatory mites were mounted on slides in Hoyer medium and then identified under a phase contrast microscope. Monthly average values for temperature and rainfall were registered.
The impact of ARM on apple yield and quality parameters was investigated in Y1 and Y2 in the same orchard adopting a randomized block design. Different mite population sizes were generated by using different pesticides (Table 1). In particular, three treatments were compared: 1) trees untreated with acaricides or fungicides having side-effects on rust mites; 2) trees treated with wettable sulphur known for having a moderate acaricide effect (four applications starting 15 days from petal fall on a week base); 3) trees treated with abamectin. Each treatment comprised four replicates of 15 plants. Samples of 40-60 leaves/treatment were taken from May to September, following previously described procedures, and starting when shoots had five to six fully developed leaves, to evaluate ARM population densities. Leaves sampled were dipped into a detergent solution (Agral® 5%, 30 cc/hl) and kept overnight (12 hours) at room temperature (Boller 1984). This solution was filtered through a 15 µm mesh cloth and the number of mites was subsequently estimated under a dissecting microscope (Höhn and Höpli 1990). The mite load was expressed as motile forms per leaf. The mean Golden Delicious leaf surface measured 48.44±12.89 cm². The cumulative mite-day parameter (CMD) was also considered (Hoyt et al. 1979, Beers and Bruner 1999), and expressed as:egincenterCMD = Σ0.5(Pa + Pb)Db-andcenterwhere Pa is the population density (mean mites per leaf) at time a, Pb is the population density at time b, Db-a is the number of days between two subsequent samplings.


The following parameters were evaluated at harvest in the three treatments: a) productivity expressed as kg of apples per tree per cm² of trunk section; b) weight (g/fruit); c) size (mm Ø: <70, 70-80 and >80); d) round colour hue (% fruit surface green); e) russeting (surface of fruits with >10% russeting). The same apples considered for the evaluation of external quality were analysed to assess the internal quality parameters, i.e. soluble solids content (°Brix), acidity (malic acid g/l) and Perlim index (PI=kg/cm2*0.5+°Brix*0.67+malic acid g/l*0.67). The influence of ARM densities on the return bloom of the subsequent year (% flower buds/total buds) was also evaluated.
The potential increase of ARM populations was evaluated following two approaches, 1) on field data collected in Y1 and Y2, or 2) on laboratory data. In the first approach, the intrinsic rate of increase was calculated from six ARM CMDs registered in Y1 and Y2. The formula adopted was:egincenterr = ln Nt – ln N0/t ndcenterwhere r is the intrinsic rate of increase, N is the population size at the start of the observations (N0) and at the time (days) considered (Nt) (Birch 1948, Turchin 2003). In the second approach, the laboratory assessment of intrinsic rates of ARM was performed in CREA-ABP facilities of Florence in 2014, using environmental chambers (Angelantoni, mod. CHL 1500) under controlled conditions. Experiments lasted 100 days from April to July. Every ten days, a constant temperature was settled up on the basis of the decade mean temperature registered in Y1 and Y2, respectively; RH ranged from 60 to 80%, while photoperiod was 16-8 LD. A total of 25 immatures collected from ARM infested apple leaves were placed on clean apple leaves, placed on wet cotton, to obtain young females. When the adult stage was reached, 10 females were left onto a clean leaf, considered an increase unit, to produce progeny for 100 days. Experiments were conducted at temperatures based on the decade mean temperatures experienced in the growing seasons Y1 and Y2. In this approach, the calculation of the intrinsic rate was developed by using the cited formula; by each Year considered, values for rm were calculated by following the increase in ARM abundance of nine increase units.When leaves were deteriorated ARM motile forms were transferred on fresh leaves.
In the study on mite seasonal abundance a Pearson correlation analysis was performed to identify an association between cumulative densities (CMD) of ARM and predatory mites. In the study on the impact of ARM on apple yield and quality a Multivariate Analysis (GLM, general Linear Model) was performed to evaluate the effect of year (Y1 and Y2) and ARM densities on external (i.e., fruit size, fruit weight, round colour hue, russeting) and internal (i.e., soluble solids and malic acid contents) apple quality parameters. The effects of ARM densities on yield efficiency and return bloom were evaluated following a similar approach. Perlim index depends on a number of above mentioned parameters and thus such records were not statistically analysed. Concerning the significant factors emerged, a post hoc comparison analysis was applied using one-way ANOVA. Percentage data were arc-sin transformed and HSD Tukey range test (P<0.05) was used for post hoc comparisons of means. Furthermore, a regression analysis (linear and exponential) was performed to highlight relationships between ARM cumulative densities and each of selected parameters. All statistical procedures were performed using the SPSS-package (SPSS, 2009) while the calculation of the intrinsic rate for population increase was performed using the spreadsheet Excel (Microsoft for Windows, 2010) program.
The monthly mean temperatures and cumulative rainfall recorded in the three experimental years are reported in Figure 1. The highest monthly temperature was registered in July (Y2 and Y3) or in August (Y1). Mean temperatures from May to July appeared to be higher in Y3 than in Y1 but an opposite situation was observed in August. Rainfall appeared to be rather homogeneous across the years with the exception of August in Y2. Regarding temperature, cumulative mean degrees appeared to be similar over the seasons (data not shown).

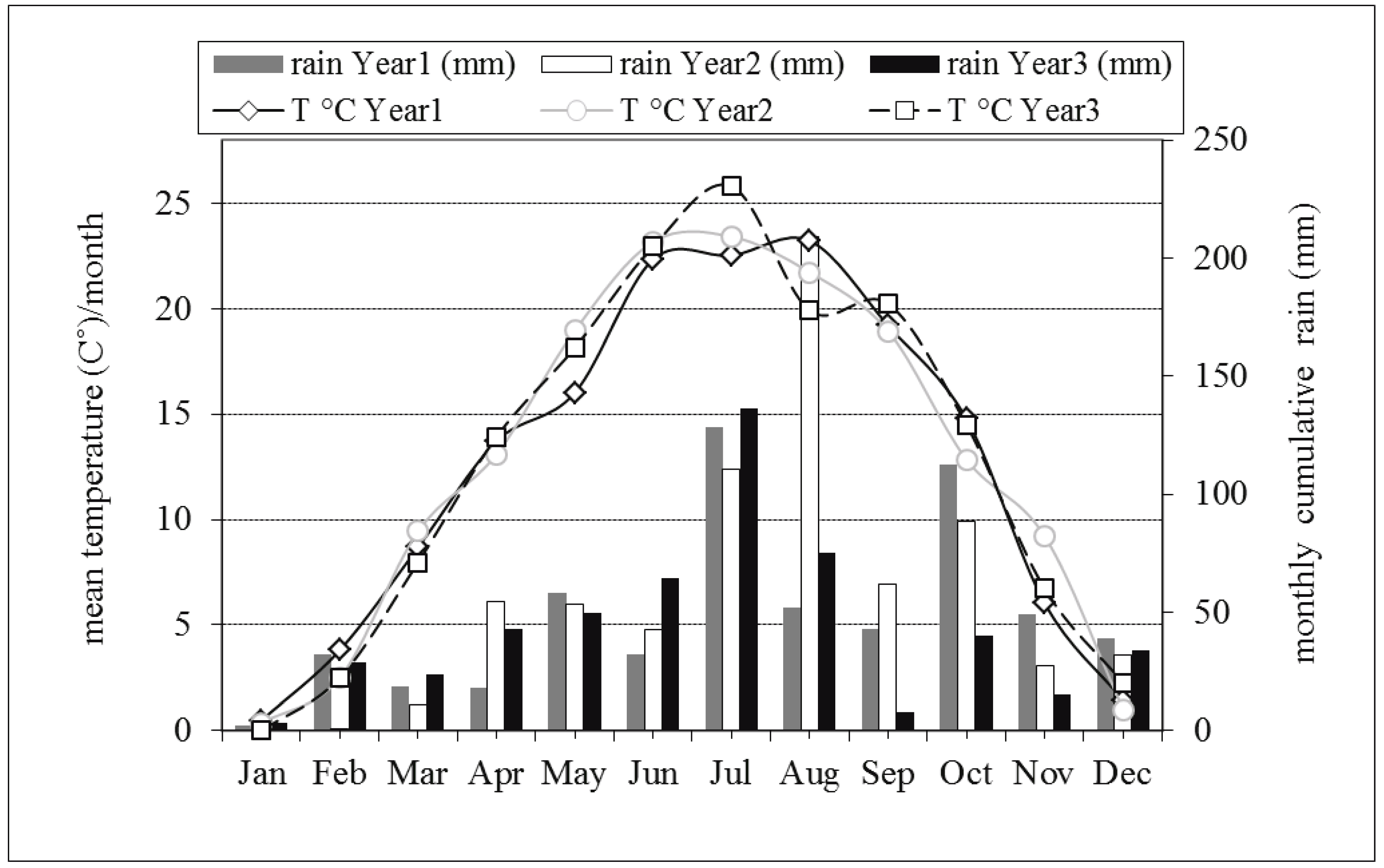
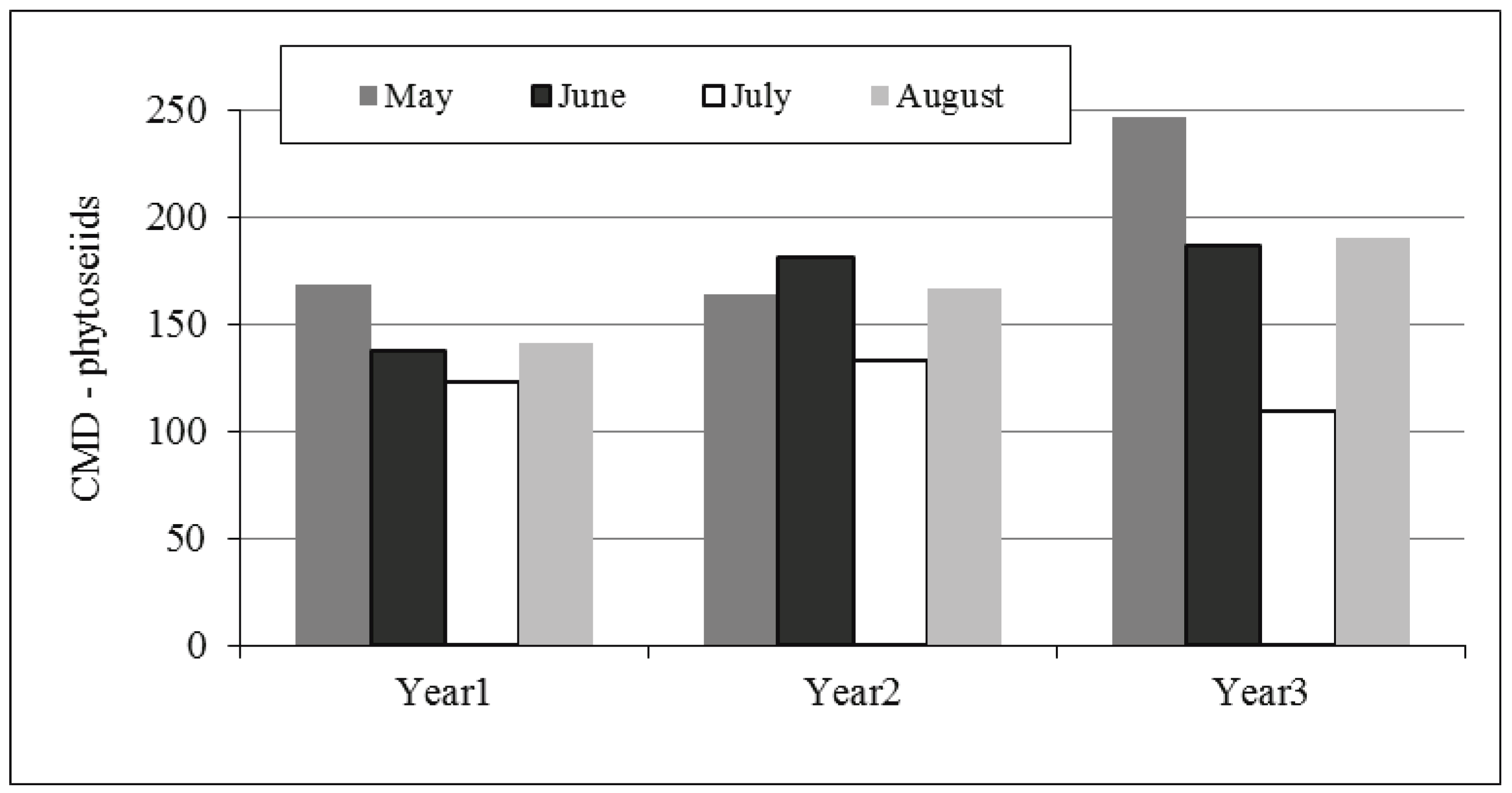
In the three years, ARM densities peaked in July (Figure 2). Mite populations showed an exponential growth in June in both Y1 and Y3 while reached low levels in Y2. ARM population densities varied significantly (F = 493.45; P < 0.001) over the seasons and cumulative densities (ARM-CMDs) were significantly higher in Y1 and Y3 than in Y2 (Table 2). There were no differences between ARM-CMDs recorded in Y1 and Y3 (F = 0.913; P = 0.426). Among predatory mites belonging to the Phytoseiidae family, Typhlodromus pyri Scheuten was largely dominant during the three years. Phytoseiid mite population levels expressed as cumulative mite-days (TP-CMDs) did not vary significantly across the seasons (F = 1.261; P = 0.329) despite differences in ARM-CMDs (Table 2). Phytoseiid mite numbers appeared to be higher in May both in Y1 and Y3 (Figure 3). The lowest phytoseiid densities were observed in July whereas they increased again in August. In Y2 the highest TP-CMDs were recorded in June. No significant linear trend within the yearly period was in the population dynamics (fit of linear regression: P 2004 = 0.35; P 2005 = 0.75; P 2006 = 0.44).
Predatory mites and ARM numbers were correlated (Pearson Correlation = – 0.58; P = 0.048; N = 36) as expected.




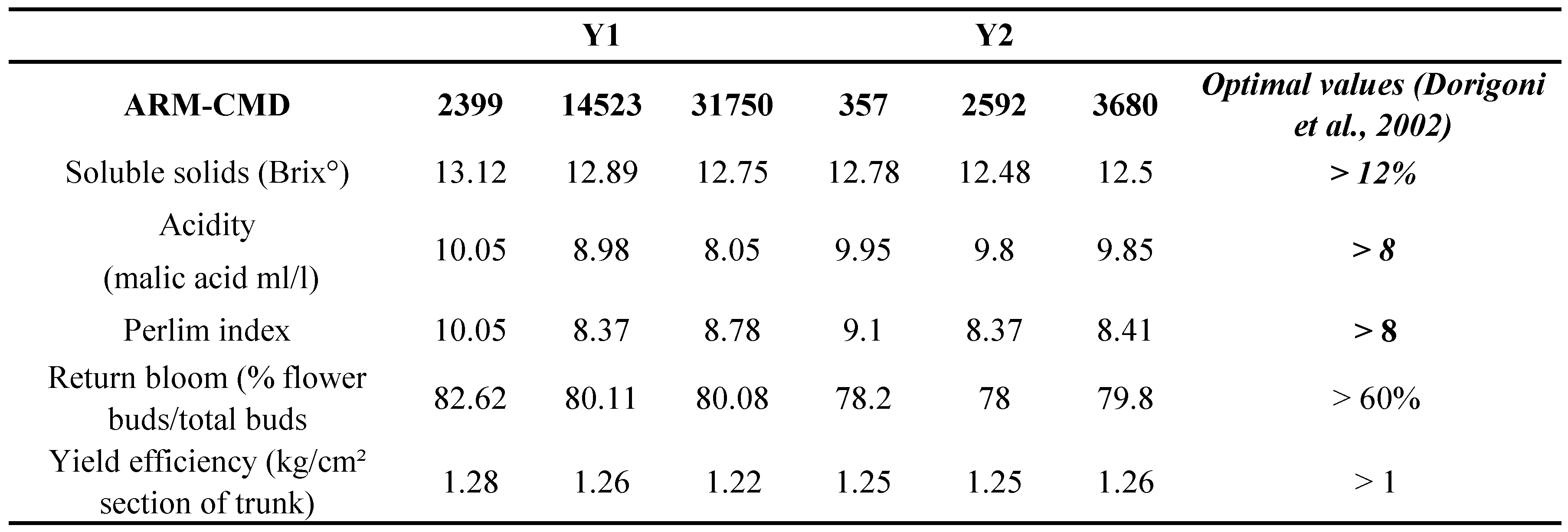
The model here adopted was highly significant ( P < 0.0001), and both year ( P < 0.0001) and ARM density ( P = 0.002) affected a number of fruit parameters, especially those related to external quality (Tables 3 and 4). In Y1, ARM-CMDs range in the three treatments was broader than in Y2, resulting 2,399 in treatment 1 (abamectin), 14,523 in treatment 2 (wettable sulphur), and 31,750 in treatment 3 (untreated control) (Table 3). In Y1, treatments 2 and 3 were associated to a lower production of apples of optimal size (Ø > 80 mm) ( F = 7.51; P = 0.002) and a higher production of apples under optimal size (Ø < 70 mm) ( F = 7.66; P < 0.001) compared to treatment 1 (Table 3). Fruit weight values showed a similar pattern ( F = 12.64; P < 0.001). The round colour hue of fruits was negatively affected in treatment 3 ( F = 10.96; P < 0.001) even if all scores exceeded the reference ones. In contrast, fruit russeting was not affected by different ARM-CMDs ( F = 0.16; P = 0.854; Table 3). A slightly decreasing trend was observed as concerns soluble solids as well as malic acid contents (Table 4); these values decreased with ARM-CMDs increased, but differences were not significant ( F = 0.745; P = 0.502; F = 1.78; P = 0.08, respectively for soluble solids and malic acid contents). A similar trend was observed for return bloom ( F = 3.52; P = 0.31). No significant effects were observed in yield efficiency ( F = 0.076; P = 0.926). However, all these parameters satisfied optimal values as, on the whole, Perlim index was always higher than the optimal value (see Dorigoni et al., 2002) (Table 3).



ARM-CMDs values in Y2 were lower than those observed in Y1, i.e. 357, 2592 and 3680 in treatments 1, 2 and 3, respectively (Table 3). A lower number of fruits with a diameter exceeding 80 mm (F = 8.3; P < 0.001) and a higher number of fruits with suboptimal size (F = 16.8; P < 0.001) were recorded in treatments 2 and 3 than in treatment 1 (Table 2). A similar tendency concerned round colour hue (F = 107.6; P < 0.0001; Table 2). Fruit weight was reduced in treatment 3 but not in treatment 2 (F = 5.97; P = 0.024; Table 2). Fruit russeting was not affecting by different ARM-CMD (F = 0.30; P = 0.743; Table 2). Soluble solids content and acidity were not significantly affected by mite densities (F = 1.234; P = 0.336 and F = 0.187; P = 0.833, respectively; Table 3) and fulfilled optimal values. Perlim index showed similar patterns. Productive efficiency and return bloom in the following year were not affected (F = 1.259; P = 0.277; F = 0.711; P = 0.585, respectively) by mite densities (Table 4).

Fitting analysis of fruit parameters with ARM-CMDs showed significant results regarding fruit size and fruit weight. The trend in fruit size varied with ARM-CMDs in a logarithmic way: the model returns that the proportion of undersized fruits exceeds 10% when ARM-CMDs overtake 2,500 (Figure 4a). Moreover, a ARM-CMD exceeding 2,900 can lower the proportion of fruits with optimal size below 30% (Figure 4b). The response of fruit weight to increasing ARM-CMDs was linear and saturation did not occur considering experimental values (Figure 4c). Most records were aggregated around 195 g/fruit and very high densities were needed for an additional weight reduction. Concerning the internal quality parameters, no significant linear or exponential trends were in fit.


The life table parameters of A. schlechtendali on apple were sensibly dissimilar considering the two approaches. As expected, the highest values of rm were those calculated in second approach (Table 5). The laboratory population projection, from an initial density of 10 ARM females/increase unit showed an intrinsic rate of 0.08 day-1 in Y1 and 0.07 day-1 in Y2.
The seasonal abundance of ARM was similar in two out three years: mite numbers were low in May, started to increase in June and peaked in July. ARM densities were much higher in Y1 and Y3 than in Y2. Differences in climatic conditions across the seasons were not so relevant to explain differences in ARM abundance. Based on the intrinsic rates estimated in our study, on the other hand, biotic factors did not show a clear impact on ARM populations. ARM and phytoseiid densities were significantly correlated but the numerical response of predators to prey densities appeared to be weak in July when ARM densities suddenly increased. This phenomenon is consistent with previous observations carried out in the same area (Duso et al. 2003, Ioriatti unpubl. data). It has been reported that T. pyri performance can be affected by high temperatures (Duso et al. 1991) and this condition was recorded in July. On the other hand T. pyri has been considered a key predator of ARM in apple orchards in Northern Europe and elsewhere (e.g., Dicke 1988, Easterbrook 1996, Fitzgerald et al. 2003, Walde et al. 1997, Hardman et al. 2003). Probably the influence of climatic conditions on relationships between ARM and T. pyri requires to be investigated more in depth.The economic importance of A. schlechtendali is not homogeneous among World apple growing areas. In Europe earlier studies attributed a major concern to russeting apple peal (Ciampolini et al. 1976). Later, ARM numbers on fruitlets were found to be correlated with the amount of russet on fruits at harvest (Easterbrook and Fuller 1986). According to these authors, calyx-end and cheek russet on fruits appear when ARM feeds in the first weeks after blossoming. Extremely high ARM numbers in spring were also suspected to cause russet on apples (Lyne 1981). In England and Ireland several cultivars cannot tolerate high ARM numbers in May-June and thus ARM is considered within the factors responsible of russet formation (Cuthbertson and Murchie 2006). In contrast, in North America there are few reports of damage to fruits caused by ARM (Leeper 1981) while ARM is considered an alternative food source for predatory mites in spring before the occurrence of P. ulmi motile forms (Croft and Hoying 1977). This contradiction could be explained by the low susceptibility of the most popular apple cultivars grown in USA. More recently, there is an agreement on the potential role of ARM as an alternative prey for predatory mites involved in the control of spider mites in fruit orchards (e.g., Easterbrook 1996, Hill and Foster 1998, Duso and Pasini 2003). It should be stressed that this concept strongly depends on cultivar susceptibility.
In the present study moderate to high ARM population densities had no detectable effects on russeting. This result can be explained by mite seasonal abundance as populations increased only one month after petal fall and very low ARM numbers colonized fruitlets in spring. On the other hand this study demonstrates that ARM can affect seriously some fruit parameters. In the first experimental season the two highest density levels reduced the size, weight and colour of fruits significantly. In contrast ARM population had no detectable effects on two important agronomic parameters such as yield efficiency and return bloom. These population sizes also had an impact on soluble solids and acidity. From the economic point of view the most serious problem is represented by the reduction in fruit size from the optimal values required by the market. Surprisingly, similar effects (i.e. reduction in fruit size and weight) were observed one year later when ARM populations were much lower. This study shows that Golden Delicious may be susceptible to ARM, and that consequences on fruit size and weight can be significant when moderate to high populations occur in early summer. These results are partially consistent with those obtained in studies conducted by Spieser et al. (1999). From an economic point of view it should be stressed that the most serious problem was the reduction in fruit size from the optimal values required by the fresh market.
Angeli G., Rizzi C., Dorigoni A., Ioriatti C.2007. Population injury levels of the apple rust mite Aculus schlechtendali (Nal) on Golden Delicious and Red Delicious apple fruits. Bull. OILB/SROP, 30(4): 255-260.
Beers E.H., Brunner J.F. 1999. Effects of low rates of esfenvalerate on pest and beneficial species of apple in comparison with a standard program. J. ournal of Tree Fruit Production, 2: 33-48. doi:10.1300/J072v02n02_04 ![]()
Birch, L.C. 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol., 17: 15–26. doi:10.2307/1605 ![]()
Boller E. 1984. Eine einfache Ausschwemm-Methode zur schnellen Erfassung von Raubmilben, Thrips und anderen Kleinarthropoden im Weinbau. Schweiz., Z., Obst-und Weinb., 120: 16-17.
Ciampolini M., Rota P.A., De Schulthaus S. 1976. Rugginosità delle mele causata dall'eriofide Aculus schlechtendali. L'Informatore Agrario, 32(40): 24243-24245.
Croft B.A., Hoying S.A. 1977. Competitive displacement of Panonychus ulmi (Acarina: Tetranychidae) by Aculus schlechtendali (Acarina:Eriophyidae) in apple orchards. Can. Entomol., 109: 1025–1034. doi:10.4039/Ent1091025-8 ![]()
Cuthbertson A.G.S., Murchie A.K. 2006. Environmental monitoring of economically important invertebrate pests in Bramley apple orchards in Northern Ireland. Int. J. Env. Science and Technol., 3: 1-7. doi:10.1007/BF03325901 ![]()
Danelski W., Badowska-Czubik T., Rozpara, E. 2015. Occurrence of the apple rust mite Aculus schlechtendali (Nal.) in organic cultivation of apple. J. Res. Appl. Agric. Engin., 60 25-27.
Dicke M. 1988. Prey preference of the phytoseiid mite Typhlodromus pyri 1. Response to volatile kairomones. Exp. Appl. Acarol., 4: 1-13. doi:10.1007/BF01213837 ![]()
Duso C., Pasqualetto C., Camporese P. 1991. Role of the predatory mites Amblyseius aberrans (Oud.), Typhlodromus pyri Scheuten and Amblyseius andersoni (Chant) (Acari, Phytoseiidae) in vineyards. II. Minimum releases of A. aberrans and T. pyri to control spider mite populations (Acari, Tetranychidae). J. Appl. Entomol., 112: 298-308. doi:10.1111/j.1439-0418.1991.tb01059.x ![]()
Duso C., Pasini M. 2003. Distribution of the predatory mite Amblyseius andersoni (Acari: Phytoseiidae) on different apple cultivars. Anzeiger für Schädlingskunde – J. Pest Sc., 76: 33-40.
Duso C., Pasini M., Pellegrini M. 2003. Distribution of the predatory mite Typhlodromus pyri (Acari: Phytoseiidae) on different apple cultivars. Biocontrol Science and Technology, 13: 671-681. doi:10.1080/09583150310001606264 ![]()
Duso C., Castagnoli M., Simoni S., Angeli G. 2010. The impact of eriophyoids on crops: recent issues on Aculus schlechtendali, Calepitrimerus vitis and Aculops lycopersici. Experimental and Applied Acarology, 51: 151-168. doi:10.1007/s10493-009-9300-0 ![]()
Easterbrook M.A. 1979. The life-history of the eriophyid mite Aculus schlechtendali on apple in South-east England. Ann. Appl. Biol., 91: 287-296. doi:10.1111/j.1744-7348.1979.tb06502.x ![]()
Easterbrook M.A. 1984. The biology and control of the rust mite Aculus schlechtendali and Epitrymerus pyri on apple and pear in England. Acarology, 6(2): 797-803.
Easterbrook M.A. 1991. The effects of apple rust mite, Aculus schlechtendali, on the apple tree and its fruit. F. Dusbábek &V. Bukva (Eds.): Modern Acarology, The Hague and Academia, Prague, Vol. 1, 13.15.
Easterbrook M.A. 1996. Damage and control of eriophyid mites in apple and pear. In: Eriophyid mites, their biology, natural enemies and control. In: Lindquist E.E., Sabelis M.W., Bruin J. (Eds). Eriophyoid mites – their biology, natural enemies and control. Amsterdam: Elsevier. pp. 527-541. doi:10.1016/S1572-4379(96)80033-9 ![]()
Easterbrook M.A., Palmer J.W. 1996. The relationship between early-season leaf feeding by apple rust mite, Aculus schlechtendali (Nal.), and fruit set and photosynthesis of apple. J. Hort. Sch., 71(6): 939-944. doi:10.1080/14620316.1996.11515478 ![]()
Easterbrook M.A., Morwenna M., Fuller M.M. 1986. Russeting of apples caused by apple rust mite Aculus schlechtendali (Acarina: Eriophyidae). Ann. Appl. Biol., 109: 1-9. doi:10.1111/j.1744-7348.1986.tb03178.x ![]()
Easterbrook M.A., Fuller M.M. 1986. Russeting of apples caused by apple rust mite Aculus schlechtendali Acarina Eriophidae. Annu. Appl. Biol., 109: 1-10. doi:10.1111/j.1744-7348.1986.tb03178.x ![]()
Fitzgerald J., Solomon M., Pepper N. 2003. Reduction of broad spectrum insectcide use in apple: implications for biocontrol of extslPanonychus ulmi
. Bulletin OILB/SROP, 26(11): 37-41.Hardman J. M., Franklin J. L., Moreau D. L., Bostanian N. J. 2003. An index for selective toxicity of miticides to phytophagous mites and their predators based on orchard trials. Pest Management Science, 59: 1324-1332. doi:10.1002/ps.769 ![]()
Herbert H.J 1974. Notes on the biology of the apple rust mite, Aculus schlechtendali (Prostigmata: Eriophyoidea), and its density on several cultivars of apple in Nova Scotia. Can. Ent. 106: 1035-1038. doi:10.4039/Ent1061035-10 ![]()
Hill T.A., Foster R. 1998. Influence of selective insecticides on population dynamics of European Red Mite (Acari: Tetranychidae), Apple Rust Mite (Acari: Eriophyidae), and their predator Amblyseius fallacis (Acari: Phytoseiidae) in Apple. Journal of Economic Entomology, 91(1): 191-199. doi:10.1093/jee/91.1.191 ![]()
Höhn H., Höpli H.U. 1990. Die Apfelrostmilbe: Oft überschätzt, aber kaum prognostizierbar!. Schweiz. Z. Obst-Weinbau, 126: 259. 266.
Hoyt S.C. 1979. Integrated chemical control of insects and biological control of mites on apple in Washington. J. Econ. Entomol., 62: 74- 86. doi:10.1093/jee/62.1.74 ![]()
Hoyt S.C., Tanigoshi L.K., Browne R.W. 1979. Economic injury level studies in relation to mites on apple. Recent Adv. Acarol., 1: 3-12.
Ioriatti C., Forti D., Angeli G., Molignoni R. 1996. L'eriofide del melo (Aculus schlechtendali): morfologia, biologia e danno. Informatore Fitopatologico, 12: 9-14.
Jeppson L.R., Keifer H.H., Baker E.W. (1975). Mites injurious to economic plants. University of California Press, Berkeley, CA, USA
Kozłowski J. 1980. Researches on the occurrence and noxiousness of apple leaf mite Aculus schlechtendali (Nal.). Prace Naukowe Instytutu Ochrony Roslin, Tom XXII. zeszyt 2:155- 162.
Kozłowski J., Zielinska L. (1997). Cytological changes in apple leaves infested with the apple rust mite Aculus schlechtendali (Nal.) (Acarina: Eriophyidae). J. Pl. Prot. Res., 37(1-2): 72-84.
Leeper J.R. 1981. Extension Based Tree and Small Fruit Pest Management Strategies. New York's and Life Sciences Bulletin, 88: 1-19.
Li Q.X., Cai R.X. 1996. Occurrence and damage of the apple rust mite Aculus schlechtendali. Plant Prot., 22(3): 16-17.
Lyne P. M. 1981. Pirimiphos-methyl for rust mite control in UK orchards. Proc. Brit. Crop Prot. Confer. – Pests and Diseases, 1: 83-88.
Solhoy T., Dybwad E., Hesjedal K., Hossain M., Hovland B. 1991. Observations on Aculus schlechtendali (Nal) (Acari: Eriophyidae) in apple orchards in western Norway. In: Dusbabek F, Bukva V (eds), Modern acarology, vol 1. Academia, Prague, SPB Academic Publishing bv, The Hague, pp 533- 538.
Spieser F., Graf B., Walther P., Noesberger J. 1998. Impact of apple rust mite (Acari: Eriophyidae) feeding on apple leaf gas exchange and leaf color associated with changes in leaf tissue. Env. Entomol., 27(5): 1149- 1156. doi:10.1093/ee/27.5.1149 ![]()
Spieser F., Graf B., Höhn H., Höpli H.U. 1999. Effects of high Apple Rust Mite population densities on gas exchange, yield, fruit quality, tree growth and flower formation. Bull. OILB/SROP, 22(7): 77-85.
SPSS Inc. Released 2009. PASW Statistics for Windows, Version 18.0. Chicago: SPSS Inc.
Turchin, P. 2003. Complex Population Dynamics: a Theoretical Empirical Synthesis. Princeton, NJ: Princeton University Press 450pp.
Walde S.J., Hardman J.M., Magagula C.N. 1997. Direct and indirect species interactions influencing within season dynamics of apple rust mite, Aculus schlechtendali (Acari: Eriophyidae). Exp. Appl. Acarol., 21(9): 587-614.



2017-04-05
Date accepted:
2018-03-29
Date published:
2018-09-28
Edited by:
Kreiter, Serge

This work is licensed under a Creative Commons Attribution 4.0 International License
2018 Simoni, Sauro; Angeli, Gino; Baldessari, Mario and Duso, Carlo
 Download article
Download articleDownload the citation
RIS with abstract
(Zotero, Endnote, Reference Manager, ProCite, RefWorks, Mendeley)
RIS without abstract
BIB
(Zotero, BibTeX)
TXT
(PubMed, Txt)